
Extracellular Vesicle MicroRNA in Malignant Pleural Effusion

 ,
,  ,
, 


Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Population
2.2. Data and Sample Collection and Analysis
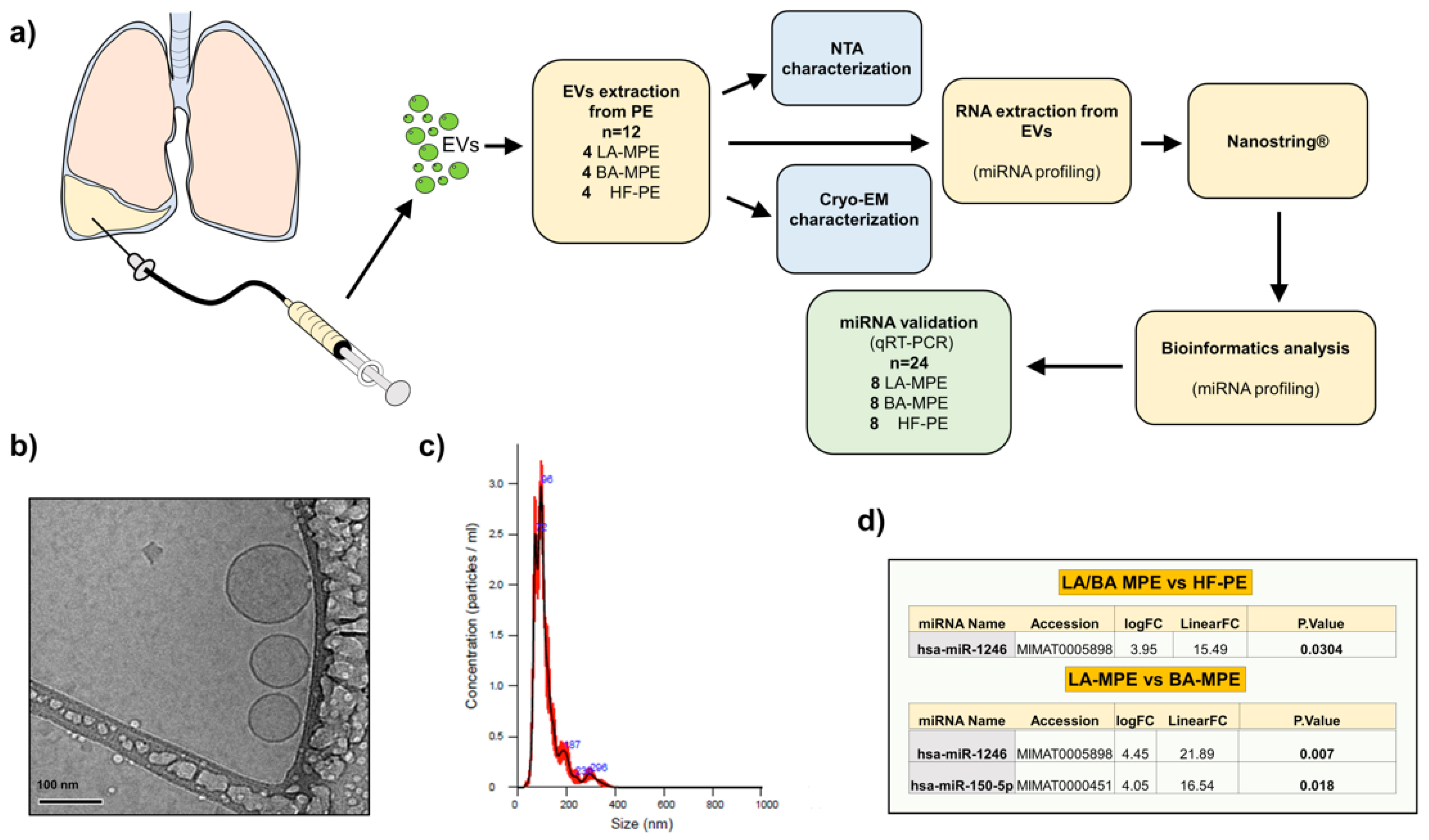
2.3. EVs Isolation
2.4. RNA Extraction
2.5. Nanoparticle Tracking Analysis
2.6. NanoString nCounter Assay
2.7. NanoString Data Analysis
2.8. RT-qPCR Analysis
2.9. Electron Microscopy
2.10. Target Prediction Analysis
2.11. Statistical Analysis
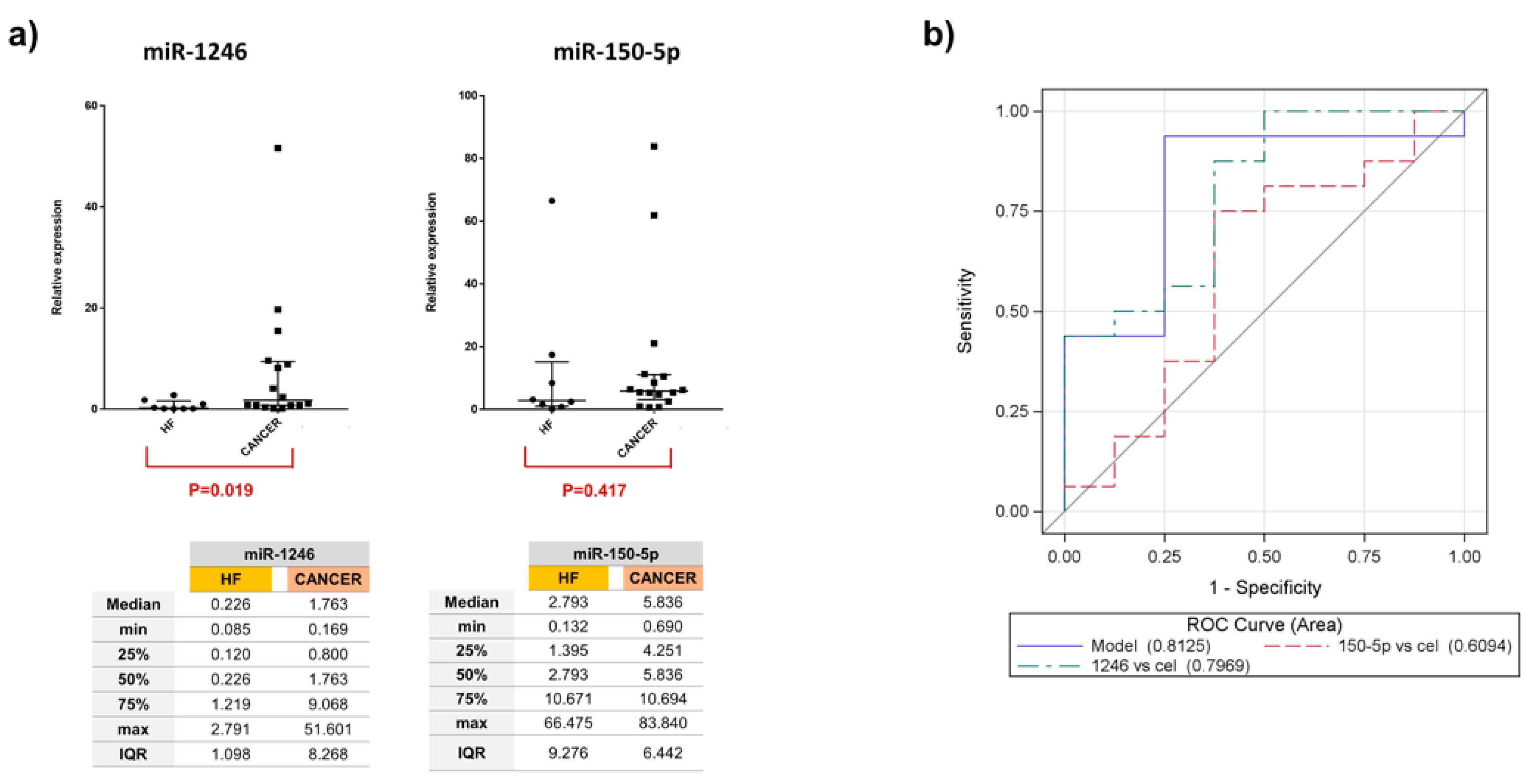
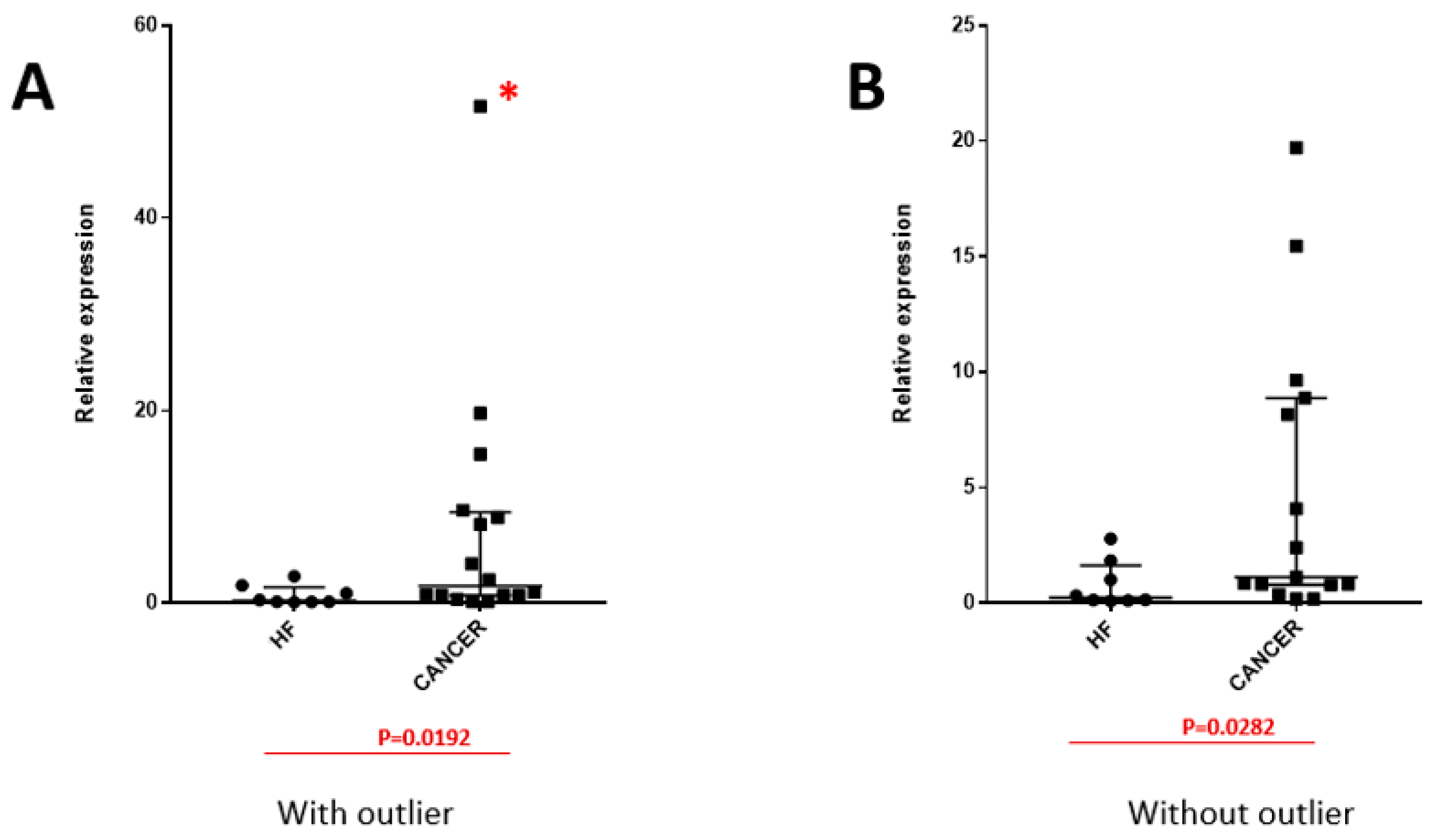
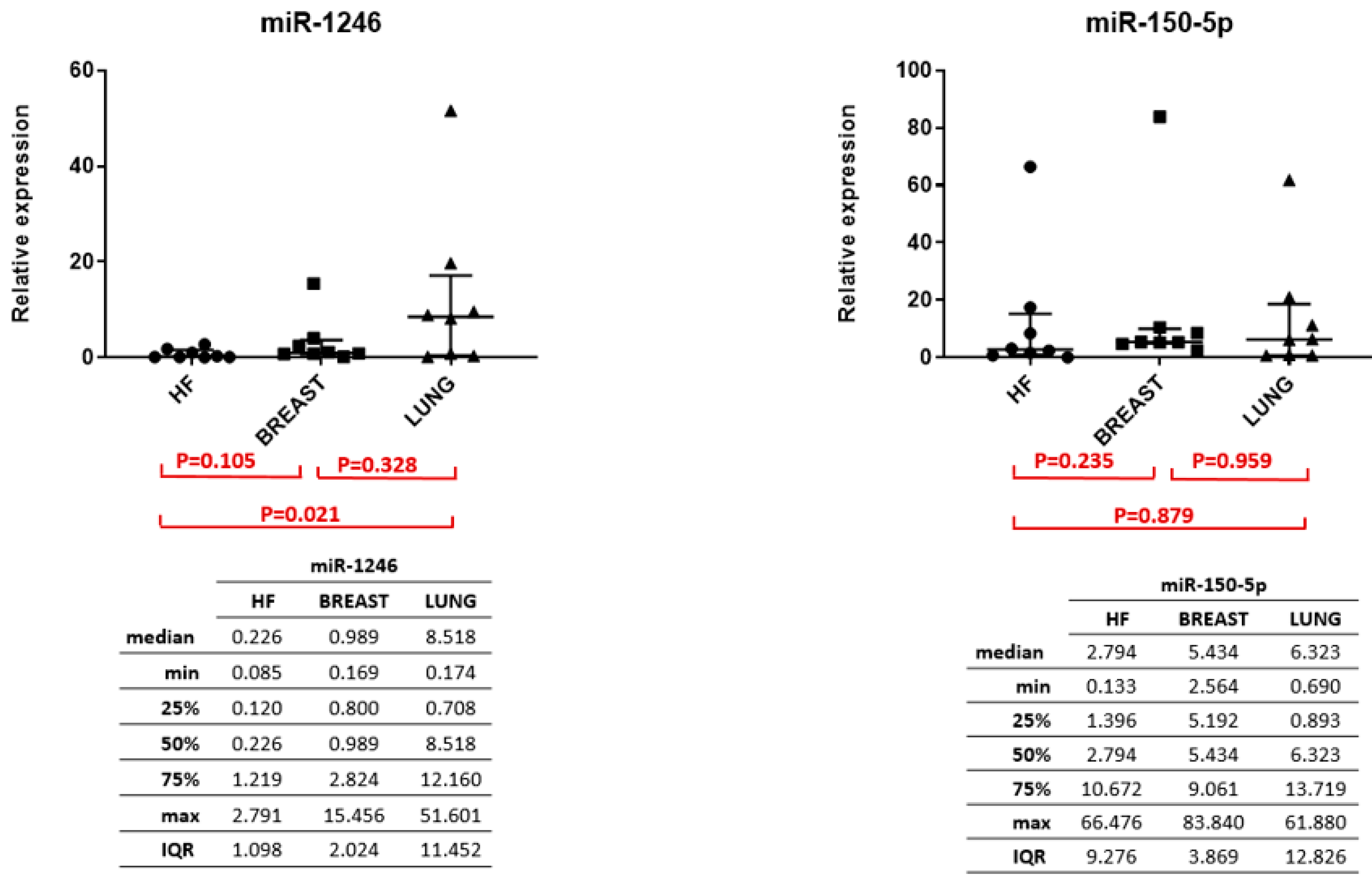
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MPE | Malignant pleural effusion |
| NMPE | Non-malignant pleural effusion |
| miRNAs | microRNAs |
| EVs | Extracellular vesicles |
| HF-PE | Heart-failure-induced effusions |
| LA-MPE | MPE due to lung adenocarcinoma |
| BA-MPE | MPE due to breast adenocarcinoma |
| SD | Standard deviation |
| IQR | Interquartile range |
| ROC | Receiver operating characteristic |
| AUC | Area under the curve |
| qRT-PCR | Quantitative Reverse Transcription Polymerase Chain Reaction |
| CI | Confidence interval |
| FNA | Fine-needle aspiration |
| NSCLC | Non-small-cell lung cancer |
Appendix A. Aspiration Procedures


References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef]
- Shojaee, S.; Roy-Chowdhuri, S.; Safi, J.; Grosu, H. Cytologic Investigations for the Diagnosis of Malignant Pleural Effusion in Non–small Cell Lung Cancer: State-of-the-art Review for Pulmonologists. J. Bronchol. Interv. Pulmonol. 2021; ahead of print. [Google Scholar] [CrossRef]
- Barger, J.F.; Rahman, M.A.; Jackson, D.; Acunzo, M.; Nana-Sinkam, S.P. Extracellular miRNAs as biomarkers in cancer. Food Chem. Toxicol. 2016, 98, 66–72. [Google Scholar] [CrossRef]
- Andre, F.; Schartz, N.E.; Movassagh, M.; Flament, C.; Pautier, P.; Morice, P.; Pomel, C.; Lhomme, C.; Escudier, B.; Le Chevalier, T.; et al. Malignant effusions and immunogenic tumour-derived exosomes. Lancet Lond Engl. 2002, 360, 295–305. [Google Scholar] [CrossRef]
- Saviana, M.; Romano, G.; Le, P.; Acunzo, M.; Nana-Sinkam, P. Extracellular Vesicles in Lung Cancer Metastasis and Their Clinical Applications. Cancers 2021, 13, 5633. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; El Andaloussi, S.; Wood, M.J.A. Exosomes and microvesicles: Extracellular vesicles for genetic information transfer and gene therapy. Hum. Mol. Genet. 2012, 21, R125–R134. [Google Scholar] [CrossRef] [PubMed]
- Hydbring, P.; De Petris, L.; Zhang, Y.; Brandén, E.; Koyi, H.; Novak, M.; Kanter, L.; Hååg, P.; Hurley, J.; Tadigotla, V.; et al. Exosomal RNA-profiling of pleural effusions identifies adenocarcinoma patients through elevated miR-200 and LCN2 expression. Lung Cancer 2018, 124, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Roman-Canal, B.; Moiola, C.P.; Gatius, S.; Bonnin, S.; Ruiz-Miró, M.; González, E.; Ojanguren, A.; Recuero, J.L.; Gil-Moreno, A.; Falcón-Pérez, J.M.; et al. EV-associated miRNAs from pleural lavage as potential diagnostic biomarkers in lung cancer. Sci. Rep. 2019, 9, 15057. [Google Scholar] [CrossRef] [PubMed]
- Romano, G.; Nigita, G.; Calore, F.; Saviana, M.; Le, P.; Croce, C.M.; Acunzo, M.; Nana-Sinkam, P. MiR-124a Regulates Extracellular Vesicle Release by Targeting GTPase Rabs in Lung Cancer. Front. Oncol. 2020, 10, 1454. Available online: https://www.frontiersin.org/articles/10.3389/fonc.2020.01454 (accessed on 28 September 2022). [CrossRef]
- McDonald, K. Handbook of Cryo-Preparation Methods for Electron Microscopy. Annie Cavalier, Danielle Spehner, and Bruno M. Humbel (Eds.). CRC Press, Boca Raton, FL; 2009, 682 pages. ISBN 978-0-8493-7227-8. Microsc. Microanal. 2009, 15, 469–470. [Google Scholar] [CrossRef]
- Gao, M.; Kim, Y.K.; Zhang, C.; Borshch, V.; Zhou, S.; Park, H.S.; Jákli, A.; Lavrentovich, O.D.; Tamba, M.G.; Kohlmeier, A.; et al. Direct observation of liquid crystals using cryo-TEM: Specimen preparation and low-dose imaging. Microsc. Res. Tech. 2014, 77, 754–772. [Google Scholar] [CrossRef]
- Distefano, R.; Nigita, G.; Veneziano, D.; Romano, G.; Croce, C.M.; Acunzo, M. isoTar: Consensus Target Prediction with Enrichment Analysis for MicroRNAs Harboring Editing Sites and Other Variations. Methods Mol. Biol. Clifton NJ 2019, 1970, 211–235. [Google Scholar] [CrossRef]
- Vejnar, C.E.; Zdobnov, E.M. MiRmap: Comprehensive prediction of microRNA target repression strength. Nucleic Acids Res. 2012, 40, 11673–11683. [Google Scholar] [CrossRef]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Kertesz, M.; Iovino, N.; Unnerstall, U.; Gaul, U.; Segal, E. The role of site accessibility in microRNA target recognition. Nat. Genet. 2007, 39, 1278–1284. [Google Scholar] [CrossRef]
- Krüger, J.; Rehmsmeier, M. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34, W451–W454. [Google Scholar] [CrossRef] [PubMed]
- John, B.; Enright, A.J.; Aravin, A.; Tuschl, T.; Sander, C.; Marks, D.S. Human MicroRNA Targets. PLoS Biol. 2004, 2, e363. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Wei, Y.K.; Kaliamurthi, S.; Cao, Y.; Nangraj, A.S.; Sui, X.; Chu, D.; Wang, H.; Wei, D.Q.; Peslherbe, G.H.; et al. Circulating miR-1246 Targeting UBE2C, TNNI3, TRAIP, UCHL1 Genes and Key Pathways as a Potential Biomarker for Lung Adenocarcinoma: Integrated Biological Network Analysis. J. Pers. Med. 2020, 10, 162. [Google Scholar] [CrossRef]
- Cui, X.; Li, Z.; Zhao, Y.; Song, A.; Shi, Y.; Hai, X.; Zhu, W. Breast cancer identification via modeling of peripherally circulating miRNAs. PeerJ 2018, 6, e4551. [Google Scholar] [CrossRef]
- Jang, J.Y.; Kim, Y.S.; Kang, K.N.; Kim, K.H.; Park, Y.J.; Kim, C.W. Multiple microRNAs as biomarkers for early breast cancer diagnosis. Mol. Clin. Oncol. 2021, 14, 31. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Qu, D. Early diagnostic and prognostic value of serum exosomal miR-1246 in non-small cell lung cancer. Int. J. Clin. Exp. Pathol. 2020, 13, 1601–1607. [Google Scholar] [PubMed]
- Huang, S.; Luo, S.; Gong, C.; Liang, L.; Xiao, Y.; Li, M.; He, J. MTTL3 upregulates microRNA-1246 to promote occurrence and progression of NSCLC via targeting paternally expressed gene 3. Mol. Ther. Nucleic Acids 2021, 24, 542–553. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Hou, D.; Liang, H.; Gong, F.; Wang, Y.; Yan, X.; Jiang, X.; Wang, C.; Zhang, J.; Zen, K.; et al. miR-150 promotes the proliferation and migration of lung cancer cells by targeting SRC kinase signalling inhibitor 1. Eur. J. Cancer 2014, 50, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Dai, F.Q.; Li, C.R.; Fan, X.Q.; Tan, L.; Wang, R.T.; Jin, H. miR-150-5p Inhibits Non-Small-Cell Lung Cancer Metastasis and Recurrence by Targeting HMGA2 and β-Catenin Signaling. Mol. Ther. Nucleic Acids 2019, 16, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Misono, S.; Seki, N.; Mizuno, K.; Yamada, Y.; Uchida, A.; Sanada, H.; Moriya, S.; Kikkawa, N.; Kumamoto, T.; Suetsugu, T.; et al. Molecular Pathogenesis of Gene Regulation by the miR-150 Duplex: miR-150-3p Regulates TNS4 in Lung Adenocarcinoma. Cancers 2019, 11, 601. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Malignant (16) | Non-Malignant (8) | ||
|---|---|---|---|---|
| Age | Mean (SD) | 61.8 (12.3) | 76.4 (7.3) | |
| Sex | ||||
| Males | 3 (19%) | 4 (50%) | ||
| Females | 13 (81%) | 4 (50%) | ||
| Race | ||||
| White | 12 (75%) | 4 (50%) | ||
| Black | 4 (25%) | 4 (50%) | ||
| Smoking Status | ||||
| Former | 6 (37%) | 4 (50%) | ||
| Never | 7 (44%) | 4 (50%) | ||
| Current | 3 (19%) | 0 (0%) | ||
| ECOG | ||||
| 0 | 1 (6%) | 0 (0%) | ||
| 1 | 6 (37%) | 2 (25%) | ||
| 2 | 5 (31%) | 3 (37%) | ||
| 3 | 3 (19%) | 3 (37%) | ||
| 4 | 1 (6%) | 0 (0%) | ||
| Etiology | ||||
| Lung Adeno. | 8 (50%) | CHF | 8 (100%) | |
| Breast Adeno | 8 (50%) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shojaee, S.; Romano, G.; Sanchez, T.M.; Yermakhanova, G.; Saviana, M.; Le, P.; Nigita, G.; Calore, F.; Guthrie, R.; Hess, K.; et al. Extracellular Vesicle MicroRNA in Malignant Pleural Effusion. Genes 2022, 13, 2159. https://doi.org/10.3390/genes13112159
Shojaee S, Romano G, Sanchez TM, Yermakhanova G, Saviana M, Le P, Nigita G, Calore F, Guthrie R, Hess K, et al. Extracellular Vesicle MicroRNA in Malignant Pleural Effusion. Genes. 2022; 13(11):2159. https://doi.org/10.3390/genes13112159
Chicago/Turabian StyleShojaee, Samira, Giulia Romano, Trinidad M. Sanchez, Gulmira Yermakhanova, Michela Saviana, Patricia Le, Giovanni Nigita, Federica Calore, Rachel Guthrie, Kathryn Hess, and et al. 2022. "Extracellular Vesicle MicroRNA in Malignant Pleural Effusion" Genes 13, no. 11: 2159. https://doi.org/10.3390/genes13112159
APA StyleShojaee, S., Romano, G., Sanchez, T. M., Yermakhanova, G., Saviana, M., Le, P., Nigita, G., Calore, F., Guthrie, R., Hess, K., Kang, L., Swift-Scanlan, T., Graham, J. T., Rahman, N. M., Nana-Sinkam, P. S., & Acunzo, M. (2022). Extracellular Vesicle MicroRNA in Malignant Pleural Effusion. Genes, 13(11), 2159. https://doi.org/10.3390/genes13112159