
The Genome of Rhyzopertha dominica (Fab.) (Coleoptera: Bostrichidae): Adaptation for Success


 , ,
, ,  ,
,  ,
,  , , ,
, , ,  , , , , ,
, , , , , 
 ,
,  ,
,  , , and
, , and
Abstract
1. Introduction
2. Methods
2.1. Insect Strain
2.2. Photography
2.3. Measurement of Genome Size
2.4. Extraction of Nucleic Acids, Sequencing, and Assembly
2.4.1. Genome Sequencing and Assembly
2.4.2. Transcriptome Sequencing and Gene Expression Analysis
2.5. Post Genome Analysis
2.5.1. Gene Prediction and Annotation
2.5.2. Manual Annotation
2.5.3. Phylogenetic Analysis
2.5.4. Expression Analysis
2.5.5. Repeat Structure Analysis
3. Results
3.1. Sequencing and Assembly
3.2. Manual Annotation
3.2.1. Detoxification Genes
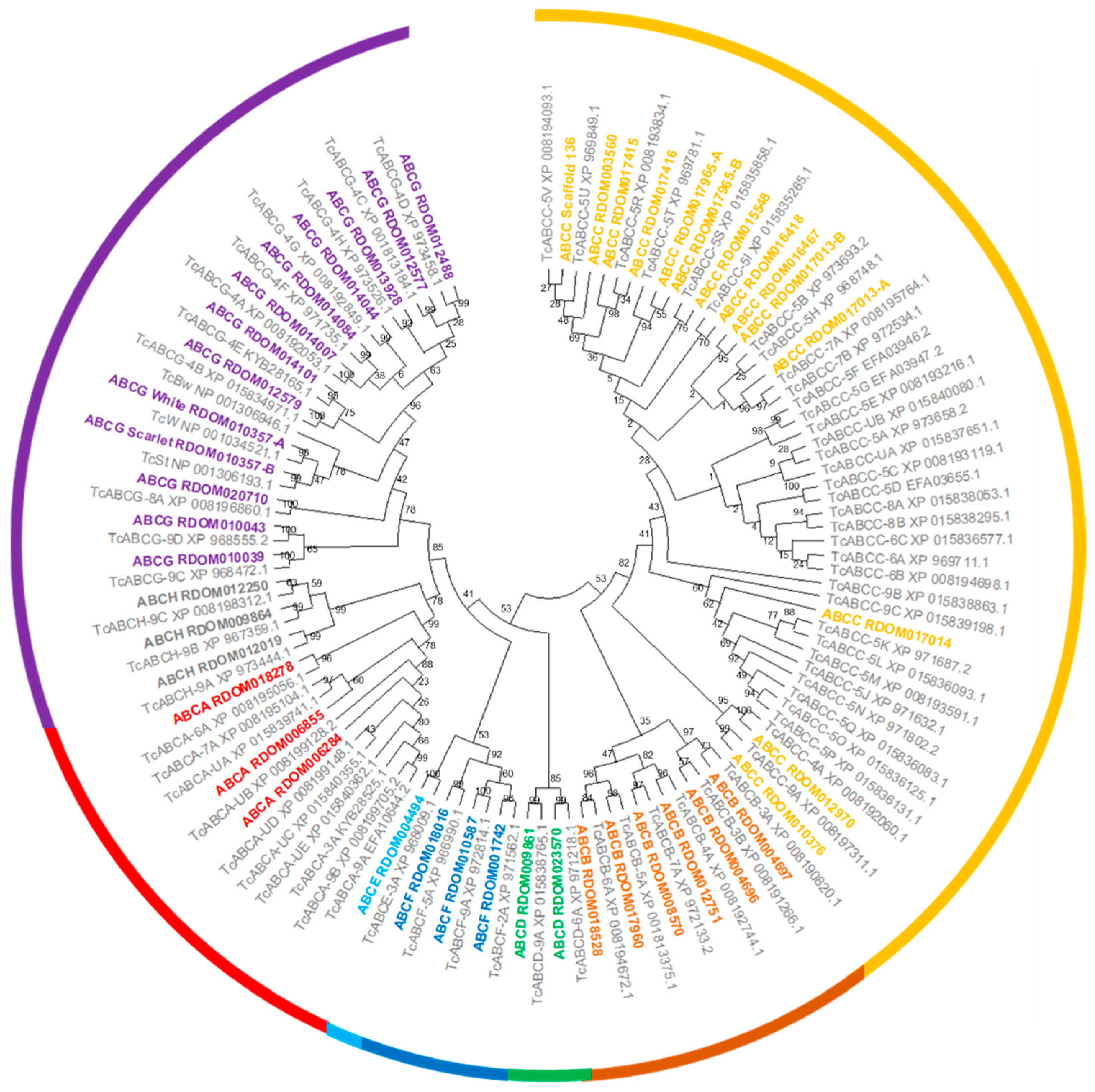
ATP Binding Cassette Transporters
Cytochrome P450s, UDP-Glucuronosyltransferases, Glutathione S-Transferases, and Carboxylesterases
3.2.2. Chemosensory Biology
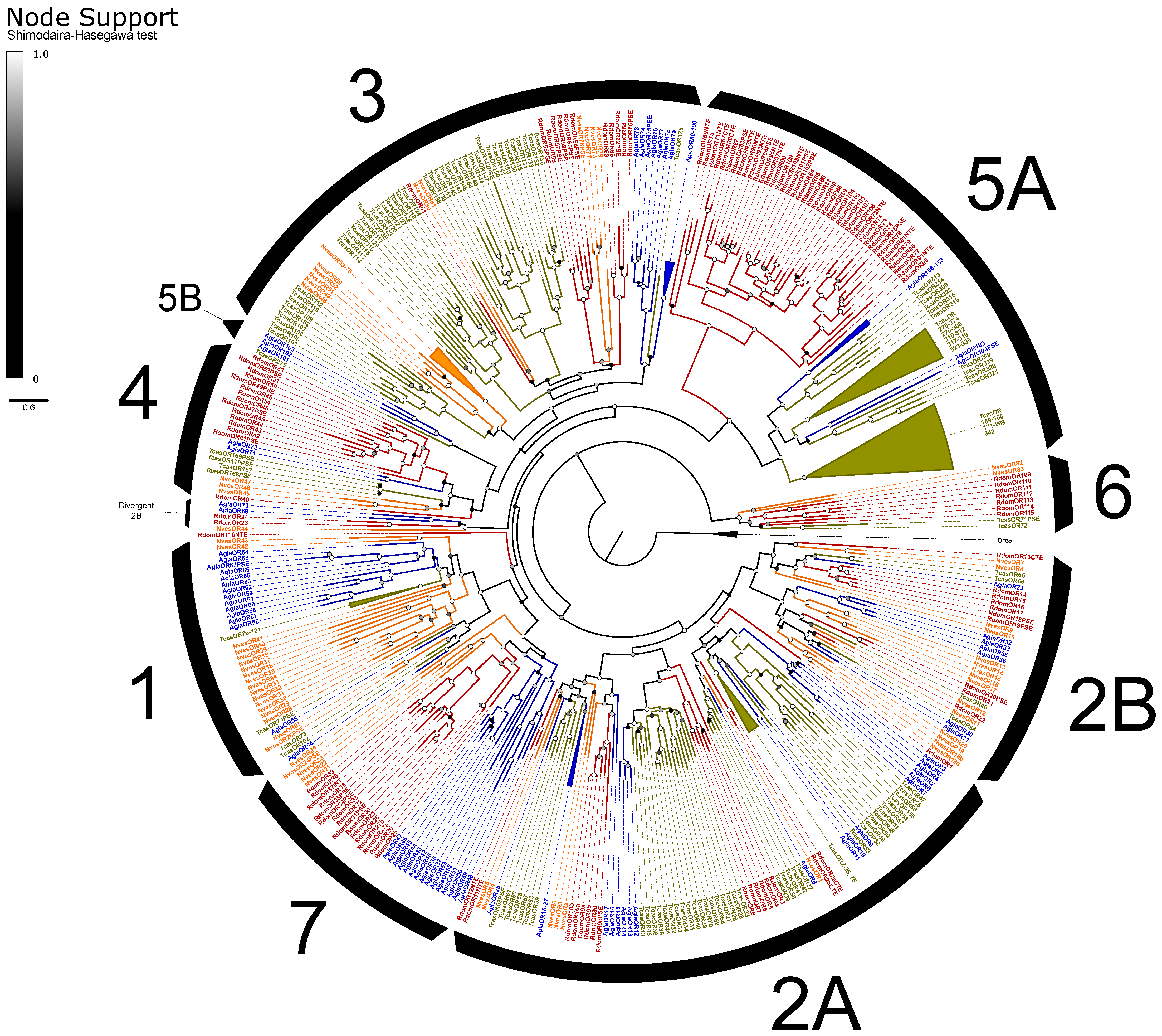
Odorant Receptors
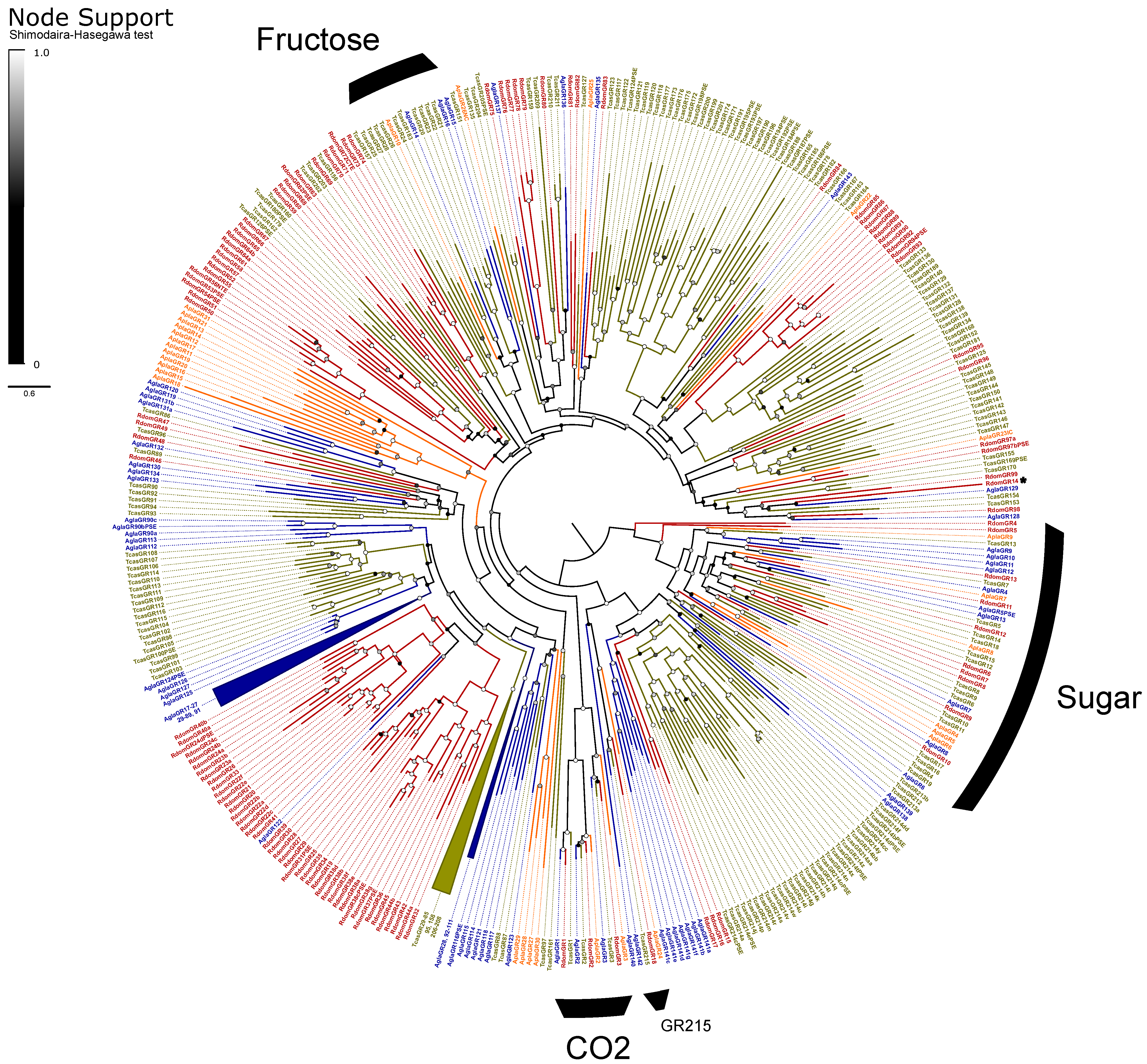
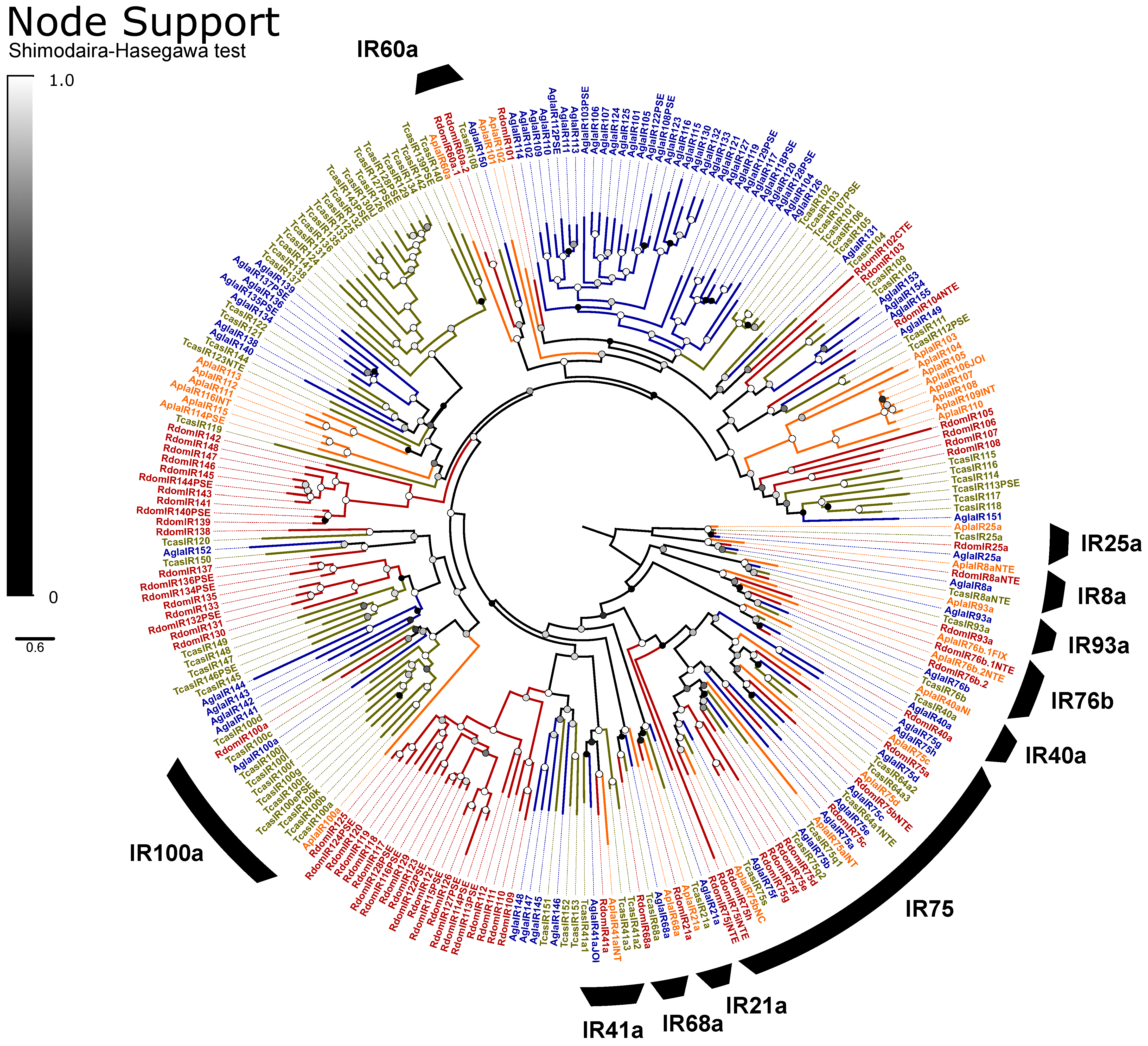
Gustatory and Ionotropic Receptors
3.2.3. Digestive Enzymes
Carbohydrases
Peptidases
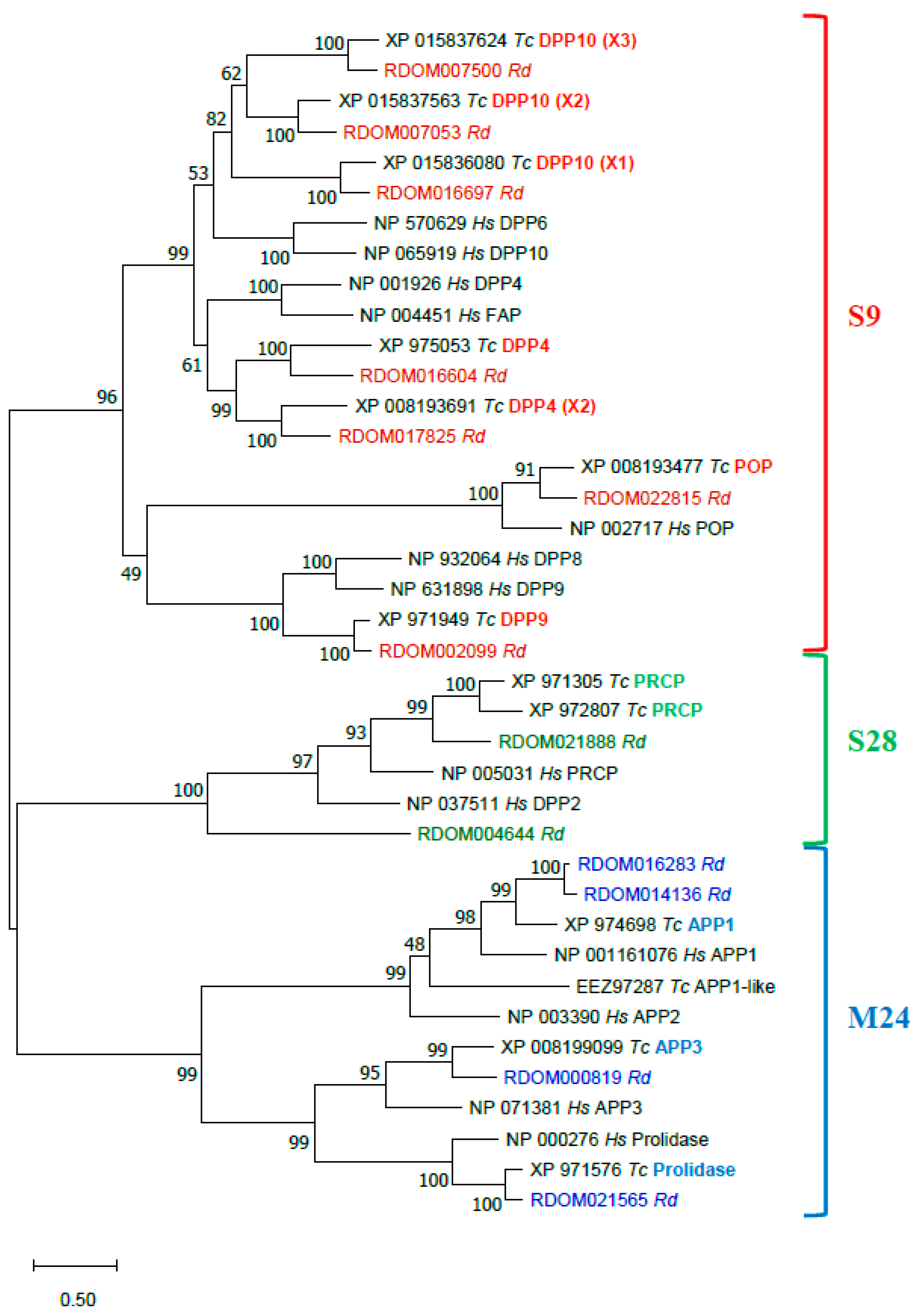
Proline-Specific Peptidases (PSPs)
Serine Peptidases
Metalloexopeptidases
3.2.4. Other Genes of Interest
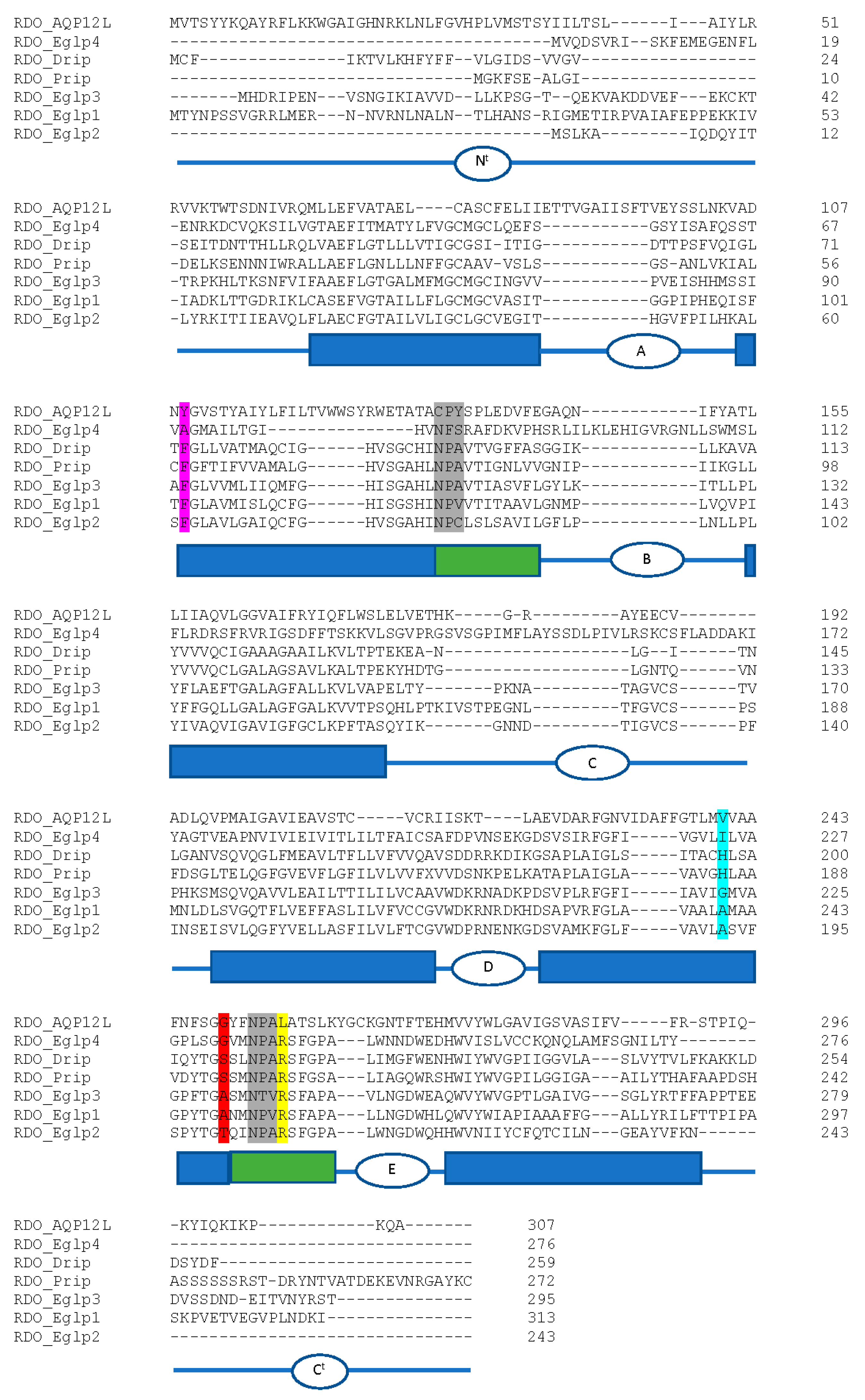
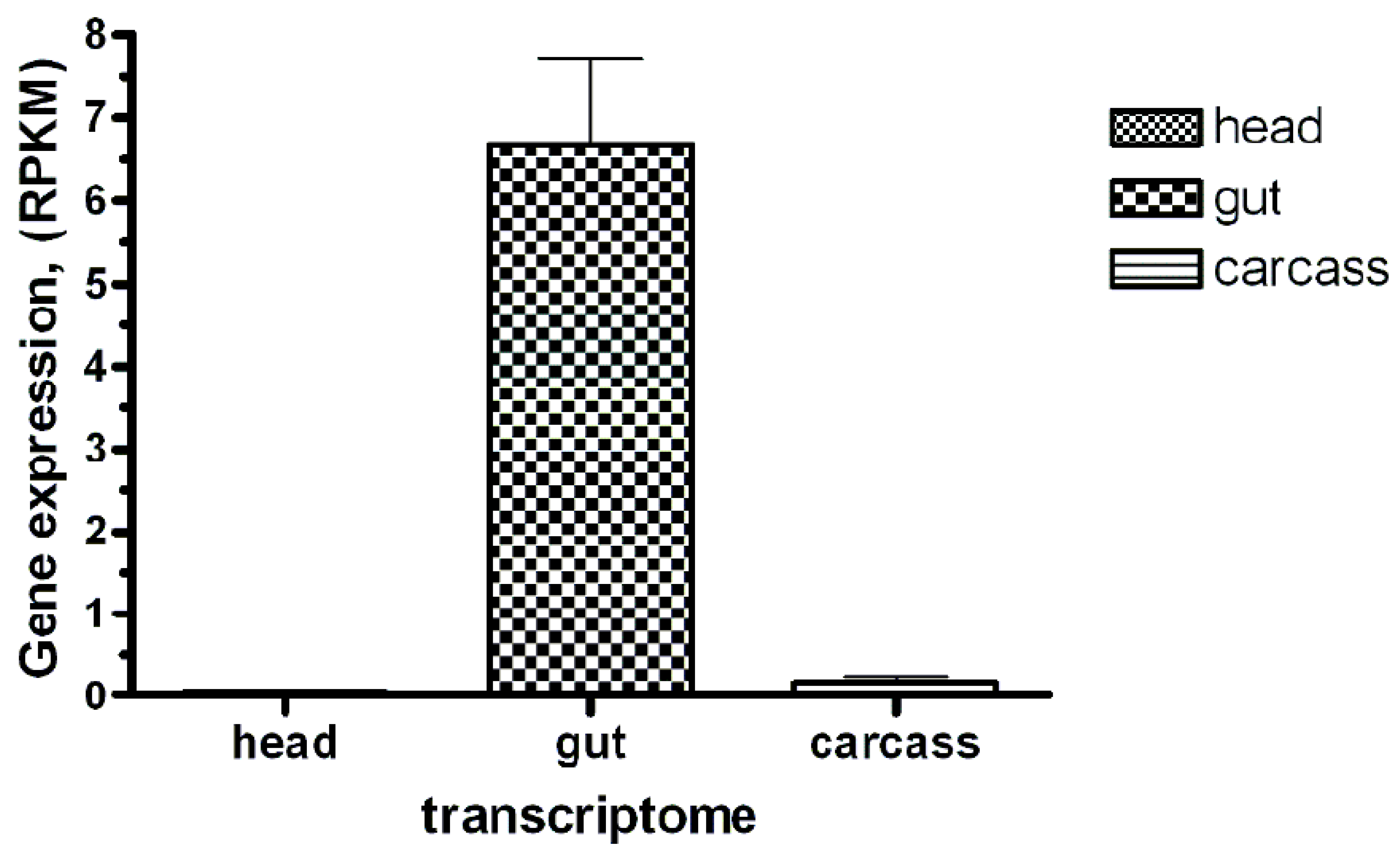
Aquaporins
RdCad1, a Putative Receptor of Insecticidal Cry Toxins
Cysteine Peptidases
Phosphine Resistance Genes
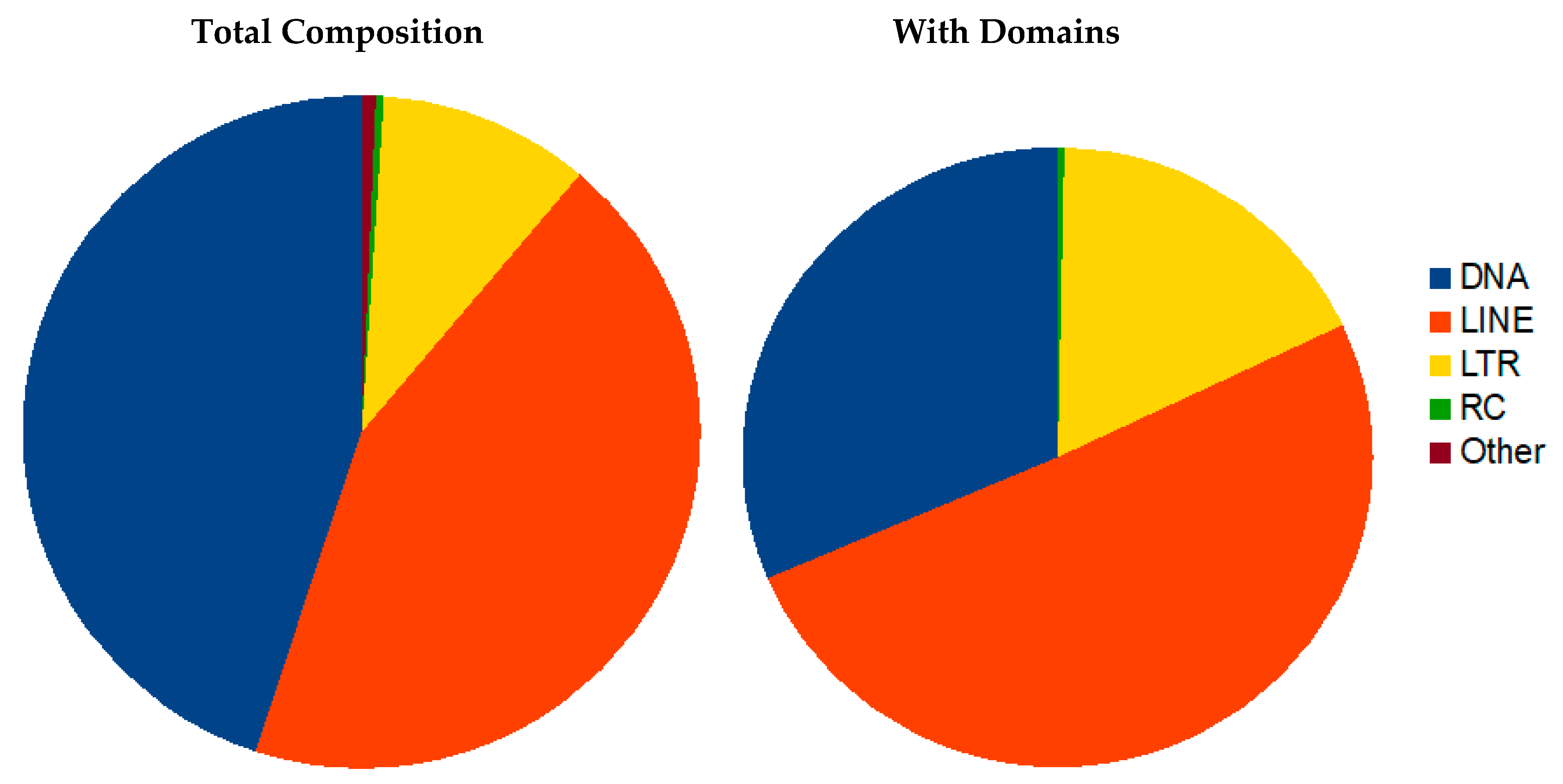
3.3. Repeat Sequence/Structure
3.3.1. Transposon Elements (TE)
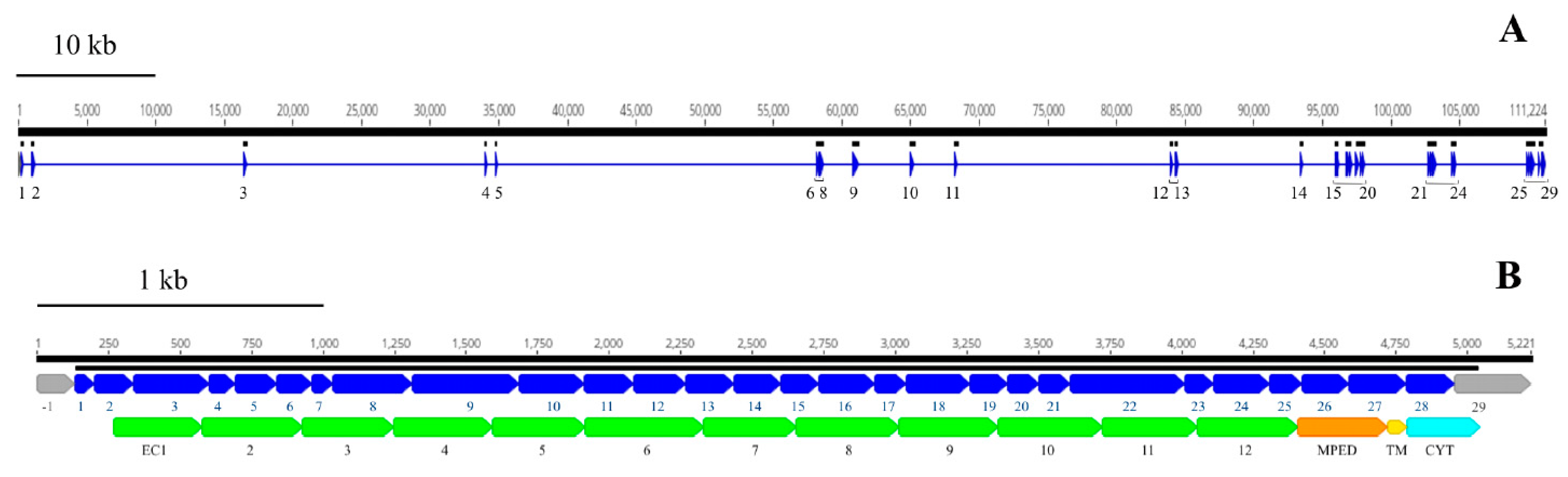
3.3.2. Identification and Characterization of Satellite DNAs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABC | ATP binding cassette |
| APP | aminopepdidase P |
| AQP | Aquaporin |
| Bib | Drosophila big-brain |
| Bt | Bacillus thuringiensis |
| BUSCO | Benchmarking Universal Single-Copy Orthologs |
| CGAHR | Center for Grain and Animal Health Research |
| COesterases | Carboxylesterases |
| CYT | Cytoplasmic |
| DLD | dihydrolipoamide dehydrogenase |
| DPP | dipeptdidylpeptidases |
| Drip | Drosophila integral protein |
| EC | Ectodomain |
| Eglps | entomoglyveroporins |
| FAP | fibroblast activation protein |
| GH | glucosyl hydrolase |
| GLPs | aquaglyceroporins |
| GR | gustatory receptor |
| INT | internal exons |
| IR | ionotropic receptor |
| LINE | long interspersed nuclear element |
| LTR | long terminal repeat; |
| MPED | membrane proximal extracellular domain |
| MRPs | multidrug resistance proteins |
| NBD | nucleotide binding domain |
| NPA motif | asparagine-proline-alanine motif |
| NTE | N-terminal |
| OR | odorant receptor |
| PCWDE | plant cell wall degrading enzymes |
| PSPs | proline-specific peptidases |
| PER | period to eliminate multiple reporting of repeats |
| POP | prolyloligopeptidase |
| PRCP | prolylcarboxypeptidase |
| Prip | Pyrocoelia rufa integral protein |
| RdCad1 | BtR1-like cadherin gene |
| RNAi | RNA interference |
| RPKM | reads per kilobase per million mapped reads |
| satDNA | satellite DNA |
| SP | single-domain serine peptidases |
| SPHs | non-catalytic SP homologs |
| TBR | toxin-binding regions |
| TE | transposable elements |
| TIR | terminal inverted repeats |
| TM/TMD | transmembrane/transmembrane domain |
| TRBD | tandem repeat database |
| UGTs | Uridine 5′-diphospho-glucuronosyltransferase |
References
- Lucas, H. Crustacés, arachnides, myriopodes et hexapodes: Exploration scientifique de l’Algérie pendant les années 1840, 1841, 1842. Sci. Phys. Zool. Hist. Nat. Animaux Articul. 1849, 1, 1–403. [Google Scholar]
- Fields, P.; Van Loon, J.; Dolinski, M.; Harris, J.; Burkholder, W. The distribution of Rhyzopertha dominica (F.) in Western Canada. Can. Entomol. 1993, 125, 317–328. [Google Scholar] [CrossRef]
- Edde, P. A review of the biology and control of Rhyzopertha dominica (F.) the lesser grain borer. J. Stored Prod. Res. 2012, 48, 1–18. [Google Scholar] [CrossRef]
- Toews, M.D.; Campbell, J.F.; Arthur, F.H.; Ramaswamy, S.B. Outdoor flight activity and immigration of Rhyzopertha dominica into seed wheat warehouses. Entomol. Exp. Appl. 2006, 121, 73–85. [Google Scholar] [CrossRef]
- Schwardt, H.H. Life history of the lesser grain borer. J. Kans. Entomol. Soc. 1993, 6, 61–66. [Google Scholar]
- Edde, P.A.; Phillips, T.W. Potential host affinities for the lesser grain borer, Rhyzopertha dominica: Behavioral responses to host odors and pheromones and reproductive ability on non-grain hosts. Entomol. Exp. Appl. 2006, 119, 255–263. [Google Scholar] [CrossRef]
- Wright, V.F.; Fleming, E.E.; Post, D.; Wright, F. Survival of Rhyzopertha dominica (Coleoptera, Bostrichidae ) on fruits and seeds collected from woodrat nests in Kansas. J. Kans. Entomol. Soc. 1990, 63, 344–347. [Google Scholar]
- Jia, F.; Toews, M.D.; Campbell, J.F.; Ramaswamy, S.B. Survival and reproduction of lesser grain borer, Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae) on flora associated with native habitats in Kansas. J. Stored Prod. Res. 2008, 44, 366–372. [Google Scholar] [CrossRef]
- Mahroof, R.M.; Edde, P.; Robertson, B.; Puckette, J.A.; Phillips, T.W. Dispersal of Rhyzopertha dominica (Coleoptera: Bostrichidae) in different habitats. Environ. Entomol. 2010, 39, 930–938. [Google Scholar] [CrossRef]
- Quellhorst, H.; Athanassiou, C.G.; Zhu, K.Y.; Morrison, W.R. The biology, ecology and management of the larger grain borer, Prostephanus truncatus (Horn) (Coleoptera: Bostrichidae). J. Stored Prod. Res. 2021, 94, 101860. [Google Scholar] [CrossRef]
- Opit, G.P.; Phillips, T.W.; Aikins, M.J.; Hasan, M.M. Phosphine resistance in Tribolium castaneum and Rhyzopertha dominica from stored wheat in Oklahoma. J. Econ. Entomol. 2012, 105, 1107–1114. [Google Scholar] [CrossRef] [PubMed]
- Lorini, I.; Collins, P.J.; Daglish, G.J.; Nayak, M.K.; Pavic, H. Detection and characterisation of strong resistance to phosphine in Brazilian Rhyzopertha dominica (F.) (Coleoptera: Bostrychidae). Pest Manag. Sci. 2007, 63, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Benhalima, H.; Chaudhry, M.; Mills, K.; Price, N. Phosphine resistance in stored-product insects collected from various grain storage facilities in Morocco. J. Stored Prod. Res. 2004, 40, 241–249. [Google Scholar] [CrossRef]
- Collins, P.J.; Daglish, G.; Bengston, M.; Lambkin, T.M.; Pavic, H. Genetics of resistance to phosphine in Rhyzopertha dominica (Coleoptera: Bostrichidae). J. Econ. Entomol. 2002, 95, 862–869. [Google Scholar] [CrossRef]
- Schlipalius, I.D.; Chen, W.; Collins, P.J.; Nguyen, T.; Reilly, P.E.B.; Ebert, P.R. Gene interactions constrain the course of evolution of phosphine resistance in the lesser grain borer, Rhyzopertha dominica. Heredity 2008, 100, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Schlipalius, D.I.; Valmas, N.; Tuck, A.G.; Jagadeesan, R.; Ma, L.; Kaur, R.; Goldinger, A.; Anderson, C.; Kuang, J.; Zuryn, S.; et al. A core metabolic enzyme mediates resistance to phosphine gas. Science 2012, 338, 807–810. [Google Scholar] [CrossRef]
- Arthur, F.H. Grain protectants: Current status and prospects for the future. J. Stored Prod. Res. 1996, 32, 293–302. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Athanassiou, C.G.; Arthur, F.H. Efficacy of deltamethrin against stored-product beetles at short exposure intervals or on a partially treated rice mass. J. Econ. Entomol. 2015, 108, 1416–1421. [Google Scholar] [CrossRef]
- Sehgal, B.; Subramanyam, B.; Arthur, F.H.; Gill, B.S. Variation in susceptibility of laboratory and field strains of three stored-grain insect species to β -cyfluthrin and chlorpyrifos-methyl plus deltamethrin applied to concrete surfaces. Pest Manag. Sci. 2014, 70, 576–587. [Google Scholar] [CrossRef]
- Arthur, F. Efficacy of methoprene for multi-year protection of stored wheat, brown rice, rough rice and corn. J. Stored Prod. Res. 2016, 68, 85–92. [Google Scholar] [CrossRef]
- Haliscak, J.P.; Beeman, R.W. Status of malathion resistance in five genera of beetles infesting farm-stored corn, wheat, and oats in the United States. J. Econ. Entomol. 1983, 76, 717–722. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Kambhampati, S.; Dover, B.A.; Zhu, K.Y. Biochemical mechanism of organophosphate resistance in Brazilian and U. S. populations of Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae). Bull. Entomol. Res. 1997, 87, 581–586. [Google Scholar] [CrossRef]
- Guedes, R.N.; Zhu, K.; Kambhampati, S.; Dover, B. Characterization of acetylcholinesterase purified from the lesser grain borer, Rhyzopertha dominica (Coleoptera: Bostrichidae). Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1998, 119, 205–210. [Google Scholar] [CrossRef]
- Wang, H.-T.; Tsai, C.-L.; Chen, M.-E. Nicotinic acetylcholine receptor subunit α6 associated with spinosad resistance in Rhyzopertha dominica (Coleoptera: Bostrichidae). Pestic. Biochem. Physiol. 2018, 148, 68–73. [Google Scholar] [CrossRef]
- Sakka, M.K.; Riga, M.; Ioannidis, P.; Baliota, G.V.; Tselika, M.; Jagadeesan, R.; Nayak, M.K.; Vontas, J.; Athanassiou, C.G. Transcriptomic analysis of s-methoprene resistance in the lesser grain borer, Rhyzopertha dominica, and evaluation of piperonyl butoxide as a resistance breaker. BMC Genom. 2021, 22, 65. [Google Scholar] [CrossRef]
- Khorramshahi, A.; Burkholder, W.E. Behavior of the lesser grain borer Rhyzopertha dominica (Coleoptera: Bostrichidae). J. Chem. Ecol. 1981, 7, 33–38. [Google Scholar] [CrossRef]
- Williams, H.J.; Silverstein, R.M.; Burkholder, W.E.; Khorramshahi, A. Dominicalure 1 and 2: Components of aggregation pheromone from male lesser grain borer Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae). J. Chem. Ecol. 1981, 7, 759–780. [Google Scholar] [CrossRef]
- Bashir, T.; Birkinshaw, L.; Hall, D.; Hodges, R. Host odours enhance the responses of adult Rhyzopertha dominica to male-produced aggregation pheromone. Entomol. Exp. Appl. 2001, 101, 273–280. [Google Scholar] [CrossRef]
- Nguyen, D.T.; Hodges, R.J.; Belmain, S.R. Do walking Rhyzopertha dominica (F.) locate cereal hosts by chance? J. Stored Prod. Res. 2008, 44, 90–99. [Google Scholar] [CrossRef]
- Edde, P.; Phillips, T.W.; Robertson, J.B.; Dillwith, J.W. Pheromone output by Rhyzopertha dominica (Coleoptera: Bostrichidae), as affected by host plant and beetle size. Ann. Entomol. Soc. Am. 2007, 100, 83–90. [Google Scholar] [CrossRef]
- Dowdy, A.K.; Howard, R.W.; Seitz, L.M.; McGaughey, W.H. Response of Rhyzopertha dominica (Coleoptera: Bostrichidae) to its aggregation pheromone and wheat volatiles. Environ. Entomol. 1993, 22, 965–970. [Google Scholar] [CrossRef]
- Cordeiro, E.M.G.; Campbell, J.F.; Phillips, T.W. Movement and orientation decision modeling of Rhyzopertha dominica (Coleoptera: Bostrichidae) in the grain mass. Environ. Entomol. 2016, 45, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Robertson, H.M. Molecular evolution of the major arthropod chemoreceptor gene families. Annu. Rev. Entomol. 2019, 64, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.F.; Andersson, M.N. Olfactory genomics of the Coleoptera. In Insect Pheromone Biochemistry and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 547–590. [Google Scholar]
- Andersson, M.N.; Keeling, C.I.; Mitchell, R.F. Genomic content of chemosensory genes correlates with host range in wood-boring beetles (Dendroctonus ponderosae, Agrilus planipennis, and Anoplophora glabripennis). BMC Genom. 2019, 20, 690. [Google Scholar] [CrossRef]
- Mitchell, R.F.; Hughes, D.T.; Luetje, C.W.; Millar, J.G.; Soriano-Agatón, F.; Hanks, L.M.; Robertson, H.M. Sequencing and characterizing odorant receptors of the cerambycid beetle Megacyllene caryae. Insect Biochem. Mol. Biol. 2012, 42, 499–505. [Google Scholar] [CrossRef]
- Antony, B.; Johny, J.; Montagné, N.; Jacquin-Joly, E.; Capoduro, R.; Cali, K.; Persaud, K.; Al-Saleh, M.A.; Pain, A. Pheromone receptor of the globally invasive quarantine pest of the palm tree, the red palm weevil (Rhynchophorus ferrugineus). Mol. Ecol. 2021, 30, 2025–2039. [Google Scholar] [CrossRef]
- Yuvaraj, J.K.; Roberts, R.E.; Sonntag, Y.; Hou, X.-Q.; Grosse-Wilde, E.; Machara, A.; Zhang, D.-D.; Hansson, B.S.; Johanson, U.; Löfstedt, C.; et al. Putative ligand binding sites of two functionally characterized bark beetle odorant receptors. BMC Biol. 2021, 19, 16. [Google Scholar] [CrossRef]
- Wang, X.; Wang, S.; Yi, J.; Li, Y.; Liu, J.; Wang, J.; Xi, J. Three host plant volatiles, hexanal, lauric acid, and tetradecane, are detected by an antenna-biased expressed odorant receptor 27 in the dark black chafer Holotrichia parallela. J. Agric. Food Chem. 2020, 68, 7316–7323. [Google Scholar] [CrossRef]
- Hou, X.Q.; Yuvaraj, J.K.; Roberts, R.E.; Zhang, D.D.; Unelius, C.R.; Löfstedt, C.; Andersson, M.N. Functional evolution of a bark beetle odorant receptor clade detecting monoterpenoids of different ecological origins. Mol. Biol. Evol. 2021, 38, 4934–4947. [Google Scholar] [CrossRef]
- Ji, T.; Xu, Z.; Jia, Q.; Wang, G.; Hou, Y. Non-palm plant volatile α-pinene is detected by antenna-biased expressed odorant receptor 6 in the Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae). Front. Physiol. 2021, 12, 701545. [Google Scholar] [CrossRef]
- Takada, T.; Sato, R.; Kikuta, S. A mannitol/sorbitol receptor stimulates dietary intake in Tribolium castaneum. PLoS ONE 2017, 12, e0186420. [Google Scholar] [CrossRef] [PubMed]
- Mandiana Diakite, M.; Wang, J.; Ali, S.; Wang, M. Identification of chemosensory gene families in Rhyzopertha dominica (Coleoptera: Bostrichidae). Can. Entomol. 2016, 148, 8–21. [Google Scholar] [CrossRef]
- Schlipalius, I.D.; Cheng, Q.; Reilly, P.E.B.; Collins, P.J.; Ebert, P.R. Genetic linkage analysis of the lesser grain borer Rhyzopertha dominica identifies two loci that confer high-level resistance to the fumigant phosphine. Genetics 2002, 161, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Johnston, J.S.; Bernardini, A.; Hjelmen, C.E. Genome size estimation and quantitative cytogenetics in insects. Program. Necrosis 2019, 1858, 15–26. [Google Scholar] [CrossRef]
- Putnam, N.H.; O’Connell, B.L.; Stites, J.C.; Rice, B.J.; Blanchette, M.; Calef, R.; Troll, C.J.; Fields, A.; Hartley, P.D.; Sugnet, C.W.; et al. Chromosome-scale shotgun assembly using an in vitro method for long-range linkage. Genome Res. 2016, 26, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed]
- Lieberman-Aiden, E.; Van Berkum, N.L.; Williams, L.; Imakaev, M.; Ragoczy, T.; Telling, A.; Amit, I.; Lajoie, B.R.; Sabo, P.J.; Dorschner, M.O.; et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science 2009, 326, 289–293. [Google Scholar] [CrossRef]
- Masumoto, H.; Masukata, H.; Muro, Y.; Nozaki, N.; Okazaki, T. A human centromere antigen (CENP-B) interacts with a short specific sequence in alphoid DNA, a human centromeric satellite. J. Cell Biol. 1989, 109, 1963–1973. [Google Scholar] [CrossRef]
- Johnson, A.D.; Handsaker, R.E.; Pulit, S.L.; Nizzari, M.M.; O’Donnell, C.J.; de Bakker, P.I.W. SNAP: A web-based tool for identification and annotation of proxy SNPs using HapMap. Bioinformatics 2008, 24, 2938–2939. [Google Scholar] [CrossRef]
- Oppert, B.; Morgan, T. Improved high-throughput bioassay for Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae). J. Stored Prod. Res. 2013, 52, 68–73. [Google Scholar] [CrossRef]
- Perkin, L.C.; Oppert, B. Gene expression in Tribolium castaneum life stages: Identifying a species-specific target for pest control applications. PeerJ 2019, 7, e6946. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.M.; Hubley, R.; Goubert, C.; Rosen, J.; Clark, A.G.; Feschotte, C.; Smit, A.F. RepeatModeler2 for automated genomic discovery of transposable element families. Proc. Natl. Acad. Sci. USA 2020, 117, 9451–9457. [Google Scholar] [CrossRef] [PubMed]
- Smit, A.F.A.; Hubley, R.; Green, P. RepeatMasker Open-4.0. Available online: http://www.repeatmasker.org (accessed on 5 January 2021).
- Stanke, M.; Steinkamp, R.; Waack, S.; Morgenstern, B. AUGUSTUS: A web server for gene finding in eukaryotes. Nucleic Acids Res. 2004, 32, W309–W312. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Cantarel, B.L.; Korf, I.; Robb, S.M.; Parra, G.; Ross, E.; Moore, B.; Holt, C.; Alvarado, A.S.; Yandell, M. MAKER: An easy-to-use annotation pipeline designed for emerging model organism genomes. Genome Res. 2007, 18, 188–196. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA genes in genomic sequences. In Springer Protocols Handbooks; Springer Science and Business Media: Cham, Switzerland, 2019; Volume 1962, pp. 1–14. [Google Scholar]
- Broehan, G.; Kroeger, T.; Lorenzen, M.; Merzendorfer, H. Functional analysis of the ATP-binding cassette (ABC) transporter gene family of Tribolium castaneum. BMC Genom. 2013, 14, 6. [Google Scholar] [CrossRef]
- Grubbs, N.; Haas, S.; Beeman, R.W.; Lorenzen, M.D. The ABCs of eye color in Tribolium castaneum: Orthologs of the Drosophila white, scarlet, and brown genes. Genetics 2015, 199, 749–759. [Google Scholar] [CrossRef]
- Lee, E.; Helt, A.G.; Reese, J.T.; Munoz-Torres, M.C.; Childers, C.P.; Buels, R.M.; Stein, L.; Holmes, I.H.; Elsik, C.G.; Lewis, E.S. Web Apollo: A web-based genomic annotation editing platform. Genome Biol. 2013, 14, R93. [Google Scholar] [CrossRef]
- Skinner, M.E.; Uzilov, A.V.; Stein, L.D.; Mungall, C.J.; Holmes, I.H. JBrowse: A next-generation genome browser. Genome Res. 2009, 19, 1630–1638. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.F.; Schneider, T.M.; Schwartz, A.M.; Andersson, M.N.; McKenna, D.D. The diversity and evolution of odorant receptors in beetles (Coleoptera). Insect Mol. Biol. 2020, 29, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J. Molecular Evolution, Phylogenetics and Epidemiology. FigTree v1.3.1. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 5 January 2021).
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Park, C.; Bennett, C.; Thornton, M.; Kim, D. Rapid and accurate alignment of nucleotide conversion sequencing reads with HISAT-3N. Genome Res. 2021, 31, 1290–1295. [Google Scholar] [CrossRef] [PubMed]
- Kovaka, S.; Zimin, A.V.; Pertea, G.M.; Razaghi, R.; Salzberg, S.L.; Pertea, M. Transcriptome assembly from long-read RNA-seq alignments with StringTie2. Genome Biol. 2019, 20, 278. [Google Scholar] [CrossRef]
- Cao, X.; Jiang, H. Integrated modeling of structural genes using MCuNovo. In Insect Genomics. Methods in Molecular Biology; Brown, S., Pfrender, M., Eds.; Humana Press: New York, NY, USA, 2019; Volume 1858, pp. 45–57. [Google Scholar] [CrossRef]
- Warburton, P.E.; Giordano, J.; Cheung, F.; Gelfand, Y.; Benson, G. Inverted repeat structure of the human genome: The X-chromosome contains a preponderance of large, highly homologous inverted repeats that contain testes genes. Genome Res. 2004, 14, 1861–1869. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef]
- Jurka, J.; Kapitonov, V.V.; Pavlicek, A.; Klonowski, P.; Kohany, O.; Walichiewicz, J. Repbase update, a database of eukaryotic repetitive elements. Cytogenet. Genome Res. 2005, 110, 462–467. [Google Scholar] [CrossRef]
- Gelfand, Y.; Rodriguez, A.; Benson, G. TRDB—The Tandem Repeats Database. Nucleic Acids Res. 2007, 35, D80–D87. [Google Scholar] [CrossRef]
- Smith, S.G.; Brower, J.H. Chromosome numbers of stored-product Coleoptera. J. Kans. Entomol. Soc. 1974, 47, 317–328. [Google Scholar]
- Guedes, R.N.C.; Dover, B.A.; Kambhampati, S. Resistance to chlorpyrifos-methyl, pirimiphos-methyl, and malathion in Brazilian and U.S. populations of Rhyzopertha dominica (Coleopera: Bostrichidae). J. Econ. Entomol. 1996, 89, 27–32. [Google Scholar] [CrossRef]
- Daglish, G.J.; Nayak, M.K. Prevalence of resistance to deltamethrin in Rhyzopertha dominica (F.) in eastern Australia. J. Stored Prod. Res. 2018, 78, 45–49. [Google Scholar] [CrossRef]
- Daglish, G.; Holloway, J.C.; Nayak, M.K. Implications of methoprene resistance for managing Rhyzopertha dominica (F.) in stored grain. J. Stored Prod. Res. 2013, 54, 8–12. [Google Scholar] [CrossRef]
- Adedipe, F.; Grubbs, N.; Coates, B.; Wiegmman, B.; Lorenzen, M. Structural and functional insights into the Diabrotica virgifera virgifera ATP-binding cassette transporter gene family. BMC Genom. 2019, 20, 899. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.D.; McKenna, D.; Scully, E.; Cook, S.C.; Dainat, B.; Egekwu, N.; Grubbs, N.; Lopez, D.; Lorenzen, M.; Reyna, S.M.; et al. Genome of the small hive beetle (Aethina tumida, Coleoptera: Nitidulidae), a worldwide parasite of social bee colonies, provides insights into detoxification and herbivory. GigaScience 2018, 7, 138. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.-J.; Niu, Y.-W.; Segraves, K.A.; Nie, R.-E.; Hao, Y.-J.; Zhang, L.-L.; Cheng, X.-C.; Zhang, X.-W.; Li, W.-Z.; Chen, R.-S.; et al. The draft genome of the specialist flea beetle Altica viridicyanea (Coleoptera: Chrysomelidae). BMC Genom. 2021, 22, 243. [Google Scholar] [CrossRef] [PubMed]
- Strauss, A.S.; Wang, D.; Stock, M.; Gretscher, R.; Groth, M.; Boland, W.; Burse, A. Tissue-specific transcript profiling for ABC transporters in the sequestering larvae of the phytophagous leaf beetle Chrysomela populi. PLoS ONE 2014, 9, e98637. [Google Scholar] [CrossRef]
- Dean, M.; Rzhetsky, A.; Allikmets, R. The human ATP-Binding Cassette (ABC) transporter superfamily. Genome Res. 2001, 11, 1156–1166. [Google Scholar] [CrossRef]
- David, J.-P.; Ismail, H.M.; Chandor-Proust, A.; Paine, M.J.I. Role of cytochrome P450s in insecticide resistance: Impact on the control of mosquito-borne diseases and use of insecticides on Earth. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120429. [Google Scholar] [CrossRef]
- Zhu, F.; Moural, T.W.; Shah, K.; Palli, S.R. Integrated analysis of cytochrome P450 gene superfamily in the red flour beetle, Tribolium castaneum. BMC Genom. 2013, 14, 174. [Google Scholar] [CrossRef]
- Jackson, C.J.; Liu, J.-W.; Carr, P.D.; Younus, F.; Coppin, C.; Meirelles, T.; Lethier, M.; Pandey, G.; Ollis, D.L.; Russell, R.J.; et al. Structure and function of an insect -carboxylesterase (Esterase7) associated with insecticide resistance. Proc. Natl. Acad. Sci. USA 2013, 110, 10177–10182. [Google Scholar] [CrossRef] [PubMed]
- Rane, R.; Walsh, T.; Pearce, S.L.; Jermiin, L.; Gordon, K.; Richards, S.; Oakeshott, J.G. Are feeding preferences and insecticide resistance associated with the size of detoxifying enzyme families in insect herbivores? Curr. Opin. Insect Sci. 2016, 13, 70–76. [Google Scholar] [CrossRef] [PubMed]
- McKenna, D.D.; Scully, E.D.; Pauchet, Y.; Hoover, K.; Kirsch, R.; Geib, S.M.; Mitchell, R.F.; Waterhouse, R.M.; Ahn, S.-J.; Arsala, D.; et al. Genome of the Asian longhorned beetle (Anoplophora glabripennis), a globally significant invasive species, reveals key functional and evolutionary innovations at the beetle–plant interface. Genome Biol. 2016, 17, 227. [Google Scholar] [CrossRef] [PubMed]
- Kaplanoglu, E.; Chapman, P.; Scott, I.; Donly, C. Overexpression of a cytochrome P450 and a UDP-glycosyltransferase is associated with imidacloprid resistance in the Colorado potato beetle, Leptinotarsa decemlineata. Sci. Rep. 2017, 7, 1762. [Google Scholar] [CrossRef]
- Scully, E.D.; Geib, S.M.; Carlson, E.J.; Tien, M.; McKenna, D.; Hoover, K. Functional genomics and microbiome profiling of the Asian longhorned beetle (Anoplophora glabripennis) reveal insights into the digestive physiology and nutritional ecology of wood feeding beetles. BMC Genom. 2014, 15, 1096. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, M.; Wang, W.; Sun, H.; Xu, Y.; Ma, L.; Sun, Y.; Zhu, C. prag01, a novel deltamethrin-resistance-associated gene from Culex pipiens pallens. Parasitol. Res. 2011, 108, 417–423. [Google Scholar] [CrossRef]
- Vosshall, L.B.; Hansson, B.S. A unified nomenclature system for the insect olfactory coreceptor. Chem. Senses 2011, 36, 497–498. [Google Scholar] [CrossRef]
- Dippel, S.; Kollmann, M.; Oberhofer, G.; Montino, A.; Knoll, C.; Krala, M.; Rexer, K.-H.; Frank, S.; Kumpf, R.; Schachtner, J.; et al. Morphological and transcriptomic analysis of a beetle chemosensory system reveals a gnathal olfactory center. BMC Biol. 2016, 14, 90. [Google Scholar] [CrossRef]
- Croset, V.; Rytz, R.; Cummins, S.F.; Budd, A.; Brawand, D.; Kaessmann, H.; Gibson, T.J.; Benton, R. Ancient protostome origin of chemosensory ionotropic glutamate receptors and the evolution of insect taste and olfaction. PLoS Genet. 2010, 6, e1001064. [Google Scholar] [CrossRef]
- Rytz, R.; Croset, V.; Benton, R. Ionotropic Receptors (IRs): Chemosensory ionotropic glutamate receptors in Drosophila and beyond. Insect Biochem. Mol. Biol. 2013, 43, 888–897. [Google Scholar] [CrossRef]
- Sánchez-Alcañiz, J.A.; Silbering, A.F.; Croset, V.; Zappia, G.; Sivasubramaniam, A.K.; Abuin, L.; Sahai, S.Y.; Münch, D.; Steck, K.; Auer, T.O.; et al. An expression atlas of variant ionotropic glutamate receptors identifies a molecular basis of carbonation sensing. Nat. Commun. 2018, 9, 4252. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Zhang, M.; Üçpunar, H.; Svensson, T.; Quillery, E.; Gompel, N.; Ignell, R.; Kadow, I.C.G. Ionotropic chemosensory receptors mediate the taste and smell of polyamines. PLoS Biol. 2016, 14, e1002454. [Google Scholar] [CrossRef] [PubMed]
- Knecht, Z.A.; Silbering, A.F.; Cruz, J.; Yang, L.; Croset, V.; Benton, R.; Garrity, P.A. Ionotropic receptor-dependent moist and dry cells control hygrosensation in Drosophila. eLife 2017, 6, e26654. [Google Scholar] [CrossRef] [PubMed]
- Pauchet, Y.; Wilkinson, P.; Chauhan, R.; Ffrench-Constant, R. Diversity of beetle genes encoding novel plant cell wall degrading enzymes. PLoS ONE 2010, 5, e15635. [Google Scholar] [CrossRef] [PubMed]
- Consortium, T.G.S.; Richards, S.; Gibbs, R.A.; Weinstock, G.M.; Brown, S.J.; Denell, R. The genome of the model beetle and pest Tribolium castaneum. Nature 2008, 452, 949–955. [Google Scholar] [CrossRef]
- Da Lage, J.-L. The amylases of insects. Int. J. Insect Sci. 2018, 10, 1179543318804783. [Google Scholar] [CrossRef]
- Da Costa-Latgé, S.G.; Bates, P.; Dillon, R.; Genta, F.A. Characterization of glycoside hydrolase families 13 and 31 reveals expansion and diversification of α-amylase genes in the phlebotomine Lutzomyia longipalpis and modulation of sandfly glycosidase activities by leishmania infection. Front. Physiol. 2021, 12, 635633. [Google Scholar] [CrossRef]
- Keeling, C.I.; Henderson, H.; Li, M.; Yuen, M.; Clark, E.L.; Fraser, J.D.; Huber, D.P.; Liao, N.Y.; Docking, T.R.; Birol, I.; et al. Transcriptome and full-length cDNA resources for the mountain pine beetle, Dendroctonus ponderosae Hopkins, a major insect pest of pine forests. Insect Biochem. Mol. Biol. 2012, 42, 525–536. [Google Scholar] [CrossRef]
- Sicker, D.; Frey, M.; Schulz, M.; Gierl, A. Role of natural benzoxazinones in the survival strategy of plants. Int. Rev. Cytol. 2000, 198, 319–346. [Google Scholar] [CrossRef]
- Hanhineva, K.; Rogachev, I.; Aura, A.-M.; Aharoni, A.; Poutanen, K.; Mykkänen, H. Qualitative characterization of benzoxazinoid derivatives in whole grain rye and wheat by LC-MS metabolite profiling. J. Agric. Food Chem. 2011, 59, 921–927. [Google Scholar] [CrossRef]
- Robert, C.A.; Zhang, X.; Machado, R.A.; Schirmer, S.; Lori, M.; Mateo, P.; Erb, M.; Gershenzon, J. Sequestration and activation of plant toxins protect the western corn rootworm from enemies at multiple trophic levels. eLife 2017, 6, e29307. [Google Scholar] [CrossRef] [PubMed]
- Davison, A.; Blaxter, M. Ancient origin of glycosyl hydrolase family 9 cellulase genes. Mol. Biol. Evol. 2005, 22, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Lo, N.; Tokuda, G.; Watanabe, H.; Rose, H.; Slaytor, M.; Maekawa, K.; Bandi, C.; Noda, H. Evidence from multiple gene sequences indicates that termites evolved from wood-feeding cockroaches. Curr. Biol. 2000, 10, 801–804. [Google Scholar] [CrossRef]
- Zhou, X.; Kovaleva, E.S.; Wu-Scharf, D.; Campbell, J.H.; Buchman, G.W.; Boucias, D.G.; Scharf, M.E. Production and characterization of a recombinant beta-1,4-endoglucanase (glycohydrolase family 9) from the termite Reticulitermes flavipes. Arch. Insect Biochem. Physiol. 2010, 74, 147–162. [Google Scholar] [CrossRef]
- Shelomi, M.; Watanabe, H.; Arakawa, G. Endogenous cellulase enzymes in the stick insect (Phasmatodea) gut. J. Insect Physiol. 2014, 60, 25–30. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef]
- Cunningham, D.F.; O’Connor, B. Proline specific peptidases. Biochim. Biophys. Acta BBA—Protein Struct. Mol. Enzym. 1997, 1343, 160–186. [Google Scholar] [CrossRef]
- Dunaevsky, Y.E.; Tereshchenkova, V.F.; Oppert, B.; Belozersky, M.A.; Filippova, I.Y.; Elpidina, E.N. Human proline specific peptidases: A comprehensive analysis. Biochim. Biophys. Acta BBA—Gen. Subj. 2020, 1864, 129636. [Google Scholar] [CrossRef]
- Shewry, P.R.; Halford, N.G. Cereal seed storage proteins: Structures, properties and role in grain utilization. J. Exp. Bot. 2002, 53, 947–958. [Google Scholar] [CrossRef]
- Shewry, P.R.; Tatham, A.S. The prolamin storage proteins of cereal seeds: Structure and evolution. Biochem. J. 1990, 267, 1–12. [Google Scholar] [CrossRef]
- Goptar, I.A.; Filippova, I.Y.; Lysogorskaya, E.N.; Oksenoit, E.S.; Vinokurov, K.S.; Zhuzhikov, D.P.; Bulushova, N.V.; Zalunin, I.A.; Dunaevsky, Y.E.; Belozersky, M.A.; et al. Localization of post-proline cleaving peptidases in Tenebrio molitor larval midgut. Biochimie 2008, 90, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Kakimoto, T.; Oshima, G.; Yeh, H.; Erdös, E. Purification of lysosomal prolylcarboxypeptidase angiotensinase C. Biochim. Biophys. Acta BBA—Enzym. 1973, 302, 178–182. [Google Scholar] [CrossRef]
- Odya, C.; Marinkovic, D.; Hammon, K.; Stewart, T.; Erdös, E. Purification and properties of prolylcarboxypeptidase (angiotensinase C) from human kidney. J. Biol. Chem. 1978, 253, 5927–5931. [Google Scholar] [CrossRef]
- Tan, F.; Morris, P.; Skidgel, R.; Erdös, E. Sequencing and cloning of human prolylcarboxypeptidase (angiotensinase C). Similarity to both serine carboxypeptidase and prolylendopeptidase families. J. Biol. Chem. 1993, 268, 16631–16638. [Google Scholar] [CrossRef]
- Tereshchenkova, V.F.; Goptar, I.A.; Kulemzina, I.A.; Zhuzhikov, D.P.; Serebryakova, M.; Belozersky, M.A.; Dunaevsky, Y.E.; Oppert, B.; Filippova, I.Y.; Elpidina, E.N. Dipeptidyl peptidase 4—An important digestive peptidase in Tenebrio molitor larvae. Insect Biochem. Mol. Biol. 2016, 76, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Di Cera, E. Serine proteases. IUBMB Life 2009, 61, 510–515. [Google Scholar] [CrossRef]
- Srinivasan, A.; Giri, A.P.; Gupta, V.S. Structural and functional diversities in lepidopteran serine proteases. Cell. Mol. Biol. Lett. 2006, 11, 132–154. [Google Scholar] [CrossRef]
- Konarev, A.V.; Fomicheva, Y.V. Cross analysis of the interaction of alpha-amylase and proteinase components of insects with protein inhibitors from wheat endosperm. Biokhimiya 1991, 56, 628–638. [Google Scholar]
- Zhu, Y.-C.; Baker, J.E. Characterization of midgut trypsin-like enzymes and three trypsinogen cDNAs from the lesser grain borer, Rhyzopertha dominica (Coleoptera: Bostrichidae). Insect Biochem. Mol. Biol. 1999, 29, 1053–1063. [Google Scholar] [CrossRef]
- Zhu, Y.-C.; Baker, J.E. Molecular cloning and characterization of a midgut chymotrypsin-like enzyme from the lesser grain borer, Rhyzopertha dominica. Arch. Insect Biochem. Physiol. 2000, 43, 173–184. [Google Scholar] [CrossRef]
- Miao, Z.; Cao, X.; Jiang, H. Digestion-related proteins in the tobacco hornworm, Manduca sexta. Insect Biochem. Mol. Biol. 2020, 126, 103457. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Jiang, H. Building a platform for predicting functions of serine protease-related proteins in Drosophila melanogaster and other insects. Insect Biochem. Mol. Biol. 2018, 103, 53–69. [Google Scholar] [CrossRef]
- Agre, P. Aquaporin water channels. Biosci. Rep. 2004, 24, 127–163. [Google Scholar] [CrossRef]
- Verkman, A. Aquaporins. Curr. Biol. 2013, 23, R52–R55. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.N.; Cerdà, J. Evolution and functional diversity of aquaporins. Biol. Bull. 2015, 229, 6–23. [Google Scholar] [CrossRef]
- Fu, D.; Libson, A.; Miercke, L.J.W.; Weitzman, C.; Nollert, P.; Krucinski, J.; Stroud, R.M. Structure of a glycerol-conducting channel and the basis for its selectivity. Science 2000, 290, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Sui, H.; Han, B.-G.; Lee, J.K.; Walian, P.J.; Jap, B.K. Structural basis of water-specific transport through the AQP1 water channel. Nature 2001, 414, 872–878. [Google Scholar] [CrossRef]
- Beitz, E.; Wu, B.; Holm, L.M.; Schultz, J.E.; Zeuthen, T. Point mutations in the aromatic/arginine region in aquaporin 1 allow passage of urea, glycerol, ammonia, and protons. Proc. Natl. Acad. Sci. USA 2006, 103, 269–274. [Google Scholar] [CrossRef]
- Fu, D.; Lu, M. The structural basis of water permeation and proton exclusion in aquaporins (review). Mol. Membr. Biol. 2007, 24, 366–374. [Google Scholar] [CrossRef]
- Almasalmeh, A.; Krenc, D.; Wu, B.; Beitz, E. Structural determinants of the hydrogen peroxide permeability of aquaporins. FEBS J. 2014, 281, 647–656. [Google Scholar] [CrossRef]
- Finn, R.N.; Chauvigné, F.; Stavang, J.A.; Belles, X.; Cerdà, J. Insect glycerol transporters evolved by functional co-option and gene replacement. Nat. Commun. 2015, 6, 7814. [Google Scholar] [CrossRef]
- Stavang, J.A.; Chauvigné, F.; Kongshaug, H.; Cerdà, J.; Nilsen, F.; Finn, R.N. Phylogenomic and functional analyses of salmon lice aquaporins uncover the molecular diversity of the superfamily in Arthropoda. BMC Genom. 2015, 16, 618. [Google Scholar] [CrossRef] [PubMed]
- Morishita, Y.; Matsuzaki, T.; Hara-Chikuma, M.; Andoo, A.; Shimono, M.; Matsuki, A.; Kobayashi, K.; Ikeda, M.; Yamamoto, T.; Verkman, A.; et al. Disruption of aquaporin-11 produces polycystic kidneys following vacuolization of the proximal tubule. Mol. Cell. Biol. 2005, 25, 7770–7779. [Google Scholar] [CrossRef] [PubMed]
- Gorelick, A.D.; Praetorius, J.; Tsunenari, T.; Nielsen, S.; Agre, P. Aquaporin-11: A channel protein lacking apparent transport function expressed in brain. BMC Biochem. 2006, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Drake, L.L.; Boudko, D.Y.; Marinotti, O.; Carpenter, V.K.; Dawe, A.L.; Hansen, I.A. The aquaporin gene family of the yellow fever mosquito, Aedes aegypti. PLoS ONE 2010, 5, e15578. [Google Scholar] [CrossRef]
- Fabrick, J.A.; Pei, J.; Hull, J.J.; Yool, A.J. Molecular and functional characterization of multiple aquaporin water channel proteins from the western tarnished plant bug, Lygus hesperus. Insect Biochem. Mol. Biol. 2014, 45, 125–140. [Google Scholar] [CrossRef]
- Van Ekert, E.; Chauvigné, F.; Finn, R.N.; Mathew, L.G.; Hull, J.J.; Cerdà, J.; Fabrick, J.A. Molecular and functional characterization of Bemisia tabaci aquaporins reveals the water channel diversity of hemipteran insects. Insect Biochem. Mol. Biol. 2016, 77, 39–51. [Google Scholar] [CrossRef]
- Yao, X.-X.; Meng, Q.-W.; Li, G.-Q. RNA interference-mediated functional characterization of aquaporin genes in Tribolium castaneum. Insect Mol. Biol. 2018, 27, 234–246. [Google Scholar] [CrossRef]
- Midboe, E.G.; Candas, M.; Bulla, L.A. Expression of a midgut-specific cadherin BT-R1 during the development of Manduca sexta larva. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2003, 135, 125–137. [Google Scholar] [CrossRef]
- Fabrick, J.A.; Wu, Y.; Wu, Y. Roles of insect midgut cadherin in Bt intoxication and resistance. In Bt Resistance: Characterization and Strategies for GM Crops Producing Bacillus Thuringiensis Toxins; Soberón, M., Gao, A., Bravo, A., Eds.; CABI Crop Protection Compendium: Boston, MA, USA, 2015; Volume 4, pp. 69–86. [Google Scholar]
- Adang, M.J.; Crickmore, N.; Jurat-Fuentes, J.L. Diversity of Bacillus thuringiensis crystal toxins and mechanism of action. Adv. Insect Physiol. 2014, 47, 39–87. [Google Scholar] [CrossRef]
- Dorsch, J.; Candas, M.; Griko, N.; Maaty, W.; Midboe, E.; Vadlamudi, R.; Bulla, L. Cry1A toxins of Bacillus thuringiensis bind specifically to a region adjacent to the membrane-proximal extracellular domain of BT-R1 in Manduca sexta: Involvement of a cadherin in the entomopathogenicity of Bacillus thuringiensis. Insect Biochem. Mol. Biol. 2002, 32, 1025–1036. [Google Scholar] [CrossRef]
- Hua, G.; Jurat-Fuentes, J.L.; Adang, M.J. Bt-R1a extracellular cadherin repeat 12 mediates Bacillus thuringiensis Cry1Ab binding and cytotoxicity. J. Biol. Chem. 2004, 279, 28051–28056. [Google Scholar] [CrossRef] [PubMed]
- Gómez, I.; Lopez, L.P.; Muñoz-Garay, C.; Fernandez, L.; Pérez, C.; Sánchez, J.; Soberón, M.; Bravo, A. Role of receptor interaction in the mode of action of insecticidal Cry and Cyt toxins produced by Bacillus thuringiensis. Peptides 2007, 28, 169–173. [Google Scholar] [CrossRef]
- Sayed, A.; Nekl, E.R.; Siqueira, H.; Wang, H.-C.; Ffrench-Constant, R.; Bagley, M.; Siegfried, B.D. A novel cadherin-like gene from western corn rootworm, Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae), larval midgut tissue. Insect Mol. Biol. 2007, 16, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Fabrick, J.; Oppert, C.; Lorenzen, M.D.; Morris, K.; Oppert, B.; Jurat-Fuentes, J.L. A novel tenebrio molitor cadherin is a functional receptor for Bacillus thuringiensis Cry3Aa toxin. J. Biol. Chem. 2009, 284, 18401–18410. [Google Scholar] [CrossRef]
- Hua, G.; Park, Y.; Adang, M.J. Cadherin AdCad1 in Alphitobius diaperinus larvae is a receptor of Cry3Bb toxin from Bacillus thuringiensis. Insect Biochem. Mol. Biol. 2014, 45, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Bel, Y.; Escriche, B. Common genomic structure for the Lepidoptera cadherin-like genes. Gene 2006, 381, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.; Armisén, D.; Gibbs, R.A.; Hering, L.; Khila, A.; Mayer, G.; Richards, S.; Niehuis, O.; Misof, B. Diversity and evolution of the transposable element repertoire in arthropods with particular reference to insects. BMC Ecol. Evol. 2019, 19, 1–15. [Google Scholar] [CrossRef]
- Gómez, I.; Oltean, D.I.; Gill, S.S.; Bravo, A.; Soberón, M. Mapping the epitope in cadherin-like receptors involved in Bacillus thuringiensis Cry1A toxin interaction using phage display. J. Biol. Chem. 2001, 276, 28906–28912. [Google Scholar] [CrossRef]
- Whalon, M.E.; Miller, D.L.; Hollingworth, R.M.; Grafius, E.J.; Miller, J.R. Selection of a Colorado potato beetle (Coleoptera: Chrysomelidae) strain resistant to Bacillus thuringiensis. J. Econ. Entomol. 1993, 86, 226–233. [Google Scholar] [CrossRef]
- Oppert, B.; Morgan, T.D.; Kramer, K.J. Efficacy of Bacillus thuringiensis Cry3Aa protoxin and protease inhibitors against coleopteran storage pests. Pest Manag. Sci. 2011, 67, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Turk, V.; Stoka, V.; Vasiljeva, O.; Renko, M.; Sun, T.; Turk, B.; Turk, D. Cysteine cathepsins: From structure, function and regulation to new frontiers. Biochim. Biophys. Acta BBA—Proteins Proteom. 2012, 1824, 68–88. [Google Scholar] [CrossRef] [PubMed]
- Terra, W.R.; Ferreira, C. Insect digestive enzymes: Properties, compartmentalization and function. Comp. Biochem. Physiol. Part B Comp. Biochem. 1994, 109, 1–62. [Google Scholar] [CrossRef]
- Wolfson, J.L.; Murdock, L.L. Diversity in digestive proteinase activity among insects. J. Chem. Ecol. 1990, 16, 1089–1102. [Google Scholar] [CrossRef]
- Terra, W.R.; Cristofoletti, P. Midgut proteinases in three divergent species of Coleoptera. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1996, 113, 725–730. [Google Scholar] [CrossRef]
- Goptar, I.; Semashko, T.; Danilenko, S.; Lysogorskaya, E.; Oksenoit, E.; Zhuzhikov, D.; Belozersky, M.; Dunaevsky, Y.; Oppert, B.; Filippova, I.; et al. Cysteine digestive peptidases function as post-glutamine cleaving enzymes in tenebrionid stored-product pests. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2012, 161, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Martynov, A.G.; Elpidina, E.N.; Perkin, L.; Oppert, B. Functional analysis of C1 family cysteine peptidases in the larval gut of Tenebrio molitor and Tribolium castaneum. BMC Genom. 2015, 16, 75. [Google Scholar] [CrossRef]
- Perkin, L.; Elpidina, E.N.; Oppert, B. Expression patterns of cysteine peptidase genes across the Tribolium castaneum life cycle provide clues to biological function. PeerJ 2016, 4, e1581. [Google Scholar] [CrossRef]
- Oppert, B.; Elpidina, E.N.; Toutges, M.; Mazumdar-Leighton, S. Microarray analysis reveals strategies of Tribolium castaneum larvae to compensate for cysteine and serine protease inhibitors. Comp. Biochem. Physiol. Part D Genom. Proteom. 2010, 5, 280–287. [Google Scholar] [CrossRef]
- Novinec, M.; Lenarčič, B. Papain-like peptidases: Structure, function, and evolution. Biomol. Concepts 2013, 4, 287–308. [Google Scholar] [CrossRef]
- Wex, T.; Lipyansky, A.; Brömme, N.C.; Wex, H.; Guan, X.Q.; Brömme, D. TIN-ag-RP, a novel catalytically inactive cathepsin B-related protein with EGF domains, is predominantly expressed in vascular smooth muscle cells. Biochemistry 2001, 40, 1350–1357. [Google Scholar] [CrossRef] [PubMed]
- Schlipalius, D.; Tuck, A.G.; Jagadeesan, R.; Nguyen, T.; Kaur, R.; Subramanian, S.; Barrero, R.; Nayak, M.; Ebert, P.R. Variant linkage analysis using de novo transcriptome sequencing identifies a conserved phosphine resistance gene in insects. Genetics 2018, 209, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Chakrabarty, S.; Jin, M.; Liu, K.; Xiao, Y. Insect ATP-Binding Cassette (ABC) transporters: Roles in xenobiotic detoxification and Bt insecticidal activity. Int. J. Mol. Sci. 2019, 20, 2829. [Google Scholar] [CrossRef] [PubMed]
- Montella, I.R.; Schama, R.; Valle, D. The classification of esterases: An important gene family involved in insecticide resistance—A review. Memórias Inst. Oswaldo Cruz 2012, 107, 437–449. [Google Scholar] [CrossRef]
- Ahn, S.-J.; Vogel, H.; Heckel, D.G. Comparative analysis of the UDP-glycosyltransferase multigene family in insects. Insect Biochem. Mol. Biol. 2012, 42, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Latief, M. A family of chemoreceptors in Tribolium castaneum (Tenebrionidae: Coleoptera). PLoS ONE 2007, 19, e1319. [Google Scholar] [CrossRef] [PubMed]
- Engsontia, P.; Sangket, U.; Robertson, H.M.; Satasook, C. Diversification of the ant odorant receptor gene family and positive selection on candidate cuticular hydrocarbon receptors. BMC Res. Notes 2015, 8, 380. [Google Scholar] [CrossRef]
- Biebl, S.; Querner, P. Transportation of wood boring beetles in wooden transport boxes, wooden pallets, and newly bought wood in museums. Stud. Conserv. 2021, 66, 44–50. [Google Scholar] [CrossRef]
- Six, D.L. A major symbiont shift supports a major niche shift in a clade of tree-killing bark beetles. Ecol. Entomol. 2020, 45, 190–201. [Google Scholar] [CrossRef]
- Konarev, A.; Dolgikh, V.; Senderskiy, I.; Konarev, A.; Kapustkina, A.; Lovegrove, A. Characterisation of proteolytic enzymes of Eurygaster integriceps Put. (Sunn bug), a major pest of cereals. J. Asia-Pac. Entomol. 2019, 22, 379–385. [Google Scholar] [CrossRef]
- McKenna, D.D.; Wild, A.L.; Kanda, K.; Bellamy, C.L.; Beutel, R.G.; Caterino, M.S.; Farnum, C.W.; Hawks, D.C.; Ivie, M.A.; Jameson, M.L.; et al. The beetle tree of life reveals that Coleoptera survived end-Permian mass extinction to diversify during the Cretaceous terrestrial revolution. Syst. Entomol. 2015, 40, 835–880. [Google Scholar] [CrossRef]
- Kaur, R.; Daniels, E.V.; Nayak, M.K.; Ebert, P.R.; Schlipalius, I.D. Determining changes in the distribution and abundance of aRhyzopertha dominicaphosphine resistance allele in farm grain storages using a DNA marker. Pest Manag. Sci. 2013, 69, 685–688. [Google Scholar] [CrossRef]
- Schlipalius, D.I.; Tuck, A.G.; Pavic, H.; Daglish, G.J.; Nayak, M.K.; Ebert, P.R. A high-throughput system used to determine frequency and distribution of phosphine resistance across large geographical regions. Pest Manag. Sci. 2019, 75, 1091–1098. [Google Scholar] [CrossRef] [PubMed]
- Nayak, M.K.; Jagadeesan, R.; Singarayan, V.T.; Nath, N.S.; Pavic, H.; Dembowski, B.; Daglish, G.J.; Schlipalius, D.I.; Ebert, P.R. First report of strong phosphine resistance in stored grain insects in a far northern tropical region of Australia, combining conventional and genetic diagnostics. J. Stored Prod. Res. 2021, 92, 101813. [Google Scholar] [CrossRef]
- Henikoff, S.; Ahmad, K.; Malik, H.S. The centromere paradox: Stable inheritance with rapidly evolving DNA. Science 2001, 293, 1098–1102. [Google Scholar] [CrossRef]
- Heslop-Harrison, J.S.; Schwarzacher, T. Nucleosomes and centromeric DNA packaging. Proc. Natl. Acad. Sci. USA 2013, 110, 19974–19975. [Google Scholar] [CrossRef] [PubMed]
- Ugarkovic, D.; Podnar, M.; Plohl, M. Satellite DNA of the red flour beetle Tribolium castaneum—Comparative study of satellites from the genus Tribolium. Mol. Biol. Evol. 1996, 13, 1059–1066. [Google Scholar] [CrossRef]
- Wang, S.; Lorenzen, M.; Beeman, R.W.; Brown, S.J. Analysis of repetitive DNA distribution patterns in the Tribolium castaneum genome. Genome Biol. 2008, 9, R61. [Google Scholar] [CrossRef]
- Saito, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783. [Google Scholar] [CrossRef]
- Schwarz, R.; Dayhoff, M. Matrices for detecting distant relationships. In Atlas of Protein Sequences; Dayhoff, M., Ed.; National Biomedical Research Foundation: Washington, DC, USA, 1979; pp. 353–358. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assembly | # Scaffolds | Total Mb | N50 1 (Mb) | L50 1 | N901 (Mb) | L90 1 | Longest Scaffold | BUSCO 2 |
|---|---|---|---|---|---|---|---|---|
| CANU | 1861 3 | 493 | 0.87 | 158 | 0.15 | 627 | 5,205,710 | 99.3 |
| Chicago/HiRise | 948 | 493 | 7.32 | 20 | 1.11 | 84 | 27,933,969 | 99.4 |
| Hybrid 4 | 336 | 479 | 7.44 | 19 | 1.48 | 74 | 27,934,817 | 97.6 |
| Hi-C/HiRise | 139 | 479 | 53.6 | 4 | 15.9 | 9 | 82,855,609 | 98.5 |
| Species | Subfamily | Total | Reference | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | D | E | F | G | H | |||
| Rhyzopertha dominica | 3 | 6 | 14 | 2 | 1 | 3 | 13 | 3 | 45 | This study |
| Diabrotica v. virgifera | 4 | 7 | 32 | 2 | 1 | 3 | 12 | 4 | 65 | [89] * |
| Aethina tumida | 4 | 6 | 24 | 2 | 1 | 3 | 13 | 3 | 56 | [90] |
| Tribolium castaneum | 10 | 6 | 35 | 2 | 1 | 3 | 14 | 3 | 74 | [61] |
| Altica viridicyanea | 8 | 9 | 37 | 2 | 1 | 3 | 8 | 1 | 69 | [91] |
| Chrysomela populi | 5 | 8 | 29 | 2 | 1 | 3 | 14 | 3 | 65 | [92] * |
| Drosophila melanogaster | 10 | 8 | 14 | 2 | 1 | 3 | 15 | 3 | 56 | [93] |
| Type | MEROPS | Peptidase * | R. dominica Sequence ID | Active Site Residues | RNA Expression (RPKM) | Signal Peptide (Amino Acid) | ||
|---|---|---|---|---|---|---|---|---|
| Gut | Carcass | Head | ||||||
| Serine | S9 | POP | RDOM022815 | SDH | 40.0 | 66.0 | 179 | n/a |
| DPP 4 | RDOM016604 | SDH | 29.0 | 15.0 | 6.00 | 13 | ||
| RDOM017825 | SDH | 13.0 | 10.0 | 9.00 | n/a | |||
| DPP 9 | RDOM002099 | SDH | 4.40 | 4.30 | 6.00 | n/a | ||
| DPP 10 | RDOM007500 | GDH | 0.60 | 1.30 | 1.30 | n/a | ||
| RDOM016697 | GDH | 0.04 | 11.0 | 26.0 | n/a | |||
| RDOM007053 | SDH | 0.50 | 1.00 | 1.00 | n/a | |||
| S28 | PRCP | RDOM021888 | SDH | 25.0 | 47.0 | 60.0 | 18 | |
| RDOM004644 | SDH | 56.0 | 105 | 79.0 | 18 | |||
| Metal-dependent | M24 | APP1 | RDOM014136 | DDHEE | 0.50 | 1.40 | 0.60 | n/a |
| RDOM016283 | DDHEE | 50.0 | 29.0 | 29.0 | n/a | |||
| APP3 | RDOM000819 | DDHEE | 246 | 222 | 88.0 | n/a | ||
| Prolidase | RDOM021565 | DDHEE | 182 | 74.0 | 71.0 | n/a | ||
| Name | Position | Intron # | CDS (bp) | Residues |
|---|---|---|---|---|
| RDO_Drip | Scaffold_97:54496632–54558762 − strand | 4 | 681 | 227 |
| RDO_Prip | Scaffold_97:54640277–54649322 + strand | 3 | 819 | 272 |
| RDO_Bigbrain | Scaffold_5:3510369–3521423 − strand | 3 | 1161 | 386 |
| RDO_AQP12L | Scaffold_2:13423224–13433006 − strand | 4 | 924 | 307 |
| RDO_Eglp1 | Scaffold_1:3563523–3572656 + strand | 5 | 942 | 313 |
| RDO_Eglp2 | Scaffold_1:34240108–34250443 + strand | 4 | 660 | 219 |
| RDO_Eglp3 | Scaffold_1:3546737–3555431 + strand | 6 | 888 | 295 |
| RDO_Eglp4 | Scaffold_1:26416818–26424599 − strand | 4 | 831 | 276 |
| Gene names | Annotation | Gene Coordinates | Expression (RPKM) | Active Site Residues | |||
|---|---|---|---|---|---|---|---|
| Adult | Gut 1 | Carcass 1 | Head 1 | ||||
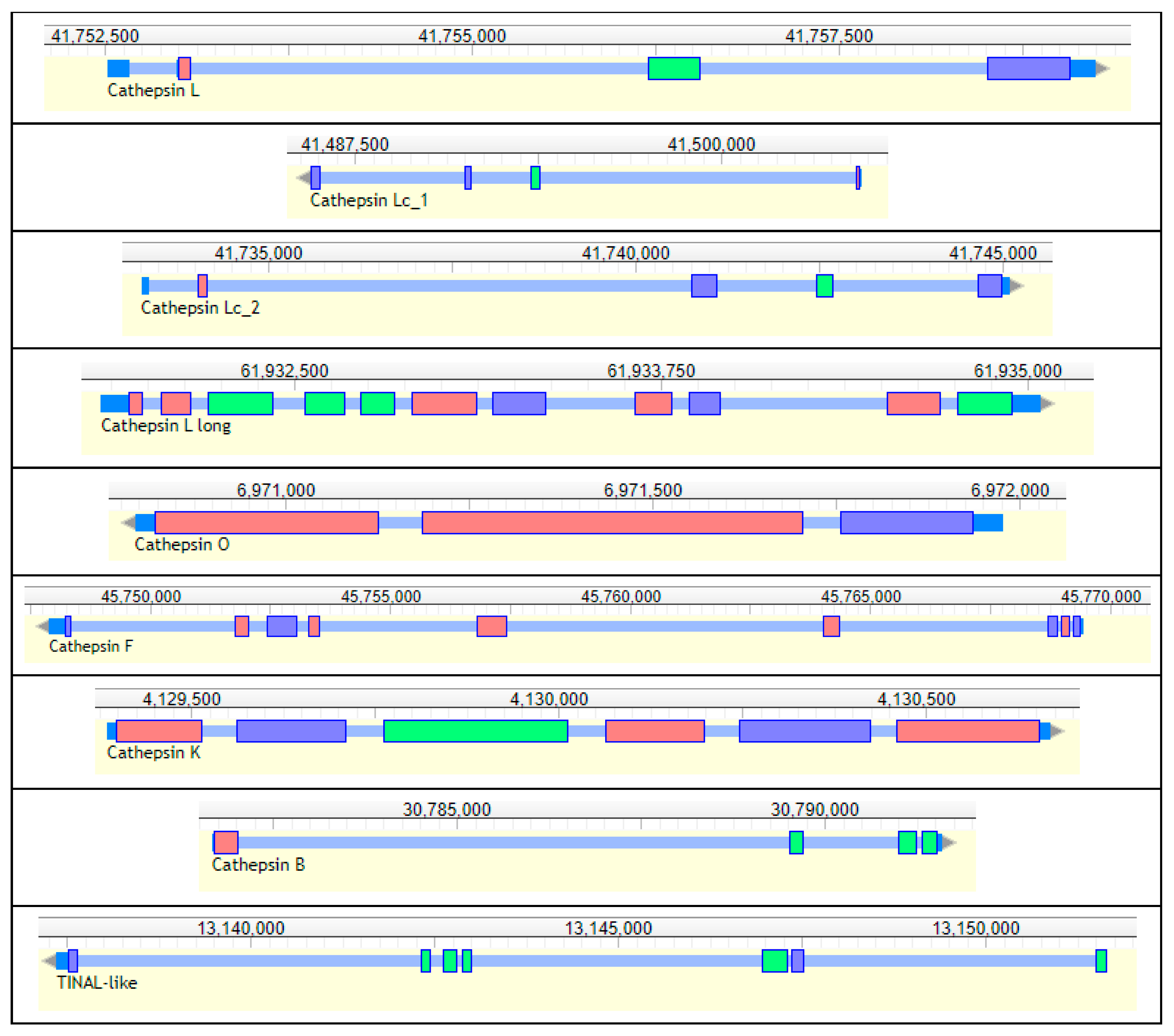
| RdL_97 | Cathepsin L | Scaffold_97:41752520–41759245 + strand | 305 | 202 | 272 | 359 | QCHN |
| RdLc1_97 | Cathepsin Lc1 | Scaffold_97:41485991–41504771 − strand | 5.79 | 0 | 2.17 | 0.49 | QCHN |
| RdLc2_97 | Cathepsin Lc2 | Scaffold_97:41733271–41745109 + strand | 115 | 14.8 | 903 | 335 | QCHN |
| RdLl_3 | Cathepsin L1 | Scaffold_3:61931842–61935045 + strand | 114 | 160 | 145 | 108 | QCHN |
| RdO_135 | Cathepsin O | Scaffold_135:6970795–6971976 − strand | 12.0 | 7.41 | 8.53 | 7.30 | QCHN |
| RdF_3 | Cathepsin F | Scaffold_3:45747905–45769454 − strand | 94.8 | 57.2 | 176 | 213 | QCHN |
| RdI_5 | Cathepsin I | Scaffold_5:4129386–4130669 + strand | 9.99 | 0.21 | 0.58 | 0.31 | QCHN |
| RdB_100 | Cathepsin B | Scaffold_100:30781674–30791599 + strand | 272 | 729 | 571 | 164 | QCHN HH |
| RdTINAL-like_3 | Cathepsin B TINAL-like | Scaffold_3:13137364-13151656 − strand | 32.3 | 27.4 | 35.3 | 43.9 | QSHN |
| Repeat Name | Consensus Monomer Length (bp) | Number of Arrays in a Cluster | Total Number of Monomers | Maximum Number of Monomers in an Array | Average Number of Monomers in an Array | AT Content of Consensus Sequence (%) | Average Monomer Similarity (%) | Genome Occupancy (bp) | % of the Assembled Genome |
|---|---|---|---|---|---|---|---|---|---|
| RD rep1 | 147 | 320 | 1836.8 | 37.1 | 5.7 | 52.4 | 93.6 | 270,614 | 0.0565 |
| RD rep2 | 123 | 308 | 3765.4 | 230.0 | 12.2 | 61.0 | 78.4 | 462,627 | 0.0965 |
| RD rep3 | 126 | 266 | 3389.4 | 190.7 | 12.7 | 63.5 | 85.2 | 418,239 | 0.0873 |
| RD rep4 | 126 | 182 | 2390.3 | 173.5 | 13.1 | 65.1 | 91.1 | 298,197 | 0.0622 |
| RD rep5 | 127 | 107 | 429.3 | 11.3 | 4.0 | 55.1 | 91.9 | 54,080 | 0.0113 |
| RD rep6 | 272 | 98 | 751.1 | 58.3 | 7.6 | 61.4 | 89.0 | 203,159 | 0.0424 |
| RD rep7 | 294 | 97 | 1350.2 | 59.7 | 13.9 | 59.5 | 86.6 | 397,984 | 0.0831 |
| RD rep8 | 110 | 97 | 260.2 | 7.0 | 2.7 | 63.6 | 96.4 | 28,933 | 0.0060 |
| RD rep9 | 135 | 95 | 734.4 | 23.7 | 7.7 | 65.2 | 89.4 | 95,988 | 0.0200 |
| RD rep10 | 153 | 89 | 898.4 | 31.7 | 10.1 | 54.9 | 93.6 | 137,075 | 0.0286 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oppert, B.; Muszewska, A.; Steczkiewicz, K.; Šatović-Vukšić, E.; Plohl, M.; Fabrick, J.A.; Vinokurov, K.S.; Koloniuk, I.; Johnston, J.S.; Smith, T.P.L.; et al. The Genome of Rhyzopertha dominica (Fab.) (Coleoptera: Bostrichidae): Adaptation for Success. Genes 2022, 13, 446. https://doi.org/10.3390/genes13030446
Oppert B, Muszewska A, Steczkiewicz K, Šatović-Vukšić E, Plohl M, Fabrick JA, Vinokurov KS, Koloniuk I, Johnston JS, Smith TPL, et al. The Genome of Rhyzopertha dominica (Fab.) (Coleoptera: Bostrichidae): Adaptation for Success. Genes. 2022; 13(3):446. https://doi.org/10.3390/genes13030446
Chicago/Turabian StyleOppert, Brenda, Anna Muszewska, Kamil Steczkiewicz, Eva Šatović-Vukšić, Miroslav Plohl, Jeffrey A. Fabrick, Konstantin S. Vinokurov, Igor Koloniuk, J. Spencer Johnston, Timothy P. L. Smith, and et al. 2022. "The Genome of Rhyzopertha dominica (Fab.) (Coleoptera: Bostrichidae): Adaptation for Success" Genes 13, no. 3: 446. https://doi.org/10.3390/genes13030446
APA StyleOppert, B., Muszewska, A., Steczkiewicz, K., Šatović-Vukšić, E., Plohl, M., Fabrick, J. A., Vinokurov, K. S., Koloniuk, I., Johnston, J. S., Smith, T. P. L., Guedes, R. N. C., Terra, W. R., Ferreira, C., Dias, R. O., Chaply, K. A., Elpidina, E. N., Tereshchenkova, V. F., Mitchell, R. F., Jenson, A. J., ... Campbell, J. F. (2022). The Genome of Rhyzopertha dominica (Fab.) (Coleoptera: Bostrichidae): Adaptation for Success. Genes, 13(3), 446. https://doi.org/10.3390/genes13030446