
Exploiting Comparative Omics to Understand the Pathogenic and Virulence-Associated Protease: Anti-Protease Relationships in the Zoonotic Parasites Fasciola hepatica and Fasciola gigantica



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fasciola spp. Databases Used to Isolate/Identify Peptidase and Peptidase Inhibitor Genes
2.2. Identification of the Gene Families Relating to the Cathepsin Peptidases and Peptidase Inhibitors
2.3. F. gigantica Genome Analysis
3. Results
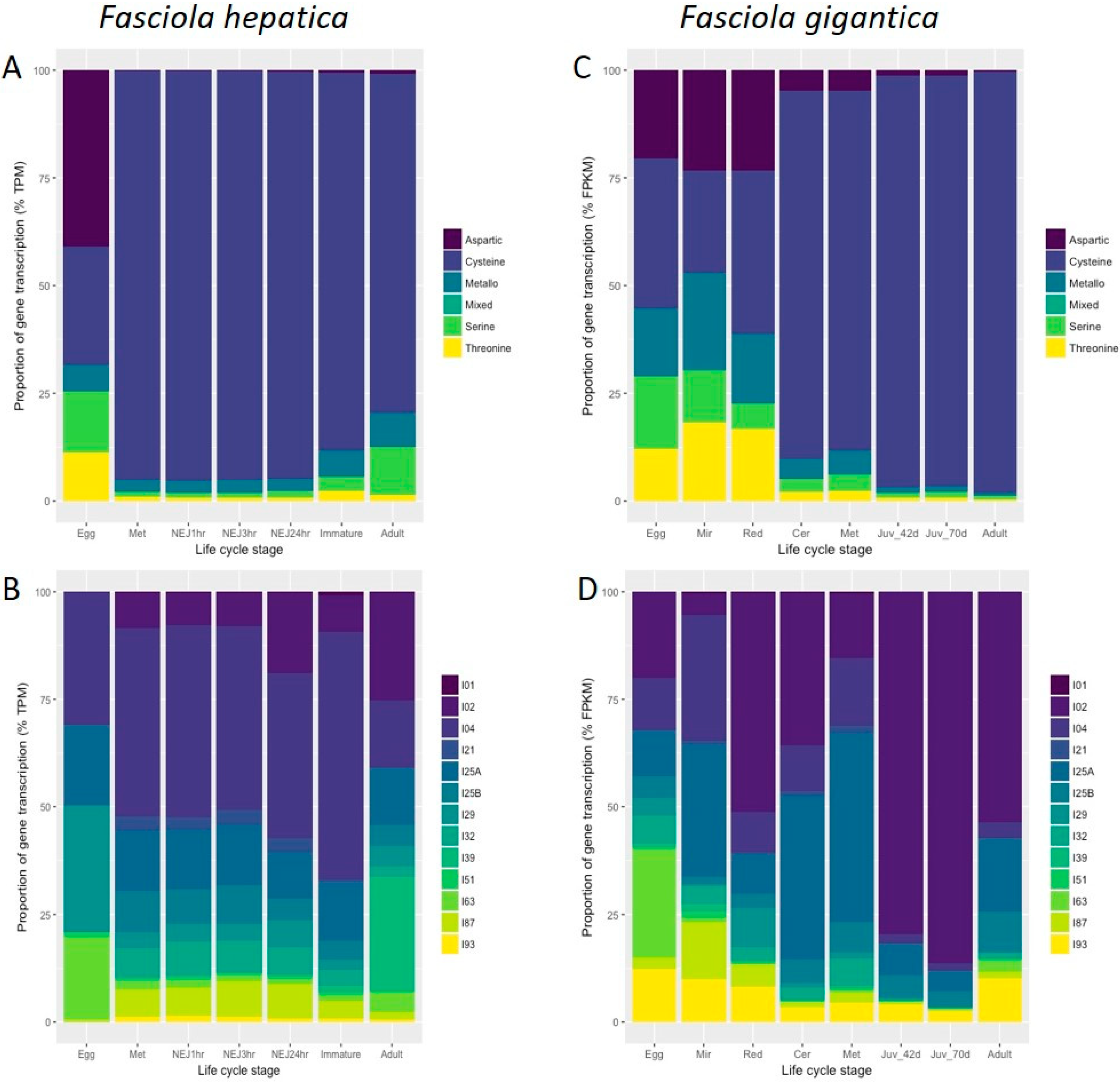
3.1. Peptidases and Peptidase Inhibitors Expressed throughout the Life Cycle of Fasciola spp.
3.2. F. hepatica Cathepsin Cysteine Peptidases
3.3. F. gigantica Cathepsin Cysteine Peptidases
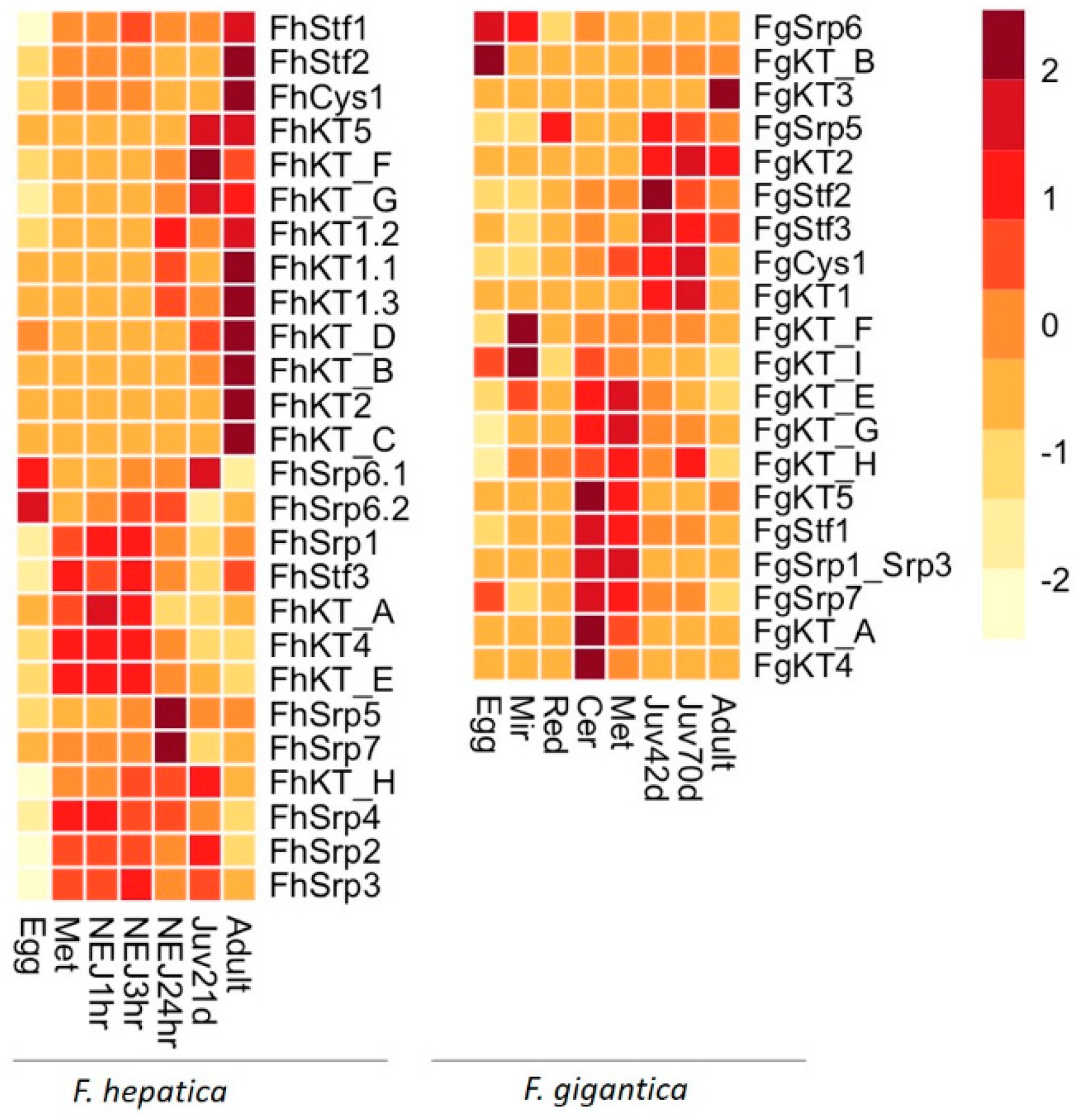
3.4. Key Fasciola spp. Peptidase Inhibitors
3.5. Chromosomal Location of Key F. gigantica Cathepsin Peptidase and Peptidase Inhibitor Genes
3.5.1. Peptidase Gene Families
3.5.2. Peptidase Inhibitor Gene Families
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Andrews, S.J.; Cwiklinski, K.; Dalton, J.P. The discovery of Fasciola hepatica and its life cycle. In Fasciolosis; Dalton, J.P., Ed.; CAB International: Wallingford, UK, 2022. [Google Scholar] [CrossRef]
- Robinson, M.W.; Dalton, J.P. Zoonotic helminth infections with particular emphasis on fasciolosis and other trematodiases. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2763–2776. [Google Scholar] [CrossRef] [PubMed]
- Lalor, R.; Cwiklinski, K.; Calvani, N.E.D.; Dorey, A.; Hamon, S.; López Corrales, J.; Dalton, J.P.; De Marco Verissimo, C. Pathogenicity and virulence of the liver flukes Fasciola hepatica and Fasciola gigantica that cause the zoonosis Fasciolosis. Virulence 2021, 12, 2839–2867. [Google Scholar] [CrossRef] [PubMed]
- Cwiklinski, K.; Donnelly, S.; Drysdale, O.; Jewhurst, H.; Smith, D.; De Marco Verissimo, C.; Pritsch, I.C.; O’Neill, S.; Dalton, J.P.; Robinson, M.W. The cathepsin-like cysteine peptidases of trematodes of the genus Fasciola. Adv. Parasitol. 2019, 104, 113–164. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Cwiklinski, K.; Jewhurst, H.; Tikhonova, I.G.; Dalton, J.P. An atypical and functionally diverse family of Kunitz-type cysteine/serine proteinase inhibitors secreted by the helminth parasite Fasciola hepatica. Sci. Rep. 2020, 10, 20657. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Tikhonova, I.G.; Jewhurst, H.L.; Drysdale, O.C.; Dvořák, J.; Robinson, M.W.; Cwiklinski, K.; Dalton, J.P. Unexpected activity of a novel Kunitz-type inhibitor: Inhibition of cysteine proteases but not serine proteases. J. Biol. Chem. 2016, 291, 19220–19234. [Google Scholar] [CrossRef] [PubMed]
- Cwiklinski, K.; Drysdale, O.; López Corrales, J.; Corripio-Miyar, Y.; De Marco Verissimo, C.; Jewhurst, H.; Smith, D.; Lalor, R.; McNeilly, T.N.; Dalton, J.P. Targeting Secreted Protease/Anti-Protease Balance as a Vaccine Strategy against the Helminth Fasciola hepatica. Vaccines 2022, 10, 155. [Google Scholar] [CrossRef] [PubMed]
- De Marco Verissimo, C.; Jewhurst, H.L.; Dobó, J.; Gál, P.; Dalton, J.P.; Cwiklinski, K. Fasciola hepatica is refractory to complement killing by preventing attachment of mannose binding lectin (MBL) and inhibiting MBL-associated serine proteases (MASPs) with serpins. PLoS Pathog. 2022, 18, e1010226. [Google Scholar] [CrossRef] [PubMed]
- De Marco Verissimo, C.; Jewhurst, H.L.; Tikhonova, I.G.; Urbanus, R.T.; Maule, A.G.; Dalton, J.P.; Cwiklinski, K. Fasciola hepatica serine protease inhibitor family (serpins): Purposely crafted for regulating host proteases. PLoS Negl. Trop. Dis. 2020, 14, e0008510. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.W.; Dalton, J.P.; Donnelly, S. Helminth pathogen cathepsin proteases: It’s a family affair. Trends Biochem. Sci. 2008, 33, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Cancela, M.; Corvo, I.; DA Silva, E.; Teichmann, A.; Roche, L.; Díaz, A.; Tort, J.F.; Ferreira, H.B.; Zaha, A. Functional characterization of single-domain cystatin-like cysteine proteinase inhibitors expressed by the trematode Fasciola hepatica. Parasitology 2017, 144, 1695–1707. [Google Scholar] [CrossRef] [PubMed]
- Cwiklinski, K.; Dalton, J.P.; Dufresne, P.J.; La Course, J.; Williams, D.J.; Hodgkinson, J.; Paterson, S. The Fasciola hepatica genome: Gene duplication and polymorphism reveals adaptation to the host environment and the capacity for rapid evolution. Genome Biol. 2015, 16, 71. [Google Scholar] [CrossRef] [PubMed]
- Cwiklinski, K.; De Marco Verissimo, C.; McVeigh, P.; Donnelly, S.; Dalton, J.P. Applying ‘Omics’ technologies to understand Fasciola spp. biology. In Fasciolosis; Dalton, J.P., Ed.; CAB International: Wallingford, UK, 2022. [Google Scholar] [CrossRef]
- Choi, Y.J.; Fontenla, S.; Fischer, P.U.; Le, T.H.; Costábile, A.; Blair, D.; Brindley, P.J.; Tort, J.F.; Cabada, M.M.; Mitreva, M. Adaptive radiation of the flukes of the Family Fasciolidae inferred from genome-wide comparisons of key species. Mol. Biol. Evol. 2020, 37, 84–99. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Cui, K.; Wang, Z.; Li, Z.; Wu, Z.; Huang, W.; Zhu, X.-Q.; Ruan, J.; Zhang, W.; Liu, Q.; et al. High-quality reference genome of Fasciola gigantica: Insights into the genomic signatures of transposon-mediated evolution and specific parasitic adaption in tropical regions. PLoS Negl. Trop. Dis. 2021, 15, e0009750. [Google Scholar] [CrossRef] [PubMed]
- Geadkaew, A.; Kosa, N.; Siricoon, S.; Grams, S.V.; Grams, R. A 170kDa multi-domain cystatin of Fasciola gigantica is active in the male reproductive system. Mol. Biochem. Parasitol. 2014, 196, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Grams, R.; Vichasri-Grams, S.; Sobhon, P.; Upatham, E.S.; Viyanant, V. Molecular cloning and characterization of cathepsin L encoding genes from Fasciola gigantica. Parasitol. Int. 2001, 50, 105–114. [Google Scholar] [CrossRef]
- Sansri, V.; Changklungmoa, N.; Chaichanasak, P.; Sobhon, P.; Meemon, K. Molecular cloning, characterization and functional analysis of a novel juvenile-specific cathepsin L of Fasciola gigantica. Acta Trop. 2013, 128, 76–84. [Google Scholar] [CrossRef]
- Sethadavit, M.; Meemon, K.; Jardim, A.; Spithill, T.W.; Sobhon, P. Identification, expression and immunolocalization of cathepsin B3, a stage-specific antigen expressed by juvenile Fasciola gigantica. Acta Trop. 2009, 112, 164–173. [Google Scholar] [CrossRef]
- Siricoon, S.; Grams, S.V.; Grams, R. Efficient inhibition of cathepsin B by a secreted type 1 cystatin of Fasciola gigantica. Mol. Biochem. Parasitol. 2012, 186, 126–133. [Google Scholar] [CrossRef]
- Tarasuk, M.; Grams, S.V.; Viyanant, V.; Grams, R. Type I cystatin (stefin) is a major component of Fasciola gigantica excretion/secretion product. Mol. Biochem. Parasitol. 2009, 167, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, H.; Mineki, R.; Murayama, K.; Ito, A.; Aoki, T. Characterisation and expression of the Fasciola gigantica cathepsin L gene. Int. J. Parasitol. 2002, 32, 1031–1042. [Google Scholar] [CrossRef]
- Meemon, K.; Grams, R.; Vichasri-Grams, S.; Hofmann, A.; Korge, G.; Viyanant, V.; Upatham, E.S.; Habe, S.; Sobhon, P. Molecular cloning and analysis of stage and tissue-specific expression of cathepsin B encoding genes from Fasciola gigantica. Mol. Biochem. Parasitol. 2004, 136, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Siricoon, S.; Grams, S.V.; Lertwongvisarn, K.; Abdullohfakeeyah, M.; Smooker, P.M.; Grams, R. Fasciola gigantica cathepsin B5 is an acidic endo- and exopeptidase of the immature and mature parasite. Biochimie 2015, 119, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Young, N.D.; Jex, A.R.; Cantacessi, C.; Hall, R.S.; Campbell, B.E.; Spithill, T.W.; Tangkawattana, S.; Tangkawattana, P.; Laha, T.; Gasser, R.B. A Portrait of the transcriptome of the neglected trematode, Fasciola gigantica—biological and biotechnological implications. PLoS Negl. Trop. Dis. 2011, 5, e1004. [Google Scholar] [CrossRef]
- Zhang, X.X.; Cwiklinski, K.; Hu, R.S.; Zheng, W.B.; Sheng, Z.A.; Zhang, F.K.; Elsheikha, H.M.; Dalton, J.P.; Zhu, X.Q. Complex and dynamic transcriptional changes allow the helminth Fasciola gigantica to adjust to its intermediate snail and definitive mammalian hosts. BMC Genom. 2019, 20, 729. [Google Scholar] [CrossRef]
- Cwiklinski, K.; Jewhurst, H.; McVeigh, P.; Barbour, T.; Maule, A.; Tort, J.F.; O’Neill, S.M.; Robinson, M.; Donnelly, S.; Dalton, J.P. Infection by the helminth parasite Fasciola hepatica requires rapid regulation of metabolic, virulence, and invasive factors to adjust to its mammalian host. Mol. Cell. Proteom. 2018, 17, 792–809. [Google Scholar] [CrossRef] [PubMed]
- Cwiklinski, K.; Robinson, M.W.; Donnelly, S.; Dalton, J.P. Complementary transcriptomic and proteomic analyses reveal the cellular and molecular processes that drive growth and development of Fasciola hepatica in the host liver. BMC Genom. 2021, 22, 46. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.; Cwiklinski, K.; Lalor, R.; O’Connell, B.; Robinson, M.W.; Gerlach, J.; Joshi, L.; Kilcoyne, M.; Dalton, J.P.; O’Neill, S.M. Fasciola hepatica Extracellular Vesicles isolated from excretory-secretory products using a gravity flow method modulate dendritic cell phenotype and activity. PLoS Negl. Trop. Dis. 2020, 14, e0008626. [Google Scholar] [CrossRef]
- Ilgová, J.; Vorel, J.; Roudnický, P.; Škorpíková, L.; Horn, M.; Kašný, M. Transcriptomic and proteomic profiling of peptidase expression in Fasciola hepatica eggs developing at host’s body temperature. Sci. Rep. 2022, 12, 10308. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, D.P. Cysteine peptidases of mammals: Their biological roles and potential effects in the oral cavity and other tissues in health and disease. Crit. Rev. Oral Biol. Med. 2002, 13, 238–275. [Google Scholar] [CrossRef]
- Tort, J.; Brindley, P.J.; Knox, D.; Wolfe, K.H.; Dalton, J.P. Proteinases and sssociated genes of parasitic helminths. Adv. Parasitol. 1999, 43, 161–266. [Google Scholar] [CrossRef] [PubMed]
- Dalton, J.P.; Brindley, P.J.; Donnelly, S.; Robinson, M.W. The enigmatic asparaginyl endopeptidase of helminth parasites. Trends Parasitol. 2009, 25, 59–61. [Google Scholar] [CrossRef] [PubMed]
- Corvo, I.; O’Donoghue, A.J.; Pastro, L.; Pi-Denis, N.; Eroy-Reveles, A.; Roche, L.; McKerrow, J.H.; Dalton, J.P.; Craik, C.S.; Caffrey, C.R.; et al. Dissecting the active site of the collagenolytic cathepsin L3 protease of the invasive stage of Fasciola hepatica. PLoS Negl. Trop. Dis. 2013, 7, e2269. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.W.; Corvo, I.; Jones, P.M.; George, A.M.; Padula, M.P.; To, J.; Cancela, M.; Rinaldi, G.; Tort, J.F.; Roche, L.; et al. Collagenolytic activities of the major secreted cathepsin L peptidases involved in the virulence of the helminth pathogen, Fasciola hepatica. PLoS Negl. Trop. Dis. 2011, 5, e1012. [Google Scholar] [CrossRef]
- Stack, C.M.; Caffrey, C.R.; Donnelly, S.M.; Seshaadri, A.; Lowther, J.; Tort, J.F.; Collins, P.R.; Robinson, M.W.; Xu, W.; McKerrow, J.H.; et al. Structural and functional relationships in the virulence-associated cathepsin L proteases of the parasitic liver fluke, Fasciola hepatica. J. Biol. Chem. 2008, 283, 9896–9908. [Google Scholar] [CrossRef]
- Lowther, J.; Robinson, M.W.; Donnelly, S.M.; Xu, W.; Stack, C.M.; Matthews, J.M.; Dalton, J.P. The importance of pH in regulating the function of the Fasciola hepatica cathepsin L1 cysteine protease. PLoS Negl. Trop. Dis. 2009, 3, e369. [Google Scholar] [CrossRef]
- Mas-Coma, S.; Valero, M.A.; Bargues, M.D. Chapter 2 Fasciola, lymnaeids and human fascioliasis, with a global overview on disease transmission, epidemiology, evolutionary genetics, molecular epidemiology and control. Adv. Parasitol. 2009, 69, 41–146. [Google Scholar] [CrossRef]
- Hussein, A.-N.A.; Hassan, I.M.; Khalifa, R.M. Development and hatching mechanism of Fasciola eggs, light and scanning electron microscopic studies. Saudi J. Biol. Sci. 2010, 17, 247–251. [Google Scholar] [CrossRef]
- Dinnik, J.A.; Dinnik, N.N. The influence of temperature on the succession of redial and cercarial generations of Fasciola gigantica in a snail host. Parasitology 1964, 54, 59–65. [Google Scholar] [CrossRef]
- Kendall, S. Relationships between the species of Fasciola and their molluscan hosts. Adv. Parasitol. 1970, 8, 251–258. [Google Scholar] [CrossRef]
- Dinnik, J.A.; Dinnik, N.N. Observations on the succession of redial generations of Fasciola gigantica Cobbold in a snail host. Z. Fur Trop. Parasitol. 1956, 7, 397–419. [Google Scholar]
- Rondelaud, D.; Belfaiza, M.; Vignoles, P.; Moncef, M.; Dreyfuss, G. Redial generations of Fasciola hepatica: A review. J. Helminthol. 2009, 83, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Hodgkinson, J.; Cwiklinski, K.; Beesley, N.J.; Paterson, S.; Williams, D.J.L. Identification of putative markers of triclabendazole resistance by a genome-wide analysis of genetically recombinant Fasciola hepatica. Parasitology 2013, 140, 1523–1533. [Google Scholar] [CrossRef] [PubMed]
- Alkhalfioui, F.; Renard, M.; Vensel, W.H.; Wong, J.; Tanaka, C.K.; Hurkman, W.J.; Buchanan, B.B.; Montrichard, F. Thioredoxin-linked proteins are reduced during germination of Medicago truncatula seeds. Plant Physiol. 2007, 144, 1559–1579. [Google Scholar] [CrossRef] [PubMed]
- Montrichard, F.; Alkhalfioui, F.; Yano, H.; Vensel, W.H.; Hurkman, W.J.; Buchanan, B.B. Thioredoxin targets in plants: The first 30 years. J. Proteom. 2009, 72, 452–474. [Google Scholar] [CrossRef] [PubMed]
- Dorey, A.; Cwiklinski, K.; Rooney, J.; De Marco Verissimo, C.; López Corrales, J.; Jewhurst, H.; Fazekas, B.; Calvani, N.E.D.; Hamon, S.; Gaughan, S.; et al. Autonomous non antioxidant roles for Fasciola hepatica secreted Thioredoxin-1 and Peroxiredoxin-1. Front. Cell. Infect. Microbiol. 2021, 11, 667272. [Google Scholar] [CrossRef] [PubMed]
- Agatsuma, T.; Arakawa, Y.; Iwagami, M.; Honzako, Y.; Cahyaningsih, U.; Kang, S.-Y.; Hong, S.-J. Molecular evidence of natural hybridization between Fasciola hepatica and F. gigantica. Parasitol. Int. 2000, 49, 231–238. [Google Scholar] [CrossRef]
- Calvani, N.E.D.; Šlapeta, J. Fasciola species introgression: Just a fluke or something more? Trends Parasitol. 2021, 37, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Omar, M.A.; Elmajdoub, L.O.; Ali, A.O.; Ibrahim, D.A.; Sorour, S.S.; Al-Wabel, M.A.; Ahmed, A.I.; Suresh, M.; Metwally, A.M. Genetic characterization and phylogenetic analysis of Fasciola species based on ITS2 gene sequence, with first molecular evidence of intermediate Fasciola from water buffaloes in Aswan, Egypt. Ann. Parasitol. 2021, 67, 55–65. [Google Scholar]
- Haridwal, S.; Malatji, M.P.; Mukaratirwa, S. Morphological and molecular characterization of Fasciola hepatica and Fasciola gigantica phenotypes from co-endemic localities in Mpumalanga and KwaZulu-Natal provinces of South Africa. Food Waterborne Parasitol. 2021, 22, e00114. [Google Scholar] [CrossRef]
- Evack, J.G.; Schmidt, R.S.; Boltryk, S.D.; Voss, T.S.; Batil, A.A.; Ngandolo, B.N.; Greter, H.; Utzinger, J.; Zinsstag, J.; Balmer, O. Molecular confirmation of a Fasciola gigantica × Fasciola hepatica hybrid in a Chadian bovine. J. Parasitol. 2020, 106, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Mucheka, V.T.; Lamb, J.M.; Pfukenyi, D.M.; Mukaratirwa, S. DNA sequence analyses reveal co-occurrence of novel haplotypes of Fasciola gigantica with F. hepatica in South Africa and Zimbabwe. Vet. Parasitol. 2015, 214, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Cwiklinski, K.; O’Neill, S.M.; Donnelly, S.; Dalton, J.P. A prospective view of animal and human Fasciolosis. Parasite Immunol. 2016, 38, 558–568. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cwiklinski, K.; Dalton, J.P. Exploiting Comparative Omics to Understand the Pathogenic and Virulence-Associated Protease: Anti-Protease Relationships in the Zoonotic Parasites Fasciola hepatica and Fasciola gigantica. Genes 2022, 13, 1854. https://doi.org/10.3390/genes13101854
Cwiklinski K, Dalton JP. Exploiting Comparative Omics to Understand the Pathogenic and Virulence-Associated Protease: Anti-Protease Relationships in the Zoonotic Parasites Fasciola hepatica and Fasciola gigantica. Genes. 2022; 13(10):1854. https://doi.org/10.3390/genes13101854
Chicago/Turabian StyleCwiklinski, Krystyna, and John Pius Dalton. 2022. "Exploiting Comparative Omics to Understand the Pathogenic and Virulence-Associated Protease: Anti-Protease Relationships in the Zoonotic Parasites Fasciola hepatica and Fasciola gigantica" Genes 13, no. 10: 1854. https://doi.org/10.3390/genes13101854
APA StyleCwiklinski, K., & Dalton, J. P. (2022). Exploiting Comparative Omics to Understand the Pathogenic and Virulence-Associated Protease: Anti-Protease Relationships in the Zoonotic Parasites Fasciola hepatica and Fasciola gigantica. Genes, 13(10), 1854. https://doi.org/10.3390/genes13101854