
MicroRNA-17-92a-1 Host Gene (MIR17HG) Expression Signature and rs4284505 Variant Association with Alopecia Areata: A Case–Control Study

 ,
,  , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Clinical Assessment
2.3. Allelic Discrimination of rs4284505 Variant
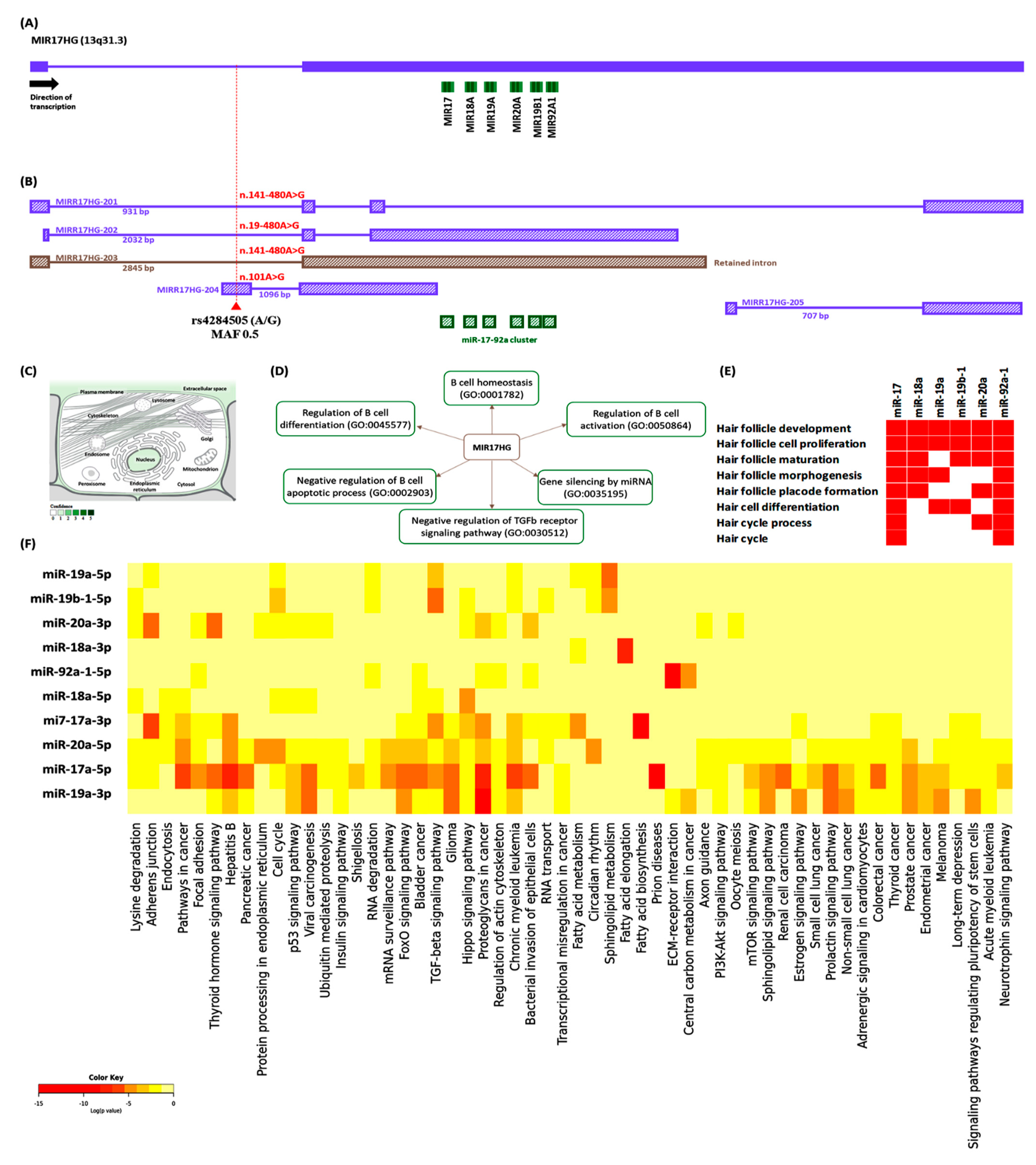
2.4. MIR17HG Expression Profiling
2.5. Statistical Analysis
3. Results
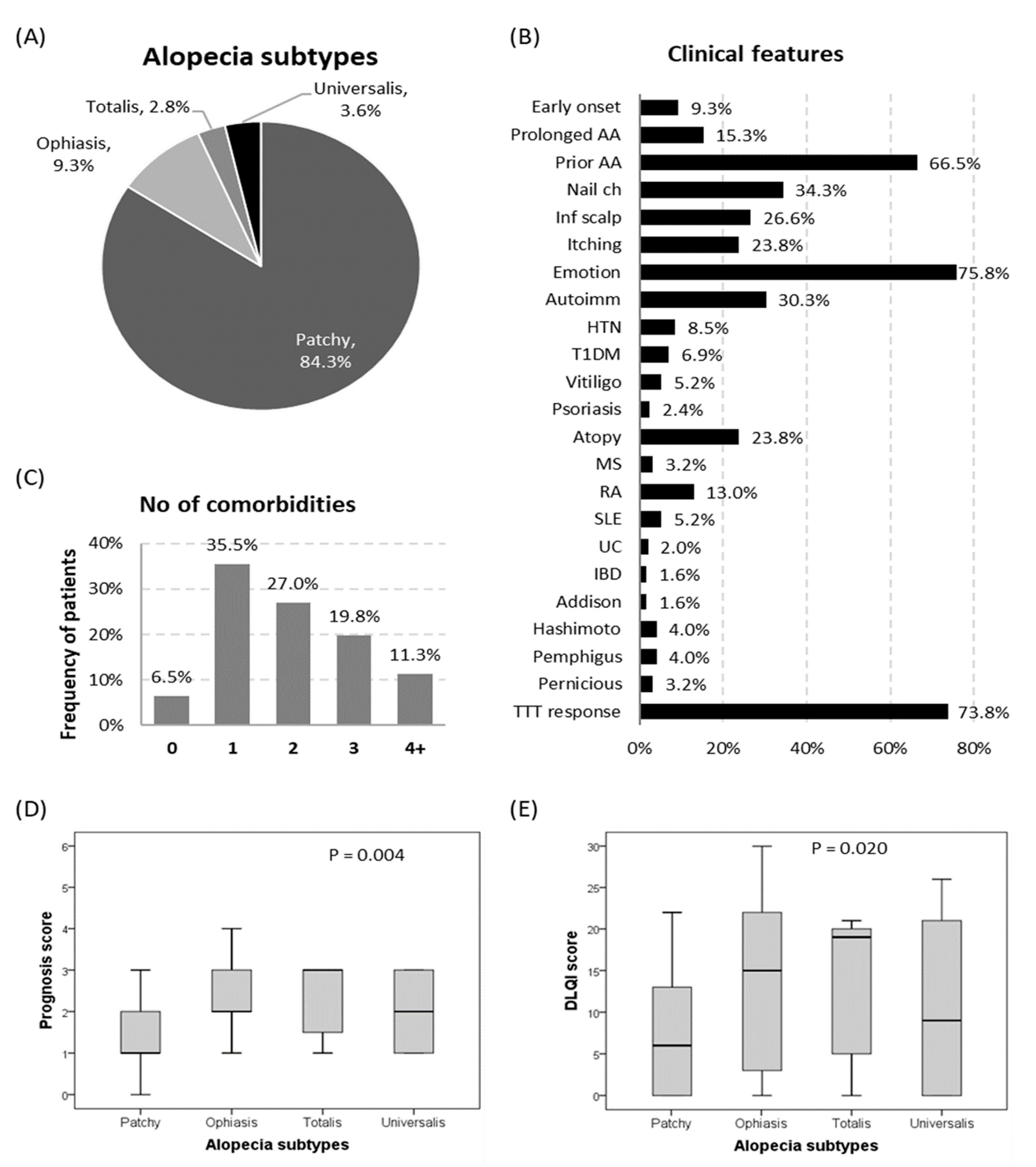
3.1. Characteristics of the Study Population
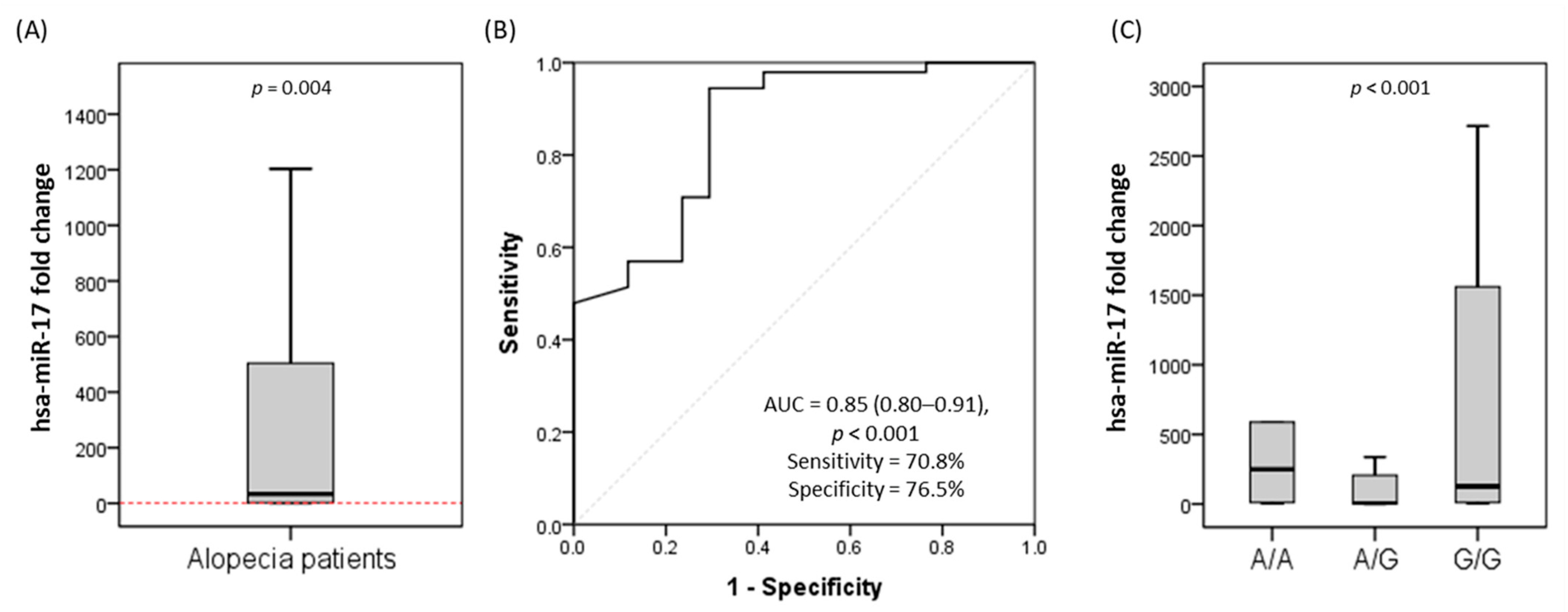
3.2. Allelic Discrimination Analysis of rs4284505 SNP
3.3. Relative Expression of MIR17HG
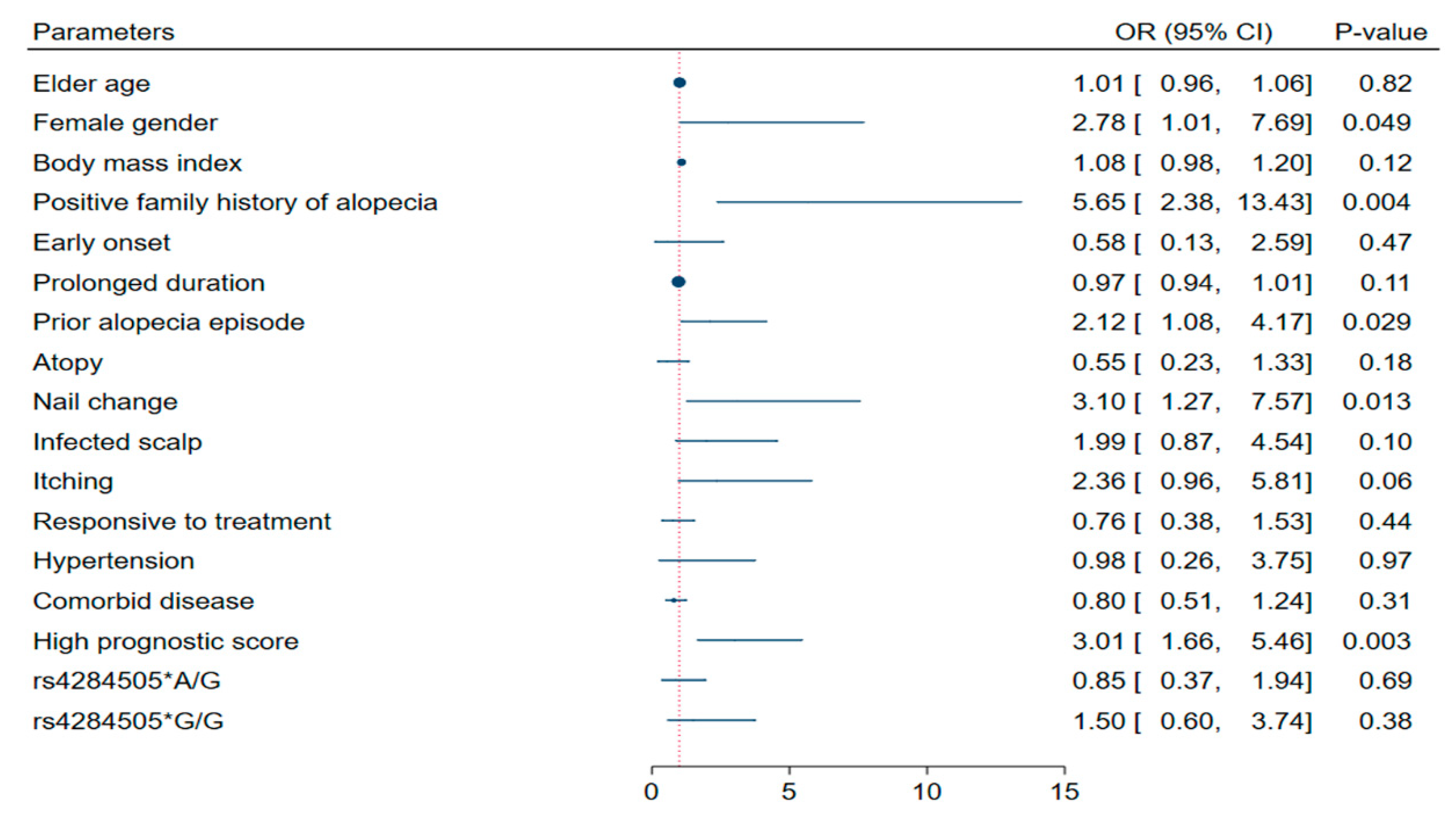
3.4. Association with Disease Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pratt, C.H.; King, L.E.; Messenger, A.G.; Christiano, A.M.; Sundberg, J.P. Alopecia areata. Nat. Rev. Dis. Primers 2017, 3, 17011. [Google Scholar] [CrossRef]
- Strazzulla, L.C.; Wang, E.H.C.; Avila, L.; Lo Sicco, K.; Brinster, N.; Christiano, A.M.; Shapiro, J. Alopecia areata: Disease characteristics, clinical evaluation, and new perspectives on pathogenesis. J. Am. Acad. Dermatol. 2018, 78, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Dai, Z.; Jabbari, A.; Cerise, J.E.; Higgins, C.A.; Gong, W.; de Jong, A.; Harel, S.; DeStefano, G.M.; Rothman, L.; et al. Alopecia areata is driven by cytotoxic T lymphocytes and is reversed by JAK inhibition. Nat. Med. 2014, 20, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Petukhova, L.; Duvic, M.; Hordinsky, M.; Norris, D.; Price, V.; Shimomura, Y.; Kim, H.; Singh, P.; Lee, A.; Chen, W.V.; et al. Genome-wide association study in alopecia areata implicates both innate and adaptive immunity. Nature 2010, 466, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Al-Eitan, L.N.; Al Momani, R.O.; Al Momani, K.K.; Al Warawrah, A.M.; Aljamal, H.A.; Alghamdi, M.A.; Muhanna, A.M.; Al-Qarqaz, F.A. Candidate Gene Analysis of Alopecia Areata In Jordanian Population of Arab Descent: A Case-Control Study. Appl. Clin. Genet. 2019, 12, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, X.; Zhang, R. Pathogenesis of Alopecia Areata Based on Bioinformatics Analysis. Indian J. Dermatol. 2019, 64, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lekka, E.; Hall, J. Noncoding RNAs in disease. FEBS Lett. 2018, 592, 2884–2900. [Google Scholar] [CrossRef]
- Andl, T.; Murchison, E.P.; Liu, F.; Zhang, Y.; Yunta-Gonzalez, M.; Tobias, J.W.; Andl, C.D.; Seykora, J.T.; Hannon, G.J.; Millar, S.E. The miRNA-processing enzyme dicer is essential for the morphogenesis and maintenance of hair follicles. Curr. Biol. 2006, 16, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Teta, M.; Choi, Y.S.; Okegbe, T.; Wong, G.; Tam, O.H.; Chong, M.M.; Seykora, J.T.; Nagy, A.; Littman, D.R.; Andl, T.; et al. Inducible deletion of epidermal Dicer and Drosha reveals multiple functions for miRNAs in postnatal skin. Development 2012, 139, 1405–1416. [Google Scholar] [CrossRef] [PubMed]
- Tavakolpour, S.; Elkaei Behjati, S. The ignored roles of microRNAs in alopecia areata. Dermatol. Ther. 2017, 30. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.H.C.; DeStefano, G.M.; Patel, A.V.; Drill, E.; Harel, S.; Cela, C.; Tavazoie, M.; Christiano, A.M. Identification of differentially expressed miRNAs in alopecia areata that target immune-regulatory pathways. Genes Immun. 2017, 18, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.; Qi, S.; Hu, R.; Zhao, J.; Rui, W.; Miao, Y.; Ma, J.; Yang, Q. Identification of blood microRNA alterations in patients with severe active alopecia areata. J. Cell Biochem. 2019, 120, 14421–14430. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Weng, T.; Zhang, W.; Zhang, M.; He, X.; Han, C.; Wang, X. The Roles of Non-coding R.N.A. in the Development and Regeneration of Hair Follicles: Current Status and Further Perspectives. Front. Cell Dev. Biol. 2021, 9, 720879. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of mammalian microRNA host genes and transcription units. Genome Res. 2004, 14, 1902–1910. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.F.; Niu, P.; Wang, L.; Vogelsang, M.; Gaur, M.; Krastins, B.; Zhao, Y.; Smagul, A.; Nussupbekova, A.; Akanov, A.A.; et al. Opposing activities of oncogenic MIR17HG and tumor suppressive MIR100HG clusters and their gene targets regulate replicative senescence in human adult stem cells. NPJ Aging Mech. Dis. 2017, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Comabella, M.; Craig, D.W.; Camiña-Tato, M.; Morcillo, C.; Lopez, C.; Navarro, A.; Rio, J.; Montalban, X.; Martin, R.; Group, B.S. Identification of a novel risk locus for multiple sclerosis at 13q31.3 by a pooled genome-wide scan of 500,000 single nucleotide polymorphisms. PLoS ONE 2008, 3, e3490. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Gawad, A.R.; Shaheen, S.; Babteen, N.A.; Toraih, E.A.; Elshazli, R.M.; Fawzy, M.S.; Gouda, N.S. Association of microRNA 17 host gene variant (rs4284505) with susceptibility and severity of systemic lupus erythematosus. Immun. Inflamm. Dis. 2020, 8, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Olsen, E.A.; Hordinsky, M.K.; Price, V.H.; Roberts, J.L.; Shapiro, J.; Canfield, D.; Duvic, M.; King, L.E.; McMichael, A.J.; Randall, V.A.; et al. Alopecia areata investigational assessment guidelines—Part II. National Alopecia Areata Foundation. J. Am. Acad. Dermatol. 2004, 51, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Toraih, E.A.; Ameen, H.M.; Hussein, M.H.; Youssef Elabd, A.A.; Mohamed, A.M.; Abdel-Gawad, A.R.; Fawzy, M.S. Association of Autoimmune Regulator Gene Rs2075876 Variant, but Not Gene Expression with Alopecia Areata in Males: A Case-control Study. Immunol. Investig. 2020, 49, 146–165. [Google Scholar] [CrossRef] [PubMed]
- Seetharam, K.A. Alopecia areata: An update. Indian J. Dermatol. Venereol. Leprol. 2013, 79, 563–575. [Google Scholar] [CrossRef]
- Kutyavin, I.V.; Afonina, I.A.; Mills, A.; Gorn, V.V.; Lukhtanov, E.A.; Belousov, E.S.; Singer, M.J.; Walburger, D.K.; Lokhov, S.G.; Gall, A.A.; et al. 3′-minor groove binder-DNA probes increase sequence specificity at PCR extension temperatures. Nucleic Acids Res. 2000, 28, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Toraih, E.A.; Aly, N.M.; Abdallah, H.Y.; Al-Qahtani, S.A.; Shaalan, A.A.; Hussein, M.H.; Fawzy, M.S. MicroRNA-target cross-talks: Key players in glioblastoma multiforme. Tumour Biol. 2017, 39, 1010428317726842. [Google Scholar] [CrossRef] [PubMed]
- Toraih, E.A.; Ibrahiem, A.T.; Fawzy, M.S.; Hussein, M.H.; Al-Qahtani, S.A.M.; Shaalan, A.A.M. MicroRNA-34a: A Key Regulator in the Hallmarks of Renal Cell Carcinoma. Oxid. Med. Cell Longev. 2017, 2017, 3269379. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Fawzy, M.S.; Hussein, M.H.; Abdelaziz, E.Z.; Yamany, H.A.; Ismail, H.M.; Toraih, E.A. Association of MicroRNA-196a2 Variant with Response to Short-Acting β2-Agonist in COPD: An Egyptian Pilot Study. PLoS ONE 2016, 11, e0152834. [Google Scholar] [CrossRef]
- Mogilyansky, E.; Rigoutsos, I. The miR-17/92 cluster: A comprehensive update on its genomics, genetics, functions and increasingly important and numerous roles in health and disease. Cell Death Differ. 2013, 20, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Huang, T.; Ye, Z.; Fu, X.; Hu, K.; Yang, X. Correlation of MicroRNA 17-92 Cluster Host Gene (MIR17HG) Polymorphisms with Susceptibility and Prognosis for Multiple Myeloma. Clin. Lymphoma Myeloma Leuk. 2019, 19, e359–e366. [Google Scholar] [CrossRef] [PubMed]
- Chacon-Cortes, D.; Smith, R.A.; Lea, R.A.; Youl, P.H.; Griffiths, L.R. Association of microRNA 17-92 cluster host gene (MIR17HG) polymorphisms with breast cancer. Tumour Biol. 2015, 36, 5369–5376. [Google Scholar] [CrossRef] [PubMed]
- Silva-Sousa, A.C.; Mazzi-Chaves, J.F.; Freitas, J.V.; Salles, A.G.; Segato, R.A.B.D.; Silva, L.A.B.D.; Antunes, L.A.A.; Antunes, L.S.; Baratto-Filho, F.; Sousa-Neto, M.D.; et al. Association between Estrogen, Vitamin D and Microrna17 Gene Polymorphisms and Periapical Lesions. Braz. Dent. J. 2020, 31, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Dimas, A.S.; Deutsch, S.; Stranger, B.E.; Montgomery, S.B.; Borel, C.; Attar-Cohen, H.; Ingle, C.; Beazley, C.; Gutierrez Arcelus, M.; Sekowska, M.; et al. Common regulatory variation impacts gene expression in a cell type-dependent manner. Science 2009, 325, 1246–1250. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Orozco, G. Identification of rheumatoid arthritis causal genes using functional genomics. Scand. J. Immunol. 2019, 89, e12753. [Google Scholar] [CrossRef]
- Yang, K.; Zhang, Y.; Mai, X.; Hu, L.; Ma, C.; Wei, J. MIR17HG genetic variations affect the susceptibility of IgA nephropathy in Chinese Han people. Gene 2021, 800, 145838. [Google Scholar] [CrossRef] [PubMed]
- Aksenenko, M.; Palkina, N.; Komina, A.; Ruksha, T. MiR-92a-1-5p and miR-328-3p Are Up-Regulated in Skin of Female Pattern Hair Loss Patients. Ann. Dermatol. 2019, 31, 256–259. [Google Scholar] [CrossRef]
- Goodarzi, H.R.; Abbasi, A.; Saffari, M.; Tabei, M.B.; Noori Daloii, M.R. MicroRNAs take part in pathophysiology and pathogenesis of Male Pattern Baldness. Mol. Biol. Rep. 2010, 37, 2959–2965. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, H.R.; Abbasi, A.; Saffari, M.; Fazelzadeh Haghighi, M.; Tabei, M.B.; Noori Daloii, M.R. Differential expression analysis of balding and nonbalding dermal papilla microRNAs in male pattern baldness with a microRNA amplification profiling method. Br. J. Dermatol. 2012, 166, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yi, X.; An, Y.; Guo, S.; Li, S.; Song, P.; Chang, Y.; Zhang, S.; Gao, T.; Wang, G.; et al. MicroRNA-17-92 cluster promotes the proliferation and the chemokine production of keratinocytes: Implication for the pathogenesis of psoriasis. Cell Death Dis. 2018, 9, 567. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Bi, X.; Qu, L.; Han, L.; Yin, C.; Deng, J.; Dong, Z.; Mi, Q.S.; Zhou, L. miRNA miR-17-92 cluster is differentially regulated in the imiqumod-treated skin but is not required for imiqumod-induced psoriasis-like dermatitis in mice. Exp. Dermatol. 2017, 26, 82–84. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.H.; Zhu, X.H.; Liang, J.; Wu, J.F.; Yang, Y.S.; Xu, J.H. The expression and significance of miR-17-92 cluster miRs in CD4+ T cells from patients with systemic lupus erythematosus. Clin. Exp. Rheumatol. 2013, 31, 472–473. [Google Scholar] [PubMed]
- Dai, R.; Zhang, Y.; Khan, D.; Heid, B.; Caudell, D.; Crasta, O.; Ahmed, S.A. Identification of a common lupus disease-associated microRNA expression pattern in three different murine models of lupus. PLoS ONE 2010, 5, e14302. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, R.L.; Hoffmann, F.; Mehling, M.; Kuhle, J.; Kappos, L. Altered expression of miR-17-5p in CD4+ lymphocytes of relapsing-remitting multiple sclerosis patients. Eur. J. Immunol. 2010, 40, 888–898. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Ohuchida, K.; Mizumoto, K.; Fujita, H.; Nakata, K.; Tanaka, M. MicroRNA miR-17-5p is overexpressed in pancreatic cancer, associated with a poor prognosis, and involved in cancer cell proliferation and invasion. Cancer Biol. Ther. 2010, 10, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, C.; Zhang, R.; Gao, X.; Qu, X.; Zhao, M.; Qiao, C.; Xu, J.; Li, J. miR-17-92 cluster microRNAs confers tumorigenicity in multiple myeloma. Cancer Lett. 2011, 309, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Rao, E.; Jiang, C.; Ji, M.; Huang, X.; Iqbal, J.; Lenz, G.; Wright, G.; Staudt, L.M.; Zhao, Y.; McKeithan, T.W.; et al. The miRNA-17∼92 cluster mediates chemoresistance and enhances tumor growth in mantle cell lymphoma via PI3K/AKT pathway activation. Leukemia 2012, 26, 1064–1072. [Google Scholar] [CrossRef]
- Chen, Q.; Si, Q.; Xiao, S.; Xie, Q.; Lin, J.; Wang, C.; Chen, L.; Wang, L. Prognostic significance of serum miR-17-5p in lung cancer. Med. Oncol. 2013, 30, 353. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Han, C.; Wu, T. MiR-17-92 cluster promotes hepatocarcinogenesis. Carcinogenesis 2015, 36, 1213–1222. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Qian, X.; Li, H.; Zhang, X. Combinations of elevated tissue miRNA-17-92 cluster expression and serum prostate-specific antigen as potential diagnostic biomarkers for prostate cancer. Oncol. Lett. 2017, 14, 6943–6949. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jepsen, R.K.; Novotny, G.W.; Klarskov, L.L.; Bang-Berthelsen, C.H.; Haakansson, I.T.; Hansen, A.; Christensen, I.J.; Riis, L.B.; Høgdall, E. Early metastatic colorectal cancers show increased tissue expression of miR-17/92 cluster members in the invasive tumor front. Hum. Pathol. 2018, 80, 231–238. [Google Scholar] [CrossRef]
- Khalifa, M.M.; Zaki, N.E.; Nazier, A.A.; Moussa, M.A.; Haleem, R.A.; Rabie, M.A.; Mansour, A.R. Prognostic significance of microRNA 17-92 cluster expression in Egyptian chronic lymphocytic leukemia patients. J. Egypt. Natl. Cancer Inst. 2021, 33, 37. [Google Scholar] [CrossRef] [PubMed]
- AlDeen, H.G.; Ramadan, A.; Awadallah, E.; Alnaggar, A.R.; Khalil, N.M.; Behiry, M.E.; Emara, N.; Nasr, S.A.; Ali, A.; Yousef, R.N. Patterns of microRNAs 142-3p, 106a, 17 and 20a expression in patients with systemic lupus erythematosus. Egypt. Rheumatol. 2022, 44, 31–35. [Google Scholar] [CrossRef]
- Carlsen, A.L.; Schetter, A.J.; Nielsen, C.T.; Lood, C.; Knudsen, S.; Voss, A.; Harris, C.C.; Hellmark, T.; Segelmark, M.; Jacobsen, S.; et al. Circulating microRNA expression profiles associated with systemic lupus erythematosus. Arthritis Rheum. 2013, 65, 1324–1334. [Google Scholar] [CrossRef]
- Cox, M.B.; Cairns, M.J.; Gandhi, K.S.; Carroll, A.P.; Moscovis, S.; Stewart, G.J.; Broadley, S.; Scott, R.J.; Booth, D.R.; Lechner-Scott, J.; et al. MicroRNAs miR-17 and miR-20a inhibit T cell activation genes and are under-expressed in MS whole blood. PLoS ONE 2010, 5, e12132. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Singh, A.K.; Ahmed, S. MicroRNA-17 Suppresses TNF-α Signaling by Interfering with TRAF2 and cIAP2 Association in Rheumatoid Arthritis Synovial Fibroblasts. J. Immunol. 2016, 197, 2219–2228. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.C.; Wade, S.; Floudas, A.; Orr, C.; McGarry, T.; Cregan, S.; Fearon, U.; Veale, D.J. Serum miRNA Signature in Rheumatoid Arthritis and “At-Risk Individuals”. Front. Immunol. 2021, 12, 633201. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Srinivasan, L.; Calado, D.P.; Patterson, H.C.; Zhang, B.; Wang, J.; Henderson, J.M.; Kutok, J.L.; Rajewsky, K. Lymphoproliferative disease and autoimmunity in mice with increased miR-17-92 expression in lymphocytes. Nat. Immunol. 2008, 9, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Li, C.; Olive, V.; Lykken, E.; Feng, F.; Sevilla, J.; Wan, Y.; He, L.; Li,, Q.J. Molecular dissection of the miR-17-92 cluster’s critical dual roles in promoting Th1 responses and preventing inducible Treg differentiation. Blood 2011, 118, 5487–5497. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Q.; Jiang, S.; Li, C.; Zhang, B.; Li,, Q.J. miR-17-92 cluster targets phosphatase and tensin homology and Ikaros Family Zinc Finger 4 to promote TH17-mediated inflammation. J. Biol. Chem. 2014, 289, 12446–12456. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.Y.; Barbi, J.; Wu, C.Y.; Zheng, Y.; Vignali, P.D.; Wu, X.; Tao, J.H.; Park, B.V.; Bandara, S.; Novack, L.; et al. MicroRNA-17 Modulates Regulatory T Cell Function by Targeting Co-regulators of the Foxp3 Transcription Factor. Immunity 2016, 45, 83–93. [Google Scholar] [CrossRef]
- Han, Y.M.; Sheng, Y.Y.; Xu, F.; Qi, S.S.; Liu, X.J.; Hu, R.M.; Miao, Y.; Huang, G.Q.; Yang, Q.P. Imbalance of T-helper 17 and regulatory T cells in patients with alopecia areata. J. Dermatol. 2015, 42, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Kuo, G.; Wu, C.Y.; Yang, H.Y. MiR-17-92 cluster and immunity. J. Formos. Med. Assoc. 2019, 118, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Pan, C.; Li, L.; Bian, Z.; Lv, Z.; Shi, L.; Zhang, J.; Li, D.; Gu, H.; Zhang, C.Y.; et al. MicroRNA-17/20a/106a modulate macrophage inflammatory responses through targeting signal-regulatory protein α. J. Allergy Clin. Immunol. 2013, 132, 426–436.e8. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Categories | Controls | AA | p-Value | OR (95% CI) |
|---|---|---|---|---|---|
| Age, years | ≤30 years | 144 (59) | 125 (50.4) | 0.05 | Reference |
| >30 years | 100 (41) | 123 (49.6) | 1.41 (0.99–2.02) | ||
| Sex | Male | 198 (81.1) | 216 (87.1) | 0.08 | Reference |
| Female | 46 (18.9) | 32 (12.9) | 0.63 (0.39–1.04) | ||
| Obesity | Negative | 234 (95.9) | 229 (92.3) | 0.12 | Reference |
| Positive | 10 (4.1) | 19 (7.7) | 1.94 (0.88–4.26) | ||
| Residence | Port-Said | 18 (7.4) | 13 (5.2) | 0.29 | Reference |
| Suez | 22 (9) | 15 (6) | 0.94 (0.35–2.48) | ||
| Ismailia | 62 (25.4) | 77 (31) | 1.71 (0.78–3.78) | ||
| Cairo | 142 (58.2) | 143 (57.7) | 1.39 (0.65–2.95) | ||
| Occupation | Student | 96 (39.3) | 107 (43.1) | 0.16 | Reference |
| Unemployed | 100 (41) | 108 (43.5) | 0.96 (0.65–1.42) | ||
| Employed | 48 (19.7) | 33 (13.3) | 0.61 (0.36–1.03) | ||
| Family history | Alopecia | 0 (0) | 108 (43.5) | N.A. | |
| Autoimmune disorders | 0 (0) | 105 (42.3) | NA. |
| Characteristics | Alopecia Areata (Patchy/Aphiasis) | Alopecia Totalis/Universalis | p-Value | OR (95% CI) | |
|---|---|---|---|---|---|
| Number | 173 (69.8) | 75 (30.2) | |||
| Demographic characteristics | |||||
| Age, years | Mean ± SD | 30.9 ± 7.1 | 30.1 ± 7.0 | 0.52 | |
| Sex | Male | 150 (86.7) | 66 (88) | 0.84 | Reference |
| Female | 23 (13.3) | 9 (12) | 0.88 (0.39–2.02) | ||
| BMI, kg/m2 | Mean ± SD | 26.0 ± 2.8 | 25.1 ± 2.7 | 0.06 | |
| Family history | Alopecia | 75 (43.4) | 33 (44) | 0.94 | 1.02 (0.59–1.77) |
| Autoimmune | 74 (42.8) | 31 (41.3) | 0.88 | 0.94 (0.54–1.63) | |
| Prior episode of alopecia | No | 57 (32.9) | 26 (34.7) | 0.88 | Reference |
| Yes | 116 (67.1) | 49 (65.3) | 0.92 (0.52–1.64) | ||
| Duration of disease, months | Mean ± SD | 4.8 ± 9.7 | 4.5 ± 8.3 | 0.91 | |
| Age at onset | Mean ± SD | 30.5 ± 7.0 | 29.7 ± 7.1 | 0.64 | |
| Disease characteristics | |||||
| Nail changes | Absent | 125 (72.3) | 38 (50.7) | 0.001 | Reference |
| Present | 48 (27.7) | 37 (49.3) | 2.53 (1.44–4.44) | ||
| Itching | Absent | 137 (79.2) | 52 (69.3) | 0.10 | Reference |
| Present | 36 (20.8) | 23 (30.7) | 1.68 (0.91–3.10) | ||
| Scalp infection | Absent | 134 (77.5) | 48 (64) | 0.041 | Reference |
| Present | 39 (22.5) | 27 (36) | 1.93 (1.07–3.49) | ||
| Atopy | Absent | 157 (75.1) | 32 (82.1) | 0.41 | Reference |
| Present | 52 (24.9) | 7 (17.9) | 0.66 (0.27–1.58) | ||
| Hypertension | Absent | 190 (90.9) | 37 (94.9) | 0.54 | Reference |
| Present | 19 (9.1) | 2 (5.1) | 0.54 (0.12–2.42) | ||
| Emotional stress | Absent | 52 (24.9) | 8 (20.5) | 0.68 | Reference |
| Present | 157 (75.1) | 31 (79.5) | 1.28 (0.55–2.96) | ||
| Concomitant autoimmune disease | Absent | 143 (68.4) | 30 (76.9) | 0.34 | Reference |
| Present | 66 (31.6) | 9 (23.1) | 0.65 (0.29–1.44) | ||
| SALT score | Mean ± SD | 9.4 ± 8.8 | 13.8 ± 10.0 | <0.001 | |
| Prognostic score | Mean ± SD | 1.61 ± 1.1 | 2.30 ± 1.02 | <0.001 | |
| DLQI score | Mean ± SD | 9.4 ± 8.8 | 13.8 ± 10.0 | 0.003 | |
| Responded to treatment | No | 51 (29.5) | 14 (18.7) | 0.08 | Reference |
| Yes | 122 (70.5) | 61 (81.3) | 1.82 (0.93–3.54) | ||
| All Subjects | Controls | Cases | |||||
|---|---|---|---|---|---|---|---|
| Variable | Count | Proportion | Count | Proportion | Count | Proportion | p-value |
| Allele | |||||||
| G | 575 | 0.58 | 291 | 0.60 | 284 | 0.57 | 0.45 |
| A | 409 | 0.42 | 197 | 0.40 | 212 | 0.43 | |
| Genotypes | |||||||
| A/A | 84 | 0.17 | 46 | 0.19 | 38 | 0.15 | 0.032 |
| G/A | 241 | 0.49 | 105 | 0.43 | 136 | 0.55 | |
| G/G | 167 | 0.34 | 93 | 0.38 | 74 | 0.30 | |
| HWE | |||||||
| p-value | 0.93 | 0.11 | 0.07 |
| Model | Genotype | Controls | Cases | Crude OR (95% CI) | p-Value | Adjusted OR (95% CI) | p-Value |
|---|---|---|---|---|---|---|---|
| Codominant | G/G | 93 (38.1%) | 74 (29.8%) | 1.00 | 0.032 | 1.00 | 0.044 |
| A/G | 105 (43%) | 136 (54.8%) | 1.63 (1.09–2.42) | 1.36 (0.84–2.20) | |||
| A/A | 46 (18.9%) | 38 (15.3%) | 1.04 (0.61–1.76) | 0.60 (0.30–1.20) | |||
| Dominant | G/G | 93 (38.1%) | 74 (29.8%) | 1.00 | 0.052 | 1.00 | 0.66 |
| A/G–A/A | 151 (61.9%) | 174 (70.2%) | 1.45 (1.00–2.11) | 1.11 (0.70–1.74) | |||
| Recessive | G/G–A/G | 198 (81.2%) | 210 (84.7%) | 1.00 | 0.30 | 1.00 | 0.03 |
| A/A | 46 (18.9%) | 38 (15.3%) | 0.78 (0.49–1.25) | 0.51 (0.27–0.96) | |||
| Overdominant | G/G–A/A | 139 (57%) | 112 (45.2%) | 1.00 | 0.008 | 1.00 | 0.043 |
| A/G | 105 (43%) | 136 (54.8%) | 1.61 (1.13–2.29) | 1.57 (1.01–2.45) | |||
| Log-additive | --- | --- | --- | 1.10 (0.86–1.42) | 0.45 | 0.88 (0.64–1.20) | 0.41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faisal, S.; Toraih, E.A.; Atef, L.M.; Hassan, R.; Fouad, M.M.; Al Ageeli, E.; Fawzy, M.S.; Abdalla, H.A. MicroRNA-17-92a-1 Host Gene (MIR17HG) Expression Signature and rs4284505 Variant Association with Alopecia Areata: A Case–Control Study. Genes 2022, 13, 505. https://doi.org/10.3390/genes13030505
Faisal S, Toraih EA, Atef LM, Hassan R, Fouad MM, Al Ageeli E, Fawzy MS, Abdalla HA. MicroRNA-17-92a-1 Host Gene (MIR17HG) Expression Signature and rs4284505 Variant Association with Alopecia Areata: A Case–Control Study. Genes. 2022; 13(3):505. https://doi.org/10.3390/genes13030505
Chicago/Turabian StyleFaisal, Salwa, Eman A. Toraih, Lina M. Atef, Ranya Hassan, Marwa M. Fouad, Essam Al Ageeli, Manal S. Fawzy, and Hussein Abdelaziz Abdalla. 2022. "MicroRNA-17-92a-1 Host Gene (MIR17HG) Expression Signature and rs4284505 Variant Association with Alopecia Areata: A Case–Control Study" Genes 13, no. 3: 505. https://doi.org/10.3390/genes13030505
APA StyleFaisal, S., Toraih, E. A., Atef, L. M., Hassan, R., Fouad, M. M., Al Ageeli, E., Fawzy, M. S., & Abdalla, H. A. (2022). MicroRNA-17-92a-1 Host Gene (MIR17HG) Expression Signature and rs4284505 Variant Association with Alopecia Areata: A Case–Control Study. Genes, 13(3), 505. https://doi.org/10.3390/genes13030505