

Ancient DNA Reveals Maternal Philopatry of the Northeast Eurasian Brown Bear (Ursus arctos) Population during the Holocene

 , , ,
, , ,  , , and
, , and 
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Brown Bear Specimen Description and 14C Dating
2.2. Ancient DNA Extraction, Library Preparation and DNA Sequencing
- Ancient DNA extraction using proteinase K in buffer containing EDTA.
- Ancient DNA enrichment on silica beads in binding buffer (contains tris(hydroxymethyl)aminomethane (TRIS), sodium acetate guanidine thiocyanate, and sodium chloride).
- Ancient DNA washing using ethanol.
- Ancient DNA elution in low-salt buffer.
2.3. Bioinformatics Analysis
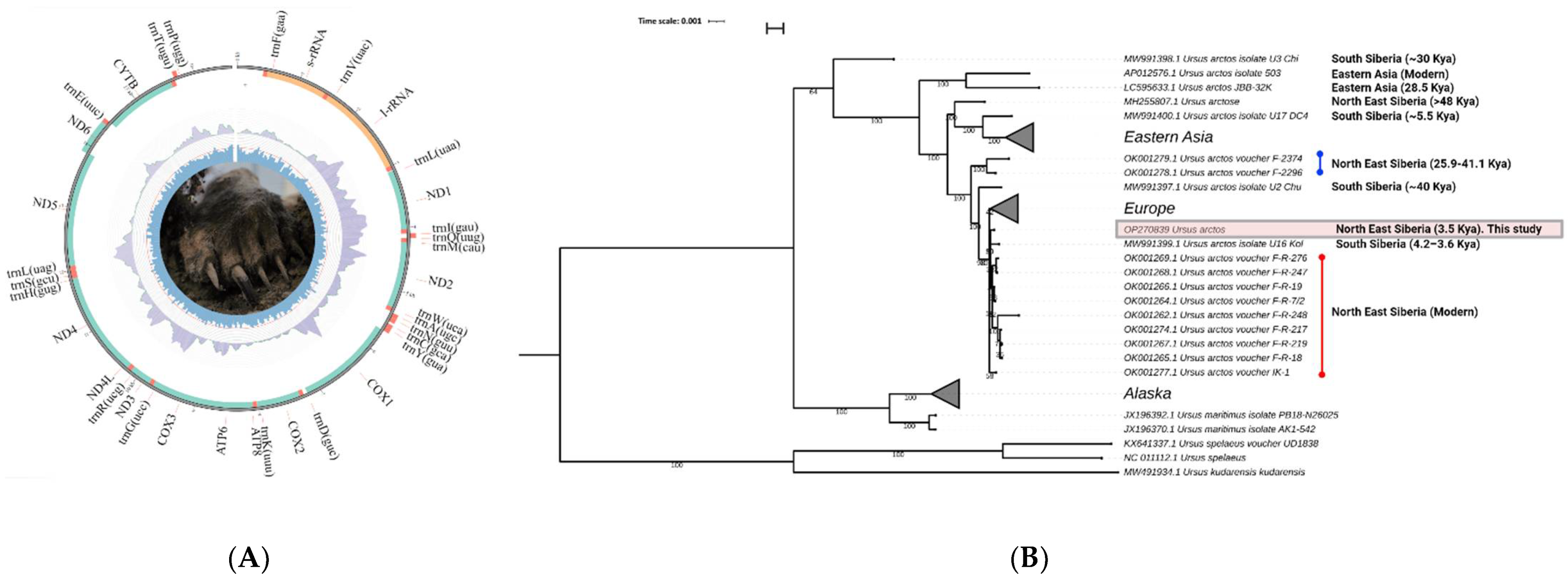
3. Results
3.1. AMS Radiocarbon Dating
3.2. Ancient DNA Sequencing and Brown Bear Mitogenome Assembly
3.3. Phylogenetic Analysis of Holocene Specimen and Other Ancient and Modern Brown Bears from Eurasia and North America Based on Their Mitochondrial Sequences
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chestin, I.E.; Gubar, Y.P.; Sokolov, V.E.; Lobachev, V.S. The brown bear (Ursus arctos L.) in the USSR: Numbers, hunting and systematics. Ann. Zool. Fenn. 1992, 29, 57–68. [Google Scholar]
- Boeskorov, G.G.; Baryshnikov, G.F.; Tikhonov, A.N.; Protopopov, A.V.; Klimovsky, A.I.; Grigoriev, S.E.; Cheprasov, M.Y.; Novgorodov, G.P.; Shchelchkova, M.V.; van der Plicht, J. New Data on Large Brown Bear (Ursus arctos L., 1758, Ursidae, Carnivora, Mammalia) from the Pleistocene in Yakutia. Dokl. Earth Sci. 2019, 486, 617–622. [Google Scholar] [CrossRef]
- Kosintsev, P.A.; Bocherens, H.; Kirillova, I.V.; Levchenko, V.A.; Zazovskaya, E.P.; Trofimova, S.S.; Lan, T.; Lindqvist, C. Palaeoecological and genetic analyses of Late Pleistocene bears in Asiatic Russia. Boreas 2022, 51, 465–480. [Google Scholar] [CrossRef]
- Davison, J.; Ho, S.Y.W.; Bray, S.C.; Korsten, M.; Tammeleht, E.; Hindrikson, M.; Østbye, K.; Østbye, E.; Lauritzen, S.-E.; Austin, J.; et al. Late-Quaternary biogeographic scenarios for the brown bear (Ursus arctos), a wild mammal model species. Quat. Sci. Rev. 2011, 30, 418–430. [Google Scholar] [CrossRef]
- Hirata, D.; Mano, T.; Abramov, A.V.; Baryshnikov, G.F.; Kosintsev, P.A.; Vorobiev, A.A.; Raichev, E.G.; Tsunoda, H.; Kaneko, Y.; Murata, K.; et al. Molecular phylogeography of the brown bear (Ursus arctos) in Northeastern Asia based on analyses of complete mitochondrial DNA sequences. Mol. Biol. Evol. 2013, 30, 1644–1652. [Google Scholar] [CrossRef]
- Molodtseva, A.S.; Makunin, A.I.; Salomashkina, V.V.; Kichigin, I.G.; Vorobieva, N.V.; Vasiliev, S.K.; Shunkov, M.V.; Tishkin, A.A.; Grushin, S.P.; Anijalg, P.; et al. Phylogeography of ancient and modern brown bears from eastern Eurasia. Biol. J. Linn. Soc. 2022, 135, 722–733. [Google Scholar] [CrossRef]
- Rey-Iglesia, A.; Garcia-Vazquez, A.; Treadaway, E.C.; van der Plicht, J.; Baryshnikov, G.F.; Szpak, P.; Bocherens, H.; Boeskorov, G.G.; Lorenzen, E.D. Evolutionary history and palaeoecology of brown bear in North-East Siberia re-examined using ancient DNA and stable isotopes from skeletal remains. Sci. Rep. 2019, 9, 4462. [Google Scholar] [CrossRef]
- Boeskorov, G.; Baryshnikov, G. Late Quaternary Carnivora of Yakutia. St.-Petersburg Nauka 2013, 210. [Google Scholar]
- Sharko, F.S.; Boulygina, E.S.; Tsygankova, S.V.; Slobodova, N.V.; Alekseev, D.A.; Krasivskaya, A.A.; Rastorguev, S.M.; Tikhonov, A.N.; Nedoluzhko, A.V. Steller’s sea cow genome suggests this species began going extinct before the arrival of Paleolithic humans. Nat. Commun. 2021, 12, 2215. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Pedersen, M.W.; Alsos, I.G.; De Sanctis, B.; Racimo, F.; Prohaska, A.; Coissac, E.; Owens, H.L.; Merkel, M.K.F.; Fernandez-Guerra, A.; et al. Late Quaternary dynamics of Arctic biota from ancient environmental genomics. Nature 2021, 600, 86–92. [Google Scholar] [CrossRef]
- Seryodkin, I.V. Problems in the “brown bear–salmon–human” relationship on the Pacific coast of Russia. Int. Res. J. 2017, 12, 49–51. [Google Scholar] [CrossRef]
- Stroganov, S.U. Carnivorous Mammals of Siberia; USSR Academy of Sciences: Moscow, Russia, 1962; p. 458. [Google Scholar]
- Orlando, L.; Ginolhac, A.; Raghavan, M.; Vilstrup, J.; Rasmussen, M.; Magnussen, K.; Steinmann, K.E.; Kapranov, P.; Thompson, J.F.; Zazula, G.; et al. True single-molecule DNA sequencing of a pleistocene horse bone. Genome Res. 2011, 21, 1705–1719. [Google Scholar] [CrossRef] [PubMed]
- Boulygina, E.; Tsygankova, S.; Sharko, F.; Slobodova, N.; Gruzdeva, N.; Rastorguev, S.; Belinsky, A.; Harke, H.; Kadieva, A.; Demidenko, S.; et al. Mitochondrial and Y-chromosome diversity of the prehistoric Koban culture of the North Caucasus. J. Archaeol. Sci. Rep. 2020, 31, 102357. [Google Scholar] [CrossRef]
- Schubert, M.; Lindgreen, S.; Orlando, L. AdapterRemoval v2: Rapid adapter trimming, identification, and read merging. BMC Res. Notes 2016, 9, 88. [Google Scholar] [CrossRef]
- Schubert, M.; Ermini, L.; Der Sarkissian, C.; Jonsson, H.; Ginolhac, A.; Schaefer, R.; Martin, M.D.; Fernandez, R.; Kircher, M.; McCue, M.; et al. Characterization of ancient and modern genomes by SNP detection and phylogenomic and metagenomic analysis using PALEOMIX. Nat. Protoc. 2014, 9, 1056–1082. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows—Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Jonsson, H.; Ginolhac, A.; Schubert, M.; Johnson, P.L.; Orlando, L. mapDamage2.0: Fast approximate Bayesian estimates of ancient DNA damage parameters. Bioinformatics 2013, 29, 1682–1684. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Bernt, M.; Donath, A.; Juhling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Putz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenetics Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML Web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef]
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef]
- Sharko, F.S.; Boulygina, E.S.; Rastorguev, S.M.; Tsygankova, S.V.; Tomkovich, P.S.; Nedoluzhko, A.V. Phylogenetic position of the presumably extinct slender-billed curlew, Numenius tenuirostris. Mitochondrial DNA Part A 2019, 30, 626–631. [Google Scholar] [CrossRef]
- Levin, B.; Simonov, E.; Gabrielyan, B.K.; Mayden, R.L.; Rastorguev, S.M.; Roubenyan, H.R.; Sharko, F.S.; Nedoluzhko, A.V. Caucasian treasure: Genomics sheds light on the evolution of half-extinct Sevan trout, Salmo ischchan, species flock. Mol. Phylogenetics Evol. 2022, 167, 107346. [Google Scholar] [CrossRef]
- Barlow, A.; Cahill, J.A.; Hartmann, S.; Theunert, C.; Xenikoudakis, G.; Fortes, G.G.; Paijmans, J.L.A.; Rabeder, G.; Frischauf, C.; Grandal-d’Anglade, A.; et al. Partial genomic survival of cave bears in living brown bears. Nat. Ecol. Evol. 2018, 2, 1563–1570. [Google Scholar] [CrossRef]
- Lan, T.; Leppälä, K.; Tomlin, C.l.; Talbot, S.L.; Sage, G.K.; Farley, S.D.; Shideler, R.T.; Bachmann, L.; Wiig, Ø.; Albert, V.A.; et al. Insights into bear evolution from a Pleistocene polar bear genome. Proc. Natl. Acad. Sci. USA 2022, 119, e2200016119. [Google Scholar] [CrossRef]
- Zou, T.; Kuang, W.; Yin, T.; Frantz, L.; Zhang, C.; Liu, J.; Wu, H.; Yu, L. Uncovering the enigmatic evolution of bears in greater depth: The hybrid origin of the Asiatic black bear. Proc. Natl. Acad. Sci. USA 2022, 119, e2120307119. [Google Scholar] [CrossRef]
- Pitulko, V.V.; Kuzmin, Y.V.; Glascock, M.D.; Pavlova, E.Y.; Grebennikov, A.V. ‘They came from the ends of the earth’: Long-distance exchange of obsidian in the High Arctic during the Early Holocene. Antiquity 2019, 93, 28–44. [Google Scholar] [CrossRef]
- Babiy, U.V.; Salomashkina, V.V.; Kulemeev, P.S.; Kholodova, M.V.; Gruzdev, A.R.; Regehr, E.V. First evidence of a brown bear on Wrangel Island, Russia. Ursus 2022, 2022, 1–8. [Google Scholar] [CrossRef]
- Pongracz, J.D.; Paetkau, D.; Branigan, M.; Richardson, E. Recent Hybridization between a Polar Bear and Grizzly Bears in the Canadian Arctic. Arctic 2017, 70, 151–160. [Google Scholar] [CrossRef]
- Wang, M.S.; Murray, G.G.R.; Mann, D.; Groves, P.; Vershinina, A.O.; Supple, M.A.; Kapp, J.D.; Corbett-Detig, R.; Crump, S.E.; Stirling, I.; et al. A polar bear paleogenome reveals extensive ancient gene flow from polar bears into brown bears. Nat. Ecol. Evol. 2022, 6, 936–944. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.; Schuster, S.C.; Welch, A.J.; Ratan, A.; Bedoya-Reina, O.C.; Zhao, F.; Kim, H.L.; Burhans, R.C.; Drautz, D.I.; Wittekindt, N.E.; et al. Polar and brown bear genomes reveal ancient admixture and demographic footprints of past climate change. Proc. Natl. Acad. Sci. USA 2012, 109, E2382–E2390. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boulygina, E.; Sharko, F.; Cheprasov, M.; Gladysheva-Azgari, M.; Slobodova, N.; Tsygankova, S.; Rastorguev, S.; Grigorieva, L.; Kopp, M.; Fernandes, J.M.O.; et al. Ancient DNA Reveals Maternal Philopatry of the Northeast Eurasian Brown Bear (Ursus arctos) Population during the Holocene. Genes 2022, 13, 1961. https://doi.org/10.3390/genes13111961
Boulygina E, Sharko F, Cheprasov M, Gladysheva-Azgari M, Slobodova N, Tsygankova S, Rastorguev S, Grigorieva L, Kopp M, Fernandes JMO, et al. Ancient DNA Reveals Maternal Philopatry of the Northeast Eurasian Brown Bear (Ursus arctos) Population during the Holocene. Genes. 2022; 13(11):1961. https://doi.org/10.3390/genes13111961
Chicago/Turabian StyleBoulygina, Eugenia, Fedor Sharko, Maksim Cheprasov, Maria Gladysheva-Azgari, Natalia Slobodova, Svetlana Tsygankova, Sergey Rastorguev, Lena Grigorieva, Martina Kopp, Jorge M. O. Fernandes, and et al. 2022. "Ancient DNA Reveals Maternal Philopatry of the Northeast Eurasian Brown Bear (Ursus arctos) Population during the Holocene" Genes 13, no. 11: 1961. https://doi.org/10.3390/genes13111961
APA StyleBoulygina, E., Sharko, F., Cheprasov, M., Gladysheva-Azgari, M., Slobodova, N., Tsygankova, S., Rastorguev, S., Grigorieva, L., Kopp, M., Fernandes, J. M. O., Novgorodov, G., Boeskorov, G., Protopopov, A., Hwang, W.-S., Tikhonov, A., & Nedoluzhko, A. (2022). Ancient DNA Reveals Maternal Philopatry of the Northeast Eurasian Brown Bear (Ursus arctos) Population during the Holocene. Genes, 13(11), 1961. https://doi.org/10.3390/genes13111961