
Identification of Genetic Effects of ACADVL and IRF6 Genes with Milk Production Traits of Holstein Cattle in China


Abstract
1. Introduction
2. Materials and Methods
2.1. Animal and Phenotype Data Collection
2.2. SNP Identification and Genotyping
2.3. Linkage Disequilibrium (LD) Estimation and Association Analyses
2.4. Biological Function Prediction
3. Results
3.1. SNP Identification
3.2. Single-Locus Association Analyses with Five Milk Traits
3.3. Haplotype-Based Association Analyses with Five Milk Traits
3.4. Functional Variation Prediction Caused by SNPs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anderson, G.H.; Tecimer, S.N.; Shah, D.; Zafar, T.A. Protein source, quantity, and time of consumption determine the effect of proteins on short-term food intake in young men. J. Nutr. 2004, 134, 3011–3015. [Google Scholar] [CrossRef] [PubMed]
- Scholz-Ahrens, K.E.; Ahrens, F.; Barth, C.A. Nutritional and health attributes of milk and milk imitations. Eur. J. Nutr. 2020, 59, 19–34. [Google Scholar] [CrossRef]
- Poppitt, S.D. Cow's Milk and Dairy Consumption: Is There Now Consensus for Cardiometabolic Health? Front. Nutr. 2020, 7, 574725. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, M.U.; Trolle, E.; Outzen, M.; Mejborn, H.; Gronberg, M.G.; Lyndgaard, C.B.; Stockmarr, A.; Veno, S.K.; Bysted, A. Intake of dairy products and associations with major atherosclerotic cardiovascular diseases: A systematic review and meta-analysis of cohort studies. Sci. Rep. 2021, 11, 1303. [Google Scholar] [CrossRef]
- Cheng, R.; Wang, Q.; Wei, L. Income growth, employment structure transition and the rise of modern markets: The impact of urbanization on residents' consumption of dairy products in China. PLoS ONE 2022, 17, e0267006. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Yuan, R.; Zhang, Y.; Jin, X. Chinese Consumers' Preferences for Attributes of Fresh Milk: A Best-Worst Approach. Int. J. Environ. Res. Public Health 2019, 16, 4286. [Google Scholar] [CrossRef] [PubMed]
- Ikonen, T.; Morri, S.; Tyriseva, A.M.; Ruottinen, O.; Ojala, M. Genetic and phenotypic correlations between milk coagulation properties, milk production traits, somatic cell count, casein content, and pH of milk. J. Dairy Sci. 2004, 87, 458–467. [Google Scholar] [CrossRef] [PubMed]
- de Haas, Y.; Janss, L.L.; Kadarmideen, H.N. Genetic and phenotypic parameters for conformation and yield traits in three Swiss dairy cattle breeds. J. Anim. Breed. Genet. 2007, 124, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef]
- Goddard, M.E.; Hayes, B.J. Genomic selection. J. Anim. Breed. Genet. 2007, 124, 323–330. [Google Scholar] [CrossRef]
- Schefers, J.M.; Weigel, K.A. Genomic selection in dairy cattle: Integration of DNA testing into breeding programs. Anim. Front. 2012, 2, 4–9. [Google Scholar] [CrossRef]
- Wiggans, G.R.; Cole, J.B.; Hubbard, S.M.; Sonstegard, T.S. Genomic Selection in Dairy Cattle: The USDA Experience. Annu. Rev. Anim. Biosci. 2017, 5, 309–327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Ober, U.; Erbe, M.; Zhang, H.; Gao, N.; He, J.; Li, J.; Simianer, H. Improving the Accuracy of Whole Genome Prediction for Complex Traits Using the Results of Genome Wide Association Studies. PLoS ONE 2014, 9, e93017. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Khatib, H. Comparison of transcriptomic landscapes of bovine embryos using RNA-Seq. BMC Genomics. 2010, 11, 711. [Google Scholar] [CrossRef] [PubMed]
- Jain, M. Next-generation sequencing technologies for gene expression profiling in plants. Briefings Funct. Genomics 2012, 11, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.; Han, B.; Li, Q.; Yuan, Y.; Li, J.; Sun, D. Using RNA sequencing to identify putative competing endogenous RNAs (ceRNAs) potentially regulating fat metabolism in bovine liver. Sci. Rep. 2017, 7, 6396. [Google Scholar] [CrossRef]
- Aoyama, T.; Souri, M.; Ushikubo, S.; Kamijo, T.; Yamaguchi, S.; Kelley, R.I.; Rhead, W.J.; Uetake, K.; Tanaka, K.; Hashimoto, T. Purification of human very-long-chain acyl-coenzyme A dehydrogenase and characterization of its deficiency in seven patients. J. Clin. Investig. 1995, 95, 2465–2473. [Google Scholar] [CrossRef]
- Malloy, V.L.; Perrone, C.E.; Mattocks, D.A.L.; Ables, G.P.; Caliendo, N.S.; Orentreich, D.S.; Orentreich, N. Methionine restriction prevents the progression of hepatic steatosis in leptin-deficient obese mice. Metabolism 2013, 62, 1651–1661. [Google Scholar] [CrossRef]
- Loor, J.J.; Dann, H.M.; Everts, R.E.; Oliveira, R.; Green, C.A.; Guretzky, N.A.J.; Rodriguez-Zas, S.L.; Lewin, H.A.; Drackley, J.K. Temporal gene expression profiling of liver from periparturient dairy cows reveals complex adaptive mechanisms in hepatic function. Physiol. Genomics 2005, 23, 217–226. [Google Scholar] [CrossRef]
- Barletta, R.V.; Maturana Filho, M.; Carvalho, P.D.; Del Valle, T.A.; Netto, A.S.; Renno, F.P.; Mingoti, R.D.; Gandra, J.R.; Mourao, G.B.; Fricke, P.M.; et al. Association of changes among body condition score during the transition period with NEFA and BHBA concentrations, milk production, fertility, and health of Holstein cows. Theriogenology 2017, 104, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, T.; Ogasawara, K.; Takaoka, A.; Tanaka, N. IRF family of transcription factors as regulators of host defense. Annu. Rev. Immunol. 2001, 19, 623–655. [Google Scholar] [CrossRef] [PubMed]
- Barnes, B.J.; Kellum, M.J.; Pinder, K.E.; Frisancho, J.A.; Pitha, P.M. Interferon regulatory factor 5, a novel mediator of cell cycle arrest and cell death. Cancer Res. 2003, 63, 6424–6431. [Google Scholar]
- Duguay, D.; Mercier, F.; Stagg, J.; Martineau, D.; Bramson, J.; Servant, M.; Lin, R.T.; Galipeau, J.; Hiscott, J. In vivo interferon regulatory factor 3 tumor suppressor activity in B16 melanoma tumors. Cancer Res. 2002, 62, 5148–5152. [Google Scholar] [PubMed]
- Hu, G.D.; Mancl, M.E.; Barnes, B.J. Signaling through IFN regulatory factor-5 sensitizes p53-deficient tumors to DNA damage-induced apoptosis and cell death. Cancer Res. 2005, 65, 7403–7412. [Google Scholar] [CrossRef]
- Hennighausen, L.; Robinson, G.W. Information networks in the mammary gland. Nat. Rev. Mol. Cell Biol. 2005, 6, 715–725. [Google Scholar] [CrossRef]
- Blum, J.W.; Baumrucker, C.R. Colostral and milk insulin-like growth factors and related substances: Mammary gland and neonatal (intestinal and systemic) targets. Domest. Anim. Endocrinol. 2002, 23, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Bennewitz, J.; Reinsch, N.; Grohs, C.; Leveziel, H.; Malafosse, A.; Thomsen, H.; Xu, N.Y.; Looft, C.; Kuhn, C.; Brockmann, G.A.; et al. Combined analysis of data from two granddaughter designs: A simple strategy for QTL confirmation and increasing experimental power in dairy cattle. Genet. Sel. Evol. 2003, 35, 319–338. [Google Scholar] [CrossRef] [PubMed]
- Bennewitz, J.; Reinsch, N.; Guiard, V.; Fritz, S.; Thomsen, H.; Looft, C.; Kuhn, C.; Schwerin, M.; Weimann, C.; Erhardt, G.; et al. Multiple quantitative trait loci mappmg with cofactors and application of alternative variants of the false discovery rate in an enlarged granddaughter design. Genetics 2004, 168, 1019–1027. [Google Scholar] [CrossRef]
- Cai, Z.; Guldbrandtsen, B.; Lund, M.S.; Sahana, G. Dissecting closely linked association signals in combination with the mammalian phenotype database can identify candidate genes in dairy cattle. BMC Genet. 2019, 20, 1–12. [Google Scholar] [CrossRef]
- Fuseya, Y.; Sakurai, T.; Miyahara, J.-I.; Sato, K.; Kaji, S.; Saito, Y.; Takahashi, M.; Nishino, I.; Fukuda, T.; Sugie, H.; et al. Adult-onset Repeat Rhabdomyolysis with a Very Long-chain Acyl-CoA Dehydrogenase Deficiency Due to Compound Heterozygous ACADVL Mutations. Intern. Med. 2020, 59, 2729–2732. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, T.; Uchida, Y.; Kelley, R.I.; Marble, M.; Hofman, K.; Tonsgard, J.H.; Rhead, W.J.; Hashimoto, T. A novel disease with deficiency of mitochondrial very-long-chain acyl-CoA dehydrogenase. Biochem. Biophys. Res. Commun. 1993, 191, 1369–1372. [Google Scholar] [CrossRef]
- Lepori, V.; Muhlhause, F.; Sewell, A.C.; Jagannathan, V.; Janzen, N.; Rosati, M.; de Sousa, F.M.M.A.; Tschopp, A.; Schupbach, G.; Matiasek, K.; et al. A Nonsense Variant in the ACADVL Gene in German Hunting Terriers with Exercise Induced Metabolic Myopathy. G3-Genes Genom. Genet. 2018, 8, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Drackley, J.K.; Overton, T.R.; Douglas, G.N. Adaptations of glucose and long-chain fatty acid metabolism in liver of dairy cows during the periparturient period. J. Dairy Sci. 2001, 84, E100–E112. [Google Scholar] [CrossRef]
- McAndrew, R.P.; Wang, Y.; Mohsen, A.-W.; He, M.; Vockley, J.; Kim, J.-J.P. Structural basis for substrate fatty acyl chain specificity-Crystal structure of human very-long-chain acyl-CoA dehydrogenase. J. Biol. Chem. 2008, 283, 9435–9443. [Google Scholar] [CrossRef]
- Tucci, S.; Primassin, S.; Ter Veld, F.; Spiekerkoetter, U. Medium-chain triglycerides impair lipid metabolism and induce hepatic steatosis in very long-chain acyl-CoA dehydrogenase (VLCAD)-deficient mice. Mol. Genet. Metab. 2010, 101, 40–47. [Google Scholar] [CrossRef]
- Ibeagha-Awemu, E.M.; Li, R.; Ammah, A.A.; Dudemaine, P.L.; Bissonnette, N.; Benchaar, C.; Zhao, X. Transcriptome adaptation of the bovine mammary gland to diets rich in unsaturated fatty acids shows greater impact of linseed oil over safflower oil on gene expression and metabolic pathways. BMC Genomics 2016, 17, 1–23. [Google Scholar] [CrossRef]
- Wiedemann, S.; Sigl, G.; Schmautz, C.; Kaske, M.; Viturro, E.; Meyer, H.H.D. Omission of dry period or milking once daily affects metabolic status and is reflected by mRNA levels of enzymes in liver and muscle of dairy cows. Livest. Sci. 2013, 154, 193–203. [Google Scholar] [CrossRef]
- do Amaral, B.C.; Connor, E.E.; Tao, S.; Hayen, J.; Bubolz, J.; Dahl, G.E. Heat-stress abatement during the dry period: Does cooling improve transition into lactation? J. Dairy Sci. 2009, 92, 5988–5999. [Google Scholar] [CrossRef]
- Bailey, C.M.; Abbott, D.E.; Margaryan, N.V.; Khalkhali-Ellis, Z.; Hendrix, M.J.C. Interferon regulatory factor 6 promotes cell cycle arrest and is regulated by the proteasome in a cell cycle-dependent manner. Mol. Cell. Biol. 2008, 28, 2235–2243. [Google Scholar] [CrossRef]
- Bailey, C.M.; Hendrix, M.J.C. IRF6 in development and disease: A mediator of quiescence and differentiation. Cell Cycle 2008, 7, 1925–1930. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.M.; Margaryan, N.V.; Abbott, D.E.; Schutte, B.C.; Yang, B.; Khalkhali-Ellis, Z.; Hendrix, M.J.C. Temporal and spatial expression patterns for the tumor suppressor Maspin and its binding partner interferon regulatory factor 6 during breast development. Dev. Growth Differ. 2009, 51, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Fuerst, C.; Solkner, J. Additive and nonadditive genetic variances for milk yield, fertility, and lifetime performance traits of dairy cattle. J Dairy Sci. 1994, 77, 1114–1125. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Christensen, O.F.; Ostersen, T.; Henryon, M.; Lund, M.S. Estimating additive and non-additive genetic variances and predicting genetic merits using genome-wide dense single nucleotide polymorphism markers. PLoS ONE 2012, 7, e45293. [Google Scholar] [CrossRef]
- Calkhoven, C.F.; Ab, G. Multiple steps in the regulation of transcription-factor level and activity. Biochem. J. 1996, 317 Pt 2, 329–342. [Google Scholar] [CrossRef]
- Kasowski, M.; Grubert, F.; Heffelfinger, C.; Hariharan, M.; Asabere, A.; Waszak, S.M.; Habegger, L.; Rozowsky, J.; Shi, M.; Urban, A.E.; et al. Variation in Transcription Factor Binding Among Humans. Science 2010, 328, 232–235. [Google Scholar] [CrossRef]
- Aigner, K.; Dampier, B.; Descovich, L.; Mikula, M.; Sultan, A.; Schreiber, M.; Mikulits, W.; Brabletz, T.; Strand, D.; Obrist, P.; et al. The transcription factor ZEB1 (deltaEF1) promotes tumour cell dedifferentiation by repressing master regulators of epithelial polarity. Oncogene 2007, 26, 6979–6988. [Google Scholar] [CrossRef]
- Watanabe, Y.; Kawakami, K.; Hirayama, Y.; Nagano, K. Transcription factors positively and negatively regulating the Na,K-ATPase alpha 1 subunit gene. J. Biochem. 1993, 114, 849–855. [Google Scholar] [CrossRef]
- Landrette, S.F.; Madera, D.; He, F.; Castilla, L.H. The transcription factor PlagL2 activates Mpl transcription and signaling in hematopoietic progenitor and leukemia cells. Leukemia 2011, 25, 655–662. [Google Scholar] [CrossRef]
- Borgmann, J.; Tuttelmann, F.; Dworniczak, B.; Ropke, A.; Song, H.W.; Kliesch, S.; Wilkinson, M.F.; Laurentino, S.; Gromoll, J. The human RHOX gene cluster: Target genes and functional analysis of gene variants in infertile men. Hum. Mol. Genet. 2016, 25, 4898–4910. [Google Scholar] [CrossRef]


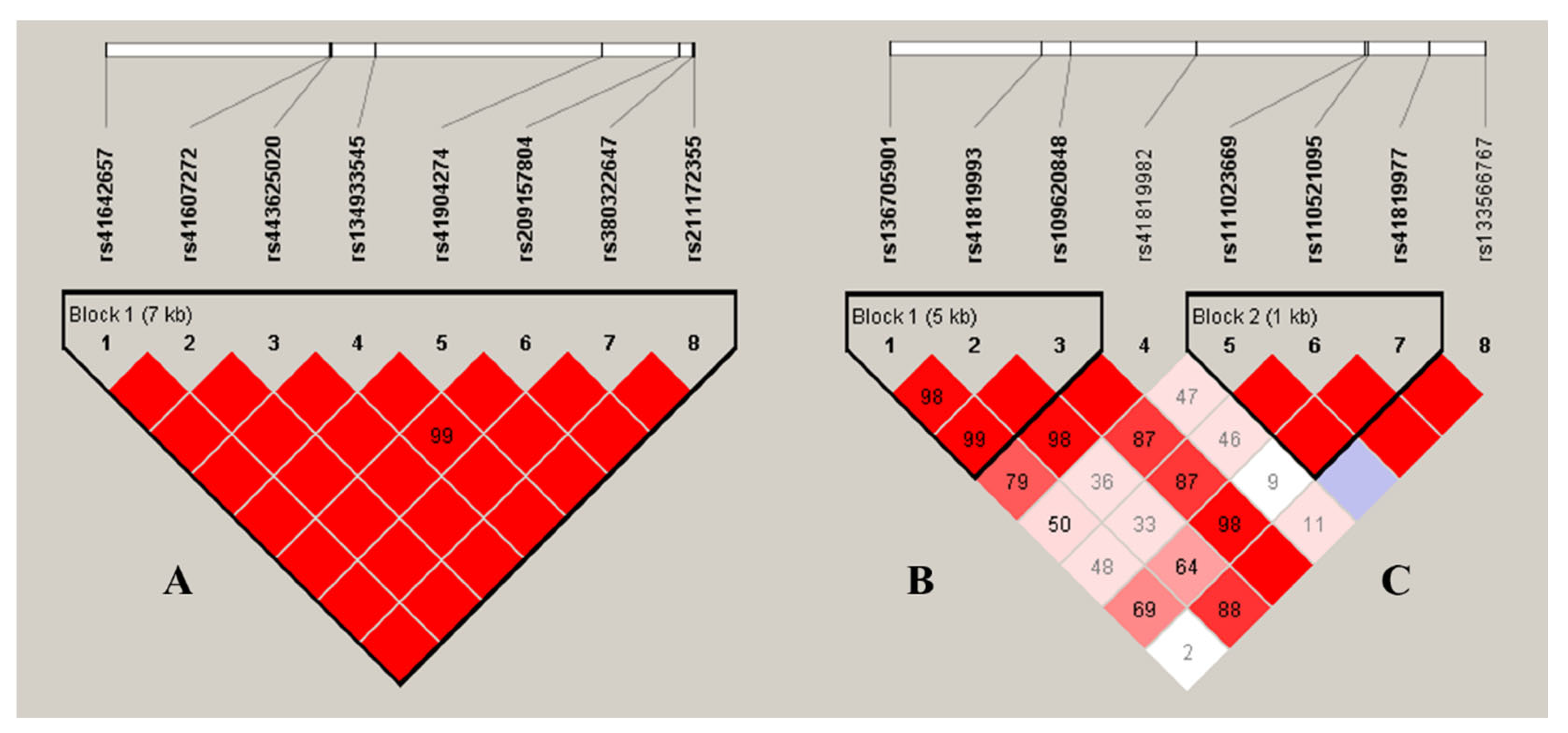{kind=link}
| Gene | SNP Name | GenBank No. | Position (ARS-UCD1.2) | Location | Allele | Allelic Frequency | Genotype | Genotypic Frequency |
|---|---|---|---|---|---|---|---|---|
| ACADVL | g.26933503T>C | rs41642657 | chr19:26933503 | 5´flanking region | G | 0.73 | AA | 0.07 |
| A | 0.27 | AG | 0.39 | |||||
| GG | 0.54 | |||||||
| g.26936476T>C | rs41607272 | chr19:26936476 | Intron-7 | C | 0.63 | CC | 0.40 | |
| T | 0.37 | CT | 0.46 | |||||
| TT | 0.14 | |||||||
| g.26936494C>G | rs443625020 | chr19:26936494 | Intron-7 | A | 0.51 | AA | 0.25 | |
| G | 0.49 | AG | 0.53 | |||||
| GG | 0.23 | |||||||
| g.26937081G>C | rs134933545 | chr19:26937081 | Intron-9 | T | 0.87 | CC | 0.02 | |
| C | 0.13 | CT | 0.23 | |||||
| TT | 0.75 | |||||||
| g.26940095G>T | rs41904274 | chr19:26940095 | 3´flanking region | C | 0.95 | CC | 0.89 | |
| T | 0.05 | CT | 0.11 | |||||
| TT | 0.00 | |||||||
| g.26941116C>A | rs209157804 | chr19:26941116 | 3´flanking region | C | 0.94 | CC | 0.89 | |
| T | 0.06 | CT | 0.11 | |||||
| TT | 0.00 | |||||||
| g.26941290A>G | rs380322647 | chr19:26941290 | 3´flanking region | C | 0.58 | CC | 0.33 | |
| G | 0.42 | CG | 0.50 | |||||
| GG | 0.17 | |||||||
| g.26941299G>A | rs211172355 | chr19:26941299 | 3´flanking region | G | 0.88 | AA | 0.01 | |
| A | 0.12 | AG | 0.21 | |||||
| GG | 0.77 | |||||||
| IRF6 | g.73501985G>A | rs136705901 | chr16:73501985 | 5´flanking region | C | 0.77 | CC | 0.60 |
| T | 0.23 | CT | 0.34 | |||||
| TT | 0.06 | |||||||
| g.73506456C>T | rs41819993 | chr16:73506456 | Intron-1 | C | 0.88 | CC | 0.77 | |
| T | 0.12 | CT | 0.22 | |||||
| TT | 0.02 | |||||||
| g.73507276G>A | rs109620848 | chr16:73507276 | Intron-3 | C | 0.94 | CC | 0.88 | |
| G | 0.06 | CG | 0.11 | |||||
| GG | 0.01 | |||||||
| g.73510990T>C | rs41819982 | chr16:73510990 | Intron-3 | G | 0.58 | CC | 0.17 | |
| C | 0.42 | CG | 0.49 | |||||
| GG | 0.33 | |||||||
| g.73515941C>T | rs111023669 | chr16:73515941 | Intron-7 | G | 0.77 | GG | 0.60 | |
| T | 0.23 | GT | 0.34 | |||||
| TT | 0.06 | |||||||
| g.73516019C>T | rs110521095 | chr16:73516019 | Exon-8 | C | 0.58 | AA | 0.17 | |
| A | 0.42 | AC | 0.49 | |||||
| CC | 0.34 | |||||||
| g.73517802G>C | rs41819977 | chr16:73517802 | Intron-9 | G | 0.77 | AA | 0.06 | |
| A | 0.23 | AG | 0.34 | |||||
| GG | 0.60 | |||||||
| g.73519434G>A | rs133566767 | chr16:73519434 | 3´flanking region | A | 0.77 | AA | 0.59 | |
| G | 0.23 | AG | 0.35 | |||||
| GG | 0.06 |
| Gene | SNP Name | Lactation | Genotype(n) | Milk Yield (kg) | Fat Yield (kg) | Fat Percentage (%) | Protein Yield (kg) | Protein Percentage (%) |
|---|---|---|---|---|---|---|---|---|
| ACADVL | g.26933503T>C | 1 | CC(676) | 8503.26 ± 113.07 | 301.73 ± 4.583 a | 3.559 ± 0.045 | 262.28 ± 3.343 | 3.092 ± 0.027 |
| CT(377) | 8572.99 ± 115.7 | 304.66 ± 4.68 ab | 3.553 ± 0.046 | 264.74 ± 3.414 | 3.091 ± 0.027 | |||
| TT(66) | 8681.75 ± 137.9 | 311.33 ± 5.504 a | 3.597 ± 0.055 | 266.98 ± 4.016 | 3.07 ± 0.031 | |||
| p value | 0.0503 | 0.0064 ** | 0.4438 | 0.0341 * | 0.4564 | |||
| 2 | CC(345) | 9080.59 ± 125.48 | 336.12 ± 5.330 | 3.717 ± 0.051 | 284.39 ± 3.884 b | 3.154 ± 0.032 | ||
| CT(233) | 9092.22 ± 128.84 | 330.73 ± 5.447 | 3.665 ± 0.052 | 284.43 ± 3.97 b | 3.154 ± 0.033 | |||
| TT(38) | 9171.66 ± 166.27 | 330.31 ± 6.832 | 3.612 ± 0.067 | 293.27 ± 4.982 a | 3.197 ± 0.04 | |||
| p value | 0.7321 | 0.0414 * | 0.0127 * | 0.024 * | 0.2271 | |||
| g.26936476T>C | 1 | CC(847) | 7999.03 ± 110.94 B | 280.56 ± 4.504 B | 3.557 ± 0.044 A | 248.09 ± 3.285 b | 3.126 ± 0.026 | |
| CT(242) | 8135.8 ± 115.55 A | 287.67 ± 4.67 A | 3.567 ± 0.046 A | 252.12 ± 3.406 a | 3.123 ± 0.027 | |||
| TT(20) | 7964.4 ± 185.93 AB | 260.64 ± 7.298 C | 3.337 ± 0.074 B | 244.55 ± 5.327 ab | 3.091 ± 0.041 | |||
| p value | 0.0172 * | <0.0001 ** | 0.0007 ** | 0.0071 ** | 0.5194 | |||
| 2 | CC(466) | 9883.54 ± 124.95 A | 350.62 ± 5.313 | 3.608 ± 0.051 | 307.33 ± 3.872 a | 3.113 ± 0.032 | ||
| CT(139) | 9664.45 ± 135.07 B | 345.24 ± 5.683 | 3.622 ± 0.055 | 302.55 ± 4.142 b | 3.142 ± 0.034 | |||
| TT(6) | 9795.21 ± 299.52 AB | 343.24 ± 11.917 | 3.567 ± 0.119 | 312.17 ± 8.697 ab | 3.186 ± 0.068 | |||
| p value | 0.004 ** | 0.0975 | 0.8024 | 0.0295 * | 0.0731 | |||
| g.26936494C>G | 1 | CC(988) | 8526.7 ± 112.65 | 294.64 ± 4.569 B | 3.454 ± 0.045 B | 265.91 ± 3.333 C | 3.125 ± 0.027 B | |
| CG(127) | 8591.62 ± 122.88 | 305.3 ± 4.942 A | 3.552 ± 0.049 A | 273.11 ± 3.606 B | 3.177 ± 0.029 A | |||
| GG(5) | 9226.25 ± 315.6 | 308.3 ± 12.238 AB | 3.276 ± 0.125 AB | 300.13 ± 8.936 A | 3.244 ± 0.067 AB | |||
| p value | 0.0397* | <0.0001 ** | 0.0001 ** | <0.0001 ** | 0.0001 ** | |||
| 2 | CC(528) | 9438.08 ± 124.32 A | 357.47 ± 5.282 B | 3.808 ± 0.051 C | 299.36 ± 3.849 A | 3.177 ± 0.032 | ||
| CG(82) | 9533.25 ± 138.85 A | 372.54 ± 5.8 A | 3.934 ± 0.056 B | 304.54 ± 4.229 A | 3.195 ± 0.035 | |||
| GG(5) | 8194.37 ± 323.04 B | 385.75 ± 12.85 AB | 4.596 ± 0.129 A | 254.37 ± 9.378 B | 3.194 ± 0.073 | |||
| p value | 0.0001 ** | <0.0001 ** | <0.0001 ** | <0.0001 ** | 0.5947 | |||
| g.26937081G>C | 1 | CC(197) | 7930.49 ± 117.93 C | 301.85 ± 4.756 Bc | 3.827 ± 0.047 A | 242.48 ± 3.47 C | 3.068 ± 0.028 | |
| CG(548) | 8183.27 ± 112.03 B | 307.3 ± 4.541 Bb | 3.755 ± 0.045 B | 250.56 ± 3.312 B | 3.067 ± 0.026 | |||
| GG(371) | 8326.4 ± 113.49 A | 315.64 ± 4.594 Aa | 3.784 ± 0.045 AB | 256.54 ± 3.352 A | 3.073 ± 0.027 | |||
| p value | <0.0001 ** | <0.0001 ** | 0.0034 ** | <0.0001 ** | 0.7694 | |||
| 2 | CC(94) | 9001.11 ± 135.56 | 317.48 ± 5.682 Bc | 3.59 ± 0.055 Bb | 283.72 ± 4.142 b | 3.176 ± 0.034 | ||
| CG(298) | 9175.35 ± 127.5 | 338.72 ± 5.399 Aa | 3.729 ± 0.052 Aa | 287.63 ± 3.935 ab | 3.154 ± 0.033 | |||
| GG(223) | 9154.33 ± 131.4 | 332.84 ± 5.541 Ab | 3.67 ± 0.053A Bb | 290.14 ± 4.038 a | 3.185 ± 0.033 | |||
| p value | 0.0956 | <0.0001 ** | <0.0001 ** | 0.0314 * | 0.0694 | |||
| g.26940095G>T | 1 | GG(676) | 8208.1 ± 113.55 | 296.19 ± 4.602 B | 3.606 ± 0.045 | 260.61 ± 3.357 | 3.188 ± 0.027 | |
| GT(375) | 8264.01 ± 114.89 | 299.46 ± 4.651A B | 3.612 ± 0.046 | 262.45 ± 3.393 | 3.186 ± 0.027 | |||
| TT(66) | 8391.01 ± 139.35 | 305.71 ± 5.558 A | 3.642 ± 0.056 | 265.69 ± 4.056 | 3.167 ± 0.032 | |||
| p value | 0.0653 | 0.0051 ** | 0.5421 | 0.054 | 0.5252 | |||
| 2 | GG(344) | 9038.88 ± 125.2 | 333.34 ± 5.319 a | 3.709 ± 0.051 | 283.55 ± 3.876 b | 3.161 ± 0.032 | ||
| GT(233) | 9035.65 ± 129.27 | 327.46 ± 5.461 b | 3.657 ± 0.053 | 283.01 ± 3.98 b | 3.16 ± 0.033 | |||
| TT(38) | 9118.61 ± 165.87 | 327.23 ± 6.815 ab | 3.606 ± 0.067 | 292.05 ± 4.97 a | 3.204 ± 0.04 | |||
| p value | 0.7733 | 0.0234 * | 0.0135 * | 0.0258 * | 0.2131 | |||
| g.26941116C>A | 1 | AA(197) | 8132.94 ± 120.04 C | 298.26 ± 4.838 B | 3.668 ± 0.048 a | 250.77 ± 3.529 C | 3.09 ± 0.028 | |
| AC(547) | 8360.68 ± 112.47 B | 303.25 ± 4.558 B | 3.604 ± 0.045 b | 258.36 ± 3.325 B | 3.095 ± 0.027 | |||
| CC(371) | 8497.37 ± 113.5 A | 312.53 ± 4.594 A | 3.648 ± 0.045 a | 264.27 ± 3.351 A | 3.102 ± 0.027 | |||
| p value | <0.0001 ** | <0.0001 ** | 0.0038 ** | <0.0001 ** | 0.5866 | |||
| 2 | AA(93) | 8998.55 ± 135.9 | 317.89 ± 5.694 b | 3.595 ± 0.055 Bb | 282.64 ± 4.151 b | 3.169 ± 0.034 | ||
| AC(297) | 9126.35 ± 127.65 | 335.43 ± 5.408 a | 3.723 ± 0.052 Aa | 285.51 ± 3.941 ab | 3.155 ± 0.033 | |||
| CC(223) | 9119.86 ± 130.63 | 332.93 ± 5.518 a | 3.689 ± 0.053 ABa | 288.62 ± 4.022 a | 3.186 ± 0.033 | |||
| p value | 0.27 | <0.0001 ** | 0.0004 ** | 0.0375 * | 0.0742 | |||
| g.26941290A>G | 1 | AA(66) | 8451.25 ± 139.28 | 301.81 ± 5.554 a | 3.564 ± 0.055 | 267.63 ± 4.053 | 3.166 ± 0.032 | |
| AG(376) | 8332.13 ± 115.26 | 295.75 ± 4.665 ab | 3.533 ± 0.046 | 264.79 ± 3.403 | 3.186 ± 0.027 | |||
| GG(676) | 8263.88 ± 113.11 | 292.48 ± 4.585 a | 3.532 ± 0.045 | 262.41 ± 3.344 | 3.187 ± 0.027 | |||
| p value | 0.0431 * | 0.0059 ** | 0.646 | 0.026 * | 0.5244 | |||
| 2 | AA(38) | 9171.66 ± 166.27 | 330.31 ± 6.832 | 3.612 ± 0.067 | 293.27 ± 4.982 a | 3.197 ± 0.04 | ||
| AG(233) | 9092.22 ± 128.84 | 330.73 ± 5.447 | 3.665 ± 0.052 | 284.43 ± 3.97 b | 3.154 ± 0.033 | |||
| GG(345) | 9080.59 ± 125.48 | 336.12 ± 5.33 | 3.717 ± 0.051 | 284.39 ± 3.884 b | 3.154 ± 0.032 | |||
| p value | 0.7321 | 0.0414 * | 0.0127 * | 0.024 * | 0.2271 | |||
| g.26941299G>A | 1 | AA(663) | 8179.63 ± 111.32 | 304.32 ± 4.516 b | 3.724 ± 0.045 | 255.6 ± 3.294 | 3.129 ± 0.026 | |
| AG(382) | 8222.8 ± 113.43 | 307.55 ± 4.594 ab | 3.734 ± 0.045 | 257 ± 3.351 | 3.126 ± 0.027 | |||
| GG(67) | 8356.95 ± 136.42 | 313.66 ± 5.441 a | 3.764 ± 0.054 | 260.29 ± 3.971 | 3.107 ± 0.031 | |||
| p value | 0.0928 | 0.006 ** | 0.4567 | 0.1049 | 0.4885 | |||
| 2 | AA(342) | 9842.47 ± 125.39 | 361.58 ± 5.331 a | 3.693 ± 0.051 a | 307.92 ± 3.885 b | 3.122 ± 0.032 | ||
| AG(231) | 9829.35 ± 130.48 | 354.34 ± 5.511 b | 3.632 ± 0.053 b | 307.58 ± 4.017 b | 3.125 ± 0.033 | |||
| GG(39) | 9931.31 ± 163.51 | 356.58 ± 6.727 ab | 3.599 ± 0.066 ab | 316.67 ± 4.906 a | 3.163 ± 0.04 | |||
| p value | 0.6871 | 0.006 ** | 0.0097 ** | 0.0219 * | 0.2575 | |||
| IRF6 | g.73501985G>A | 1 | AA(79) | 8354.21 ± 133.24 | 298.43 ± 5.331 | 3.593 ± 0.053 A | 261.08 ± 3.89 | 3.148 ± 0.031 |
| AG(436) | 8424.55 ± 115.21 | 294.49 ± 4.663 | 3.517 ± 0.046 AB | 261.44 ± 3.401 | 3.12 ± 0.027 | |||
| GG(601) | 8432.88 ± 113.56 | 293.38 ± 4.601 | 3.492 ± 0.045 B | 262.83 ± 3.356 | 3.132 ± 0.027 | |||
| p value | 0.6302 | 0.2679 | 0.0059 ** | 0.4531 | 0.1866 | |||
| 2 | AA(70) | 9454.76 ± 145.55 A | 346.64 ± 6.059 A | 3.682 ± 0.059 ABa | 303.04 ± 4.418 A | 3.218 ± 0.036 | ||
| AG(237) | 9154.08 ± 128.85 B | 333.65 ± 5.452 A | 3.69 ± 0.052 Aa | 292.1 ± 3.973 B | 3.204 ± 0.033 | |||
| GG(307) | 9293.21 ± 126.94 AB | 328.18 ± 5.385 B | 3.59 ± 0.052 Bb | 294.64 ± 3.924 B | 3.186 ± 0.033 | |||
| p value | 0.0029 ** | <0.0001 ** | <0.0001 ** | 0.0003 ** | 0.1812 | |||
| g.73506456C>T | 1 | CC(445) | 7897.68 ± 112.44 ab | 295.58 ± 4.556 AB | 3.756 ± 0.045 | 248.36 ± 3.323 ABb | 3.153 ± 0.027 | |
| CT(516) | 7965.9 ± 114.21 a | 299.01 ± 4.623 AB | 3.773 ± 0.046 | 249.84 ± 3.372 Aa | 3.146 ± 0.027 | |||
| TT(155) | 7799.49 ± 121.56 b | 290.23 ± 4.895 B | 3.724 ± 0.049 | 243.81 ± 3.571 Bb | 3.129 ± 0.028 | |||
| p value | 0.0191 * | 0.0007 ** | 0.1217 | 0.0024 ** | 0.1864 | |||
| 2 | CC(223) | 9722.67 ± 129.26 a | 325.65 ± 5.465 a | 3.408 ± 0.053 | 305.81 ± 3.983 Aa | 3.141 ± 0.033 | ||
| CT(275) | 9462.99 ± 127.27 b | 319.56 ± 5.391 b | 3.447 ± 0.052 | 298.89 ± 3.929 Bb | 3.155 ± 0.033 | |||
| TT(114) | 9467.5 ± 140.44 b | 324.78 ± 5.881 ab | 3.48 ± 0.057 | 299.54 ± 4.287 ABb | 3.164 ± 0.035 | |||
| p value | 0.0001 ** | 0.0332 * | 0.0689 | 0.0005 ** | 0.378 | |||
| g.73507276G>A | 1 | AA(278) | 8172.47 ± 116.63 | 295.15 ± 4.708 A | 3.644 ± 0.047 A | 255.84 ± 3.434 Aa | 3.134 ± 0.027 a | |
| AG(589) | 8125.38 ± 111.75 | 295.57 ± 4.53 A | 3.676 ± 0.045 A | 253.91 ± 3.304 ABb | 3.127 ± 0.026 ab | |||
| GG(248) | 8063.09 ± 116.45 | 286.89 ± 4.706 B | 3.59 ± 0.047 B | 249.75 ± 3.433 Bb | 3.102 ± 0.027 b | |||
| p value | 0.1877 | <0.0001 ** | 0.0001 ** | 0.0012 ** | 0.0256 * | |||
| 2 | AA(131) | 9601.09 ± 137.6 A | 323.29 ± 5.763 A | 3.437 ± 0.056 | 307.73 ± 4.201 A | 3.213 ± 0.035 | ||
| AG(301) | 9202.86 ± 126.33 B | 309.87 ± 5.354 B | 3.452 ± 0.051 | 291.54 ± 3.902 B | 3.18 ± 0.032 | |||
| GG(181) | 9249.49 ± 130.95 B | 318.18 ± 5.531 A | 3.51 ± 0.053 | 294.65 ± 4.031 B | 3.2 ± 0.033 | |||
| p value | <0.0001 ** | <0.0001 ** | 0.0428 * | <0.0001 ** | 0.087 | |||
| g.73510990T>C | 1 | CC(18) | 7571.84 ± 192.55 | 286.46 ± 7.585 | 3.771 ± 0.076 ab | 234.72 ± 5.537 | 3.101 ± 0.043 | |
| CT(253) | 7875.23 ± 118.2 | 291.32 ± 4.769 | 3.729 ± 0.047 a | 243.51 ± 3.479 | 3.095 ± 0.028 | |||
| TT(844) | 7938.29 ± 111.14 | 290.62 ± 4.51 | 3.681 ± 0.045 b | 245.68 ± 3.289 | 3.099 ± 0.026 | |||
| p value | 0.0467 * | 0.7327 | 0.0237 * | 0.0249 * | 0.9216 | |||
| 2 | CC(14) | 9239.48 ± 223.03 AB | 286.5 ± 8.962 B | 3.221 ± 0.089 Bc | 290.84 ± 6.538 AB | 3.157 ± 0.052 | ||
| CT(161) | 9185.21 ± 134.07 B | 315.46 ± 5.642 A | 3.507 ± 0.054 Aa | 292.34 ± 4.113 B | 3.193 ± 0.034 | |||
| TT(437) | 9426.39 ± 124.47 A | 317.57 ± 5.289 A | 3.441 ± 0.051 ABb | 298.7 ± 3.855 A | 3.173 ± 0.032 | |||
| p value | 0.0008 ** | 0.0002 ** | 0.0002 ** | 0.0014 ** | 0.3069 | |||
| g.73515941C>T | 1 | CC(1003) | 8506.3 ± 110.67 | 310.91 ± 4.492 b | 3.647 ± 0.044 B | 267.41 ± 3.277 B | 3.158 ± 0.026 B | |
| CT(113) | 8370.39 ± 125.12 | 316.06 ± 5.024 ab | 3.817 ± 0.05 A | 265.64 ± 3.665 B | 3.187 ± 0.029 B | |||
| TT(1) | 9251.82 ± 670.07 | 373.98 ± 26.155 a | 4.061 ± 0.265 AB | 330 ± 19.096 A | 3.616 ± 0.146 A | |||
| p value | 0.063 | 0.0079 ** | <0.0001 ** | 0.0024 ** | 0.0009 ** | |||
| 2 | CC(528) | 9532.29 ± 123.49 Aa | 346.44 ± 5.252 a | 3.668 ± 0.05 Bb | 303.16 ± 3.828 Ac | 3.193 ± 0.032 | ||
| CT(84) | 9105.47 ± 146.94 Bb | 338.49 ± 6.127 b | 3.776 ± 0.059 Aa | 292.99 ± 4.467 Bb | 3.229 ± 0.037 | |||
| TT(1) | 10887 ± 683.11A Ba | 327.89 ± 26.719 ab | 3.098 ± 0.27 ABb | 352.94 ± 19.506 Aa | 3.27 ± 0.149 | |||
| p value | <0.0001 ** | 0.0389 * | 0.0004 ** | <0.0001 ** | 0.1249 | |||
| g.73516019C>T | 1 | CC(998) | 8372.13 ± 110.55 | 298.85 ± 4.486 b | 3.55 ± 0.044 B | 261.19 ± 3.273 B | 3.115 ± 0.026 Bb | |
| CT(118) | 8257.85 ± 124.97 | 303.54 ± 5.019b | 3.697 ± 0.05 A | 260.44 ± 3.663 B | 3.144 ± 0.029 ABb | |||
| TT(1) | 9084.23 ± 670.37 | 367.82 ± 26.168 a | 4.04 ± 0.265 AB | 322.11 ± 19.105 A | 3.568 ± 0.146 Aa | |||
| p value | 0.1163 | 0.0057 ** | <0.0001 ** | 0.0049 ** | 0.001 ** | |||
| 2 | CC(527) | 9409.02 ± 123.29 Aa | 341.74 ± 5.24 a | 3.675 ± 0.05 Bb | 297.8 ± 3.818 Ab | 3.179 ± 0.032 | ||
| CT(86) | 8957.16 ± 142.76 Bb | 332.59 ± 5.949 b | 3.779 ± 0.058 Aa | 286.29 ± 4.337 Bc | 3.209 ± 0.036 | |||
| TT(1) | 10705 ± 683.12A Ba | 322.07 ± 26.723 ab | 3.111 ± 0.27 ABb | 345.24 ± 19.509 Aa | 3.249 ± 0.149 | |||
| p value | <0.0001 ** | 0.0143 * | 0.0005** | <0.0001 ** | 0.2275 | |||
| g.73517802G>C | 1 | CC(373) | 8212.21 ± 113.87 | 302.64 ± 4.609 a | 3.695 ± 0.046 A | 256.64 ± 3.362 a | 3.131 ± 0.027 a | |
| CG(561) | 8131.65 ± 111.93 | 300.75 ± 4.541 ab | 3.714 ± 0.045 A | 253.28 ± 3.312 b | 3.12 ± 0.026 ab | |||
| GG(180) | 8252.11 ± 119.05 | 296.37 ± 4.803 b | 3.595 ± 0.048 B | 255 ± 3.504 ab | 3.094 ± 0.028 b | |||
| p value | 0.0514 | 0.0311 * | <0.0001 ** | 0.0243 * | 0.0198 * | |||
| 2 | CC(184) | 9705.43 ± 132.79 A | 343.25 ± 5.587 A | 3.584 ± 0.054 ab | 305.31 ± 4.073 A | 3.148 ± 0.034 | ||
| CG(295) | 9479.69 ± 126.72 B | 332.01 ± 5.372 B | 3.558 ± 0.052 b | 297.11 ± 3.915 B | 3.138 ± 0.033 | |||
| GG(134) | 9547.89 ± 132.15 AB | 342.17 ± 5.564 A | 3.628 ± 0.054 a | 300.45 ± 4.056 AB | 3.152 ± 0.034 | |||
| p value | 0.0025 ** | <0.0001 ** | 0.0465 * | <0.0001 ** | 0.6168 | |||
| g.73519434G>A | 1 | AA(15) | 7710.59 ± 205.32 C | 275.39 ± 8.024 B | 3.583 ± 0.081 AB | 249.02 ± 5.858 b | 3.188 ± 0.045 AB | |
| AG(232) | 8428.7 ± 116.43 A | 299.28 ± 4.703 B | 3.533 ± 0.047 B | 263.13 ± 3.431 a | 3.123 ± 0.027 B | |||
| GG(869) | 8234.17 ± 111.35 B | 303.25 ± 4.517 A | 3.686 ± 0.045 A | 261.29 ± 3.295 a | 3.173 ± 0.026 A | |||
| p value | <0.0001 ** | <0.0001 ** | <0.0001 ** | 0.0129 * | <0.0001 ** | |||
| 2 | AA(8) | 9177.3 ± 271.61 AB | 380.34 ± 10.81 A | 4.259 ± 0.108 A | 278.06 ± 7.888 B | 3.029 ± 0.062 B | ||
| AG(135) | 9272.43 ± 131.41 B | 321.17 ± 5.527 C | 3.517 ± 0.053 C | 290.74 ± 4.028 B | 3.155 ± 0.033 AB | |||
| GG(469) | 9481.65 ± 125.07 A | 346.1 ± 5.312 B | 3.69 ± 0.051 B | 300.95 ± 3.871 A | 3.178 ± 0.032 A | |||
| p value | 0.0061 ** | <0.0001 ** | <0.0001 ** | <0.0001 ** | 0.0068 ** |
| Block | Lactation | Haplotype Combination | Milk Yield, kg | Fat Yield, kg | Fat Percentage, % | Protein Yield, kg | Protein Percentage, % |
|---|---|---|---|---|---|---|---|
| ACADVL | 1 | H1H1(192) | 8235.7 ± 61.825 B | 293.51 ± 5.492 Bb | 3.663 ± 0.054 Aa | 251.72 ± 4.005 B | 3.122 ± 0.032 |
| H1H3(155) | 8492.22 ± 66.286 A | 295.75 ± 5.484 ABb | 3.561 ± 0.054 Bb | 258.89 ± 4 A | 3.115 ± 0.032 | ||
| H1H4(131) | 8352.11 ± 68.215 AB | 294.85 ± 5.521 ABb | 3.6 ± 0.054 ABab | 255.12 ± 4.027 AB | 3.127 ± 0.032 | ||
| H1H5(48) | 8348.01 ± 95.774 AB | 296.47 ± 6.265 ABab | 3.626 ± 0.062 ABab | 257.62 ± 4.571 AB | 3.154 ± 0.036 | ||
| H2H2(66) | 8540 ± 87.169 A | 306.78 ± 5.961 Aa | 3.676 ± 0.059 ABa | 261.82 ± 4.348 A | 3.114 ± 0.035 | ||
| p value | 0.0004 ** | 0.0125 * | 0.0031 ** | 0.0007 ** | 0.5268 | ||
| 2 | H1H1(90) | 9383.29 ± 80.757 B | 366.84 ± 6.781 Bb | 3.89 ± 0.065 B | 312.3 ± 4.942 Bbc | 3.268 ± 0.041 | |
| H1H3(76) | 9727.49 ± 90.196 A | 394.13 ± 7.159 Aa | 4.067 ± 0.069 A | 322.87 ± 5.218 Aa | 3.259 ± 0.043 | ||
| H1H4(68) | 9318.77 ± 91.263 B | 382.73 ± 7.046 Aa | 4.074 ± 0.068 A | 311.2 ± 5.135 Bbc | 3.297 ± 0.043 | ||
| H1H5(27) | 9650.12 ± 129.36 AB | 394.51 ± 8.449 Aa | 4.05 ± 0.082 AB | 324.37 ± 6.161 ABac | 3.289 ± 0.05 | ||
| H2H2(38) | 9635.5 ± 113.85 AB | 382.3 ± 7.618 ABa | 3.951 ± 0.074 AB | 326.06 ± 5.554 Aa | 3.319 ± 0.045 | ||
| p value | 0.000 ** | <0.0001 ** | <0.0001** | <0.0001 ** | 0.257 | ||
| IRF6-1 | 1 | H1H1(276) | 8264.24 ± 63.688 ACa | 321.07 ± 5.247 A | 3.84 ± 0.052 BC | 266.8 ± 3.827 ACac | 3.188 ± 0.031 A |
| H1H3(156) | 8075.66 ± 65.82 ABb | 316.55 ± 5.245 A | 3.87 ± 0.052 AC | 260.67 ± 3.826 Ad | 3.186 ± 0.031 A | ||
| H1H4(125) | 8322.49 ± 75.374 Aa | 321.73 ± 5.605 A | 3.8 ± 0.056 ABC | 268.6 ± 4.089 Cc | 3.179 ± 0.032 A | ||
| H2H2(78) | 8158.7 ± 84.982 ABCab | 322.14 ± 5.684 A | 3.906 ± 0.056 A | 264.01 ± 4.147 ACacd | 3.201 ± 0.033 A | ||
| H2H4(65) | 7922.56 ± 89.565 Bb | 297.97 ± 5.929 B | 3.691 ± 0.059 B | 250.17 ± 4.326 Bb | 3.103 ± 0.034 B | ||
| p value | <0.0001 ** | <0.0001 ** | <0.0001 ** | <0.0001 ** | 0.0004 ** | ||
| 2 | H1H1(130) | 9827.31 ± 78.36 Aa | 374.79 ± 6.39 A | 3.877 ± 0.061 ABa | 315.65 ± 4.658 Aa | 3.247 ± 0.039 A | |
| H1H3(84) | 9559.73 ± 80.128 ABbc | 352.78 ± 6.44 B | 3.751 ± 0.062 Bb | 297.81 ± 4.694 Bb | 3.156 ± 0.039 B | ||
| H1H4(70) | 9357.05 ± 90.068 BCc | 349 ± 6.668 B | 3.808 ± 0.064 ABab | 295.94 ± 4.861 BCbc | 3.207 ± 0.04 AB | ||
| H2H2(69) | 9791.05 ± 92.607 Aab | 380.57 ± 6.88 A | 3.916 ± 0.067 Aa | 313.28 ± 5.015 Aa | 3.241 ± 0.041 A | ||
| H2H4(39) | 8972.54 ± 112.15 Cd | 345.57 ± 7.327 B | 3.892 ± 0.071 ABab | 283.41 ± 5.343 Cd | 3.193 ± 0.043 AB | ||
| p value | <0.0001 ** | <0.0001 ** | 0.0018 ** | <0.0001* * | 0.0007 ** | ||
| IRF6-2 | 1 | H1H1(372) | 8173.41 ± 56.522 | 298.45 ± 4.67 Bc | 3.64 ± 0.046 Bb | 262.72 ± 3.407 a | 3.219 ± 0.027 Aa |
| H1H2(484) | 8098.25 ± 52.078 | 293.75 ± 4.565 Bb | 3.619 ± 0.045 BCb | 258.97 ± 3.33 b | 3.205 ± 0.027 ABa | ||
| H1H3(73) | 8025.17 ± 84.435 | 309.74 ± 5.333 Aa | 3.887 ± 0.053 Aa | 261.11 ± 3.892 ab | 3.244 ± 0.031 Aa | ||
| H2H2(139) | 8250.48 ± 66.859 | 295.07 ± 4.967 Bbc | 3.542 ± 0.049 Cc | 261.96 ± 3.624 ab | 3.167 ± 0.029 Bb | ||
| p value | 0.0118 * | <0.0001 ** | <0.0001 ** | 0.0263 * | 0.0003 ** | ||
| 2 | H1H1(183) | 9580.26 ± 66.324 Aa | 369.98 ± 5.608 Ac | 3.851 ± 0.054 BCbc | 305.81 ± 4.088 A | 3.172 ± 0.034 | |
| H1H2(249) | 9378.78 ± 59.293 BCb | 355.61 ± 5.465 Bb | 3.782 ± 0.053 Cc | 297.64 ± 3.983 B | 3.156 ± 0.033 | ||
| H1H3(44) | 9123.09 ± 105.82 Cb | 371.19 ± 6.736 Aac | 4.079 ± 0.066 Aa | 294.72 ± 4.913 B | 3.208 ± 0.04 | ||
| H2H2(92) | 9608.01 ± 78.942 ABa | 380.66 ± 5.933 Aa | 3.936 ± 0.057 ABb | 306.66 ± 4.325 A | 3.171 ± 0.035 | ||
| p value | <0.0001 ** | <0.0001 ** | <0.0001 ** | <0.0001 ** | 0.199 |
| Gene | SNPs | Allele | Transcription | Relative Score (≥0.90) | Predicted Binding Site Sequence |
|---|---|---|---|---|---|
| ACADVL | 19:g.26933503T>C | T | ZEB1 | 0.91 | CACCTT |
| C | |||||
| IRF6 | 16:g.73501985G>A | G | PLAGL2 | 0.98 | TGGGCCCCCC |
| A | RHOXF1 | 0.97 | CTGAGCCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, P.; Liu, Y.; Zheng, W.; Han, B.; Wang, K.; Sun, D. Identification of Genetic Effects of ACADVL and IRF6 Genes with Milk Production Traits of Holstein Cattle in China. Genes 2022, 13, 2393. https://doi.org/10.3390/genes13122393
Peng P, Liu Y, Zheng W, Han B, Wang K, Sun D. Identification of Genetic Effects of ACADVL and IRF6 Genes with Milk Production Traits of Holstein Cattle in China. Genes. 2022; 13(12):2393. https://doi.org/10.3390/genes13122393
Chicago/Turabian StylePeng, Peng, Yanan Liu, Weijie Zheng, Bo Han, Kun Wang, and Dongxiao Sun. 2022. "Identification of Genetic Effects of ACADVL and IRF6 Genes with Milk Production Traits of Holstein Cattle in China" Genes 13, no. 12: 2393. https://doi.org/10.3390/genes13122393
APA StylePeng, P., Liu, Y., Zheng, W., Han, B., Wang, K., & Sun, D. (2022). Identification of Genetic Effects of ACADVL and IRF6 Genes with Milk Production Traits of Holstein Cattle in China. Genes, 13(12), 2393. https://doi.org/10.3390/genes13122393