
Expression of HK2, PKM2, and PFKM Is Associated with Metastasis and Late Disease Onset in Breast Cancer Patients

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Inclusion and Exclusion Criteria
2.2. Tissue and Data Collection
2.3. RNA Extraction and cDNA Synthesis
2.4. Primer Designing
2.5. Quantitative Real Time PCR
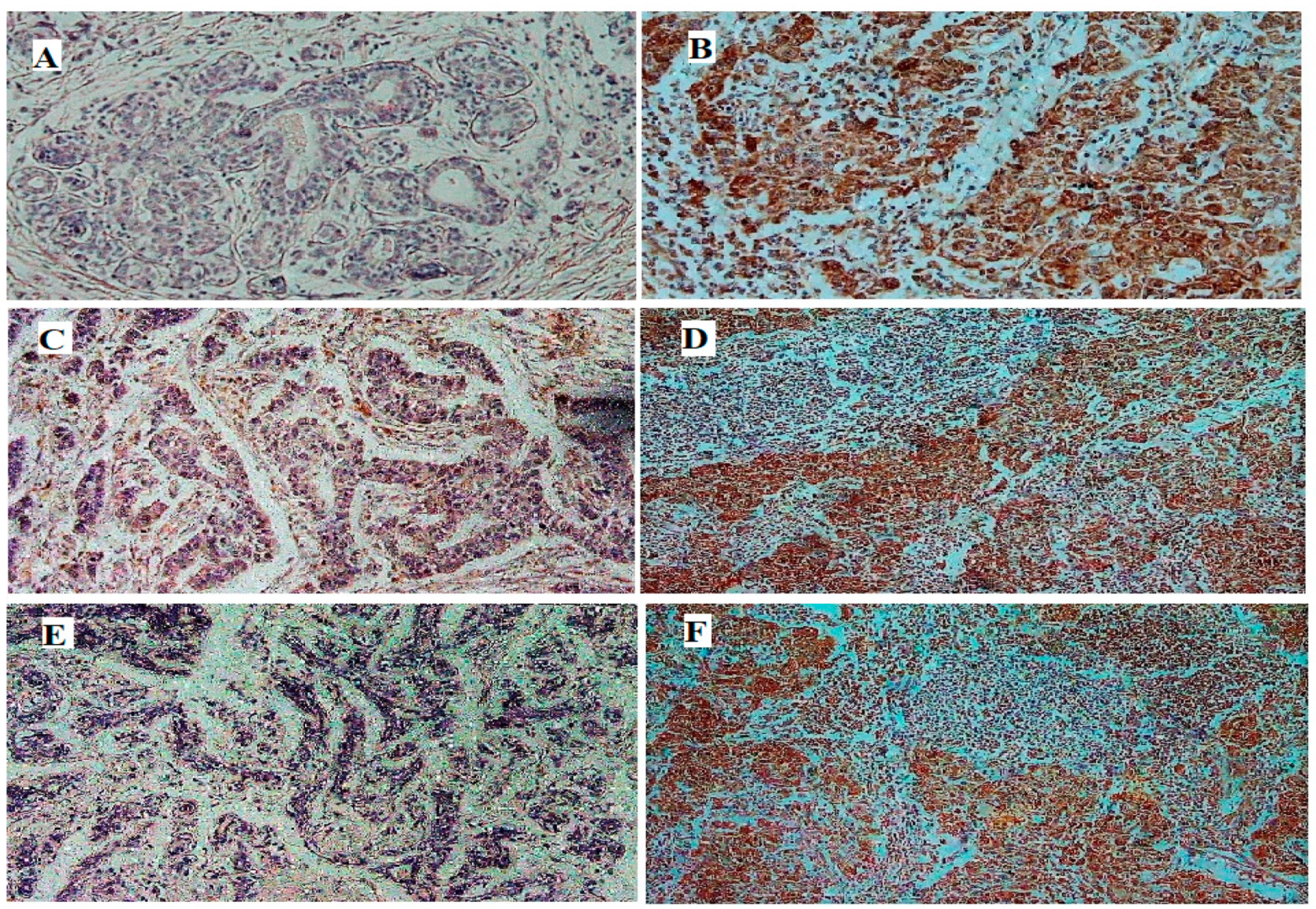
2.6. Immuno-Histochemistry (IHC)
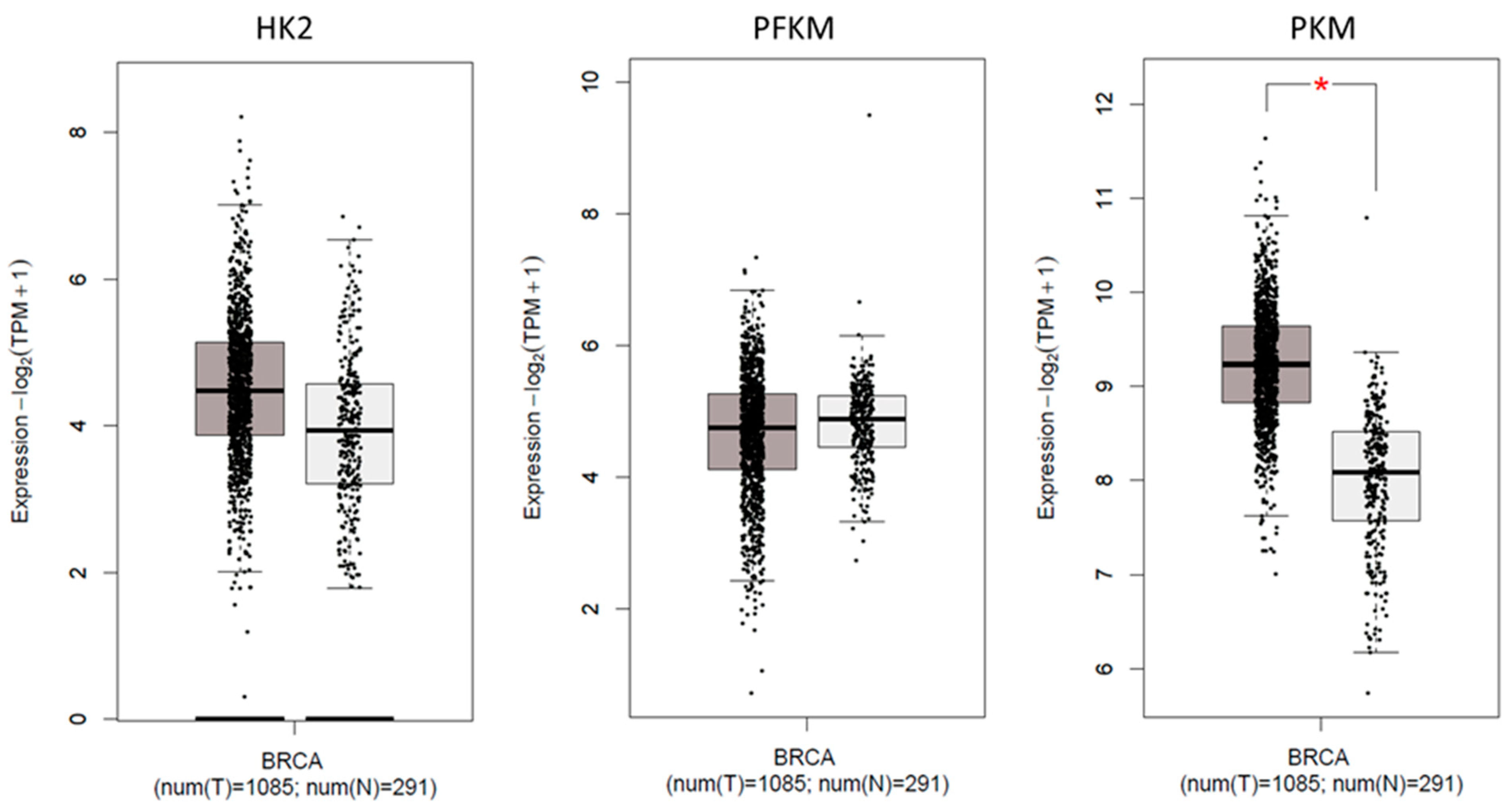
2.7. Expression of Glycolytic Genes in TCGA Cohort
2.8. Statistical Evaluation of Data
3. Results
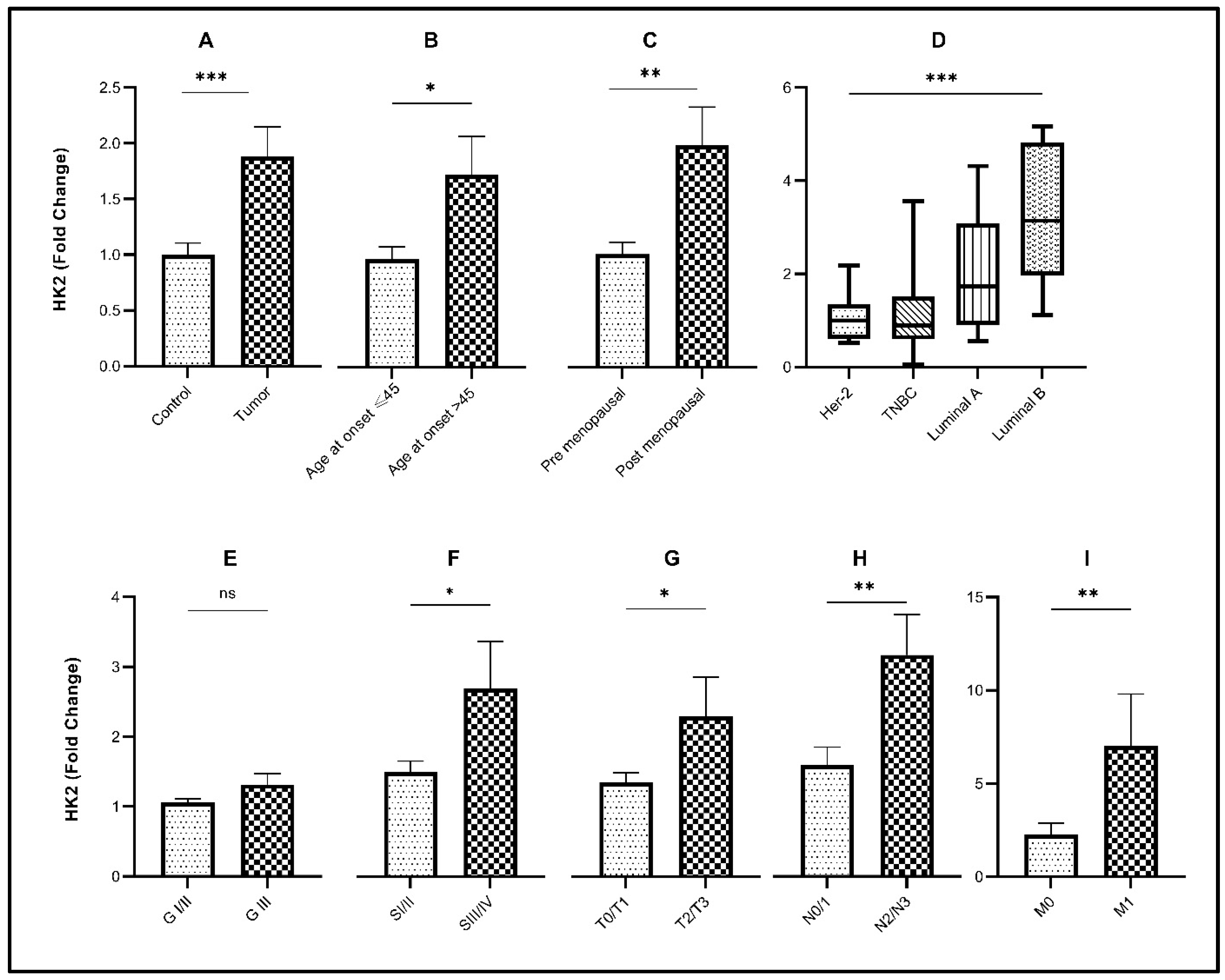
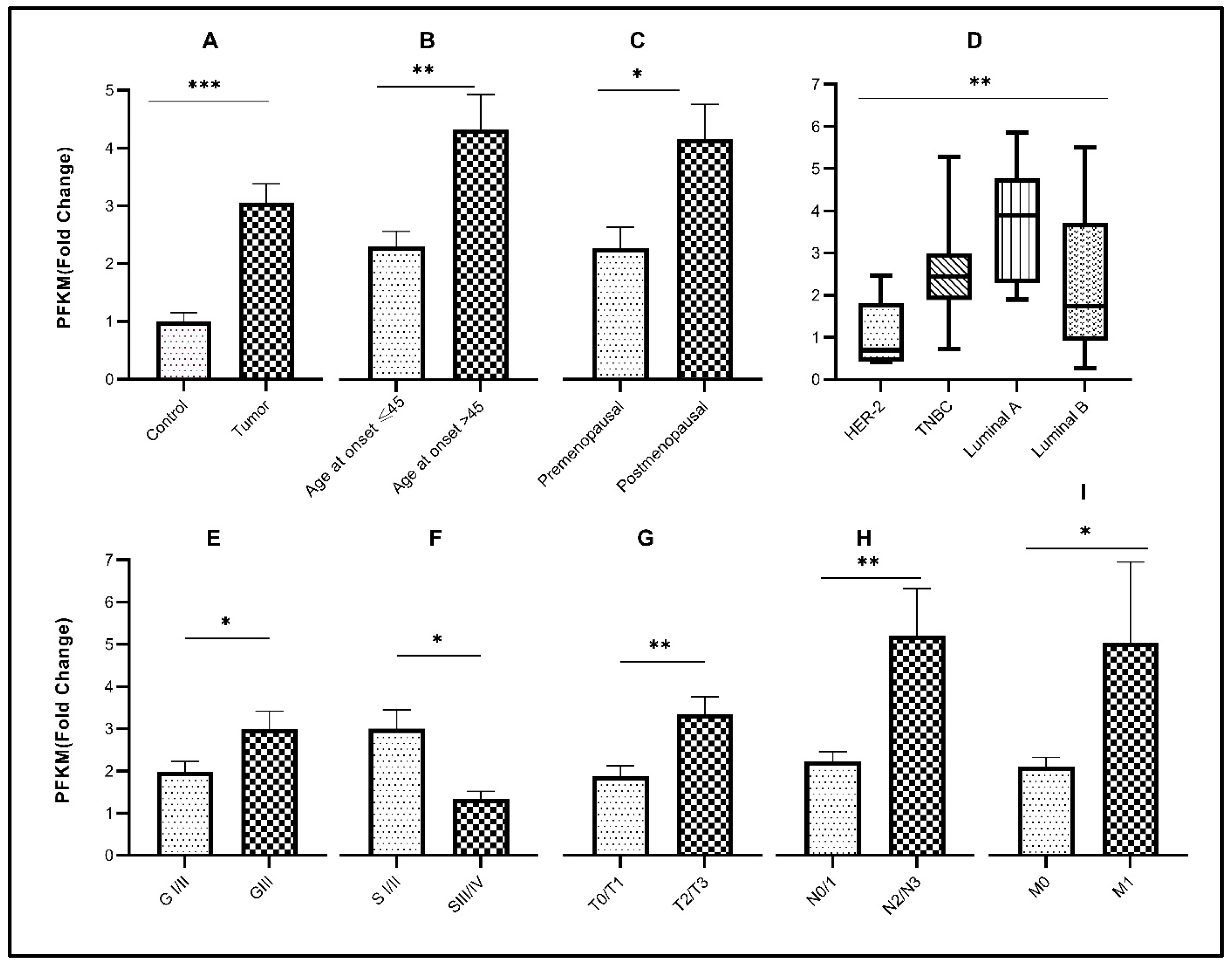
3.1. Association of Glycolytic Gene Expression with Demographic Characteristics
3.2. Relative Expression of Glycolytic Genes in Breast Cancer Study Cohort
3.3. Association of Glycolytic Genes with Clinicopathological Characteristics of the Study Cohort
3.4. Association of Glycolytic Genes with Molecular Subtypes of Breast Cancer
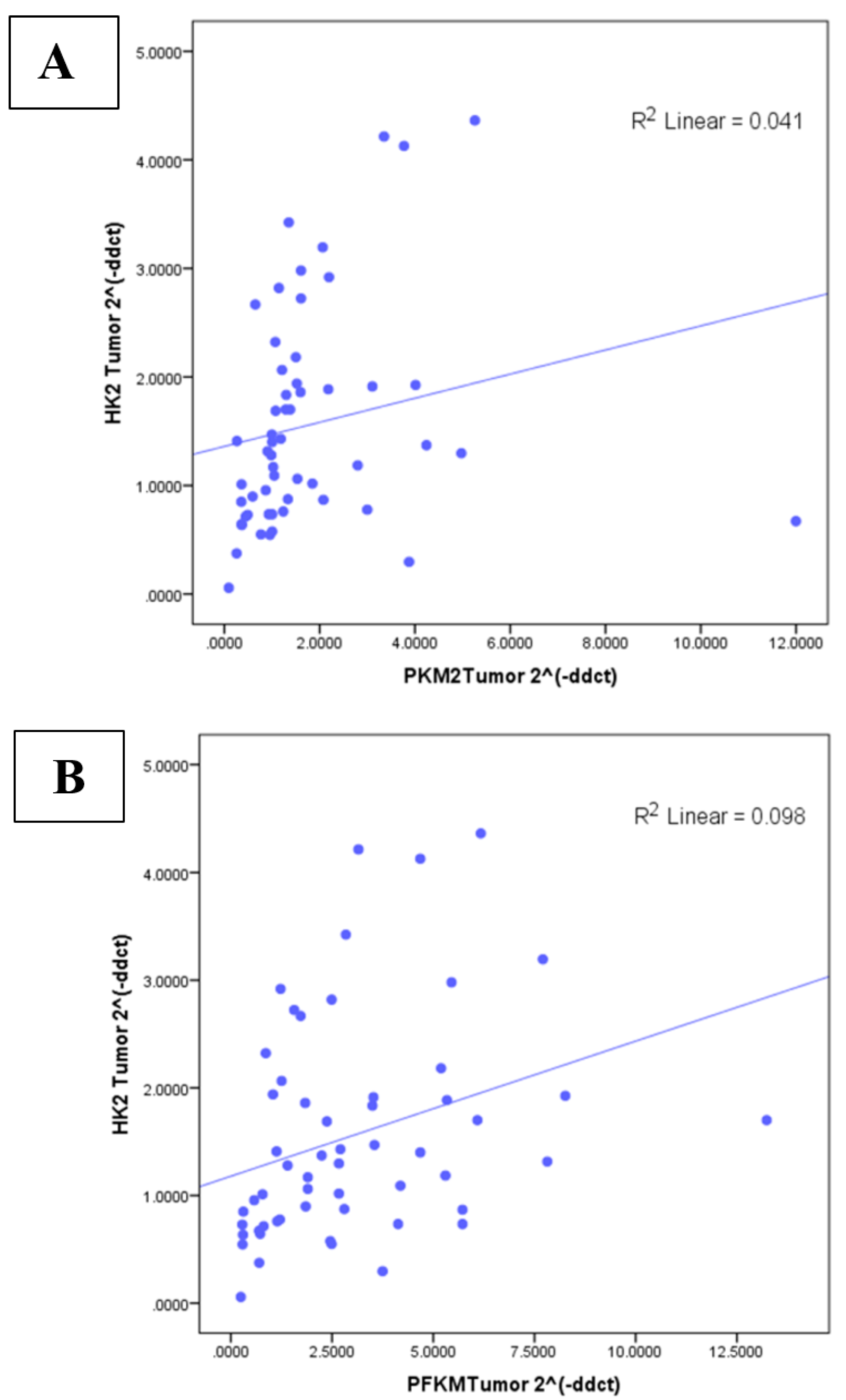
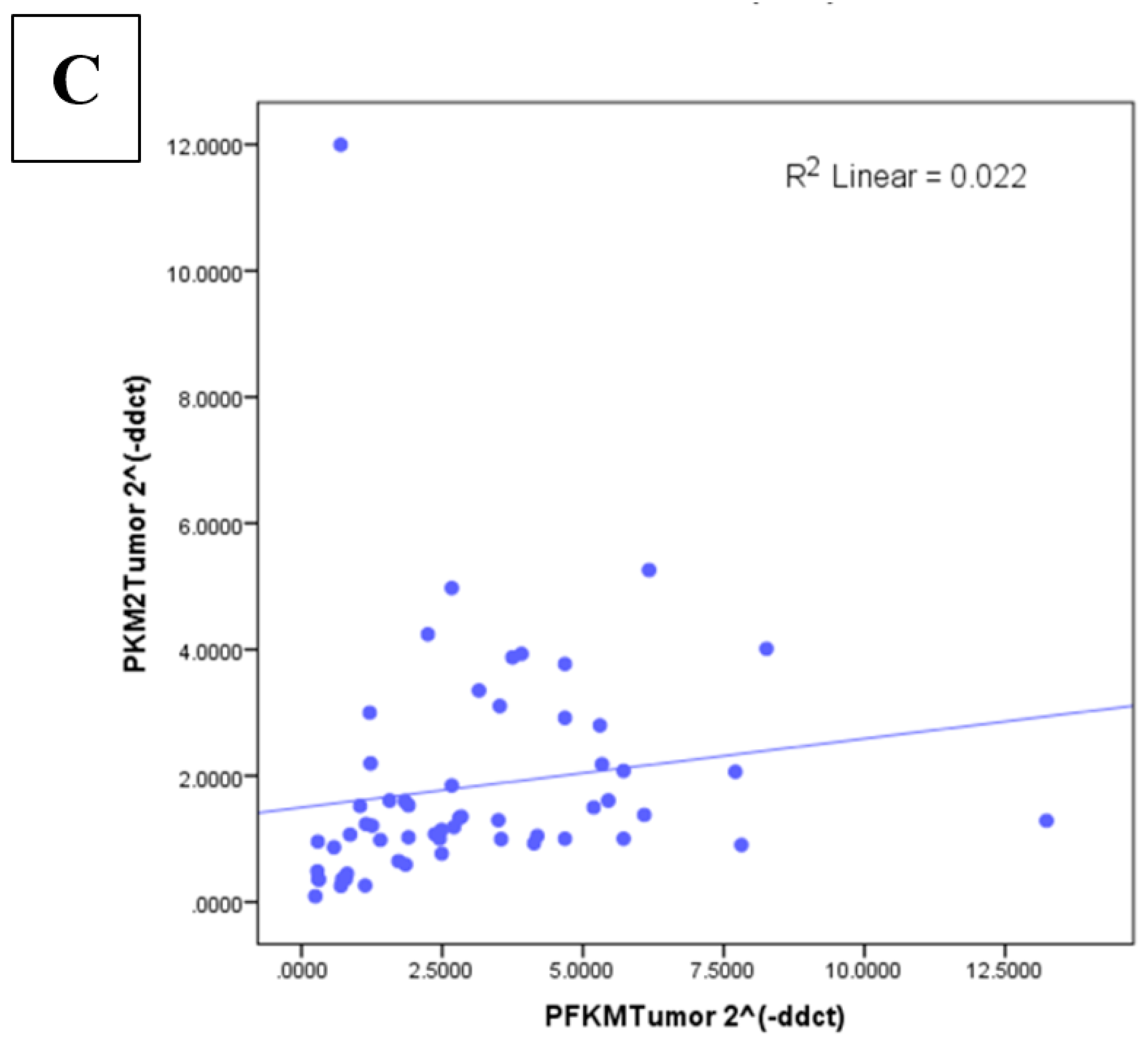
3.5. Correlation between Glycolytic Genes and Ki67 at mRNA Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| OXPHOS | Oxidative phosphorylation |
| HK | Hexokinase |
| PFK | Phosphofructokinase |
| PK | Pyruvate kinase |
| G-6P | Glucose-6-phosphate |
| PFKM | Phosphofructokinase muscle |
| PKM2 | Pyruvate kinase isozyme M2 |
| HK2 | Hexokinase2 |
| IHC | Immunohistochemistry |
| qPCR | Quantitative polymerase chain reaction |
| Ki67 | Marker of proliferation |
| HIF1 | Hypoxia inducible factor |
| cDNA | Complimentary DNA |
| mRNA | Messenger RNA |
| bps | Base pairs |
| TCGA | The Cancer Genome Atlas |
| TNBC | Triple-negative breast cancer |
References
- Tseng, P.L.; Chen, C.W.; Hu, K.H.; Cheng, H.C.; Lin, Y.H.; Tsai, W.H.; Cheng, T.J.; Wu, W.H.; Yeh, C.W.; Lin, C.C.; et al. The decrease of glycolytic enzyme hexokinase 1 accelerates tumor malignancy via deregulating energy metabolism but sensitizes cancer cells to 2-deoxyglucose inhibition. Oncotarget 2018, 9, 18949–18969. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Salvador, M.M.; de Cedrón, M.G.; Rubio, J.M.; Martínez, S.F.; Martínez, R.S.; Casado, E.; de Molina, A.R.; Sereno, M. Lipid metabolism and lung cancer. Crit. Rev. Oncol./Hematol. 2017, 112, 31–40. [Google Scholar] [CrossRef]
- Kalyanaraman, B. Teaching the basics of cancer metabolism: Developing antitumor strategies by exploiting the differences between normal and cancer cell metabolism. Redox Biol. 2017, 12, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Fadaka, A.; Ajiboye, B.; Ojo, O.; Adewale, O.; Olayide, I.; Emuowhochere, R. Biology of glucose metabolization in cancer cells. J. Oncol. Sci. 2017, 3, 45–51. [Google Scholar] [CrossRef]
- Wu, Z.; Wu, J.; Zhao, Q.; Fu, S.; Jin, J. Emerging roles of aerobic glycolysis in breast cancer. Clin. Transl. Oncol. 2020, 22, 631–646. [Google Scholar] [CrossRef] [PubMed]
- Perrin-Cocon, L.; Vidalain, P.-O.; Jacquemin, C.; Aublin-Gex, A.; Olmstead, K.; Panthu, B.; Rautureau, G.J.P.; André, P.; Nyczka, P.; Hütt, M.-T.; et al. A hexokinase isoenzyme switch in human liver cancer cells promotes lipogenesis and enhances innate immunity. Commun. Biol. 2021, 4, 217. [Google Scholar] [CrossRef] [PubMed]
- Hamabe, A.; Yamamoto, H.; Konno, M.; Uemura, M.; Nishimura, J.; Hata, T.; Takemasa, I.; Mizushima, T.; Nishida, N.; Kawamoto, K.; et al. Combined evaluation of hexokinase 2 and phosphorylated pyruvate dehydrogenase-E1α in invasive front lesions of colorectal tumors predicts cancer metabolism and patient prognosis. Cancer Sci. 2014, 105, 1100–1108. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, N.; Zhao, J.; Shi, S. High expression of hexokinase 2 promotes lung cancer proliferation and metastasis. Arch. Med. Sci. 2020, 16, 1–13. [Google Scholar] [CrossRef]
- Wu, J.; Hu, L.; Hu, F.; Zou, L.; He, T. Poor prognosis of hexokinase 2 overexpression in solid tumors of digestive system: A meta-analysis. Oncotarget 2017, 8, 32332. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Tan, Z.; Peng, C.; Yi, W. HK2 is associated with the Warburg effect and proliferation in liver cancer: Targets for effective therapy with glycyrrhizin Corrigendum in/10.3892/mmr. 2021.12143. Mol. Med. Rep. 2021, 23, 343. [Google Scholar] [CrossRef]
- Liu, C.; Wang, X.; Zhang, Y. The roles of HK2 on tumorigenesis of cervical cancer. Technol. Cancer Res. Treat. 2019, 18, 1533033819871306. [Google Scholar] [CrossRef]
- Ausina, P.; Da Silva, D.; Majerowicz, D.; Zancan, P.; Sola-Penna, M. Insulin specifically regulates expression of liver and muscle phosphofructokinase isoforms. Biomed. Pharmacother. 2018, 103, 228–233. [Google Scholar] [CrossRef]
- Li, X.B.; Gu, J.D.; Zhou, Q.H. Review of aerobic glycolysis and its key enzymes–new targets for lung cancer therapy. Thoracic cancer 2015, 6, 17–24. [Google Scholar] [CrossRef]
- Imbert-Fernandez, Y.; Clem, B.F.; O’Neal, J.; Kerr, D.A.; Spaulding, R.; Lanceta, L.; Clem, A.L.; Telang, S.; Chesney, J. Estradiol stimulates glucose metabolism via 6-phosphofructo-2-kinase (PFKFB3). J. Biol. Chem. 2014, 289, 9440–9448. [Google Scholar] [CrossRef]
- Rani, Y.; Kaur, K.; Sharma, M.; Kalia, N. In silico analysis of SNPs in human phosphofructokinase, Muscle (PFKM) gene: An apparent therapeutic target of aerobic glycolysis and cancer. Gene Rep. 2020, 21, 100920. [Google Scholar] [CrossRef]
- Li, X.; Kim, W.; Arif, M.; Gao, C.; Hober, A.; Kotol, D.; Strandberg, L.; Forsström, B.; Åsa, S.; Oksvold, P.; et al. Discovery of functional alternatively spliced PKM transcripts in human cancers. Cancers 2021, 13, 348. [Google Scholar] [CrossRef]
- Walls, J.F.; Subleski, J.J.; Palmieri, E.M.; Gonzalez-Cotto, M.; Gardiner, C.M.; McVicar, D.W.; Finlay, D.K. Metabolic but not transcriptional regulation by PKM2 is important for natural killer cell responses. Elife 2020, 9, e59166. [Google Scholar] [CrossRef]
- Hitosugi, T.; Kang, S.; Heiden, M.G.V.; Chung, T.-W.; Elf, S.; Lythgoe, K.; Dong, S.; Lonial, S.; Wang, X.; Chen, G.Z.; et al. Tyrosine phosphorylation inhibits PKM2 to promote the Warburg effect and tumor growth. Sci. Signal. 2009, 2, ra73. [Google Scholar] [CrossRef]
- Desai, S.; Ding, M.; Wang, B.; Lu, Z.; Zhao, Q.; Shaw, K.; Yung, W.A.; Weinstein, J.N.; Tan, M.; Yao, J. Tissue-specific isoform switch and DNA hypomethylation of the pyruvate kinase PKM gene in human cancers. Oncotarget 2014, 5, 8202. [Google Scholar] [CrossRef]
- Chen, X.; Chen, S.; Yu, D. Protein kinase function of pyruvate kinase M2 and cancer. Cancer Cell Int. 2020, 20, 523. [Google Scholar] [CrossRef]
- Zhang, X.; He, C.; He, C.; Chen, B.; Liu, Y.; Kong, M.; Wang, C.; Lin, L.; Dong, Y.; Sheng, H. Nuclear PKM2 expression predicts poor prognosis in patients with esophageal squamous cell carcinoma. Pathol. Res. Pract. 2013, 209, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Marbaniang, C.; Kma, L. Dysregulation of glucose metabolism by oncogenes and tumor suppressors in cancer cells. Asian Pac. J. Cancer Prev. APJCP 2018, 19, 2377. [Google Scholar] [PubMed]
- Yan, X.; Hu, Y.; Wang, B.; Wang, S.; Zhang, X. Metabolic dysregulation contributes to the progression of Alzheimer’s disease. Front. Neurosci. 2020, 14, 1107. [Google Scholar] [CrossRef] [PubMed]
- Riaz, S.K.; Khan, J.S.; Shah, S.T.A.; Wang, F.; Ye, L.; Jiang, W.G.; Malik, M.F.A. Involvement of hedgehog pathway in early onset, aggressive molecular subtypes and metastatic potential of breast cancer. Cell Commun. Signal. 2018, 16, 3. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Yu, L.; Chen, X.; Sun, X.; Wang, L.; Chen, S. The glycolytic switch in tumors: How many players are involved? J. Cancer 2017, 8, 3430. [Google Scholar] [CrossRef]
- Epstein, T.; Gatenby, R.A.; Brown, J.S. The Warburg effect as an adaptation of cancer cells to rapid fluctuations in energy demand. PLoS ONE 2017, 12, e0185085. [Google Scholar] [CrossRef]
- Rho, M.; Kim, J.; Jee, C.D.; Lee, Y.M.; Lee, H.E.; Kim, M.A.; Lee, H.S.; Kim, W.H. Expression of type 2 hexokinase and mito chondria-related genes in gastric carcinoma tissues and cell lines. Anticancer Res. 2007, 27, 251–258. [Google Scholar]
- Anderson, M.; Marayati, R.; Moffitt, R.; Yeh, J.J. Hexokinase 2 promotes tumor growth and metastasis by regulating lactate production in pancreatic cancer. Oncotarget 2017, 8, 56081–56094. [Google Scholar] [CrossRef]
- Liu, H.; Liu, N.; Cheng, Y.; Jin, W.; Zhang, P.; Wang, X.; Yang, H.; Xu, X.; Wang, Z.; Tu, Y. Hexokinase 2 (HK2), the tumor promoter in glioma, is downregulated by miR-218/Bmi1 pathway. PLoS ONE 2017, 12, e0189353. [Google Scholar]
- Lockney, N.; Zhang, M.; Lu, Y.; Sopha, S.C.; Washington, M.K.; Merchant, N.B.; Zhao, Z.; Shyr, Y.; Chakravarthy, A.B.; Xia, F. Pyruvate kinase muscle isoenzyme 2 (PKM2) expression is associated with overall survival in pancreatic ductal adenocarcinoma. J. Gastrointest. Cancer 2015, 46, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Zhang, L.; Chen, Y.; Zhou, C.; Wang, X.; Wang, D.; Liu, Z. PKM2 Promotes Breast Cancer Progression by Regulating Epithelial Mesenchymal Transition. Anal. Cell. Pathol. 2020, 2020, 8396023. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, H.; Halpern, J.; Kibriya, M.G.; Pierce, B.; Tong, L.; Gamazon, E.; McGuire, V.; Felberg, A.; Shi, J.; Jasmine, F.; et al. A genome-wide association study of early-onset breast cancer identifies PFKM as a novel breast cancer gene and supports a common genetic spectrum for breast cancer at any age. Cancer Epidemiol. Prev. Biomark. 2014, 23, 658–669. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, H.; Nagano, H.; Konno, M.; Eguchi, H.; Koseki, J.; Kawamoto, K.; Nishida, N.; Colvin, H.; Tomokuni, A.; Tomimaru, Y.; et al. The combination of the expression of hexokinase 2 and pyruvate kinase M2 is a prognostic marker in patients with pancreatic cancer. Mol. Clin. Oncol. 2015, 3, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kaarniranta, K. Glycolysis links p53 function with NF-κB signaling: Impact on cancer and aging process. J. Cell. Physiol. 2010, 224, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Mandrup, C.M.; Roland, C.B.; Egelund, J.; Nyberg, M.; Enevoldsen, L.H.; Kjaer, A.; Clemmensen, A.; Christensen, A.N.; Suetta, C.; Frikke-Schmidt, R.; et al. Effects of High-Intensity Exercise Training on Adipose Tissue Mass, Glucose Uptake and Protein Content in Pre-and Post-menopausal Women. Front. Sports Act. Living 2020, 2, 60. [Google Scholar] [CrossRef] [PubMed]
- Boonyaratanakornkit, V.; Pateetin, P. The role of ovarian sex steroids in metabolic homeostasis, obesity, and postmenopausal breast cancer: Molecular mechanisms and therapeutic implications. BioMed Res. Int. 2015, 2015, 140196. [Google Scholar] [CrossRef]
- Chen, J.-Q.; Brown, T.R.; Russo, J. Regulation of energy metabolism pathways by estrogens and estrogenic chemicals and potential implications in obesity associated with increased exposure to endocrine disruptors. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2009, 1793, 1128–1143. [Google Scholar] [CrossRef]
- Serrano-Carbajal, E.A.; Espinal-Enriquez, J.; Hernández-Lemus, E. Targeting metabolic deregulation landscapes in breast cancer subtypes. Front. Oncol. 2020, 10, 97. [Google Scholar] [CrossRef]
- Martínez-Ordoñez, A.; Seoane, S.; Avila, L.; Eiro, N.; Macía, M.; Arias, E.; Pereira, F.; García-Caballero, T.; Gómez-Lado, N.; Aguiar, P.; et al. POU1F1 transcription factor induces metabolic reprogramming and breast cancer progression via LDHA regulation. Oncogene 2021, 40, 2725–2740. [Google Scholar] [CrossRef]
- Li, W.C.; Huang, C.H.; Hsieh, Y.T.; Chen, T.Y.; Cheng, L.H.; Chen, C.Y.; Liu, C.J.; Chen, H.M.; Huang, C.L.; Lo, J.F.; et al. Regulatory Role of Hexokinase 2 in Modulating Head and Neck Tumorigenesis. Front. Oncol. 2020, 10, 176. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yan, M.; Wu, X.; Wang, Y.; Huang, L. Expression and clinical significance of pyruvate kinase M2 in breast cancer: A protocol for meta-analysis and bioinformatics validation analysis. Medicine 2021, 100, e25545. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Gene Name | Forward/Reverse Primer Seq | Product Size |
|---|---|---|---|
| 1 | HK2 | GAGTTTGACCTGGATGTGGTTGC (Forward) | 130 bps |
| CCTCCATGTAGCAGGCATTGCT (Reverse) | |||
| 2 | PKM2 | AGGACCTGAGATCCGAACTG(Forward) | 132 bps |
| AGCCACAGGATGTTCTCGTC (Reverse) | |||
| 3 | PFKM | ATGACCCATGAAGAGCACCA (Forward) | 137 bps |
| GCACCGGTGAAGATACCAAC (Reverse) | |||
| 4 | β-actin | CTGAACCCCAAGGCCAAC (Forward) | 108 bps |
| AGAGGCGTACAGGGATAGCA (Reverse) | |||
| 5 | Ki-67 | GCCTTGGTCTCTTGGGAATAC (Forward) | 123 bps |
| GGAGATTAGGAGCCAGTTTGAG (Reverse) |
| Variables | Total (%) | HK2high | PKM2high | PFKMhigh |
|---|---|---|---|---|
| Age of disease onset ≤45 | 30(50) | 19 | 21 | 21 |
| Age of disease onset >45 | 30(50) | 22 | 23 | 27 |
| Premenopausal status | 25(42) | 14 | 20 | 17 |
| Postmenopausal status | 35(58) | 27 | 24 | 31 |
| Laterality (left) | 30(50) | 19 | 21 | 23 |
| Laterality (right) | 30(50) | 22 | 23 | 25 |
| Grade I/II | 45(75) | 28 | 32 | 35 |
| Grade III | 15(25) | 13 | 12 | 13 |
| Stage I/II | 41(68) | 26 | 30 | 32 |
| Stage III/IV | 19(22) | 15 | 14 | 16 |
| N0/N1 | 49(82) | 32 | 36 | 38 |
| N2/N3 | 11(18) | 9 | 8 | 10 |
| No metastasis(M0) | 57(95) | 38 | 41 | 45 |
| Distant metastasis(M1) | 3(5) | 3 | 3 | 3 |
| T1/T2 | 40(67) | 28 | 29 | 30 |
| T3/T4 | 20(33) | 13 | 15 | 18 |
| Variables | Total | HK2 | PKM2 | PFKM | |||
|---|---|---|---|---|---|---|---|
| Mean ± SEM | p-Value | Mean ± SEM | p-Value | Mean ± SEM | p-Value | ||
| Age of disease onset <45 | 30 | 1.197 ± 0.1129 | 0.0423 # | 2.112 ± 0.4516 | 0.5593 # | 2.393 ± 0.2688 | 0.0038 # |
| Age of disease onset >45 | 30 | 1.786 ± 0.3399 | 1.599 ± 0.2239 | 4.495 ± 0.6518 | |||
| Premenopausal status | 25 | 1.011 ± 0.1066 | 0.0233 # | 0.9701 ± 0.1295 | 0.0192 # | 2.229 ± 0.3476 | 0.0183 # |
| Postmenopausal status | 35 | 1.912 ± 0.3129 | 1.741 ± 0.2095 | 4.073 ± 0.5835 | |||
| Laterality (left) | 30 | 1.553 ± 0.3226 | 0.1257 # | 1.044 ± 0.1162 | 0.0488 # | 2.584 ± 0.3395 | 0.5137 # |
| Laterality (right) | 30 | 1.909 ± 0.3522 | 2.233 ± 0.4911 | 3.368 ± 0.5807 | |||
| Tumor | 60 | 1.948 ± 0.2848 | <0.0001 ¥ | 1.960 ± 0.09343 | <0.0001 ¥ | 3.056 ± 0.3269 | <0.0001 ¥ |
| Control | 60 | 1.000 ± 0.1055 | 1.000 ± 0.2483 | 1.000 ± 0.15330 | |||
| Grade I/II | 45 | 1.058 ± 0.05400 | 0.1121 # | 1.309 ± 0.1353 | 0.2237 # | 1.976 ± 0.2425 | 0.0403 # |
| Grade III | 15 | 1.314 ± 0.1560 | 2.165 ± 0.6173 | 2.987 ± 0.4276 | |||
| Stage I/II | 41 | 1.496 ± 0.1572 | 0.0459 # | 1.161 ± 0.1164 | 0.0302 # | 2.786 ± 0.4199 | 0.0459 # |
| Stage III/IV | 19 | 2.694 ± 0.6708 | 1.881 ± 0.2609 | 1.241 ± 0.1621 | |||
| N0/N1 | 49 | 1.595 ± 0.2565 | 0.0010 # | 1.582 ± 0.1863 | 0.0084 # | 2.123 ± 0.2261 | 0.0078 # |
| N2/N3 | 11 | 3.171 ± 0.5819 | 3.529 ± 0.8762 | 4.985 ± 1.071 | |||
| No metastasis (M0) | 57 | 2.259 ± 0.6133 | 0.0039 # | 1.620 ± 0.1726 | 0.0088 # | 2.015 ± 0.2073 | 0.0432 # |
| Distant metastasis (M1) | 3 | 7.009 ± 1.794 | 4.066 ± 0.6310 | 4.816 ± 1.835 | |||
| T Stage: T1/T2 | 40 | 1.349 ± 0.1343 | 0.0499 # | 1.652 ± 0.2400 | 0.0090 # | 1.801 ± 0.2317 | 0.0026 # |
| T Stage: T3/T4 | 20 | 2.299 ± 0.5588 | 2.480 ± 0.4631 | 3.205 ± 0.3991 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishfaq, M.; Bashir, N.; Riaz, S.K.; Manzoor, S.; Khan, J.S.; Bibi, Y.; Sami, R.; Aljahani, A.H.; Alharthy, S.A.; Shahid, R. Expression of HK2, PKM2, and PFKM Is Associated with Metastasis and Late Disease Onset in Breast Cancer Patients. Genes 2022, 13, 549. https://doi.org/10.3390/genes13030549
Ishfaq M, Bashir N, Riaz SK, Manzoor S, Khan JS, Bibi Y, Sami R, Aljahani AH, Alharthy SA, Shahid R. Expression of HK2, PKM2, and PFKM Is Associated with Metastasis and Late Disease Onset in Breast Cancer Patients. Genes. 2022; 13(3):549. https://doi.org/10.3390/genes13030549
Chicago/Turabian StyleIshfaq, Mehreen, Nabiha Bashir, Syeda Kiran Riaz, Shumaila Manzoor, Jahangir Sarwar Khan, Yamin Bibi, Rokayya Sami, Amani H. Aljahani, Saif A. Alharthy, and Ramla Shahid. 2022. "Expression of HK2, PKM2, and PFKM Is Associated with Metastasis and Late Disease Onset in Breast Cancer Patients" Genes 13, no. 3: 549. https://doi.org/10.3390/genes13030549
APA StyleIshfaq, M., Bashir, N., Riaz, S. K., Manzoor, S., Khan, J. S., Bibi, Y., Sami, R., Aljahani, A. H., Alharthy, S. A., & Shahid, R. (2022). Expression of HK2, PKM2, and PFKM Is Associated with Metastasis and Late Disease Onset in Breast Cancer Patients. Genes, 13(3), 549. https://doi.org/10.3390/genes13030549