
The Role of NEDD4 E3 Ubiquitin–Protein Ligases in Parkinson’s Disease



Abstract
1. Introduction
2. E3 Ligases Ubiquitinating α-Synuclein
3. NEDD4-1 and NEDD4-2 Expression
4. NEDD4-1 and NEDD4-2 Structure und Posttranslational Modifications
5. NEDD4-1 and NEDD4-2 Substrates, Adaptors, Regulators, and Function
6. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goedert, M.; Spillantini, M.G.; Del Tredici, K.; Braak, H. 100 years of Lewy pathology. Nat. Rev. Neurol. 2013, 9, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Singleton, A.; Gwinn-Hardy, K. Parkinson’s disease and dementia with Lewy bodies: A difference in dose? Lancet 2004, 364, 1105–1107. [Google Scholar] [CrossRef]
- Lassot, I.; Mora, S.; Lesage, S.; Zieba, B.A.; Coque, E.; Condroyer, C.; Bossowski, J.P.; Mojsa, B.; Marelli, C.; Soulet, C.; et al. The E3 Ubiquitin Ligases TRIM17 and TRIM41 Modulate α-Synuclein Expression by Regulating ZSCAN21. Cell Rep. 2018, 25, 2484–2496.e2489. [Google Scholar] [CrossRef] [PubMed]
- Abeywardana, T.; Lin, Y.H.; Rott, R.; Engelender, S.; Pratt, M.R. Site-specific differences in proteasome-dependent degradation of monoubiquitinated α-synuclein. Chem. Biol. 2013, 20, 1207–1213. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; Li, J.-D. The Roles of Post-translational Modifications on α-Synuclein in the Pathogenesis of Parkinson’s Diseases. Front. Neurosci. 2019, 13, 381. [Google Scholar] [CrossRef]
- Hasegawa, M.; Fujiwara, H.; Nonaka, T.; Wakabayashi, K.; Takahashi, H.; Lee, V.M.; Trojanowski, J.Q.; Mann, D.; Iwatsubo, T. Phosphorylated α-synuclein is ubiquitinated in α-synucleinopathy lesions. J. Biol. Chem. 2002, 277, 49071–49076. [Google Scholar] [CrossRef]
- Tofaris, G.K.; Razzaq, A.; Ghetti, B.; Lilley, K.S.; Spillantini, M.G. Ubiquitination of α-synuclein in Lewy bodies is a pathological event not associated with impairment of proteasome function. J. Biol. Chem. 2003, 278, 44405–44411. [Google Scholar] [CrossRef]
- Rott, R.; Szargel, R.; Haskin, J.; Bandopadhyay, R.; Lees, A.J.; Shani, V.; Engelender, S. α-Synuclein fate is determined by USP9X-regulated monoubiquitination. Proc. Natl. Acad. Sci. USA 2011, 108, 18666–18671. [Google Scholar] [CrossRef]
- Deng, L.; Meng, T.; Chen, L.; Wei, W.; Wang, P. The role of ubiquitination in tumorigenesis and targeted drug discovery. Signal Transduct. Target. Ther. 2020, 5, 11. [Google Scholar] [CrossRef]
- Metzger, M.B.; Hristova, V.A.; Weissman, A.M. HECT and RING finger families of E3 ubiquitin ligases at a glance. J. Cell Sci. 2012, 125, 531. [Google Scholar] [CrossRef]
- Shimura, H.; Schlossmacher, M.G.; Hattori, N.; Frosch, M.P.; Trockenbacher, A.; Schneider, R.; Mizuno, Y.; Kosik, K.S.; Selkoe, D.J. Ubiquitination of a new form of α-synuclein by parkin from human brain: Implications for Parkinson’s disease. Science 2001, 293, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Kitada, T.; Asakawa, S.; Hattori, N.; Matsumine, H.; Yamamura, Y.; Minoshima, S.; Yokochi, M.; Mizuno, Y.; Shimizu, N. Mutations in the parkin gene cause autosomal recessive juvenile parkinsonism. Nature 1998, 392, 605–608. [Google Scholar] [CrossRef] [PubMed]
- Dawson, T.M.; Dawson, V.L. The role of parkin in familial and sporadic Parkinson’s disease. Mov. Disord. 2010, 25, S32–S39. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liao, X.; Rao, X.; Wang, B.; Zhang, J.; Xu, G.; Jiang, X.; Qin, X.; Chen, C.; Zou, Z. The lysosomal membrane protein LAMP-2 is dispensable for PINK1/Parkin-mediated mitophagy. FEBS Lett. 2020, 594, 823–840. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.K.K.; Zhang, Y.; Lim, K.L.; Tanaka, Y.; Huang, H.; Gao, J.; Ross, C.A.; Dawson, V.L.; Dawson, T.M. Parkin ubiquitinates the α-synuclein–interacting protein, synphilin-1: Implications for Lewy-body formation in Parkinson disease. Nat. Med. 2001, 7, 1144–1150. [Google Scholar] [CrossRef]
- Ribeiro, C.S.; Carneiro, K.; Ross, C.A.; Menezes, J.R.L.; Engelender, S. Synphilin-1 Is Developmentally Localized to Synaptic Terminals, and Its Association with Synaptic Vesicles Is Modulated by α-Synuclein. J. Biol. Chem. 2002, 277, 23927–23933. [Google Scholar] [CrossRef] [PubMed]
- Wilkaniec, A.; Lenkiewicz, A.M.; Czapski, G.A.; Jęśko, H.M.; Hilgier, W.; Brodzik, R.; Gąssowska-Dobrowolska, M.; Culmsee, C.; Adamczyk, A. Extracellular A-Synuclein Oligomers Induce Parkin S-Nitrosylation: Relevance to Sporadic Parkinson’s Disease Etiopathology. Mol. Neurobiol. 2019, 56, 125–140. [Google Scholar] [CrossRef]
- Wheeler, T.C.; Chin, L.-S.; Li, Y.; Roudabush, F.L.; Li, L. Regulation of Synaptophysin Degradation by Mammalian Homologues of Seven in Absentia. J. Biol. Chem. 2002, 277, 10273–10282. [Google Scholar] [CrossRef]
- Liani, E.; Eyal, A.; Avraham, E.; Shemer, R.; Szargel, R.; Berg, D.; Bornemann, A.; Riess, O.; Ross, C.A.; Rott, R.; et al. Ubiquitylation of synphilin-1 and α-synuclein by SIAH and its presence in cellular inclusions and Lewy bodies imply a role in Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2004, 101, 5500–5505. [Google Scholar] [CrossRef]
- Lee, J.T.; Wheeler, T.C.; Li, L.; Chin, L.S. Ubiquitination of -synuclein by Siah-1 promotes -synuclein aggregation and apoptotic cell death. Hum. Mol. Genet. 2007, 17, 906–917. [Google Scholar] [CrossRef]
- Rott, R.; Szargel, R.; Haskin, J.; Shani, V.; Shainskaya, A.; Manov, I.; Liani, E.; Avraham, E.; Engelender, S. Monoubiquitylation of α-synuclein by seven in absentia homolog (SIAH) promotes its aggregation in dopaminergic cells. J. Biol. Chem. 2008, 283, 3316–3328. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.; Klucken, J.; Patterson, C.; Hyman, B.T.; McLean, P.J. The co-chaperone carboxyl terminus of Hsp70-interacting protein (CHIP) mediates α-synuclein degradation decisions between proteasomal and lysosomal pathways. J. Biol. Chem. 2005, 280, 23727–23734. [Google Scholar] [CrossRef] [PubMed]
- Tetzlaff, J.E.; Putcha, P.; Outeiro, T.F.; Ivanov, A.; Berezovska, O.; Hyman, B.T.; McLean, P.J. CHIP targets toxic α-Synuclein oligomers for degradation. J. Biol. Chem. 2008, 283, 17962–17968. [Google Scholar] [CrossRef] [PubMed]
- Kalia, L.V.; Kalia, S.K.; Chau, H.; Lozano, A.M.; Hyman, B.T.; McLean, P.J. Ubiquitinylation of α-Synuclein by Carboxyl Terminus Hsp70-Interacting Protein (CHIP) Is Regulated by Bcl-2-Associated Athanogene 5 (BAG5). PLoS ONE 2011, 6, e14695. [Google Scholar] [CrossRef]
- Gerez, J.A.; Prymaczok, N.C.; Rockenstein, E.; Herrmann, U.S.; Schwarz, P.; Adame, A.; Enchev, R.I.; Courtheoux, T.; Boersema, P.J.; Riek, R.; et al. A cullin-RING ubiquitin ligase targets exogenous α-synuclein and inhibits Lewy body-like pathology. Sci. Transl. Med. 2019, 11, 6722. [Google Scholar] [CrossRef]
- Scheffner, M.; Kumar, S. Mammalian HECT ubiquitin-protein ligases: Biological and pathophysiological aspects. Biochim. Biophys. Acta 2014, 1843, 61–74. [Google Scholar] [CrossRef]
- Mund, T.; Masuda-Suzukake, M.; Goedert, M.; Pelham, H.R. Ubiquitination of α-synuclein filaments by Nedd4 ligases. PLoS ONE 2018, 13, e0200763. [Google Scholar] [CrossRef]
- Tofaris, G.K.K.; Hourez, R.; Jung, J.; Kim, K.P.; Goldberg, A.L. Ubiquitin ligase Nedd4 promotes α-synuclein degradation by the endosomal–lysosomal pathway. Proc. Natl. Acad. Sci. USA 2011, 108, 17004–17009. [Google Scholar] [CrossRef]
- Sugeno, N.; Hasegawa, T.; Tanaka, N.; Fukuda, M.; Wakabayashi, K.; Oshima, R.; Konno, M.; Miura, E.; Kikuchi, A.; Baba, T.; et al. Lys-63-linked ubiquitination by E3 ubiquitin ligase Nedd4-1 facilitates endosomal sequestration of internalized α-synuclein. J. Biol. Chem. 2014, 289, 18137–18151. [Google Scholar] [CrossRef]
- Davies, S.E.; Hallett, P.J.; Moens, T.; Smith, G.; Mangano, E.; Kim, H.T.; Goldberg, A.L.; Liu, J.-L.; Isacson, O.; Tofaris, G.K. Enhanced ubiquitin-dependent degradation by Nedd4 protects against α-synuclein accumulation and toxicity in animal models of Parkinson’s disease. Neurobiol. Dis. 2014, 64, 79–87. [Google Scholar] [CrossRef]
- Akkaya, B.G.; Zolnerciks, J.K.; Ritchie, T.K.; Bauer, B.; Hartz, A.M.; Sullivan, J.A.; Linton, K.J. The multidrug resistance pump ABCB1 is a substrate for the ubiquitin ligase NEDD4-1. Mol. Membr. Biol. 2015, 32, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Platta, H.W.; Abrahamsen, H.; Thoresen, S.B.; Stenmark, H. Nedd4-dependent lysine-11-linked polyubiquitination of the tumour suppressor Beclin 1. Biochem. J. 2012, 441, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, S.K.; Xiao, K.; Venkataramanan, V.; Snyder, P.M.; Freedman, N.J.; Weissman, A.M. Nedd4 Mediates Agonist-dependent Ubiquitination, Lysosomal Targeting, and Degradation of the β2-Adrenergic Receptor. J. Biol. Chem. 2008, 283, 22166–22176. [Google Scholar] [CrossRef] [PubMed]
- Simonin, A.; Fuster, D. Nedd4-1 and β-Arrestin-1 Are Key Regulators of Na+/H+ Exchanger 1 Ubiquitylation, Endocytosis, and Function. J. Biol. Chem. 2010, 285, 38293–38303. [Google Scholar] [CrossRef] [PubMed]
- Harvey, K.F.; Kumar, S. Nedd4-like proteins: An emerging family of ubiquitin-protein ligases implicated in diverse cellular functions. Trends Cell Biol. 1999, 9, 166–169. [Google Scholar] [CrossRef]
- Jiang, C.; Kawabe, H.; Rotin, D. The Ubiquitin Ligase Nedd4L Regulates the Na/K/2Cl Co-transporter NKCC1/SLC12A2 in the Colon. J. Biol. Chem. 2017, 292, 3137–3145. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, S.; Sun, Z.; Guo, X.; Zhou, H. E3 ubiquitin ligase Nedd4 is a key negative regulator for non-canonical inflammasome activation. Cell Death Differ. 2019, 26, 2386–2399. [Google Scholar] [CrossRef]
- Guo, J.; Wang, T.; Li, X.; Shallow, H.; Yang, T.; Li, W.; Xu, J.; Fridman, M.D.; Yang, X.; Zhang, S. Cell Surface Expression of Human Ether-a-go-go-related Gene (hERG) Channels Is Regulated by Caveolin-3 Protein via the Ubiquitin Ligase Nedd4-2*. J. Biol. Chem. 2012, 287, 33132–33141. [Google Scholar] [CrossRef]
- Yang, B.; Gay, D.L.; Macleod, M.K.L.; Cao, X.; Hala, T.; Sweezer, E.M.; Kappler, J.; Marrack, P.; Oliver, P.M. Nedd4 augments the adaptive immune response by promoting ubiquitin-mediated degradation of Cbl-b in activated T cells. Nat. Immunol. 2008, 9, 1356–1363. [Google Scholar] [CrossRef]
- Pham, N.; Rotin, D. Nedd4 regulates ubiquitination and stability of the guanine-nucleotide exchange factor CNrasGEF. J. Biol. Chem. 2001, 276, 46995–47003. [Google Scholar] [CrossRef]
- Zhang, H.; Nie, W.; Zhang, X.; Zhang, G.; Li, Z.; Wu, H.; Shi, Q.; Chen, Y.; Ding, Z.; Zhou, X.; et al. NEDD4-1 regulates migration and invasion of glioma cells through CNrasGEF ubiquitination in vitro. PLoS ONE 2013, 8, e82789. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Kumar, S. Nedd4 and Nedd4-2: Closely related ubiquitin-protein ligases with distinct physiological functions. Cell Death Differ. 2010, 17, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Persaud, A.; Alberts, P.; Mari, S.; Tong, J.; Murchie, R.; Maspero, E.; Safi, F.; Moran, M.F.; Polo, S.; Rotin, D. Tyrosine phosphorylation of NEDD4 activates its ubiquitin ligase activity. Sci. Signal. 2014, 7, ra95. [Google Scholar] [CrossRef] [PubMed]
- Bakkers, J.; Camacho-Carvajal, M.; Nowak, M.; Kramer, C.; Danger, B.; Hammerschmidt, M. Destabilization of Δα; by Nedd4-Mediated Ubiquitination Ubc9-Mediated Sumoylation, and Its Implications on Dorsoventral Patterning of the Zebrafish Embryo. Cell Cycle 2005, 4, 790–800. [Google Scholar] [CrossRef]
- Leonard, M.K.; Hill, N.T.; Grant, E.D.; Kadakia, M.P. ΔNp63α represses nuclear translocation of PTEN by inhibition of NEDD4-1 in keratinocytes. Arch. Derm. Res. 2013, 305, 733–739. [Google Scholar] [CrossRef]
- Woelk, T.; Oldrini, B.; Maspero, E.; Confalonieri, S.; Cavallaro, E.; Di Fiore, P.P.; Polo, S. Molecular mechanisms of coupled monoubiquitination. Nat. Cell Biol. 2006, 8, 1246–1254. [Google Scholar] [CrossRef]
- Persaud, A.; Alberts, P.; Hayes, M.; Guettler, S.; Clarke, I.; Sicheri, F.; Dirks, P.; Ciruna, B.; Rotin, D. Nedd4-1 binds and ubiquitylates activated FGFR1 to control its endocytosis and function. EMBO J. 2011, 30, 3259–3273. [Google Scholar] [CrossRef]
- Vittal, V.; Stewart, M.D.; Brzovic, P.S.; Klevit, R.E. Regulating the Regulators: Recent Revelations in the Control of E3 Ubiquitin Ligases. J. Biol. Chem. 2015, 290, 21244–21251. [Google Scholar] [CrossRef]
- Blot, V.; Perugi, F.; Gay, B.; PréVost, M.-C.; Briant, L.; Tangy, F.D.R.; Abriel, H.; Staub, O.; DokhéLar, M.-C.; Pique, C. Nedd4.1-mediated ubiquitination and subsequent recruitment of Tsg101 ensure HTLV-1 Gag trafficking towards the multivesicular body pathway prior to virus budding. J. Cell Sci. 2004, 117, 2357–2367. [Google Scholar] [CrossRef]
- Rost, M.; Döring, T.; Prange, R. γ2-Adaptin, a ubiquitin-interacting adaptor, is a substrate to coupled ubiquitination by the ubiquitin ligase Nedd4 and functions in the endosomal pathway. J. Biol. Chem. 2008, 283, 32119–32130. [Google Scholar] [CrossRef]
- Prange, R. Host factors involved in hepatitis B virus maturation, assembly, and egress. Med. Microbiol. Immunol. 2012, 201, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Wan, T.; Lei, Z.; Tu, B.; Wang, T.; Wang, J.; Huang, F. NEDD4 Induces K48-Linked Degradative Ubiquitination of Hepatitis B Virus X Protein and Inhibits HBV-Associated HCC Progression. Front. Oncol. 2021, 11, 625169. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Choi, B.K.; Mujoo, K.; Fan, X.; Fa, M.; Mukherjee, S.; Owiti, N.; Zhang, N.; An, Z. The E3 ubiquitin ligase NEDD4 negatively regulates HER3/ErbB3 level and signaling. Oncogene 2015, 34, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, L.; Zhou, J.; Luo, S.; Huang, R.; Zhao, C.; Diao, A. Nedd4 E3 ubiquitin ligase promotes cell proliferation and autophagy. Cell Prolif. 2015, 48, 338–347. [Google Scholar] [CrossRef]
- Katz, M.; Shtiegman, K.; Tal-Or, P.; Yakir, L.; Mosesson, Y.; Harari, D.; Machluf, Y.; Asao, H.; Jovin, T.; Sugamura, K.; et al. Ligand-Independent Degradation of Epidermal Growth Factor Receptor Involves Receptor Ubiquitylation and Hgs, an Adaptor Whose Ubiquitin-Interacting Motif Targets Ubiquitylation by Nedd4. Traffic 2002, 3, 740–751. [Google Scholar] [CrossRef]
- Chesarino, N.M.; McMichael, T.M.; Yount, J.S. E3 Ubiquitin Ligase NEDD4 Promotes Influenza Virus Infection by Decreasing Levels of the Antiviral Protein IFITM3. PLoS Pathog. 2015, 11, e1005095. [Google Scholar] [CrossRef]
- Yan, C.; Zhao, M.; Li, S.; Liu, T.; Xu, C.; Liu, L.; Geng, T.; Gong, D. Increase of E3 ubiquitin ligase NEDD4 expression leads to degradation of its target proteins PTEN/IGF1R during the formation of goose fatty liver. J. Anim. Sci. 2020, 98, 270. [Google Scholar] [CrossRef]
- Sun, L.; Amraei, R.; Rahimi, N. NEDD4 regulates ubiquitination and stability of the cell adhesion molecule IGPR-1 via lysosomal pathway. J. Biomed. Sci. 2021, 28, 731. [Google Scholar] [CrossRef]
- Fukushima, T.; Yoshihara, H.; Furuta, H.; Kamei, H.; Hakuno, F.; Luan, J.; Duan, C.; Saeki, Y.; Tanaka, K.; Iemura, S.; et al. Nedd4-induced monoubiquitination of IRS-2 enhances IGF signalling and mitogenic activity. Nat. Commun. 2015, 6, 6780. [Google Scholar] [CrossRef]
- Hu, Y.; Hong, X.Y.; Yang, X.F.; Ma, R.H.; Wang, X.; Zhang, J.F.; Feng, Q.; Li, X.G.; Sun, D.S.; Li, X.; et al. Inflammation-dependent ISG15 upregulation mediates MIA-induced dendrite damages and depression by disrupting NEDD4/Rap2A signaling. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1477–1489. [Google Scholar] [CrossRef]
- Fan, J.-B.; Arimoto, K.-L.; Motamedchaboki, K.; Yan, M.; Wolf, D.A.; Zhang, D.-E. Identification and characterization of a novel ISG15-ubiquitin mixed chain and its role in regulating protein homeostasis. Sci. Rep. 2015, 5, 12704. [Google Scholar] [CrossRef] [PubMed]
- Fan, J. Investigating the Crosstalk between Nedd4 Ubiquitin Ligases and PIAS3 SUMO Ligase. Ph.D. Thesis, University of London, London, UK, 2017. [Google Scholar]
- Kumar, S.; Behera, A.; Saha, P.; Kumar Srivastava, A. The role of Krüppel-like factor 8 in cancer biology: Current research and its clinical relevance. Biochem. Pharm. 2021, 183, 114351. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cao, L.; Lin, J.T.; Yuan, Y.; Cao, Z.L.; Jia, J.D. Upregulated miRNA-1236-3p in osteosarcoma inhibits cell proliferation and induces apoptosis via targeting KLF8. Eur. Rev. Med. Pharm. Sci. 2019, 23, 6053–6061. [Google Scholar] [CrossRef]
- Salah, Z.; Cohen, S.; Itzhaki, E.; Aqeilan, R.I. NEDD4 E3 ligase inhibits the activity of the Hippo pathway by targeting LATS1 for degradation. Cell Cycle 2013, 12, 3817–3823. [Google Scholar] [CrossRef]
- Sun, A.; Wei, J.; Childress, C.; Shaw, J.H.t.; Peng, K.; Shao, G.; Yang, W.; Lin, Q. The E3 ubiquitin ligase NEDD4 is an LC3-interactive protein and regulates autophagy. Autophagy 2017, 13, 522–537. [Google Scholar] [CrossRef]
- Liu, Z.; Huo, X.; Zhao, S.; Yang, J.; Shi, W.; Jing, L.; Li, W.; Li, Y.; Ma, L.; Gao, Y.; et al. Low density lipoprotein receptor class A domain containing 4 (LDLRAD4) promotes tumorigenesis of hepatic cancer cells. Exp. Cell Res. 2017, 360, 189–198. [Google Scholar] [CrossRef]
- Ikeda, M.; Ikeda, A.; Longan, L.C.; Longnecker, R. The Epstein-Barr virus latent membrane protein 2A PY motif recruits WW domain-containing ubiquitin-protein ligases. Virology 2000, 268, 178–191. [Google Scholar] [CrossRef]
- Xu, C.; Fan, C.D.; Wang, X. Regulation of Mdm2 protein stability and the p53 response by NEDD4-1 E3 ligase. Oncogene 2015, 34, 281–289. [Google Scholar] [CrossRef]
- Sharma, P.; Murillas, R.; Zhang, H.; Kuehn, M.R. N4BP1 is a newly identified nucleolar protein that undergoes SUMO-regulated polyubiquitylation and proteasomal turnover at promyelocytic leukemia nuclear bodies. J. Cell Sci. 2010, 123, 1227–1234. [Google Scholar] [CrossRef]
- Murillas, R.; Simms, K.S.; Hatakeyama, S.; Weissman, A.M.; Kuehn, M.R. Identification of developmentally expressed proteins that functionally interact with Nedd4 ubiquitin ligase. J. Biol. Chem. 2002, 277, 2897–2907. [Google Scholar] [CrossRef]
- Hatstat, A.K.; Ahrendt, H.D.; Foster, M.W.; Mayne, L.; Moseley, M.A.; Englander, S.W.; McCafferty, D.G. Characterization of Small-Molecule-Induced Changes in Parkinson’s-Related Trafficking via the Nedd4 Ubiquitin Signaling Cascade. Cell Chem. Biol. 2021, 28, 14–25.e19. [Google Scholar] [CrossRef] [PubMed]
- Tardiff, D.F.; Jui, N.T.; Khurana, V.; Tambe, M.A.; Thompson, M.L.; Chung, C.Y.; Kamadurai, H.B.; Kim, H.T.; Lancaster, A.K.; Caldwell, K.A.; et al. Yeast Reveal a “Druggable” Rsp5/Nedd4 Network that Ameliorates -Synuclein Toxicity in Neurons. Science 2013, 342, 979–983. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Jin, S.; Wu, Y.; Xian, H.; Tian, S.; Liu, D.A.; Guo, Z.; Cui, J. Auto-ubiquitination of NEDD4-1 Recruits USP13 to Facilitate Autophagy through Deubiquitinating VPS34. Cell Rep. 2020, 30, 2807–2819.e2804. [Google Scholar] [CrossRef] [PubMed]
- Quirit, J.G.; Lavrenov, S.N.; Poindexter, K.; Xu, J.; Kyauk, C.; Durkin, K.A.; Aronchik, I.; Tomasiak, T.; Solomatin, Y.A.; Preobrazhenskaya, M.N.; et al. Indole-3-carbinol (I3C) analogues are potent small molecule inhibitors of NEDD4-1 ubiquitin ligase activity that disrupt proliferation of human melanoma cells. Biochem. Pharm. 2017, 127, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.Y.; Xu, N.; Malyukova, A.; Scarlett, C.J.; Sun, Y.T.; Zhang, X.D.; Ling, D.; Su, S.P.; Nelson, C.; Chang, D.K.; et al. The histone deacetylase SIRT2 stabilizes Myc oncoproteins. Cell Death Differ. 2013, 20, 503–514. [Google Scholar] [CrossRef]
- Sakata, T.; Sakaguchi, H.; Tsuda, L.; Higashitani, A.; Aigaki, T.; Matsuno, K.; Hayashi, S. Drosophila Nedd4 regulates endocytosis of notch and suppresses its ligand-independent activation. Curr. Biol. 2004, 14, 2228–2236. [Google Scholar] [CrossRef]
- Wilkin, M.B.; Carbery, A.M.; Fostier, M.; Aslam, H.; Mazaleyrat, S.L.; Higgs, J.; Myat, A.; Evans, D.A.; Cornell, M.; Baron, M. Regulation of notch endosomal sorting and signaling by Drosophila Nedd4 family proteins. Curr. Biol. 2004, 14, 2237–2244. [Google Scholar] [CrossRef]
- Koncarevic, A.; Jackman, R.W.; Kandarian, S.C. The ubiquitin-protein ligase Nedd4 targets Notch1 in skeletal muscle and distinguishes the subset of atrophies caused by reduced muscle tension. FASEB J. 2007, 21, 427–437. [Google Scholar] [CrossRef][Green Version]
- Wang, L.; Zhu, B.; Wang, S.; Wu, Y.; Zhan, W.; Xie, S.; Shi, H.; Yu, R. Regulation of glioma migration and invasion via modification of Rap2a activity by the ubiquitin ligase Nedd4-1. Oncol. Rep. 2017, 37, 2565–2574. [Google Scholar] [CrossRef]
- Zeng, T.; Wang, Q.; Fu, J.; Lin, Q.; Bi, J.; Ding, W.; Qiao, Y.; Zhang, S.; Zhao, W.; Lin, H.; et al. Impeded Nedd4-1-mediated Ras degradation underlies Ras-driven tumorigenesis. Cell Rep. 2014, 7, 871–882. [Google Scholar] [CrossRef]
- Hyndman, B.D.; Crupi, M.J.F.; Peng, S.; Bone, L.N.; Rekab, A.N.; Lian, E.Y.; Wagner, S.M.; Antonescu, C.N.; Mulligan, L.M. Differential recruitment of E3 ubiquitin ligase complexes regulates RET isoform internalization. J. Cell Sci. 2017, 130, 3282–3296. [Google Scholar] [CrossRef] [PubMed]
- Anindya, R.; Aygün, O.; Svejstrup, J.Q. Damage-induced ubiquitylation of human RNA polymerase II by the ubiquitin ligase Nedd4, but not Cockayne syndrome proteins or BRCA1. Mol Cell 2007, 28, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Harreman, M.; Taschner, M.; Sigurdsson, S.; Anindya, R.; Reid, J.; Somesh, B.; Kong, S.E.; Banks, C.A.; Conaway, R.C.; Conaway, J.W.; et al. Distinct ubiquitin ligases act sequentially for RNA polymerase II polyubiquitylation. Proc. Natl. Acad. Sci. USA 2009, 106, 20705–20710. [Google Scholar] [CrossRef] [PubMed]
- Canal, M.; Martín-Flores, N.; Pérez-Sisqués, L.; Romaní-Aumedes, J.; Altas, B.; Man, H.-Y.; Kawabe, H.; Alberch, J.; Malagelada, C. Loss of NEDD4 contributes to RTP801 elevation and neuron toxicity: Implications for Parkinson’s disease. Oncotarget 2016, 7, 58813–58831. [Google Scholar] [CrossRef]
- Zhou, W.; Xu, J.; Zhao, Y.; Sun, Y. SAG/RBX2 is a novel substrate of NEDD4-1 E3 ubiquitin ligase and mediates NEDD4-1 induced chemosensitization. Oncotarget 2014, 5, 6746–6755. [Google Scholar] [CrossRef]
- Aoh, Q.L.; Castle, A.M.; Hubbard, C.H.; Katsumata, O.; Castle, J.D. SCAMP3 Negatively Regulates Epidermal Growth Factor Receptor Degradation and Promotes Receptor Recycling. Mol. Biol. Cell 2009, 20, 1816–1832. [Google Scholar] [CrossRef]
- Al Sorkhy, M.; Craig, R.; Market, B.; Ard, R.; Porter, L.A. The cyclin-dependent kinase activator, Spy1A, is targeted for degradation by the ubiquitin ligase NEDD4. J. Biol. Chem. 2009, 284, 2617–2627. [Google Scholar] [CrossRef]
- Fifield, B.-A.; Qemo, I.; Kirou, E.; Cardiff, R.D.; Porter, L.A. The atypical cyclin-like protein Spy1 overrides p53-mediated tumour suppression and promotes susceptibility to breast tumourigenesis. Breast Cancer Res. 2019, 21, 1211. [Google Scholar] [CrossRef]
- Lin, Q.; Dai, Q.; Meng, H.; Sun, A.; Wei, J.; Peng, K.; Childress, C.; Chen, M.; Shao, G.; Yang, W. The HECT E3 ubiquitin ligase NEDD4 interacts with and ubiquitylates SQSTM1 for inclusion body autophagy. J. Cell Sci. 2017, 130, 3839–3850. [Google Scholar] [CrossRef]
- Murdaca, J.; Treins, C.; Monthouël-Kartmann, M.N.; Pontier-Bres, R.; Kumar, S.; Van Obberghen, E.; Giorgetti-Peraldi, S. Grb10 prevents Nedd4-mediated vascular endothelial growth factor receptor-2 degradation. J. Biol. Chem. 2004, 279, 26754–26761. [Google Scholar] [CrossRef]
- Bhalla, V.; Daidié, D.e.; Li, H.; Pao, A.C.; LaGrange, L.P.; Wang, J.; Vandewalle, A.; Stockand, J.D.; Staub, O.; Pearce, D. Serum- and Glucocorticoid-Regulated Kinase 1 Regulates Ubiquitin Ligase Neural Precursor Cell-Expressed, Developmentally Down-Regulated Protein 4-2 by Inducing Interaction with 14-3-3. Mol. Endocrinol. 2005, 19, 3073–3084. [Google Scholar] [CrossRef] [PubMed]
- Pohl, P.; Joshi, R.; Petrvalska, O.; Obsil, T.; Obsilova, V. 14-3-3-protein regulates Nedd4-2 by modulating interactions between HECT and WW domains. Commun. Biol. 2021, 4, 2419. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, K.; Foot, N.J.; Anand, S.; Dalton, H.E.; Chaudhary, N.; Collins, B.M.; Mathivanan, S.; Kumar, S. Regulation of the divalent metal ion transporter via membrane budding. Cell Discov. 2016, 2, 16011. [Google Scholar] [CrossRef] [PubMed]
- Shea, F.F.; Rowell, J.L.; Li, Y.; Chang, T.H.; Alvarez, C.E. Mammalian α arrestins link activated seven transmembrane receptors to Nedd4 family e3 ubiquitin ligases and interact with β arrestins. PLoS ONE 2012, 7, e50557. [Google Scholar] [CrossRef]
- Bhalla, V.; Oyster, N.M.; Fitch, A.C.; Wijngaarden, M.A.; Neumann, D.; Schlattner, U.; Pearce, D.; Hallows, K.R. AMP-activated Kinase Inhibits the Epithelial Na+ Channel through Functional Regulation of the Ubiquitin Ligase Nedd4-2*. J. Biol. Chem. 2006, 281, 26159–26169. [Google Scholar] [CrossRef]
- Hatanaka, T.; Hatanaka, Y.; Setou, M. Regulation of Amino Acid Transporter ATA2 by Ubiquitin Ligase Nedd4-2*. J. Biol. Chem. 2006, 281, 35922–35930. [Google Scholar] [CrossRef]
- Fu, L.; Rab, A.; Tang, L.; Bebok, Z.; Rowe, S.M.; Bartoszewski, R.; Collawn, J.F. ΔF508 CFTR surface stability is regulated by DAB2 and CHIP-mediated ubiquitination in post-endocytic compartments. PLoS ONE 2015, 10, e0123131. [Google Scholar] [CrossRef]
- Ahsan, M.K.; Figueroa-Hall, L.; Baratta, V.; Garcia-Milian, R.; Lam, T.T.; Hoque, K.; Salas, P.J.; Ameen, N.A. Glucocorticoids and serum- and glucocorticoid-inducible kinase 1 are potent regulators of CFTR in the native intestine: Implications for stress-induced diarrhea. Am. J. Physiol. Gastrointest Liver Physiol. 2020, 319, G121–G132. [Google Scholar] [CrossRef]
- Caohuy, H.; Jozwik, C.; Pollard, H.B. Rescue of ΔF508-CFTR by the SGK1/Nedd4-2 Signaling Pathway*. J. Biol. Chem. 2009, 284, 25241–25253. [Google Scholar] [CrossRef]
- Yamada, H.; Imajoh-Ohmi, S.; Haga, T. The high-affinity choline transporter CHT1 is regulated by the ubiquitin ligase Nedd4-2. Biomed. Res. 2012, 33, 1–8. [Google Scholar] [CrossRef]
- Hryciw, D.H.; Ekberg, J.; Lee, A.; Lensink, I.L.; Kumar, S.; Guggino, W.B.; Cook, D.I.; Pollock, C.A.; Poronnik, P. Nedd4-2 Functionally Interacts with ClC-5: Involvement in Constitutive Albumin Endocytosis in Proximal Tubule Cells*. J. Biol. Chem. 2004, 279, 54996–55007. [Google Scholar] [CrossRef] [PubMed]
- Nanami, M.; Pham, T.D.; Kim, Y.H.; Yang, B.; Sutliff, R.L.; Staub, O.; Klein, J.D.; Lopez-Cayuqueo, K.I.; Chambrey, R.; Park, A.Y.; et al. The Role of Intercalated Cell Nedd4-2 in BP Regulation, Ion Transport, and Transporter Expression. J. Am. Soc. Nephrol. 2018, 29, 1706–1719. [Google Scholar] [CrossRef] [PubMed]
- Arévalo, J.C. Nedd4-2 regulation of voltage-gated ion channels: An update on structure–function relationships and the pathophysiological consequences of dysfunction. J. Recept. Ligand Channel Res. 2015, 53, 52534. [Google Scholar] [CrossRef]
- Embark, H.M.; Böhmer, C.; Palmada, M.; Rajamanickam, J.; Wyatt, A.W.; Wallisch, S.; Capasso, G.; Waldegger, P.; Seyberth, H.W.; Waldegger, S.; et al. Regulation of CLC-Ka/barttin by the ubiquitin ligase Nedd4-2 and the serum- and glucocorticoid-dependent kinases. Kidney Int. 2004, 66, 1918–1925. [Google Scholar] [CrossRef]
- Jentsch, T.J.; Pusch, M. CLC Chloride Channels and Transporters: Structure, Function, Physiology, and Disease. Physiol. Rev. 2018, 98, 1493–1590. [Google Scholar] [CrossRef]
- Kim, Y.H.; Yoo, H.; Hong, A.R.; Kwon, M.; Kang, S.W.; Kim, K.; Song, Y. NEDD4L limits cAMP signaling through ubiquitination of CREB-regulated transcription coactivator 3. FASEB J. 2018, 32, 4053–4062. [Google Scholar] [CrossRef]
- Vina-Vilaseca, A.; Sorkin, A. Lysine 63-linked Polyubiquitination of the Dopamine Transporter Requires WW3 and WW4 Domains of Nedd4-2 and UBE2D Ubiquitin-conjugating Enzymes*. J. Biol. Chem. 2010, 285, 7645–7656. [Google Scholar] [CrossRef]
- Howitt, J.; Putz, U.; Lackovic, J.; Doan, A.; Dorstyn, L.; Cheng, H.; Yang, B.; Chan-Ling, T.; Silke, J.; Kumar, S.; et al. Divalent metal transporter 1 (DMT1) regulation by Ndfip1 prevents metal toxicity in human neurons. Proc. Natl. Acad. Sci. USA 2009, 106, 15489–15494. [Google Scholar] [CrossRef]
- Ding, Y.; Zhang, Y.; Xu, C.; Tao, Q.-H.; Chen, Y.-G. HECT Domain-containing E3 Ubiquitin Ligase NEDD4L Negatively Regulates Wnt Signaling by Targeting Dishevelled for Proteasomal Degradation. J. Biol. Chem. 2013, 288, 8289–8298. [Google Scholar] [CrossRef]
- Huang, X.; Chen, J.; Cao, W.; Yang, L.; Chen, Q.; He, J.; Yi, Q.; Huang, H.; Zhang, E.; Cai, Z. The many substrates and functions of NEDD4-1. Cell Death Dis. 2019, 10, 904. [Google Scholar] [CrossRef]
- Boehmer, C.; Henke, G.; Schniepp, R.; Palmada, M.; Rothstein, J.D.; Bröer, S.; Lang, F. Regulation of the glutamate transporter EAAT1 by the ubiquitin ligase Nedd4-2 and the serum and glucocorticoid-inducible kinase isoforms SGK1/3 and protein kinase B. J. Neurochem. 2003, 86, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- Boehmer, C.; Palmada, M.; Rajamanickam, J.; Schniepp, R.; Amara, S.; Lang, F. Post-translational regulation of EAAT2 function by co-expressed ubiquitin ligase Nedd4-2 is impacted by SGK kinases. J. Neurochem. 2006, 97, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, X.; Meng, X.; Wu, X.; Tong, H.; Zhang, X.; Qu, S. Regulation of glutamate transporter trafficking by Nedd4-2 in a Parkinson’s disease model. Cell Death Dis 2017, 8, e2574. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Lee, K.Y.; Jewett, K.A.; Man, H.-Y.; Chung, H.J.; Tsai, N.-P. Epilepsy-associated gene Nedd4-2 mediates neuronal activity and seizure susceptibility through AMPA receptors. PLoS Genet. 2017, 13, e1006634. [Google Scholar] [CrossRef]
- Albesa, M.; Grilo, L.S.; Gavillet, B.; Abriel, H. Nedd4-2-dependent ubiquitylation and regulation of the cardiac potassium channel hERG1. J. Mol. Cell Cardiol. 2011, 51, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Edinger, R.S.; Lebowitz, J.; Li, H.; Alzamora, R.; Wang, H.; Johnson, J.P.; Hallows, K.R. Functional Regulation of the Epithelial Na+ Channel by IκB Kinase-β Occurs via Phosphorylation of the Ubiquitin Ligase Nedd4-2*. J. Biol. Chem. 2009, 284, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Hallows, K.R.; Bhalla, V.; Oyster, N.M.; Wijngaarden, M.A.; Lee, J.K.; Li, H.; Chandran, S.; Xia, X.; Huang, Z.; Chalkley, R.J.; et al. Phosphopeptide Screen Uncovers Novel Phosphorylation Sites of Nedd4-2 That Potentiate Its Inhibition of the Epithelial Na+ Channel*. J. Biol. Chem. 2010, 285, 21671–21678. [Google Scholar] [CrossRef]
- Ekberg, J.; Schuetz, F.; Boase, N.A.; Conroy, S.-J.; Manning, J.; Kumar, S.; Poronnik, P.; Adams, D.J. Regulation of the Voltage-gated K+ Channels KCNQ2/3 and KCNQ3/5 by Ubiquitination: Novel Role for Nedd4-2*. J. Biol. Chem. 2007, 282, 12135–12142. [Google Scholar] [CrossRef]
- Jespersen, T.; Membrez, M.; Nicolas, C.S.; Pitard, B.; Staub, O.; Olesen, S.-P.; Baró, I.; Abriel, H. The KCNQ1 potassium channel is down-regulated by ubiquitylating enzymes of the Nedd4/Nedd4-like family. Cardiovasc. Res. 2007, 74, 64–74. [Google Scholar] [CrossRef]
- Alzamora, R.; Gong, F.; Rondanino, C.; Lee, J.K.; Smolak, C.; Pastor-Soler, N.M.; Hallows, K.R. AMP-activated protein kinase inhibits KCNQ1 channels through regulation of the ubiquitin ligase Nedd4-2 in renal epithelial cells. Am. J. Physiol. Ren. Physiol. 2010, 299, F1308–F1319. [Google Scholar] [CrossRef]
- Rougier, J.-S.; Bemmelen, M.X.v.; Bruce, M.C.; Jespersen, T.; Gavillet, B.; Apothéloz, F.; Cordonier, S.; Staub, O.; Rotin, D.; Abriel, H. Molecular determinants of voltage-gated sodium channel regulation by the Nedd4/Nedd4-like proteins. Am. J. Physiol.-Cell Physiol. 2005, 288, C692–C701. [Google Scholar] [CrossRef] [PubMed]
- Laedermann, C.J.; Cachemaille, M.; Kirschmann, G.; Pertin, M.; Gosselin, R.D.; Chang, I.; Albesa, M.; Towne, C.; Schneider, B.L.; Kellenberger, S.; et al. Dysregulation of voltage-gated sodium channels by ubiquitin ligase NEDD4-2 in neuropathic pain. J. Clin. Invest. 2013, 123, 3002–3013. [Google Scholar] [CrossRef] [PubMed]
- Gasser, A.; Cheng, X.; Gilmore, E.S.; Tyrrell, L.; Waxman, S.G.; Dib-Hajj, S.D. Two Nedd4-binding Motifs Underlie Modulation of Sodium Channel Nav1.6 by p38 MAPK*. J. Biol. Chem. 2010, 285, 26149–26161. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.P.; Lagnaz, D.; Ronzaud, C.; Vázquez, N.; Ko, B.S.; Moddes, L.; Ruffieux-Daidié, D.; Hausel, P.; Koesters, R.; Yang, B.; et al. Nedd4-2 modulates renal Na+-Cl- cotransporter via the aldosterone-SGK1-Nedd4-2 pathway. J. Am. Soc. Nephrol. 2011, 22, 1707–1719. [Google Scholar] [CrossRef]
- Rotin, D.; Staub, O. Nedd4-2 and the regulation of epithelial sodium transport. Front. Physiol. 2012, 3, 212. [Google Scholar] [CrossRef]
- Bruce, M.C.; Kanelis, V.; Fouladkou, F.; Debonneville, A.; Staub, O.; Rotin, D. Regulation of Nedd4-2 self-ubiquitination and stability by a PY motif located within its HECT-domain. Biochem. J. 2008, 415, 155–163. [Google Scholar] [CrossRef]
- Escobedo, A.; Gomes, T.; Aragón, E.; Martín-Malpartida, P.; Ruiz, L.; Macias, M.J. Structural basis of the activation and degradation mechanisms of the E3 ubiquitin ligase Nedd4L. Structure 2014, 22, 1446–1457. [Google Scholar] [CrossRef]
- Zhao, C.; Jia, M.; Song, H.; Yu, Z.; Wang, W.; Li, Q.; Zhang, L.; Zhao, W.; Cao, X. The E3 Ubiquitin Ligase TRIM40 Attenuates Antiviral Immune Responses by Targeting MDA5 and RIG-I. Cell Rep. 2017, 21, 1613–1623. [Google Scholar] [CrossRef]
- No, Y.R.; He, P.; Yoo, B.K.; Yun, C.C. Unique Regulation of Human Na+/H+ Exchanger 3 (NHE3) by Nedd4-2 Ligase That Differs from Non-primate NHE3s. J. Biol. Chem. 2014, 289, 18360–18372. [Google Scholar] [CrossRef]
- Xu, D.; Wang, H.; You, G. An Essential Role of Nedd4-2 in the Ubiquitination, Expression, and Function of Organic Anion Transporter-3. Mol. Pharm. 2016, 13, 621–630. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, J.; Zhang, Q.; Fan, Y.; Liu, C.; You, G. PKC/Nedd4-2 Signaling Pathway Regulates the Cell Surface Expression of Drug Transporter hOAT1. Drug Metab. Dispos. 2017, 45, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Raikwar, N.S.; Vandewalle, A.; Thomas, C.P. Nedd4–2 interacts with occludin to inhibit tight junction formation and enhance paracellular conductance in collecting duct epithelia. Am. J. Physiol.-Ren. Physiol. 2010, 299, F436–F444. [Google Scholar] [CrossRef] [PubMed]
- Snyder, P.M.; Olson, D.R.; Kabra, R.; Zhou, R.; Steines, J.C. cAMP and serum and glucocorticoid-inducible kinase (SGK) regulate the epithelial Na(+) channel through convergent phosphorylation of Nedd4-2. J. Biol. Chem. 2004, 279, 45753–45758. [Google Scholar] [CrossRef]
- Wiemuth, D.; Lott, J.S.; Ly, K.; Ke, Y.; Teesdale-Spittle, P.; Snyder, P.M.; McDonald, F.J. Interaction of Serum- and Glucocorticoid Regulated Kinase 1 (SGK1) with the WW-Domains of Nedd4-2 Is Required for Epithelial Sodium Channel Regulation. PLoS ONE 2010, 5, e12163. [Google Scholar] [CrossRef] [PubMed]
- Ingham, R.J.; Gish, G.; Pawson, T. The Nedd4 family of E3 ubiquitin ligases: Functional diversity within a common modular architecture. Oncogene 2004, 23, 1972–1984. [Google Scholar] [CrossRef]
- Dieter, M.; Palmada, M.; Rajamanickam, J.; Aydin, A.; Busjahn, A.; Boehmer, C.; Luft, F.C.; Lang, F. Regulation of Glucose Transporter SGLT1 by Ubiquitin Ligase Nedd4-2 and Kinases SGK1, SGK3, and PKB. Obes. Res. 2004, 12, 862–870. [Google Scholar] [CrossRef]
- Inoue, Y.; Imamura, T. Regulation of TGF-β family signaling by E3 ubiquitin ligases. Cancer Sci. 2008, 99, 2107–2112. [Google Scholar] [CrossRef]
- Morén, A.; Imamura, T.; Miyazono, K.; Heldin, C.-H.; Moustakas, A. Degradation of the Tumor Suppressor Smad4 by WW and HECT Domain Ubiquitin Ligases. J. Biol. Chem. 2005, 280, 22115–22123. [Google Scholar] [CrossRef]
- Kuratomi, G.; Komuro, A.; Goto, K.; Shinozaki, M.; Miyazawa, K.; Miyazono, K.; Imamura, T. NEDD4-2 (neural precursor cell expressed, developmentally down-regulated 4-2) negatively regulates TGF-β (transforming growth factor-β) signalling by inducing ubiquitin-mediated degradation of Smad2 and TGF-β type I receptor. Biochem. J. 2005, 386, 461–470. [Google Scholar] [CrossRef]
- Gao, S.; Alarcón, C.; Sapkota, G.; Rahman, S.; Chen, P.-Y.; Goerner, N.; Macias, M.J.; Erdjument-Bromage, H.; Tempst, P.; Massagué, J. Ubiquitin Ligase Nedd4L Targets Activated Smad2/3 to Limit TGF-β Signaling. Mol. Cell 2009, 36, 457–468. [Google Scholar] [CrossRef]
- Conkright, J.J.; Apsley, K.S.; Martin, E.P.; Ridsdale, R.; Rice, W.R.; Na, C.-L.; Yang, B.; Weaver, T.E. Nedd4-2–Mediated Ubiquitination Facilitates Processing of Surfactant Protein–C. Am. J. Respir. Cell Mol. Biol. 2010, 42, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Feng, Z.; Liao, Y.; Yang, M.; Bai, Y.; He, Z. Diminution of microRNA-98 alleviates renal fibrosis in diabetic nephropathy by elevating Nedd4L and inactivating TGF-β/Smad2/3 pathway. Cell Cycle 2020, 19, 3406–3418. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Sánchez, J.; Arévalo, J. A Review on Ubiquitination of Neurotrophin Receptors: Facts and Perspectives. Int. J. Mol. Sci. 2017, 18, 630. [Google Scholar] [CrossRef] [PubMed]
- Arévalo, J.C.; Waite, J.; Rajagopal, R.; Beyna, M.; Chen, Z.-Y.; Lee, F.S.; Chao, M.V. Cell Survival through Trk Neurotrophin Receptors Is Differentially Regulated by Ubiquitination. Neuron 2006, 50, 549–559. [Google Scholar] [CrossRef]
- Georgieva, M.V.; de Pablo, Y.; Sanchis, D.; Comella, J.X.; Llovera, M. Ubiquitination of TrkA by Nedd4-2 regulates receptor lysosomal targeting and mediates receptor signaling. J. Neurochem. 2011, 117, 479–493. [Google Scholar] [CrossRef]
- He, Y.; Hryciw, D.H.; Carroll, M.L.; Myers, S.A.; Whitbread, A.K.; Kumar, S.; Poronnik, P.; Hooper, J.D. The Ubiquitin-Protein Ligase Nedd4-2 Differentially Interacts with and Regulates Members of the Tweety Family of Chloride Ion Channels*. J. Biol. Chem. 2008, 283, 24000–24010. [Google Scholar] [CrossRef]
- Nazio, F.; Carinci, M.; Valacca, C.; Bielli, P.; Strappazzon, F.; Antonioli, M.; Ciccosanti, F.; Rodolfo, C.; Campello, S.; Fimia, G.M.; et al. Fine-tuning of ULK1 mRNA and protein levels is required for autophagy oscillation. J. Cell Biol. 2016, 215, 841–856. [Google Scholar] [CrossRef]
- Oberfeld, B.; Ruffieux-Daidié, D.; Vitagliano, J.-J.; Pos, K.M.; Verrey, F.; Staub, O. Ubiquitin-specific protease 2-45 (Usp2-45) binds to epithelial Na+ channel (ENaC)-ubiquitylating enzyme Nedd4-2. Am. J. Physiol.-Ren. Physiol. 2011, 301, F189–F196. [Google Scholar] [CrossRef]
- Roy, A.; Al-Qusairi, L.; Donnelly, B.F.; Ronzaud, C.; Marciszyn, A.L.; Gong, F.; Chang, Y.P.C.; Butterworth, M.B.; Pastor-Soler, N.M.; Hallows, K.R.; et al. Alternatively spliced proline-rich cassettes link WNK1 to aldosterone action. J. Clin. Investig. 2015, 125, 3433–3448. [Google Scholar] [CrossRef]
- Lin, Q.; Wang, J.; Childress, C.; Sudol, M.; Carey, D.J.; Yang, W. HECT E3 ubiquitin ligase Nedd4-1 ubiquitinates ACK and regulates epidermal growth factor (EGF)-induced degradation of EGF receptor and ACK. Mol. Cell Biol. 2010, 30, 1541–1554. [Google Scholar] [CrossRef]
- Chan, W.; Tian, R.; Lee, Y.-F.; Sit, S.T.; Lim, L.; Manser, E. Down-regulation of Active ACK1 Is Mediated by Association with the E3 Ubiquitin Ligase Nedd4-2*. J. Biol. Chem. 2009, 284, 8185–8194. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.D.; Lum, M.A.; Xu, C.; Black, J.D.; Wang, X. Ubiquitin-dependent regulation of phospho-AKT dynamics by the ubiquitin E3 ligase, NEDD4-1, in the insulin-like growth factor-1 response. J. Biol. Chem. 2013, 288, 1674–1684. [Google Scholar] [CrossRef] [PubMed]
- Persaud, A.; Alberts, P.; Amsen, E.M.; Xiong, X.; Wasmuth, J.; Saadon, Z.; Fladd, C.; Parkinson, J.; Rotin, D. Comparison of substrate specificity of the ubiquitin ligases Nedd4 and Nedd4-2 using proteome arrays. Mol. Syst. Biol. 2009, 5, 333. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.-H.; Dinudom, A.; Sanchez-Perez, A.; Kumar, S.; Cook, D.I. Akt Mediates the Effect of Insulin on Epithelial Sodium Channels by Inhibiting Nedd4-2*. J. Biol. Chem. 2007, 282, 29866–29873. [Google Scholar] [CrossRef] [PubMed]
- Scudder, S.L.; Goo, M.S.; Cartier, A.E.; Molteni, A.; Schwarz, L.A.; Wright, R.; Patrick, G.N. Synaptic strength is bidirectionally controlled by opposing activity-dependent regulation of Nedd4-1 and USP8. J. Neurosci. Off. J. Soc. Neurosci. 2014, 34, 16637–16649. [Google Scholar] [CrossRef]
- Schwarz, L.A.; Hall, B.J.; Patrick, G.N. Activity-dependent ubiquitination of GluA1 mediates a distinct AMPA receptor endocytosis and sorting pathway. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 16718–16729. [Google Scholar] [CrossRef]
- Lin, A.; Hou, Q.; Jarzylo, L.; Amato, S.; Gilbert, J.; Shang, F.; Man, H.-Y. Nedd4-mediated AMPA receptor ubiquitination regulates receptor turnover and trafficking. J. Neurochem. 2011, 119, 27–39. [Google Scholar] [CrossRef]
- Kim, J.E.; Lee, D.S.; Park, H.; Kim, T.H.; Kang, T.C. AMPA Receptor Antagonists Facilitate NEDD4-2-Mediated GRIA1 Ubiquitination by Regulating PP2B-ERK1/2-SGK1 Pathway in Chronic Epilepsy Rats. Biomedicines 2021, 9, 1069. [Google Scholar] [CrossRef]
- Jewett, K.A.; Zhu, J.; Tsai, N.P. The tumor suppressor p53 guides GluA1 homeostasis through Nedd4-2 during chronic elevation of neuronal activity. J. Neurochem. 2015, 135, 226–233. [Google Scholar] [CrossRef]
- Trimpert, C.; Wesche, D.; de Groot, T.; Pimentel Rodriguez, M.M.; Wong, V.; van den Berg, D.T.M.; Cheval, L.; Ariza, C.A.; Doucet, A.; Stagljar, I.; et al. NDFIP allows NEDD4/NEDD4L-induced AQP2 ubiquitination and degradation. PLoS ONE 2017, 12, e0183774. [Google Scholar] [CrossRef]
- Claude; Eitelhuber, A.; Christian; Giallonardo, P.; Gegg, M.; Oller, H.; Seth; Krappmann, D.; Ueffing, M.; Lickert, H. Dlg3 Trafficking and Apical Tight Junction Formation Is Regulated by Nedd4 and Nedd4-2 E3 Ubiquitin Ligases. Dev. Cell 2011, 21, 479–491. [Google Scholar] [CrossRef]
- Novellasdemunt, L.; Kucharska, A.; Jamieson, C.; Prange-Barczynska, M.; Baulies, A.; Antas, P.; van der Vaart, J.; Gehart, H.; Maurice, M.M.; Li, V.S. NEDD4 and NEDD4L regulate Wnt signalling and intestinal stem cell priming by degrading LGR5 receptor. EMBO J. 2020, 39, e102771. [Google Scholar] [CrossRef] [PubMed]
- Dinudom, A.; Harvey, K.F.; Komwatana, P.; Young, J.A.; Kumar, S.; Cook, D.I. Nedd4 mediates control of an epithelial Na+ channel in salivary duct cells by cytosolic Na+. Proc. Natl. Acad. Sci. USA 1998, 95, 7169–7173. [Google Scholar] [CrossRef] [PubMed]
- Staub, O.; Dho, S.; Henry, P.; Correa, J.; Ishikawa, T.; McGlade, J.; Rotin, D. WW domains of Nedd4 bind to the proline-rich PY motifs in the epithelial Na+ channel deleted in Liddle’s syndrome. EMBO J. 1996, 15, 2371–2380. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Patel, S.V.; Snyder, P.M. Nedd4-2 catalyzes ubiquitination and degradation of cell surface ENaC. J. Biol. Chem. 2007, 282, 20207–20212. [Google Scholar] [CrossRef]
- Kanelis, V.; Rotin, D.; Forman-Kay, J.D. Solution structure of a Nedd4 WW domain-ENaC peptide complex. Nat. Struct. Biol. 2001, 8, 407–412. [Google Scholar] [CrossRef]
- Monami, G.; Emiliozzi, V.; Morrione, A. Grb10/Nedd4-mediated multiubiquitination of the insulin-like growth factor receptor regulates receptor internalization. J. Cell Physiol. 2008, 216, 426–437. [Google Scholar] [CrossRef]
- Nakayama, M.; Berger, P. Coordination of VEGF receptor trafficking and signaling by coreceptors. Exp. Cell Res. 2013, 319, 1340–1347. [Google Scholar] [CrossRef]
- Cheng, W.; Huang, P.C.; Chao, H.M.; Jeng, Y.M.; Hsu, H.C.; Pan, H.W.; Hwu, W.L.; Lee, Y.M. Glypican-3 induces oncogenicity by preventing IGF-1R degradation, a process that can be blocked by Grb10. Oncotarget 2017, 8, 80429–80442. [Google Scholar] [CrossRef]
- Vélez, P.; Schwartz, A.B.; Iyer, S.R.; Warrington, A.; Fadool, D.A. Ubiquitin ligase Nedd4-2 modulates Kv1.3 current amplitude and ion channel protein targeting. J. Neurophysiol. 2016, 116, 671–685. [Google Scholar] [CrossRef]
- Plant, P.J.; Correa, J.; Goldenberg, N.; Bain, J.; Batt, J. The inositol phosphatase MTMR4 is a novel target of the ubiquitin ligase Nedd4. Biochem. J. 2009, 419, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, P.; Plant, P.J.; Correa, J.; Bain, A.; Takeda, M.; Kawabe, H.; Rotin, D.; Bain, J.R.; Batt, J.A.E. The ubiquitin ligase Nedd4-1 participates in denervation-induced skeletal muscle atrophy in mice. PLoS ONE 2012, 7, e46427. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Sala, L.; Mura, M.; Rocchetti, M.; Pedrazzini, M.; Ran, X.; Mak, T.S.H.; Crotti, L.; Sham, P.C.; Torre, E.; et al. MTMR4 SNVs modulate ion channel degradation and clinical severity in congenital long QT syndrome: Insights in the mechanism of action of protective modifier genes. Cardiovasc. Res. 2021, 117, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Fotia, A.B.; Dinudom, A.; Shearwin, K.E.; Koch, J.-P.; Korbmacher, C.; Cook, D.I.; Kumar, S. The role of individual Nedd4–2 (KIAA0439) WW domains in binding and regulating epithelial sodium channels. FASEB J. 2003, 17, 70–72. [Google Scholar] [CrossRef] [PubMed]
- Howitt, J.; Lackovic, J.; Low, L.H.; Naguib, A.; Macintyre, A.; Goh, C.P.; Callaway, J.K.; Hammond, V.; Thomas, T.; Dixon, M.; et al. Ndfip1 regulates nuclear Pten import in vivo to promote neuronal survival following cerebral ischemia. J. Cell Biol. 2012, 196, 29–36. [Google Scholar] [CrossRef]
- Trotman, L.C.; Wang, X.; Alimonti, A.; Chen, Z.; Teruya-Feldstein, J.; Yang, H.; Pavletich, N.P.; Carver, B.S.; Cordon-Cardo, C.; Erdjument-Bromage, H.; et al. Ubiquitination regulates PTEN nuclear import and tumor suppression. Cell 2007, 128, 141–156. [Google Scholar] [CrossRef]
- Wang, X.; Trotman, L.C.; Koppie, T.; Alimonti, A.; Chen, Z.; Gao, Z.; Wang, J.; Erdjument-Bromage, H.; Tempst, P.; Cordon-Cardo, C.; et al. NEDD4-1 is a proto-oncogenic ubiquitin ligase for PTEN. Cell 2007, 128, 129–139. [Google Scholar] [CrossRef]
- Amodio, N.; Scrima, M.; Palaia, L.; Salman, A.N.; Quintiero, A.; Franco, R.; Botti, G.; Pirozzi, P.; Rocco, G.; De Rosa, N.; et al. Oncogenic role of the E3 ubiquitin ligase NEDD4-1, a PTEN negative regulator, in non-small-cell lung carcinomas. Am. J. Pathol. 2010, 177, 2622–2634. [Google Scholar] [CrossRef]
- Drinjakovic, J.; Jung, H.; Campbell, D.S.; Strochlic, L.; Dwivedy, A.; Holt, C.E. E3 ligase Nedd4 promotes axon branching by downregulating PTEN. Neuron 2010, 65, 341–357. [Google Scholar] [CrossRef]
- Fouladkou, F.; Landry, T.; Kawabe, H.; Neeb, A.; Lu, C.; Brose, N.; Stambolic, V.; Rotin, D. The ubiquitin ligase Nedd4-1 is dispensable for the regulation of PTEN stability and localization. Proc. Natl. Acad. Sci. USA 2008, 105, 8585–8590. [Google Scholar] [CrossRef]
- Hsia, H.-E.; Kumar, R.; Luca, R.; Takeda, M.; Courchet, J.; Nakashima, J.; Wu, S.; Goebbels, S.; An, W.; Eickholt, B.J.; et al. Ubiquitin E3 ligase Nedd4-1 acts as a downstream target of PI3K/PTEN-mTORC1 signaling to promote neurite growth. Proc. Natl. Acad. Sci. USA 2014, 111, 13205. [Google Scholar] [CrossRef] [PubMed]
- Giasson, B.I.; Murray, I.V.J.; Trojanowski, J.Q.; Lee, V.M.Y. A Hydrophobic Stretch of 12 Amino Acid Residues in the Middle of α-Synuclein Is Essential for Filament Assembly. J. Biol. Chem. 2001, 276, 2380–2386. [Google Scholar] [CrossRef] [PubMed]
- Conway, K.A.; Harper, J.D.; Lansbury, P.T. Accelerated in vitro fibril formation by a mutant α-synuclein linked to early-onset Parkinson disease. Nat. Med. 1998, 4, 1318–1320. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.Y.; Khurana, V.; Auluck, P.K.; Tardiff, D.F.; Mazzulli, J.R.; Soldner, F.; Baru, V.; Lou, Y.; Freyzon, Y.; Cho, S.; et al. Identification and Rescue of -Synuclein Toxicity in Parkinson Patient-Derived Neurons. Science 2013, 342, 983–987. [Google Scholar] [CrossRef] [PubMed]
- Itier, J.M. Parkin gene inactivation alters behaviour and dopamine neurotransmission in the mouse. Hum. Mol. Genet. 2003, 12, 2277–2291. [Google Scholar] [CrossRef]
- Dickins, R.A.; Frew, I.J.; House, C.M.; O’Bryan, M.K.; Holloway, A.J.; Haviv, I.; Traficante, N.; De Kretser, D.M.; Bowtell, D.D.L. The Ubiquitin Ligase Component Siah1a Is Required for Completion of Meiosis I in Male Mice. Mol. Cell. Biol. 2002, 22, 2294–2303. [Google Scholar] [CrossRef]
- Frew, I.J.; Dickins, R.A.; Cuddihy, A.R.; Del Rosario, M.; Reinhard, C.; O’Connell, M.J.; Bowtell, D.D.L. Normal p53 Function in Primary Cells Deficient for Siah Genes. Mol. Cell. Biol. 2002, 22, 8155–8164. [Google Scholar] [CrossRef]
- Frew, I.J.; Hammond, V.E.; Dickins, R.A.; Quinn, J.M.W.; Walkley, C.R.; Sims, N.A.; Schnall, R.; Della, N.G.; Holloway, A.J.; Digby, M.R.; et al. Generation and Analysis of Siah2 Mutant Mice. Mol. Cell. Biol. 2003, 23, 9150–9161. [Google Scholar] [CrossRef]
- Schisler, J.C.; Rubel, C.E.; Zhang, C.; Lockyer, P.; Cyr, D.M.; Patterson, C. CHIP protects against cardiac pressure overload through regulation of AMPK. J. Clin. Investig. 2013, 123, 3588–3599. [Google Scholar] [CrossRef]
- Moroishi, T.; Nishiyama, M.; Takeda, Y.; Iwai, K.; Keiichi. The FBXL5-IRP2 Axis Is Integral to Control of Iron Metabolism In Vivo. Cell Metab. 2011, 14, 339–351. [Google Scholar] [CrossRef]
- Perry, W.L.; Hustad, C.M.; Swing, D.A.; O’Sullivan, T.N.; Jenkins, N.A.; Copeland, N.G. The itchy locus encodes a novel ubiquitin protein ligase that is disrupted in a18H mice. Nat. Genet. 1998, 18, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Ying, S.-X.; Zhang, G.-M.; Li, C.; Cheng, S.Y.; Deng, C.-X.; Zhang, Y.E. Ubiquitin Ligase Smurf1 Controls Osteoblast Activity and Bone Homeostasis by Targeting MEKK2 for Degradation. Cell 2005, 121, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Narimatsu, M.; Bose, R.; Pye, M.; Zhang, L.; Miller, B.; Ching, P.; Sakuma, R.; Luga, V.; Roncari, L.; Attisano, L.; et al. Regulation of Planar Cell Polarity by Smurf Ubiquitin Ligases. Cell 2009, 137, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Zhi, X.; Zhou, Z.; Chen, C. TAZ antagonizes the WWP1-mediated KLF5 degradation and promotes breast cell proliferation and tumorigenesis. Carcinogenesis 2012, 33, 59–67. [Google Scholar] [CrossRef]
- Zou, W.; Chen, X.; Shim, J.-H.; Huang, Z.; Brady, N.; Hu, D.; Drapp, R.; Sigrist, K.; Glimcher, L.H.; Jones, D. The E3 ubiquitin ligase Wwp2 regulates craniofacial development through mono-ubiquitylation of Goosecoid. Nat. Cell Biol. 2011, 13, 59–65. [Google Scholar] [CrossRef]
- Kawabe, H.; Neeb, A.; Dimova, K.; Young, S.M., Jr.; Takeda, M.; Katsurabayashi, S.; Mitkovski, M.; Malakhova, O.A.; Zhang, D.E.; Umikawa, M.; et al. Regulation of Rap2A by the ubiquitin ligase Nedd4-1 controls neurite development. Neuron 2010, 65, 358–372. [Google Scholar] [CrossRef]
- Boase, N.A.; Rychkov, G.Y.; Townley, S.L.; Dinudom, A.; Candi, E.; Voss, A.K.; Tsoutsman, T.; Semsarian, C.; Melino, G.; Koentgen, F.; et al. Respiratory distress and perinatal lethality in Nedd4-2-deficient mice. Nat. Commun. 2011, 2, 287. [Google Scholar] [CrossRef]
- Leitz, D.; Duerr, J.; Dvornikov, D.; Szczygiel, M.; Konietzke, P.; Rotin, D.; Wielpütz, M.; Kawabe, H.; Beers, M.; Klingmüller, U.; et al. Development of pulmonary fibrosis in conditional Nedd4-2 deficient mice. Eur. Respir. J. 2018, 52, OA5364. [Google Scholar] [CrossRef]
- Frank, C.; Pari, G.; Rossiter, J.P. Approach to diagnosis of Parkinson disease. Can. Fam. Phys. 2006, 52, 862–868. [Google Scholar]
- Nalls, M.A.; Blauwendraat, C.; Vallerga, C.L.; Heilbron, K.; Bandres-Ciga, S.; Chang, D.; Tan, M.; Kia, D.A.; Noyce, A.J.; Xue, A.; et al. Identification of novel risk loci, causal insights, and heritable risk for Parkinson’s disease: A meta-analysis of genome-wide association studies. Lancet Neurol. 2019, 18, 1091–1102. [Google Scholar] [CrossRef]
- Foo, J.N.; Chew, E.G.Y.; Chung, S.J.; Peng, R.; Blauwendraat, C.; Nalls, M.A.; Mok, K.Y.; Satake, W.; Toda, T.; Chao, Y.; et al. Identification of Risk Loci for Parkinson Disease in Asians and Comparison of Risk between Asians and Europeans. JAMA Neurol. 2020, 77, 746. [Google Scholar] [CrossRef] [PubMed]
- Putz, U.; Howitt, J.; Lackovic, J.; Foot, N.; Kumar, S.; Silke, J.; Tan, S.S. Nedd4 family-interacting protein 1 (Ndfip1) is required for the exosomal secretion of Nedd4 family proteins. J. Biol. Chem. 2008, 283, 32621–32627. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Harvey, K.F.; Kinoshita, M.; Copeland, N.G.; Noda, M.; Jenkins, N.A. cDNA cloning, expression analysis, and mapping of the mouse Nedd4 gene. Genomics 1997, 40, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ross, C.A.; Wang, N.; Huo, Y.; Mackinnon, D.F.; Potash, J.B.; Simpson, S.G.; McMahon, F.J.; Depaulo, J.J.R.; McInnis, M.G. NEDD4L on human chromosome 18q21 has multiple forms of transcripts and is a homologue of the mouse Nedd4-2 gene. Eur. J. Hum. Genet. 2001, 9, 922–930. [Google Scholar] [CrossRef]
- Itani, O.A.; Campbell, J.R.; Herrero, J.; Snyder, P.M.; Thomas, C.P. Alternate promoters and variable splicing lead to hNedd4-2 isoforms with a C2 domain and varying number of WW domains. Am. J. Physiol. Ren. Physiol. 2003, 285, F916–F929. [Google Scholar] [CrossRef]
- Itani, O.A.; Stokes, J.B.; Thomas, C.P. Nedd4-2 isoforms differentially associate with ENaC and regulate its activity. Am. J. Physiol. Ren. Physiol. 2005, 289, F334–F346. [Google Scholar] [CrossRef]
- Ronzaud, C.; Loffing-Cueni, D.; Hausel, P.; Debonneville, A.; Malsure, S.R.; Fowler-Jaeger, N.; Boase, N.A.; Perrier, R.; Maillard, M.; Yang, B.; et al. Renal tubular NEDD4-2 deficiency causes NCC-mediated salt-dependent hypertension. J. Clin. Investig. 2013, 2013, 61110. [Google Scholar] [CrossRef]
- Harvey, K.F.; Dinudom, A.; Cook, D.I.; Kumar, S. The Nedd4-like protein KIAA0439 is a potential regulator of the epithelial sodium channel. J. Biol. Chem. 2001, 276, 8597–8601. [Google Scholar] [CrossRef]
- Kumar, S.; Tomooka, Y.; Noda, M. Identification of a set of genes with developmentally down-regulated expression in the mouse brain. Biochem. Biophys. Res. Commun. 1992, 185, 1155–1161. [Google Scholar] [CrossRef]
- Fagerberg, L.; Hallström, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the Human Tissue-specific Expression by Genome-wide Integration of Transcriptomics and Antibody-based Proteomics. Mol. Cell. Proteom. 2014, 13, 397–406. [Google Scholar] [CrossRef]
- Dumitriu, A.; Latourelle, J.C.; Hadzi, T.C.; Pankratz, N.; Garza, D.; Miller, J.P.; Vance, J.M.; Foroud, T.; Beach, T.G.; Myers, R.H. Gene Expression Profiles in Parkinson Disease Prefrontal Cortex Implicate FOXO1 and Genes under Its Transcriptional Regulation. PLoS Genet. 2012, 8, e1002794. [Google Scholar] [CrossRef] [PubMed]
- Plant, P.J.; Yeger, H.; Staub, O.; Howard, P.; Rotin, D. The C2 Domain of the Ubiquitin Protein Ligase Nedd4 Mediates Ca2+-dependent Plasma Membrane Localization. J. Biol. Chem. 1997, 272, 32329–32336. [Google Scholar] [CrossRef] [PubMed]
- Goel, P.; Manning, J.A.; Kumar, S. NEDD4-2 (NEDD4L): The ubiquitin ligase for multiple membrane proteins. Gene 2015, 557, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Donovan, P.; Poronnik, P. Nedd4 and Nedd4-2: Ubiquitin ligases at work in the neuron. Int. J. Biochem. Cell Biol. 2013, 45, 706–710. [Google Scholar] [CrossRef] [PubMed]
- Fotia, A.B.; Cook, D.I.; Kumar, S. The ubiquitin-protein ligases Nedd4 and Nedd4-2 show similar ubiquitin-conjugating enzyme specificities. Int. J. Biochem. Cell Biol. 2006, 38, 472–479. [Google Scholar] [CrossRef]
- De Bie, P.; Ciechanover, A. Ubiquitination of E3 ligases: Self-regulation of the ubiquitin system via proteolytic and non-proteolytic mechanisms. Cell Death Differ. 2011, 18, 1393–1402. [Google Scholar] [CrossRef]
- Mari, S.; Ruetalo, N.; Maspero, E.; Stoffregen, M.C.; Pasqualato, S.; Polo, S.; Wiesner, S. Structural and functional framework for the autoinhibition of Nedd4-family ubiquitin ligases. Structure 2014, 22, 1639–1649. [Google Scholar] [CrossRef]
- Wang, J.; Peng, Q.; Lin, Q.; Childress, C.; Carey, D.; Yang, W. Calcium activates Nedd4 E3 ubiquitin ligases by releasing the C2 domain-mediated auto-inhibition. J. Biol. Chem. 2010, 285, 12279–12288. [Google Scholar] [CrossRef]
- Mund, T.; Pelham, H.R.B. Control of the activity of WW-HECT domain E3 ubiquitin ligases by NDFIP proteins. EMBO Rep. 2009, 10, 501–507. [Google Scholar] [CrossRef]
- Song, L.; Luo, Z.-Q. Post-translational regulation of ubiquitin signaling. J. Cell Biol. 2019, 218, 1776–1786. [Google Scholar] [CrossRef]
- Noyes, N.C.; Hampton, B.; Migliorini, M.; Strickland, D.K. Regulation of Itch and Nedd4 E3 Ligase Activity and Degradation by LRAD3. Biochemistry 2016, 55, 1204–1213. [Google Scholar] [CrossRef] [PubMed]
- Husnjak, K.; Dikic, I. EGFR trafficking: Parkin’ in a jam. Nat. Cell Biol. 2006, 8, 787–788. [Google Scholar] [CrossRef] [PubMed]
- Attali, I.; Tobelaim, W.S.; Persaud, A.; Motamedchaboki, K.; Simpson-Lavy, K.J.; Mashahreh, B.; Levin-Kravets, O.; Keren-Kaplan, T.; Pilzer, I.; Kupiec, M.; et al. Ubiquitylation-dependent oligomerization regulates activity of Nedd4 ligases. EMBO J. 2017, 36, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Shao, G.; Wang, R.; Sun, A.; Wei, J.; Peng, K.; Dai, Q.; Yang, W.; Lin, Q. The E3 ubiquitin ligase NEDD4 mediates cell migration signaling of EGFR in lung cancer cells. Mol. Cancer 2018, 17, 24. [Google Scholar] [CrossRef]
- Tu, J.; Zhang, B.; Fang, G.; Chang, W.; Zhao, Y. Neddylation-mediated Nedd4-2 activation regulates ubiquitination modification of renal NBCe1. Exp. Cell Res. 2020, 390, 111958. [Google Scholar] [CrossRef]
- Grimsey, N.J.; Narala, R.; Rada, C.C.; Mehta, S.; Stephens, B.S.; Kufareva, I.; Lapek, J.; Gonzalez, D.J.; Handel, T.M.; Zhang, J.; et al. A Tyrosine Switch on NEDD4-2 E3 Ligase Transmits GPCR Inflammatory Signaling. Cell Rep. 2018, 24, 3312–3323.e3315. [Google Scholar] [CrossRef]
- Debonneville, C.; Flores, S.Y.; Kamynina, E.; Plant, P.J.; Tauxe, C.; Thomas, M.A.; Münster, C.; Chraïbi, A.; Pratt, J.H.; Horisberger, J.-D.; et al. Phosphorylation of Nedd4-2 by Sgk1 regulates epithelial Na+ channel cell surface expression. EMBO J. 2001, 20, 7052–7059. [Google Scholar] [CrossRef]
- Snyder, P.M.; Olson, D.R.; Thomas, B.C. Serum and Glucocorticoid-regulated Kinase Modulates Nedd4-2-mediated Inhibition of the Epithelial Na+Channel. J. Biol. Chem. 2002, 277, 5–8. [Google Scholar] [CrossRef]
- Chandran, S.; Li, H.; Dong, W.; Krasinska, K.; Adams, C.; Alexandrova, L.; Chien, A.; Hallows, K.R.; Bhalla, V. Neural Precursor Cell-expressed Developmentally Down-regulated Protein 4-2 (Nedd4-2) Regulation by 14-3-3 Protein Binding at Canonical Serum and Glucocorticoid Kinase 1 (SGK1) Phosphorylation Sites. J. Biol. Chem. 2011, 286, 37830–37840. [Google Scholar] [CrossRef]
- Nagaki, K.; Yamamura, H.; Shimada, S.; Saito, T.; Hisanaga, S.-I.; Taoka, M.; Isobe, T.; Ichimura, T. 14-3-3 Mediates Phosphorylation-Dependent Inhibition of the Interaction between the Ubiquitin E3 Ligase Nedd4-2 and Epithelial Na+ Channels. Biochemistry 2006, 45, 6733–6740. [Google Scholar] [CrossRef]
- Zhou, R.; Snyder, P.M. Nedd4-2 phosphorylation induces serum and glucocorticoid-regulated kinase (SGK) ubiquitination and degradation. J. Biol. Chem. 2005, 280, 4518–4523. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Chiba, T.; Nishiyama, S.; Kakiuchi, T.; Tsukada, H.; Hatano, T.; Fukuda, T.; Yasoshima, Y.; Kai, N.; Kobayashi, K.; et al. Decline of striatal dopamine release in parkin-deficient mice shown by ex vivo autoradiography. J. Neurosci. Res. 2006, 84, 1350–1357. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Kawabe, H.; Jiang, C.; Zhang, W.; Xiang, Y.-Y.; Lu, C.; Salter, M.W.; Brose, N.; Lu, W.-Y.; Rotin, D. Deletion of the ubiquitin ligase Nedd4L in lung epithelia causes cystic fibrosis-like disease. Proc. Natl. Acad. Sci. USA 2011, 108, 3216–3221. [Google Scholar] [CrossRef] [PubMed]
- Lakso, M.; Pichel, J.G.; Gorman, J.R.; Sauer, B.; Okamoto, Y.; Lee, E.; Alt, F.W.; Westphal, H. Efficient in vivo manipulation of mouse genomic sequences at the zygote stage. Proc. Natl. Acad. Sci. USA 1996, 93, 5860–5865. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.P.; Cao, X.R.; Sweezer, E.M.; Kinney, T.S.; Williams, N.R.; Husted, R.F.; Nair, R.; Weiss, R.M.; Williamson, R.A.; Sigmund, C.D.; et al. Salt-sensitive hypertension and cardiac hypertrophy in mice deficient in the ubiquitin ligase Nedd4-2. Am. J. Physiol. Ren. Physiol. 2008, 295, F462–F470. [Google Scholar] [CrossRef]
- Dinudom, A.; Fotia, A.B.; Lefkowitz, R.J.; Young, J.A.; Kumar, S.; Cook, D.I. The kinase Grk2 regulates Nedd4/Nedd4-2-dependent control of epithelial Na+ channels. Proc. Natl. Acad. Sci. USA 2004, 101, 11886–11890. [Google Scholar] [CrossRef]
- Pronin, A.N.; Morris, A.J.; Surguchov, A.; Benovic, J.L. Synucleins are a novel class of substrates for G protein-coupled receptor kinases. J. Biol. Chem. 2000, 275, 26515–26522. [Google Scholar] [CrossRef]
- Howitt, J.; Gysbers, A.M.; Ayton, S.; Carew-Jones, F.; Putz, U.; Finkelstein, D.I.; Halliday, G.M.; Tan, S.-S. Increased Ndfip1 in the Substantia Nigra of Parkinsonian Brains Is Associated with Elevated Iron Levels. PLoS ONE 2014, 9, e87119. [Google Scholar] [CrossRef]
- Guo, M.; Wang, J.; Zhao, Y.; Feng, Y.; Han, S.; Dong, Q.; Cui, M.; Tieu, K. Microglial exosomes facilitate α-synuclein transmission in Parkinson’s disease. Brain 2020, 143, 1476–1497. [Google Scholar] [CrossRef]
- Oliver, P.M.; Cao, X.; Worthen, G.S.; Shi, P.; Briones, N.; Macleod, M.; White, J.; Kirby, P.; Kappler, J.; Marrack, P.; et al. Ndfip1 Protein Promotes the Function of Itch Ubiquitin Ligase to Prevent T Cell Activation and T Helper 2 Cell-Mediated Inflammation. Immunity 2006, 25, 929–940. [Google Scholar] [CrossRef]
- O’Leary, C.E.; Riling, C.R.; Spruce, L.A.; Ding, H.; Kumar, S.; Deng, G.; Liu, Y.; Seeholzer, S.H.; Oliver, P.M. Ndfip-mediated degradation of Jak1 tunes cytokine signalling to limit expansion of CD4+ effector T cells. Nat. Commun. 2016, 7, 11226. [Google Scholar] [CrossRef] [PubMed]
- Behrends, C.; Sowa, M.E.; Gygi, S.P.; Harper, J.W. Network organization of the human autophagy system. Nature 2010, 466, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sun, R.Q.; Camera, D.; Zeng, X.Y.; Jo, E.; Chan, S.M.H.; Herbert, T.P.; Molero, J.C.; Ye, J.M. Endoplasmic reticulum stress up-regulates Nedd4-2 to induce autophagy. FASEB J. 2016, 30, 2549–2556. [Google Scholar] [CrossRef] [PubMed]
- Conway, J.A.; Kramer, E.R. Is activation of GDNF/RET signaling the answer for successful treatment of Parkinson’s disease? A discussion of data from the culture dish to the clinic. Neural Regen. Res. 2022, 17, 1462. [Google Scholar] [CrossRef]
- Meng, B.; Lever, A.M.L. The Interplay between ESCRT and Viral Factors in the Enveloped Virus Life Cycle. Viruses 2021, 13, 324. [Google Scholar] [CrossRef]
- Malakhova, O.A.; Zhang, D.-E. ISG15 Inhibits Nedd4 Ubiquitin E3 Activity and Enhances the Innate Antiviral Response*. J. Biol. Chem. 2008, 283, 8783–8787. [Google Scholar] [CrossRef]
- Okumura, A.; Pitha, P.M.; Harty, R.N. ISG15 inhibits Ebola VP40 VLP budding in an L-domain-dependent manner by blocking Nedd4 ligase activity. Proc. Natl. Acad. Sci. USA 2008, 105, 3974–3979. [Google Scholar] [CrossRef]
- Riederer, P.; Berg, D.; Casadei, N.; Cheng, F.; Classen, J.; Dresel, C.; Jost, W.; Krüger, R.; Müller, T.; Reichmann, H.; et al. α-Synuclein in Parkinson’s disease: Causal or bystander? J. Neural Transm. 2019, 126, 815–840. [Google Scholar] [CrossRef]
- Kramer, E.R.; Liss, B. GDNF-Ret signaling in midbrain dopaminergic neurons and its implication for Parkinson disease. FEBS Lett. 2015, 589, 3760–3772. [Google Scholar] [CrossRef]
- Plaza-Menacho, I.; Barnouin, K.; Barry, R.; Borg, A.; Orme, M.; Chauhan, R.; Mouilleron, S.; Martínez-Torres, R.J.; Meier, P.; McDonald, N.Q. RET Functions as a Dual-Specificity Kinase that Requires Allosteric Inputs from Juxtamembrane Elements. Cell Rep. 2016, 17, 3319–3332. [Google Scholar] [CrossRef]
- Conway, J.A.; Ince, S.; Black, S.; Kramer, E.R. GDNF/RET signaling in dopamine neurons in vivo. Cell Tissue Res. 2020, 382, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Kramer, E.R.; Aron, L.; Ramakers, G.M.J.; Seitz, S.; Zhuang, X.; Beyer, K.; Smidt, M.P.; Klein, R. Absence of Ret Signaling in Mice Causes Progressive and Late Degeneration of the Nigrostriatal System. PLoS Biol. 2007, 5, e39. [Google Scholar] [CrossRef] [PubMed]
- Kowsky, S.; Pöppelmeyer, C.; Kramer, E.R.; Falkenburger, B.H.; Kruse, A.; Klein, R.; Schulz, J.B. RET signaling does not modulate MPTP toxicity but is required for regeneration of dopaminergic axon terminals. Proc. Natl. Acad. Sci. USA 2007, 104, 20049–20054. [Google Scholar] [CrossRef] [PubMed]
- Drinkut, A.; Tillack, K.; Meka, D.P.; Schulz, J.B.; Kügler, S.; Kramer, E.R. Ret is essential to mediate GDNF’s neuroprotective and neuroregenerative effect in a Parkinson disease mouse model. Cell Death Dis. 2016, 7, e2359. [Google Scholar] [CrossRef] [PubMed]
- Haglund, K.; Dikic, I. The role of ubiquitylation in receptor endocytosis and endosomal sorting. J. Cell Sci. 2012, 125, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Crupi, M.J.; Yoganathan, P.; Bone, L.N.; Lian, E.; Fetz, A.; Antonescu, C.N.; Mulligan, L.M. Distinct Temporal Regulation of RET Isoform Internalization: Roles of Clathrin and AP2. Traffic 2015, 16, 1155–1173. [Google Scholar] [CrossRef] [PubMed]
- Richardson, D.S.; Rodrigues, D.M.; Hyndman, B.D.; Crupi, M.J.; Nicolescu, A.C.; Mulligan, L.M. Alternative splicing results in RET isoforms with distinct trafficking properties. Mol. Biol. Cell 2012, 23, 3838–3850. [Google Scholar] [CrossRef]
- Grice, G.L.; Nathan, J.A. The recognition of ubiquitinated proteins by the proteasome. Cell Mol. Life Sci. 2016, 73, 3497–3506. [Google Scholar] [CrossRef]
- Vecchione, A.; Marchese, A.; Henry, P.; Rotin, D.; Morrione, A. The Grb10/Nedd4 complex regulates ligand-induced ubiquitination and stability of the insulin-like growth factor I receptor. Mol. Cell Biol. 2003, 23, 3363–3372. [Google Scholar] [CrossRef]
- Kwak, Y.D.; Wang, B.; Li, J.J.; Wang, R.; Deng, Q.; Diao, S.; Chen, Y.; Xu, R.; Masliah, E.; Xu, H.; et al. Upregulation of the E3 ligase NEDD4-1 by Oxidative Stress Degrades IGF-1 Receptor Protein in Neurodegeneration. J. Neurosci. 2012, 32, 10971–10981. [Google Scholar] [CrossRef]
- Cao, X.R.; Lill, N.L.; Boase, N.; Shi, P.P.; Croucher, D.R.; Shan, H.; Qu, J.; Sweezer, E.M.; Place, T.; Kirby, P.A.; et al. Nedd4 controls animal growth by regulating IGF-1 signaling. Sci. Signal. 2008, 1, ra5. [Google Scholar] [CrossRef] [PubMed]
- Giorgetti-Peraldi, S.; Murdaca, J.; Mas, J.C.; Van Obberghen, E. The adapter protein, Grb10, is a positive regulator of vascular endothelial growth factor signaling. Oncogene 2001, 20, 3959–3968. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hurley, J.H. Nipped in the bud: How the AMSH MIT domain helps deubiquitinate lysosome-bound cargo. Structure 2011, 19, 1033–1035. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Shields, S.B.; Piper, R.C. How ubiquitin functions with ESCRTs. Traffic 2011, 12, 1306–1317. [Google Scholar] [CrossRef]
- Roxrud, I.; Raiborg, C.; Pedersen, N.M.; Stang, E.; Stenmark, H. An endosomally localized isoform of Eps15 interacts with Hrs to mediate degradation of epidermal growth factor receptor. J. Cell Biol. 2008, 180, 1205–1218. [Google Scholar] [CrossRef]
- Hasegawa, T.; Konno, M.; Baba, T.; Sugeno, N.; Kikuchi, A.; Kobayashi, M.; Miura, E.; Tanaka, N.; Tamai, K.; Furukawa, K.; et al. The AAA-ATPase VPS4 regulates extracellular secretion and lysosomal targeting of α-synuclein. PLoS ONE 2011, 6, e29460. [Google Scholar] [CrossRef]
- Tanikawa, S.; Mori, F.; Tanji, K.; Kakita, A.; Takahashi, H.; Wakabayashi, K. Endosomal sorting related protein CHMP2B is localized in Lewy bodies and glial cytoplasmic inclusions in α-synucleinopathy. Neurosci. Lett. 2012, 527, 16–21. [Google Scholar] [CrossRef]
- Kurashige, T.; Takahashi, T.; Yamazaki, Y.; Hiji, M.; Izumi, Y.; Yamawaki, T.; Matsumoto, M. Localization of CHMP2B-immunoreactivity in the brainstem of Lewy body disease. Neuropathology 2013, 33, 237–245. [Google Scholar] [CrossRef]
- Jing, W.; Wang, G.; Cui, Z.; Xiong, G.; Jiang, X.; Li, Y.; Li, W.; Han, B.; Chen, S.; Shi, B. FGFR3 Destabilizes PD-L1 via NEDD4 to Control T-cell-Mediated Bladder Cancer Immune Surveillance. Cancer Res. 2022, 82, 114–129. [Google Scholar] [CrossRef]
- Yu, T.; Calvo, L.; Anta, B.; López-Benito, S.; López-Bellido, R.; Vicente-García, C.; Tessarollo, L.; Rodriguez, R.E.; Arévalo, J.C. In vivo regulation of NGF-mediated functions by Nedd4-2 ubiquitination of TrkA. J. Neurosci. 2014, 34, 6098–6106. [Google Scholar] [CrossRef]
- Yanpallewar, S.; Wang, T.; Koh, D.C.; Quarta, E.; Fulgenzi, G.; Tessarollo, L. Nedd4-2 haploinsufficiency causes hyperactivity and increased sensitivity to inflammatory stimuli. Sci. Rep. 2016, 6, 32957. [Google Scholar] [CrossRef] [PubMed]
- Melchior, B.; Nerrière-Daguin, V.; Laplaud, D.-A.; Remy, S.; Wiertlewski, S.; Neveu, I.; Naveilhan, P.; Meakin, S.; Brachet, P. Ectopic expression of the TrkA receptor in adult dopaminergic mesencephalic neurons promotes retrograde axonal NGF transport and NGF-dependent neuroprotection. Exp. Neurol. 2003, 183, 367–378. [Google Scholar] [CrossRef]
- Ding, Y.-X.; Xia, Y.; Jiao, X.-Y.; Duan, L.; Yu, J.; Wang, X.; Chen, L.-W. The TrkB-Positive Dopaminergic Neurons are Less Sensitive to MPTP Insult in the Substantia Nigra of Adult C57/BL Mice. Neurochem. Res. 2011, 36, 1759–1766. [Google Scholar] [CrossRef] [PubMed]
- Heldin, C.H.; Moustakas, A. Signaling Receptors for TGF-β Family Members. Cold Spring Harb. Perspect. Biol. 2016, 8, 22053. [Google Scholar] [CrossRef]
- Vander Ark, A.; Cao, J.; Li, X. TGF-β receptors: In and beyond TGF-β signaling. Cell. Signal. 2018, 52, 112–120. [Google Scholar] [CrossRef]
- Luo, S.X.; Huang, E.J. Dopaminergic Neurons and Brain Reward Pathways: From Neurogenesis to Circuit Assembly. Am. J. Pathol. 2016, 186, 478–488. [Google Scholar] [CrossRef]
- Sorkina, T.; Miranda, M.; Dionne, K.R.; Hoover, B.R.; Zahniser, N.R.; Sorkin, A. RNA Interference Screen Reveals an Essential Role of Nedd4-2 in Dopamine Transporter Ubiquitination and Endocytosis. J. Neurosci. 2006, 26, 8195–8205. [Google Scholar] [CrossRef]
- Efimova, E.V.; Gainetdinov, R.R.; Budygin, E.A.; Sotnikova, T.D. Dopamine transporter mutant animals: A translational perspective. J. Neurogenet. 2016, 30, 5–15. [Google Scholar] [CrossRef]
- García-Tardón, N.; González-González, I.M.; Martínez-Villarreal, J.; Fernández-Sánchez, E.; Giménez, C.; Zafra, F. Protein Kinase C (PKC)-promoted Endocytosis of Glutamate Transporter GLT-1 Requires Ubiquitin Ligase Nedd4-2-dependent Ubiquitination but Not Phosphorylation. J. Biol. Chem. 2012, 287, 19177–19187. [Google Scholar] [CrossRef]
- Yu, Z.; Liu, C.; Zhang, J.; Liang, Z.; You, G. Protein kinase C regulates organic anion transporter 1 through phosphorylating ubiquitin ligase Nedd4-2. BMC Mol. Cell Biol. 2021, 22, 53. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, C.; You, G. AG490, a JAK2-specific inhibitor, downregulates the expression and activity of organic anion transporter-3. J. Pharm. Sci. 2018, 136, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Malagelada, C.; Ryu, E.J.; Biswas, S.C.; Jackson-Lewis, V.; Greene, L.A. RTP801 Is Elevated in Parkinson Brain Substantia Nigral Neurons and Mediates Death in Cellular Models of Parkinson’s Disease by a Mechanism Involving Mammalian Target of Rapamycin Inactivation. J. Neurosci. 2006, 26, 9996–10005. [Google Scholar] [CrossRef] [PubMed]
- Canal, M.; Romaní-Aumedes, J.; Martín-Flores, N.; Pérez-Fernández, V.; Malagelada, C. RTP801/REDD1: A stress coping regulator that turns into a troublemaker in neurodegenerative disorders. Front. Cell Neurosci. 2014, 8, 313. [Google Scholar] [CrossRef] [PubMed]
- Grace, A.; Bunney, B. The control of firing pattern in nigral dopamine neurons: Single spike firing. J. Neurosci. 1984, 4, 2866–2876. [Google Scholar] [CrossRef] [PubMed]
- Grace, A.; Bunney, B. The control of firing pattern in nigral dopamine neurons: Burst firing. J. Neurosci. 1984, 4, 2877–2890. [Google Scholar] [CrossRef]
- Lammel, S.; Hetzel, A.; Häckel, O.; Jones, I.; Liss, B.; Roeper, J. Unique Properties of Mesoprefrontal Neurons within a Dual Mesocorticolimbic Dopamine System. Neuron 2008, 57, 760–773. [Google Scholar] [CrossRef]
- Margolis, E.B.; Coker, A.R.; Driscoll, J.R.; Lemaître, A.-I.; Fields, H.L. Reliability in the Identification of Midbrain Dopamine Neurons. PLoS ONE 2010, 5, e15222. [Google Scholar] [CrossRef]
- Drion, G.; Massotte, L.; Sepulchre, R.; Seutin, V. How Modeling Can Reconcile Apparently Discrepant Experimental Results: The Case of Pacemaking in Dopaminergic Neurons. PLoS Comput. Biol. 2011, 7, e1002050. [Google Scholar] [CrossRef]
- Branch, S.Y.; Sharma, R.; Beckstead, M.J. Aging decreases L-type calcium channel currents and pacemaker firing fidelity in substantia nigra dopamine neurons. J. Neurosci. 2014, 34, 9310–9318. [Google Scholar] [CrossRef]
- Rice, M.E.; Patel, J.C.; Cragg, S.J. Dopamine release in the basal ganglia. Neuroscience 2011, 198, 112–137. [Google Scholar] [CrossRef]
- Van Wart, A.; Matthews, G. Impaired Firing and Cell-Specific Compensation in Neurons Lacking Nav1.6 Sodium Channels. J. Neurosci. 2006, 26, 7172–7180. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lin, Y.; Liu, W.; Kuang, P.; Lao, W.; Ji, Y.; Zhu, H. Voltage-Gated Sodium Channels Are Involved in Cognitive Impairments in Parkinson’s Disease- like Rats. Neuroscience 2019, 418, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Hansen, H.H.; Ebbesen, C.; Mathiesen, C.; Weikop, P.; Rønn, L.C.; Waroux, O.; Scuvée-Moreau, J.; Seutin, V.; Mikkelsen, J.D. The KCNQ channel opener retigabine inhibits the activity of mesencephalic dopaminergic systems of the rat. J. Pharm. Exp. 2006, 318, 1006–1019. [Google Scholar] [CrossRef] [PubMed]
- Cooper, E.C.; Harrington, E.; Jan, Y.N.; Jan, L.Y. M channel KCNQ2 subunits are localized to key sites for control of neuronal network oscillations and synchronization in mouse brain. J. Neurosci. 2001, 21, 9529–9540. [Google Scholar] [CrossRef] [PubMed]
- Weber, Y.G.; Geiger, J.; Kämpchen, K.; Landwehrmeyer, B.; Sommer, C.; Lerche, H. Immunohistochemical analysis of KCNQ2 potassium channels in adult and developing mouse brain. Brain Res. 2006, 1077, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Martire, M.; D’Amico, M.; Panza, E.; Miceli, F.; Viggiano, D.; Lavergata, F.; Iannotti, F.A.; Barrese, V.; Preziosi, P.; Annunziato, L.; et al. Involvement of KCNQ2 subunits in [3H]dopamine release triggered by depolarization and pre-synaptic muscarinic receptor activation from rat striatal synaptosomes. J. Neurochem. 2007, 102, 179–193. [Google Scholar] [CrossRef]



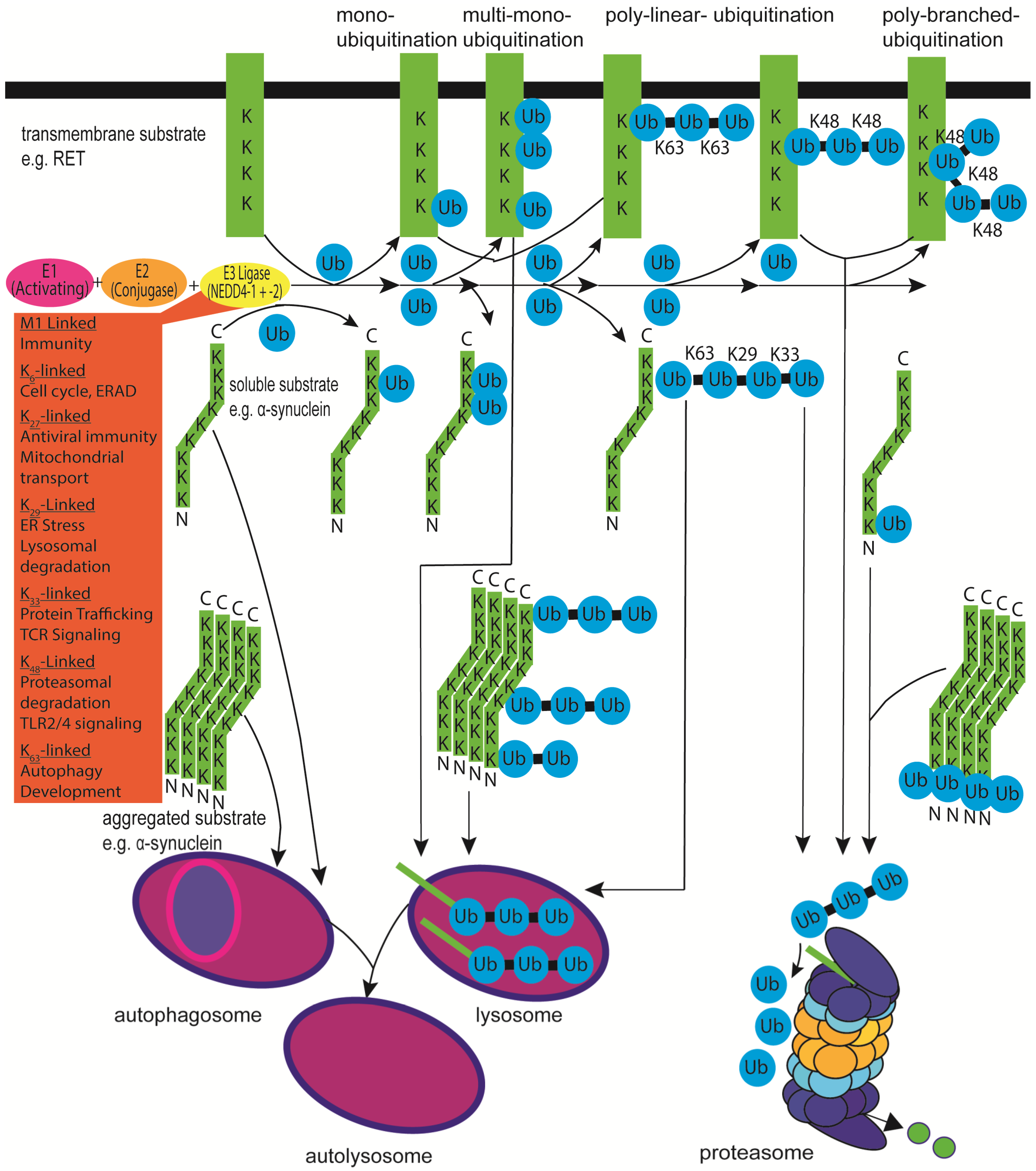{kind=link}
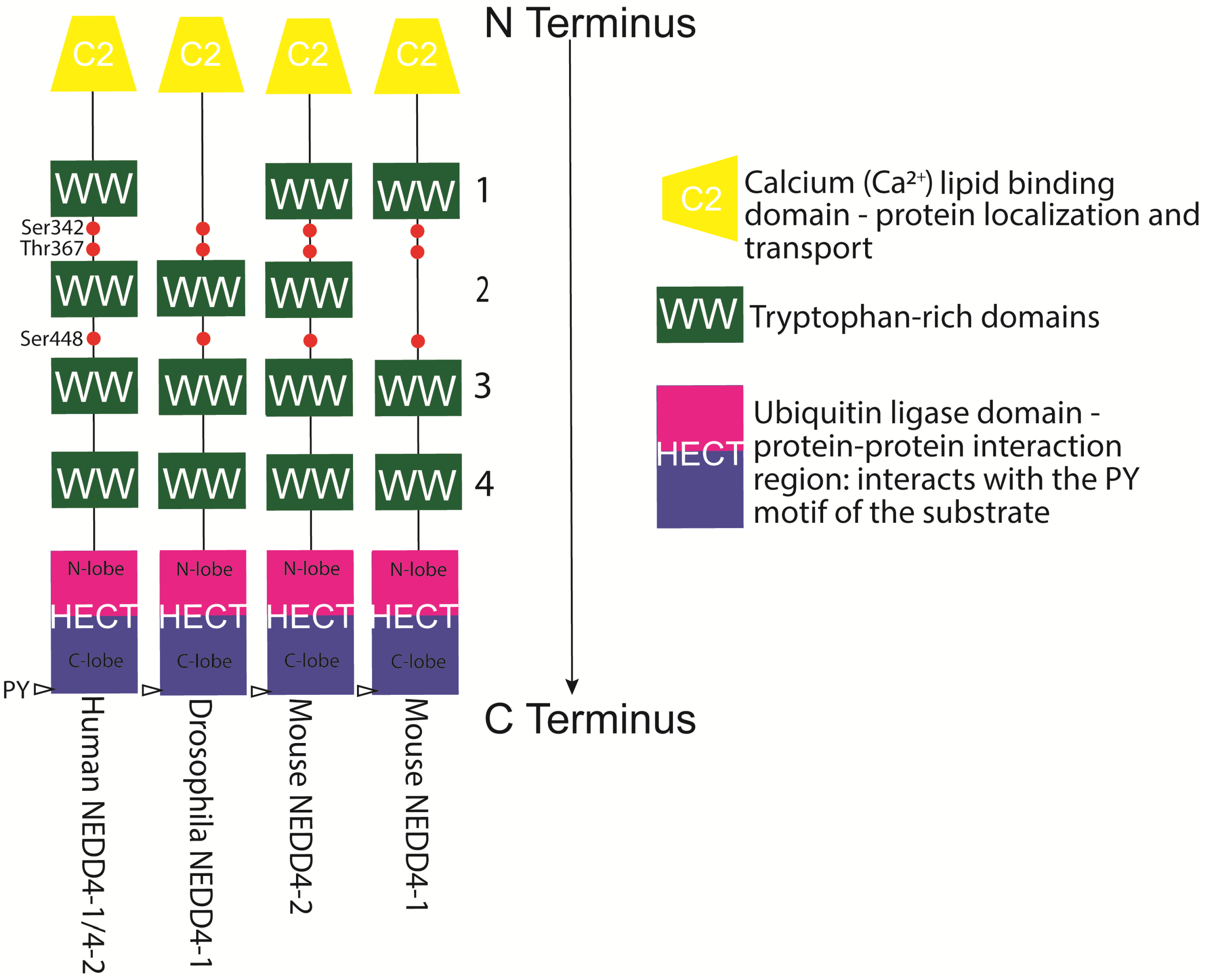{kind=link}
| Name | Substrate | Adaptor | Modifier/Regulator | Binding Motifs | Modification | Function | Experimental Model Used | References |
|---|---|---|---|---|---|---|---|---|
| ABCB1 (ATP-binding cassette sub-family B member 1/P-Glycoprotein) | 🗸 | PDY | Polyubiquitination | An ATP Binding Cassette transporter that exports β-amyloid from Blood-brain barrier endothelial cells. Potential for intervention in Alzheimer’s disease | In vitro: Sf21 cells | [31] | ||
| α-synuclein | 🗸 | PDNEAYEMP, PLY, PPLP, PPSP, PFY | Monoubiquitination and Polyubiquitination (K63–linked) | Lysosomal degradation. Potential protection mechanism against Parkinson’s Disease pathogenesis | In vitro: SH–SY5Y cells In vivo: Drosophila and Rat | [28,29,30] | ||
| Beclin 1 | 🗸 | LPLY | Beclin 1: polyubiquitination (K63 and K11–linked) | Subunit of the class III phosphatidylinositol 3-kinase complex. Autophagy-related protein. Proteasomal degradation. Inhibited autophagy and cell survival | In vitro: HeLa cells | [32] | ||
| β2-AR (β2-Arrestin Receptor) | 🗸 | Ubiquitination | Involved in internalised receptor degradation and lysosomal trafficking. Degraded via the lysosome | In vitro: HEK293 cells | [33] | |||
| β-arrestin 1 | 🗸 | Adaptor protein for NHE1 ubiquitination | In vitro: HEK293 cells, mouse embryonic fibroblasts | [34] | ||||
| β-arrestin 2 | 🗸 | 🗸 | Adaptor protein for β2-adrenergic receptor ubiquitination | In vitro: HEK293 cells | [33] | |||
| Caspase-1, -3, -4, -6, -7, -11 | 🗸 | 🗸 | Truncation | NEDD4-1 can be cleaved by caspases and can K48-polyubiquitinate caspase 11 | In vitro: Jurkat cells A549 | [14,35,36,37] | ||
| Cbl-b (Casitas B-lineage Lymphoma b) | 🗸 | 🗸 | Polyubiquitination | Impedes NEDD4-1 interaction with PTEN and also polyubiquitinates Cbl-b for degradation | In vivo: Cbl-bC373A and NEDD4Gt(IRESβgeo)249Lex mice | [38,39] | ||
| CNrasGEF (Cyclic Nucleotide rat sarcoma virus Guanine nucleotide Exchange Factor) | 🗸 | PPGY, PPDY | Polyubiquitination | RAS guanine nucleotide exchange factors that are degraded via the proteasome. NEDD4-1 overexpression promoted migration and invasion of glioma cells | In vitro: HEK293T and Glioma cells | [40,41] | ||
| Connexin43 | 🗸 | PPGY | Ubiquitination | Gap junction protein. Proteasome and lysosome degradation | In vitro: WB-F344 rat liver epithelial cells | [42] | ||
| c-Src (Proto-oncogene tyrosine-protein kinase Src) | 🗸 | Tyrosine kinase that activates NEDD4-1 through phosphorylation of its HECT and C2 domains. Phosphorylation inhibits auto-regulation | In vitro: HeLa, HEK293T and Platinum E cells | [43] | ||||
| Δ Np63 transcriptional target | 🗸 | 🗸 | PPPY | Ubiquitination and polyubiquitination | A homologue of p53 tumour suppressor. Protein destabilisation of ∆Np63α and degradation. Downregulates NEDD4-1 leading to the suppression of nuclear PTEN in basal layer keratinocytes | In vitro: HEK293-EBNA, HaCaT, A431 andH1299 cells In vivo: Zebrafish embryos | [44,45] | |
| EPS15 (Epithelial growth factor receptor substrate 15) | 🗸 | Monoubiquitination and polyubiquitination | An endocytic protein that is targeted for degradation by NEDD4-1 | In vitro: HeLa and B82L cells | [46] | |||
| FGFR1 (Fibroblast Growth Factor Receptor 1) | 🗸 | 🗸 | VLLVRPSRLSSSG | Ubiquitination | FGFR1 is a tyrosine kinase involved in cell proliferation and differentiation during development. Inhibited neural stem cell differentiation. Activates c-Src that subsequently activates NEDD4-1 | In vitro: Human embryonic stem cells In vivo: Zebrafish embryos | [47,48] | |
| GAG (group specific antigen) protein | 🗸 | PPPY, PTAP | Monoubiquitination | From the HTLV-1 (human T-lymphotropic virus type 1) involved in hijacking mutivesicular body (MVB) pathway proteins required for viral budding | In vitro: HEK293T cells | [49] | ||
| γ2-adaptin | 🗸 | 🗸 | PPAY | Monoubiquitination and multi-polyubiquitination | A member of the clathrin adaptor protein family. Forms a complex with NEDD4-1 and is involved in endosomal/multivesicular body (MVB) pathway and the assembly and release of the HBV | In vitro: HuH-7 and HeLa cells | [50,51] | |
| HBV X protein (Hepatitis B virus) | 🗸 | Ubiquitination | A multifunctional regulator that is encoded by the HBV genome. It is degraded via the K48 proteasomal pathway | In vitro: HEK293T, HBV-related HCC cell lines - HepG2.215, HepG3B, SNU182, SNU387, PLC/PRF/5, and MHCC97H | [52] | |||
| HER3 (human epidermal growth factor receptor 3) | 🗸 | PPRY | Polyubiquitination | A member of the EGRF family. Degraded via the proteasome. Inhibited cancer cell proliferation and tumour growth. NEDD4-1 knockdown induces apoptosis in DU145 cells | In vivo: Chinese Hamster ovary cells In vitro: MDA-MB-453, MCF-7, and DU145 cells | [53,54] | ||
| HGS (Hepatocyte growth factor-regulated tyrosine kinase substrate) | 🗸 | PPEY | Ubiquitination | Binding partner for NEDD4-1 involved in EGFR lysosome degradation | In vitro: Chinese hamster ovary and HeLa cells | [55] | ||
| IFITM3 (Interferon (IFN)-induced transmembrane protein 3) | 🗸 | PPNY | Polyubiquitination | A cell-intrinsic factor that limits influenza virus and other viral infections such as SARS | In vitro: HEK293T, A549, NCI-H358, NCI-H2009 cells and MEFs | [56] | ||
| IGF1R (Insulin-like growth factor 1 receptor) | 🗸 | Ubiquitination | A tyrosine kinase receptor. Expression can be downregulated by NEDD4-1 through the indirect effect on the oxidisation of very-low-density lipoproteins. Ubiquitination and degradation require a C1060 site | In vitro: Hepatocytes from Landes goose embryos In vivo: Intracerebral haemorrhage mice, Sprague Dawley and Tg2576 mice | [57] | |||
| IGPR-1 (Immunoglobulin and Proline-rich receptor-1, also known as TMIGD2/CD28H) | 🗸 | PPR | Polyubiquitination | A cell adhesion molecule involved in, for example, autophagy, angiogenesis and cell adhesion. Is degraded via the lysosomal pathway | In vitro: HEK293 cells | [58] | ||
| IRS-2 (insulin receptor substrate) | 🗸 | Monoubiquitination | NEDD4-1 recruits IRS-2 to the membrane to enhance IGF signalling | In vivo: Zebrafish embryos In vitro: HEK293 cells | [59] | |||
| ISG15 (Interferon-stimulated gene 15) | 🗸 | ISGylation | Can attach to NEDD4-1 inhibiting its ubiquitination properties | In vitro: HEK293, HeLa cells and MEFs | [60,61,62] | |||
| KLF8 (Krueppel-like factor 8) | 🗸 | Ubiquitination | The function of this transcription factor is regulated by NEDD4-1 | In vitro: HEK293 cells | [63,64] | |||
| LATS1 (large tumour suppressor kinase 1) | 🗸 | Ubiquitination | A serine/threonine kinase involved in the suppression of tumours | In vitro: HEK293 cells | [65] | |||
| LC3 (Microtubule-associated protein 1A/1B-light chain 3) | 🗸 | WEII, WVVL, WFFL, WDKL | An autophagy-related protein. LC3 binds to NEDD4-1, but is not a ubiquitination substrate of NEDD4-1 | In vitro: HEK293 cells | [66] | |||
| LDLRAD4 (Low density lipoprotein receptor class A domain containing 4) | 🗸 | Ubiquitination | Degraded via the lysosome and is a negative regulator of TGF-β signalling | In vivo: Female BALB/c nude mice In vitro: L02 and HepG2 cells | [67] | |||
| LMP2A (Latent membrane protein 2A) | 🗸 | PPPPY | Ubiquitination | A latent Epstein–Barr virus (EBV) infection protein involved in B cell signal transduction | In vitro: BJAB, Ramos, Raji, Jurkat, HPB.ALL and M12 cells | [68] | ||
| MDM2 (Mouse double minute 2 homolog) | 🗸 | Polyubiquitination (K63-linked) | Is an E3 ubiquitin ligase involved in negative regulation of p53. MDM2 is stabilised via NEDD4-1 interacting with its RING domain. NEDD4-1 overexpression reduces p53 levels | In vivo: NEDD4-1 KO mouse embryonic fibroblasts | [69] | |||
| MEKK5 (mitogen-activated protein kinase 5) (Apoptosis Signal-regulating Kinase 1 (ASK1) | 🗸 | A serine/threonine kinase that regulates NEDD4-1 cell migration signalling in lung cancer | In vitro: HEK293T, NCI-H1650, and A549 cells | [58] | ||||
| N4BP (NEDD4-binding protein) | 🗸 | PPLP, PPEY, PPPY | Monouniquitination and Polyubiquitination | N4BP is degraded via the proteasome. NEDD4-1 regulates N4BP1 at promyelocytic leukaemia nuclear bodies | In vitro: HEK293 cells and MEFs | [70,71] | ||
| NAB (N-aryl benzdimidazole) | 🗸? | NAB2 reduces the ratio of K63-linked ubiquitin chains on A53T α-synuclein by an unknown mechanism. NAB2 binds NEDD4-1 but does not alter conformation or enzymatic activity. | In vitro: SH-SH5Y cells | [72,73] | ||||
| NEDD4-1 (Neural precursor cell-expressed developmentally-down-regulated protein 4-1) | 🗸 | 🗸 | Autoubiquitination | K29-linked autoubiquitination. C2 and HECT domains bind resulting in autoubiquitination | In vitro: HEK293T, HeLa, THP-1 and A549 cells | [74,75] | ||
| NHE1 (Sodium-Hydrogen antiporter 1) | 🗸 | Multi-monoubiquitination and polyubiquitination | The Na(+)/H(+) exchanger 1 is ubiquitinated for degradation by NEDD4-1 but requires β-arrestin 1 | In vitro: HEK293 cells | [34] | |||
| N-Myc (N-myc proto-oncogene protein/ basic helix-loop-helix protein 37) | 🗸 | Polyubiquitination | An oncoprotein that is degraded via the proteasome. NEDD4-1 suppresses neuroblastoma and pancreatic cancer cell proliferation | In vitro: Neuroblastoma BE(2)-C, CHP134, pancreatic cancer MiaPaca-2 and HEK293 cells | [76] | |||
| Notch | 🗸 | PPSY | Polyubiquitination | A plasma membrane receptor that is ubiquitinated for degradation via the proteasome | In vivo: Drosophila and Conditional NEDD4-1 overexpression in Wistar Rat | [77,78,79] | ||
| Rap2a (RAS-related protein 2a) | 🗸 | Monoubiquitination of K63 | Rap2a Is a member of the RAS-related protein family. NEDD4-1 Inhibits GTP-Rap2a activity subsequently promoting the migration and invasiveness of glioma cells | In vitro: Human glioma cell lines U251 and U87 | [80] | |||
| RAS (Rat sarcoma virus) | 🗸 | PPGY, PPDY | Polyubiquitination and monoubiquitination | Small guanosine triphosphatases involved in a multitude of different cellular processes by acting as a molecular switch. RAS is regulated via NEDD4-1ubiquitination sending it for degradation to the lysosome. This regulation suppressed tumorigenesis | In vitro: HEK293T, HeLa, NIH 3T3, MEF and HepG2 cells | [40,81] | ||
| RET (Rearranged during transfection) | 🗸 | Polyubiquitination | A receptor tyrosine kinase. The short form (Ret9) becomes localised and internalised into the endosomal network through clathrin-coated pits following NEDD4-1 ubiquitination. This causes inhibition of Ret9-mediated neurotrophic signalling at the cell surface and promotion of post-internalisation signalling. This mechanism could potentially impact neurotrophic signalling of dopaminergic neurons and play a role in Parkinson’s disease | In vitro: HEK293 and SH-SY5Y cells | [82] | |||
| RNAPII (Ribonucleic acid Polymerase II) | 🗸 | Monouniquitination and polyubiquitination | A multiprotein involved in the transcription of DNA into mRNA that is degraded via the proteasome after being ubiquitinated by NEDD4-1. This ubiquitination is dependent on NEDD4-1 interacting with the ElonginA/B/C-Cullin 5 complex | In vitro: HEK293, MRC5 and S. cerevisiae | [83,84] | |||
| RTP801/REDD1 | 🗸 | Polyubiquitination | A pro-apoptotic protein that is targeted for degradation by NEDD4-1via K63 ubiquitin linkages. NEDD4-1 loss may elevate RTP801 proteins leading to an increase in neuronal death in Parkinson’s disease | In vivo: NEDD4-1f/f, Emx1Cre mice In vitro: PC12, HEK293 cells and rat primary cortical neurons | [85] | |||
| SAG (S-Arrestin) | 🗸 | Polyubiquitination | An anti-apoptotic cellular survival protein that is degraded by the proteasome. NEDD4-1 reduction of SAG resulted in etoposide-induced apoptosis in cancer cells. SAG does not bind to WW domains as it lacks PY motifs but interacts with NEDD4-1 via its RING domain | In vitro: HEK293T | [86] | |||
| SCAMP3 (Secretory Carrier Membrane Protein 3) | 🗸 | PPAY, PSAP, PTEP | Multi-monoubiquitination | Integral membrane proteins involved in the cell surface recycling system. SCAMP3 is a NEDD4-1 substrate that is involved in the degradation of EGFR via the lysosome | In vitro: HeLa and HEK293T cells | [87] | ||
| Spy1A | 🗸 | Polyubiquitination | A cyclin-like protein that is needed for a cell to progress through the G1/S phase. Spy1A is required for p53-mediated tumour suppression. Spy1A is degraded in a cell cycle-dependent manner during mitosis via the ubiquitin-proteasome system | In vitro: Human mammary breast cancer, MCF7, and HEK293cells | [88,89] | |||
| SQSTM1 (p62) | 🗸 | Polyubiquitination (K63-type) | An autophagy-related protein. NEDD4-1 ubiquitinates its PB1 domain. Lack of NEDD4-1 leads to accumulation of aberrant enlarged inclusion bodies | In vitro: HEK293T, HEK293A and A549 cells | [66,90] | |||
| VEGF-R2 (vascular endothelial growth factor receptor-2) | 🗸 | Monoubiquitination | This receptor is degraded by NEDD4-1 but Grb10 regulates this process by interacting with NEDD4-1 | In vitro: HEK-293 EBNA cells | [91] |
| Name | Substrate | Adaptor | Modifier/Regulator | Binding Motifs | Modification | Function | Experimental Model Used | References |
|---|---|---|---|---|---|---|---|---|
| 14-3-3 | 🗸 | 🗸 | 14-3-3 is an inhibitory binding partner for NEDD4-2 through a PI3-kinase/SGK1-dependent manner. This interaction is dependent on the phosphorylation of key residues Ser342 and Ser448 on NEDD4-2 | In vitro: Kidney tubule epithelial mpkCCDc14, HECT293 and E. coli BL21(DE3) cells | [92,93] | |||
| α-Arrestins | 🗸 | 🗸 | PPLP, PPEY, PPLY, PPSY, PPNY, PPPY | Ubiquitination; adaptor/regulator | α-Arrestins are scaffolding molecules involved in regulating receptor trafficking and cell signalling. These can be both substrates and adaptors for NEDD4-2. α-arrestins are implicated in the regulation of DMT1. It is activated through polymerisation or membrane tethering and is ubiquitinated by NEDD4-2 | In vitro: CHO, Caco-2, HepG2 and HEK293T cells In vivo: Arrdc1tm1(KOMP)Vlcg (VG17312) and Arrdc4tm1(KOMP)Vlcg (VG18749) embryonic stem cells | [27,94,95] | |
| AMPK (AMP-activated protein kinase) | 🗸 | AMPK is a metabolic sensor that inhibits ENaC. It activates NEDD4-2 by phosphorylation, promotes ENaC-NEDD4-2 interaction and subsequent ENaC degradation. AMPK also signals in the mTOR pathway where it plays a role in cell death. | In vitro: HEK293 cells In vivo: Xenopus oocytes | [96] | ||||
| ATA2 (Analogue Terminal Adaptor II) | 🗸 | Polyubiquitination | NEDD4-2 regulates amino acid transporter ATA2 activity on the cell surface by proteasomal degradation | In vitro: 3T3-L1 adipocytes Ex vivo: Xenopus oocytes | [97] | |||
| CFTR (Cystic fibrosis transmembrane conductance regulator) | 🗸 | Ubiquitination | CFTR is downregulated by NEDD4-2 possibly via the proteasome and lysosome degradation (contested by [98]). Ref. [99], however, show that CFTR is degraded via NEDD4-2 but has to interact with 14-3-3ε and be activated by SGK1 phosphorylation | In vitro: CFPAC-1 (ΔF508) and CFBE41o-ΔF cells | [98,99,100] | |||
| CHT1 high-affinity choline transporter 1) | 🗸 | Ubiquitination | NEDD4-2-mediated ubiquitination regulates cell surface expression of CHT1 thereby impeding choline uptake and HC-3 binding | In vitro: HEK293 Cells | [101] | |||
| CLC-5 (Chloride Voltage-Gated Channel 5) | 🗸 | PPLPPY | Ubiquitination | Voltage-gated channels that function as dimers. NEDD4-2 decreases cell surface expression of ClC-5 through ubiquitination | Ex vivo: Xenopus oocytes In vivo: NEDD4–2 null mice | [102,103,104] | ||
| CLC-K (Chloride channel protein ClC-Ka) /barttin | 🗸 | PPYVRL (located on barttin) | Ubiquitination | A chloride channel that requires barttin to be functional. Downregulation of ClC-Ka/barttin comes as a result of NEDD4-2-mediated ubiquitination | Ex vivo: Xenopus oocytes | [105,106] | ||
| CRTC3 (CREB Regulated Transcription Coactivator 3) | 🗸 | PPPY | Polyubiquitination | NEDD4-2 is responsible for the downregulation of CRTC3 in a proteasome–dependant manner in response to prolonged cAMP signalling | In vitro: HEK293T cells | [107] | ||
| DAT (Dopamine Transporter) | 🗸 | Polyubiquitination | A transporter for dopamine. PKC-dependent DAT ubiquitination by NEDD4-2 requires its WW3 and WW4 domains. May have implications in Parkinson’s disease | In vitro: HEK 293 and PAE cells | [108] | |||
| DMT1 (Divalent metal transporter 1) | 🗸 | Polyubiquitination | A metal transporter that is ubiquitinated by NEDD4-2 but requires the adaptor protein Ndfip1 | In vitro: SH-SY5Y and HEK293T cells | [109] | |||
| DVL2 (Dishevelled 2) | 🗸 | PPPY | Polyubiquitination | NEDD4-2 negatively regulates Wnt signalling by targeting dishevelled for proteasomal degradation. Wnt5a induces JNK-mediated phosphorylation of NEDD4-2, which in turn promotes Dvl2 degradation | In vitro: HEK293T and HeLa cells | [110,111] | ||
| EAAT1/2 (Excitatory amino acid transporter 1 and 2. Also known as Glutamate transporter 1 and 2) | 🗸 | PPPD | Ubiquitination | Regulation of EAAT1/2 through NEDD4-2 depends on SGK kinases. NEDD4-2 knockdown with shRNA decreases GLT-1 ubiquitination, promoting glutamate uptake and increases GLT-1 expression. This may play a role in glutamatergic signalling in dementia | Ex vivo: Xenopus oocytes In vitro: MPP+ treated astrocytes | [112,113,114] | ||
| GluA1 (Glutamate receptor 1) | 🗸 | PKY | Ubiquitination | NEDD4-2 ubiquitinates GluA1 at lysine-868 and mediates its surface expression. This may play a role in glutamatergic signalling in dementia | In vitro: HEK293 cells In vivo: NEDD4-2andi mice | [115] | ||
| hERG(1) (human ether-à-gogo-related gene (1)) | 🗸 | PPAY | Monoubiquitination and polyubiquitination | The human ether-a-go-go-related gene protein (hERG) is a voltage-gated cardiac potassium channel. Caveolin-3 (Cav3), hERG, and NEDD4-2 form a complex. hERG expression in the plasma membrane is regulated by Cav3 through NEDD4-2 ubiquitination | In vitro: HEK293 cells (Patch clamp) In vivo: Guinea pig | [38,116] | ||
| IKKβ (inhibitor of nuclear factor kappa-B kinase subunit β) | 🗸 | 🗸 | IKKβ activates NEDD4-2 via phosphorylation that results in the regulation of ENaC | In vitro: HEK-293, HEK-293T and mpkCCDc14 cells. Ex vivo: Xenopus oocytes (TEV) | [117] | |||
| JNK1 (Janus Kinase 1) | 🗸 | 🗸 | JNK1 activates NEDD4-2 via phosphorylation that results in the regulation of ENaC | In vitro: HEK293 cells and mpkCCDc14 cells. Ex vivo: Xenopus oocytes (TEV) | [118] | |||
| KCNQ (Voltage-Gated Potassium Channel Subunits) 1, 2/3, 3/5 | 🗸 | PPDPPY | Polyubiquitination | Amplitude of K+ currents mediated by KCNQ2/3 and KCNQ3/5 were reduced by NEDD4-2. NEDD4-2 is activated by AMPK leading to reduced KCNQ1 expression | In vitro: HEK293 cells Ex vivo: Xenopus oocytes | [116,119,120,121] | ||
| Navs (Voltage-gated sodium channels) 1.2, 1.3, 1.5, 1.6, 1.7, 1.8 | 🗸 | PPSY, LPSY PGSP | Ubiquitination | Are vital in creating and propagating action potentials and reside in the membrane. NEDD4-2 interaction inhibits activity of multiple Navs, including the cardiac (Nav1.5) and neuronal Navs (Nav1.2, Nav1.7, and Nav1.8) | In vitro: HEK293 cells In vivo: SNS-NEDD4-2–/– mice, Pulldown of mouse brain lysates Ex vivo: Xenopus oocytes | [122,123,124] | ||
| NCC (Sodium Chloride symporter) | 🗸 | Ubiquitination | NCC ubiquitination at the cell surface Is achieved by NEDD4-2 and its deficiency upregulate NCC. NEDD4-2 may require another protein to achieve this | In vitro: HEK293 and mDCT15 cells Ex vivo: Xenopus Oocyte In vivo: Pax8-rtTA and TRE-LC1 transgenic mice | [125,126] | |||
| NEDD4-2(Neural precursor cell-expressed developmentally-downregulated gene/protein 4-2) | 🗸 | 🗸 | LPPY | Inhibitory self-ubiquitination of NEDD4–2 | Promotes NEDD4-2 stabilisation through auto-ubiquitination involving its own PY motif located on its HECT domain. This interaction may be between an active and non-active form | In vitro: HEK293 cells Ex vivo: Xenopus oocytes | [104,127,128] | |
| NEDD8 (Neural precursor cell-expressed developmentally-downregulated gene/protein 8) | 🗸 | Neddylation | Neddylation is a process whereby the ubiquitin-like protein, Nedd8, is conjugated to NEDD4-2 resulting in its degradation | In vitro: Mouse M1 kidney and mouse NCTC1469 liver cells In vivo: CYP4F2 transgenic mice | [129] | |||
| NHE3 (Sodium–hydrogen antiporter 3) | 🗸 | PPNY | Ubiquitination | An Na+/H+ exchanger that is expressed within the kidney where it is involved in blood pressure regulation through NaCl and HCO3− absorption. NHE is ubiquitinated by NEDD4-2. Disruption of NEDD4-2 interaction elevates human NHE3 expression and activity | In vitro: HEK 293 and PS120 cells | [130] | ||
| NKCC1/2 (Na-K-Cl cotransporter ½) | 🗸 | An Na+/K+/2Cl− co-transporter where NEDD4-2 is involved in its downregulation. NEDD4-2 indirectly suppresses NKCC1 expression | In vitro: HEK293T In vivo: Tam-induced NEDD4-2f/f; Vil-CreERT2 mice | [36] | ||||
| OAT (Organic ion transporters) 1/3 | 🗸 | Polyubiquitination and multiubiquitination | NEDD4-2 regulates cell surface OAT1/3 expression and its transport activities | In vitro: COS-7 and HEK293T cells | [131,132] | |||
| Occludin | 🗸 | PPPY | Polyubiquitination | An integral membrane protein that NEDD4–2 ubiquitinates. NEDD4-2 overexpression reduced occluding at tight junctions | In vitro: HEK293 and mplCCDc14 cells | [133] | ||
| PKA (Protein Kinase A) | 🗸 | Inhibitory phosphorylation of NEDD4-2. cAMP regulates ENaC through phosphorylation & inhibition of NEDD4-2 | In vitro: COS-7, FRT epithelial cells | [134] | ||||
| SCAMP3 (Secretory Carrier Membrane Protein 3) | 🗸 | PPAY | Multi-monoubiquitination | SCAMP3 ubiquitination is involved in the degradation of EGFR via the lysosome | In vitro HeLa and HEK293T cells | [87] | ||
| SGK1 (Serum/Glucocorticoid Regulated Kinase 1) | 🗸 | 🗸 | PPFY | Regulates the activity of several ion transport proteins. Inhibitory Phosphorylation of NEDD4-2 causes its interaction with 14-3-3 and subsequent degradation | In vitro: Kidney tubule epithelial and COS7 cells | [92,135,136] | ||
| SGLT1 (sodium-glucose linked transporter 1) | 🗸 | Ubiquitination | NEDD4-2 ubiquitinates and downregulates SGLT1 | Ex vivo: Xenopus oocytes | [137] | |||
| Smad (Mothers against decapentaplegic homolog) 2, 3, 4, 7 | 🗸 | PPPY | Polyubiquitination | NEDD4–2, interacts with Smads, inducing their polyubiquitination and degradation. This is not the case for Smad3 | In vitro: COS7 (Smad2) and HEK293T (Smad 2,3,4&7) cells | [136,138,139,140,141] | ||
| SP-C (Surfactant Protein C) | 🗸 | PPDY | Monoubiquitination or biubiquitination at K6 | NEDD4-2–mediated ubiquitination regulates luminal relocation of SP-C, leading to its processing and secretion | In vitro: HEK293 cells | [142] | ||
| TGF-βR1 (Transforming Growth Factor-β Receptor 1) | 🗸 | Plays a role in epithelial-mesenchymal transition via phosphorylation of small mothers against decapentaplegic (SMAD). NEDD4-2 suppresses its signalling | In vivo: Sprague Dawley rats. In vitro: NRK-52E cells | [143] | ||||
| TrkA(Tropomyosin-related kinase Trk A) | 🗸 | PPVY, PPSY, PPIY | Multi-monoubiquitination | Tropomyosin-related kinase (Trk) A is a receptor specifically for nerve growth factor and is downregulated by NEDD4-2 | In vitro: PC12-615 cells and primary cortical neurons | [144,145,146] | ||
| Tweety | 🗸 | PPTY | Ubiquitination | A family of chloride ion channels. NEDD4-2-mediated ubiquitination of TTYH2 regulates both cell surface and total levels of Tweety proteins | In vitro: HEK293 cells | [147] | ||
| ULK1 (Unc-51 like autophagy activating kinase 1) | 🗸 | Polyubiquitination | A serine–threonine kinase involved in autophagy. NEDD4-2 ubiquitinates ULK1 and targets it for proteasomal degradation | In vitro: HeLa cells | [148] | |||
| Usp2-45 (Ubiquitin-specific protease 2-45) | 🗸 | Adaptor of NEDD4-2 for ENaC Ubiquitination | In vitro: HEK293 cells | [149] | ||||
| WNK1 (With No Lysine Kinase) | 🗸 | PPQY, PFY | Ubiquitination | Serine-threonine kinases that regulate potassium, sodium, and blood-pressure homeostasis. Hormonal (Aldosterone, insulin and vasopressin) regulation of NEDD4-2 and WNK to regulate NCC (thiazide-sensitive NaCl cotransporter) | In vitro: mpkCCDc14 and HEK294T Cells In vivo: NEDD4-2fl/fl Pax8-rtTATRE-LC1 (Renal-specific NEDD4-2 KO) mice | [150] |
| Name | Substrate | Adaptor | Modifier/Regulator | Binding Motifs | Modification | Function | Experimental Model Used | References |
|---|---|---|---|---|---|---|---|---|
| ACK-1 (activated Cdc42-associated kinase 1) (NEDD4-1) | 🗸 | PPAY | Monoubiquitination, Polyubiquitination | ACK-1 is a cytoplasmic tyrosine kinase and is a NEDD4-1 and NEDD4-2 substrate. Its degradation through the proteasome results in downregulation of ACK-1. These authors suggest only NEDD4-1 to ubiquitinate ACK-1 | In vitro: COS7, HEK293T, HeLa, T47D, and A549 cells | [151] | ||
| ACK-1 (activated Cdc42-associated kinase 1) (NEDD4-2) | 🗸 | PPAY | Polyubiquitination | Its degradation through the proteasome results in downregulation of ACK-1 although this is contested [151] | In vitro: HeLa and COS7 cells | [152] | ||
| α-synuclein filaments (NEDD4-1) | 🗸 | PDNEAYEMP, PLY, PPLP PPSP, PFY | Monoubiquitination and Polyubiquitination (K63-linked) | Degraded via the lysosome. Potential protection mechanism against Parkinson’s Disease pathogenesis | In vitro: SH-SY5Y In vivo: Drosophila and Rat | [28,29,30] | ||
| α-synuclein filaments (NEDD4-2) | 🗸 | Polyubiquitination | In vitro: SH-SY5Y cells | [29] | ||||
| AKT (Protein Kinase B) (NEDD4-1) | 🗸 | LPEY, LPFY | Polyubiquitination (K63 and K48-linked) and multi-monoubiquitination | A critical effector kinase that regulates numerous cellular processes such as cell growth, death, differentiation, and migration.NEDD4-1 regulates nuclear trafficking of the activated form of AKT via the proteasome & enhances bortezomib sensitivity | In vitro: MCF-7, HeLa, and NEDD4+/+, and NEDD4−/− cells and MEFs | [153,154] | ||
| AKT (Protein Kinase B) (NEDD4-2) | 🗸 | Inhibitory phosphorylation of NEDD4-2. Inhibits NEDD4-2, increasing ENaC expression and Na+ absorption | In vitro: FRT cells | [155] | ||||
| AMPAR (NEDD4-1) | 🗸 | Ubiquitination | An ionotropic glutamate receptor. Ubiquitination leads to AMPAR internalisation and subsequent degradation | In vivo: Rat dissociated hippocampal or cortical neurons In vitro: HEK293 cells | [156,157,158] | |||
| AMPAR (NEDD4-2) | 🗸 | Ubiquitination | Ubiquitinates the GRIA1 subunit of AMPAR thereby mediating neuronal excitation | In vivo: Sprague Dawley (SD) rats, Nedd4-2andi and GluA1 knockout mice In vitro: HEK293 cells | [115,159,160] | |||
| AQP2 (Aquaporin 2) (NEDD4-1) | 🗸 | Polyubiquitination | Ubiquitinates and degrades AQP2 but requires NDFIP1 and NDFIP2 adaptors | In vitro: HEK293 and mpkCCD cells | [161] | |||
| AQP2 (Aquaporin 2) (NEDD4-2) | 🗸 | Polyubiquitination | Ubiquitinates and degrades AQP2 but requires NDFIP1 and NDFIP2 adaptors | In vitro: HEK293 and mpkCCD cells | [161] | |||
| Ca2+ (Calcium ions) (NEDD4-1) | 🗸 | Binds to the C2 domain of NEDD4-1 leading to the activation of its ligase activity and inhibition of auto-regulation. PIP2/IP3 ratio dictates its function in either the proximity of the membrane (PIP2) or cytoplasm (IP3) | Used biophysical techniques to complement the literature (NMR Spectroscopy) | [128] | ||||
| DLG3 Discs Large MAGUK Scaffold Protein 3) (NEDD4-1 and NEDD4-2) | 🗸 | PPGY, PPDY | Monoubiquitination | NEDD4-1 interaction results in Dlg3 monoubiquitination, apical membrane recruitment, and tight junction consolidation | In vitro: MDCK cells In vivo: Co-IP of mouse brain lysates | [162] | ||
| DVL2 (NEDD4-1 and NEDD4-2) | 🗸 | Ubiquitination | A protein involved in the Wnt pathway. Reported to be ubiquitinated for degradation via NEDD4-1 and NEDD4-2 | In vivo: NEDD4-1fl/fl and NEDD4-2fl/fl mice In vitro: HEK293T, DLD1 and HCT116 cells | [163] | |||
| ENaC (Epithelial Sodium Channel) (NEDD4-1) | 🗸 | PPNY, PPRY, PPAY | Ubiquitination of α and γ subunits | The epithelial Na+ channel is downregulated through proteosome degradation. Lack of binding motif in Liddle’s syndrome causes hyperactivity | In vitro: rat foetal distal lung epithelial and Xenopus-derived A6 cells Ex vivo: Xenopus oocyte | [164,165] | ||
| ENaC (Epithelial Sodium Channel) (NEDD4-2) | 🗸 | PPAY, PPNY, PPRY, PPKY | Monoubiquitination | NEDD4-2 catalyses ubiquitination and reduces expression of ENaC at the cell surface through lysosomal degradation. NEDD4-2 also targets Na+/Cl− cotransporter (NCC) in the kidney | In vitro: HEK293T cells | [116,126,166,167] | ||
| Grb10 (Growth Factor Receptor Bound Protein 10) (NEDD4-1) | 🗸 | An adaptor for NEDD4-1 to ubiquitinate and degrade IGF-IR. Required for indirect ubiquitination and degradation of VEGFR2 | In vitro: p6, p6/Grb10, HEK-293 EBNA and HUVEC cells | [42,168,169,170] | ||||
| Grb10 (Growth Factor Receptor Bound Protein 10) (NEDD4-2) | 🗸 | 🗸 | PQTPF | Associates with NEDD4-2 leading to its regulation and inhibition of ubiquitination and degradation of Kv1.3 channels | In vitro: HEK293 cells | [171] | ||
| LGR5 (Leucine-rich repeat containing G protein-coupled receptor 5) (NEDD4-1 and NEDD4-2) | 🗸 | Ubiquitination | A receptor for R-spondin and is a protein involved in the Wnt pathway. It is apparently degraded via the lysosome and proteasome involving NEDD4-1 and NEDD4-2 | In vivo: NEDD4-1fl/fl and NEDD4-2lfl/fl mice In vitro: HEK293T, DLD1 and HCT116 cells | [163] | |||
| MTMR4 (NEDD4-1) | 🗸 | PPLY | Ubiquitination | Myotubularin-related protein 4, an inositol phosphatase that regulates endosomal signalling | In vitro: HEK293 or HeLa cells | [172,173] | ||
| MTMR4 (NEDD4-2) | 🗸 | PPLY | MTMR4 reduces NEDD4-2-mediated proteasome degradation of wild type and mutant KCNQ1 and hERG channels via dephosphorylation | In vitro: Induced pluripotent stem cardiomyocytes from human dermal fibroblasts | [174] | |||
| NaV (Voltage-gated Sodium channels)1.2 and 1.7 (NEDD4-1) | 🗸 | PPSY (Proline Proline Serine Tyrosine) | Ubiquitination | Regulates sodium channels through lysosomal degradation | Ex vivo: Xenopus Oocytes | [175] | ||
| Navs (Voltage-gated Sodium channels) 1.2, 1.3,1.5, 1.6, 1.7, 1.8 (NEDD4-2) | 🗸 | PPSY, LPSY | Ubiquitination | NEDD4-2 ubiquitination inhibits activity of multiple Navs, including the cardiac (Nav1.5) and neuronal Navs (Nav1.2, Nav1.7, and Nav1.8) | In vitro: HEK-293 cells. In vivo: SNS-NEDD4-2–/– mice, Pulldown of mouse brain lysates Ex vivo: Xenopus oocytes | [42,116,122,123,124] | ||
| Ndfip1/2 (NEDD4 family-interacting proteins) (NEDD4-1) | 🗸 | 🗸 | NEDD4 family–interacting protein that activates the HECT domain of NEDD4-1. Ndfip1 binds substrates such as PTEN to act as an adaptor for NEDD4-1 ligase activity | In vitro: HEK-293T, SH-SY5Y, PC3 cells and primary MEFs In vivo: C57BL/6J, Emx1-Cre (C57BL/6) and Nestin-Cre mice (B6.Cg-Tg(Nes-cre)1Kln/J) | [176] | |||
| Ndfip1/2 (NEDD4 family-interacting proteins) (NEDD4-2 | 🗸 | 🗸 | PPPY, PPSY, LPTY, PSY, PTY | Ndfip activates HECT domain of NEDD4-2 | Ex vivo: Xenopus oocytes | [42,161,176] | ||
| OAT1 (organic anion transporter 1) (NEDD4-1) | 🗸 | Ubiquitination | Organic anion transporter (OAT). NEDD4-1 is an important regulator for hOAT1 ubiquitination, expression, and function via its WW2 and WW3 domains | In vitro: COS-7 and HEK293T cells | [131] | |||
| OAT1/3 (Organic ion transporters 1/3) (NEDD4-2) | 🗸 | Ubiquitination | NEDD4-2 ubiquitination regulates cell surface OAT1/3 expression with their WW3 and WW4 domains | In vitro: COS-7 and HEK293T cells | [131,132] | |||
| PTEN (Phosphatase and tensin homolog) (NEDD4-1& NEDD4-2) | 🗸? | 🗸 | PRR | Multi-monoubiquitinationPolyubiquitination(Ubiquitination independent of NEDD4-1) | PTEN is a tumour suppressor. Nuclear importation occurs via monoubiquitinationNEDD4-1 single and NEDD4-1 & NEDD4-2 double knock out mice do not exhibit stability, subcellular activity or localisation differences of PTEN | In vitro: PC3 and HEK293T cells In vivo: Xenopus tectumIn vivo: Murine | [111,176,177,178,179,180,181,182] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conway, J.A.; Kinsman, G.; Kramer, E.R. The Role of NEDD4 E3 Ubiquitin–Protein Ligases in Parkinson’s Disease. Genes 2022, 13, 513. https://doi.org/10.3390/genes13030513
Conway JA, Kinsman G, Kramer ER. The Role of NEDD4 E3 Ubiquitin–Protein Ligases in Parkinson’s Disease. Genes. 2022; 13(3):513. https://doi.org/10.3390/genes13030513
Chicago/Turabian StyleConway, James A., Grant Kinsman, and Edgar R. Kramer. 2022. "The Role of NEDD4 E3 Ubiquitin–Protein Ligases in Parkinson’s Disease" Genes 13, no. 3: 513. https://doi.org/10.3390/genes13030513
APA StyleConway, J. A., Kinsman, G., & Kramer, E. R. (2022). The Role of NEDD4 E3 Ubiquitin–Protein Ligases in Parkinson’s Disease. Genes, 13(3), 513. https://doi.org/10.3390/genes13030513