

Molecular Dissection of Phagocytosis by Proteomic Analysis in Entamoeba histolytica


Abstract
1. Introduction
2. Two General Methods for Phagosome Isolation from E. histolytica Trophozoites
3. Phagosomal Proteins Detected in All Proteomic Studies Represent a Core Set of Constitutive Proteins Necessary for Phagosome Biogenesis
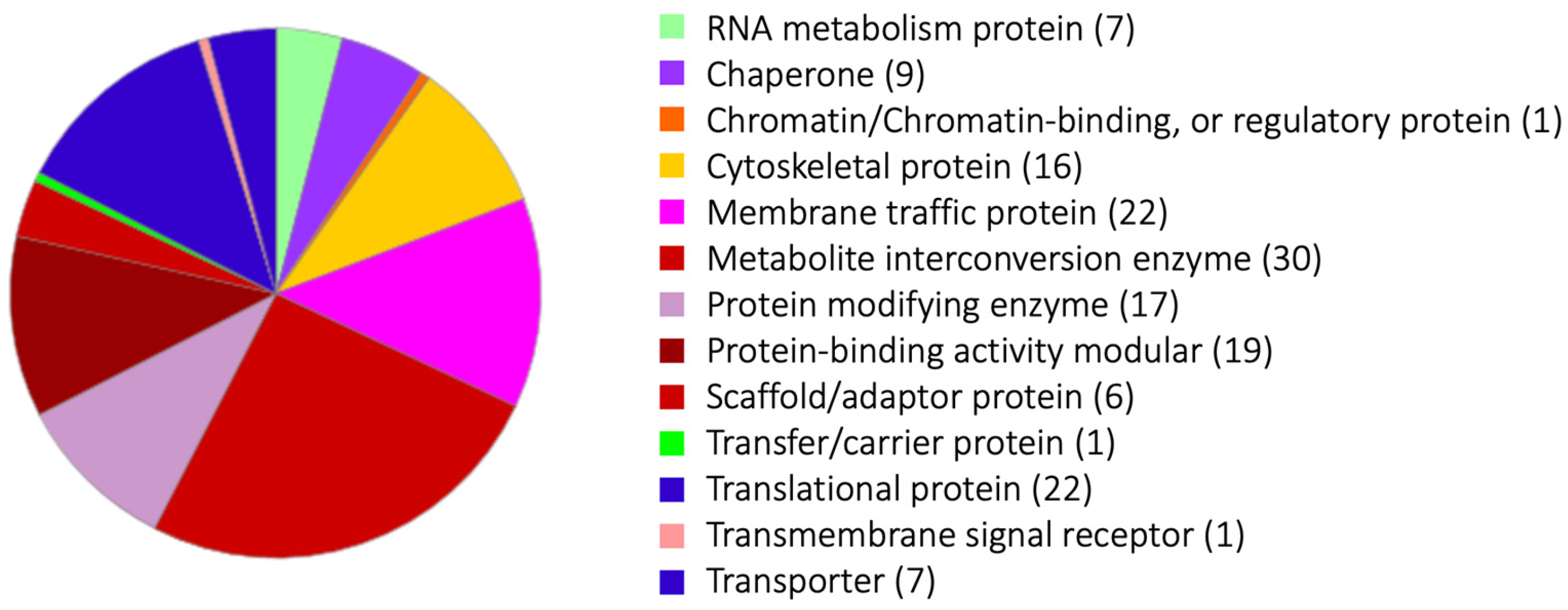
4. Phagosome Proteins Detected Exclusively by the Magnetic Bead Isolation Method, Combined with Bead Serum Coating and Chemical Cross-Linking, Represent Those Involved in Serum-Dependent Phagocytosis or Transiently Associated with Phagosomes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Okada, M.; Nozaki, T. New insights into molecular mechanisms of phagocytosis in Entamoeba histolytica by proteomic analysis. Arch. Med. Res. 2006, 37, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Okada, M.; Huston, C.D.; Oue, M.; Mann, B.J.; Petri, W.A.; Kita, K.; Nozaki, T. Kinetics and strain variation of phagosome proteins of Entamoeba histolytica by proteomic analysis. Mol. Biochem. Parasitol. 2006, 145, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Marion, S.; Laurent, C.; Guillén, N. Signalization and cytoskeleton activity through myosin IB during the early steps of phagocytosis in Entamoeba histolytica: A proteomic approach. Cell. Microbiol. 2005, 7, 1504–1518. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Nakada-Tsukui, K.; Nozaki, T. Two isotypes of phosphatidylinositol 3-phosphate-binding sorting nexins play distinct roles in trogocytosis in Entamoeba histolytica. Cell Microbiol. 2020, 22, e13144. [Google Scholar] [CrossRef]
- Nakada-tsukui, K.; Watanabe, N.; Shibata, K.; Wahyuni, R.; Miyamoto, E.; Nozaki, T. Proteomic analysis of Atg8-dependent recruitment of phagosomal proteins in the enteric protozoan parasite Entamoeba histolytica. Front. Cell Infect. Microbiol. 2022, 12, 1–14. [Google Scholar] [CrossRef]
- Furukawa, A.; Nakada-Tsukui, K.; Nozaki, T. Cysteine protease-binding protein family 6 mediates the trafficking of amylases to phagosomes in the enteric protozoan entamoeba histolytica. Infect. Immun. 2013, 81, 1820–1829. [Google Scholar] [CrossRef]
- Okada, M.; Huston, C.D.; Mann, B.J.; Petri, W.A.; Kita, K.; Nozaki, T. Proteomic analysis of phagocytosis in the enteric protozoan parasite Entamoeba histolytica. Eukaryot. Cell 2005, 4, 827–831. [Google Scholar] [CrossRef]
- Nakada-Tsukui, K.; Saito-Nakano, Y.; Ali, V.; Nozaki, T. A Retromerlike Complex Is a Novel Rab7 Effector That Is Involved in the Transport of the Virulence Factor Cysteine Protease in the Enteric Protozoan Parasite Entamoeba histolytica Kumiko. Mol. Biol. Cell 2005, 16, 5294–5303. [Google Scholar] [CrossRef]
- Saito-Nakano, Y.; Wahyuni, R.; Nakada-Tsukui, K.; Tomii, K.; Nozaki, T. Rab7D small GTPase is involved in phago-, trogocytosis and cytoskeletal reorganization in the enteric protozoan Entamoeba histolytica. Cell Microbiol. 2021, 23, e13267. [Google Scholar] [CrossRef]
- Mitra, B.N.; Saito-Nakano, Y.; Nakada-Tsukui, K.; Sato, D.; Nozaki, T. Rab11B small GTPase regulates secretion of cysteine proteases in the enteric protozoan parasite Entamoeba histolytica. Cell Microbiol. 2007, 9, 2112–2125. [Google Scholar] [CrossRef]
- Verma, K.; Saito-Nakano, Y.; Nozaki, T.; Datta, S. Insights into endosomal maturation of human holo-transferrin in the enteric parasite Entamoeba histolytica: Essential roles of Rab7A and Rab5 in biogenesis of giant early endocytic vacuoles. Cell Microbiol. 2015, 17, 1779–1796. [Google Scholar] [CrossRef]
- Saito-nNkano, Y.; Mitra, B.N.; Nakada-tsukui, K.; Sato, D.; Nozaki, T. Two Rab7 isotypes, Eh Rab7A and Eh Rab7B, play distinct roles in biogenesis of lysosomes and phagosomes in the enteric protozoan parasite Entamoeba histolytica. Cell Microbiol. 2007, 9, 1796–1808. [Google Scholar] [CrossRef]
- Saito-Nakano, Y.; Yasuda, T.; Nakada-Tsukui, K.; Leippe, M.; Nozaki, T. Rab5-associated vacuoles play a unique role in phagocytosis of the enteric protozoan parasite Entamoeba histolytica. J. Biol. Chem. 2004, 279, 49497–49507. [Google Scholar] [CrossRef]
- Herman, E.; Siegesmund, M.A.; Bottery, M.J.; Van Aerle, R.; Shather, M.M.; Caler, E.; Dacks, J.B.; Van Der Giezen, M. Membrane Trafficking Modulation during Entamoeba Encystation. Sci. Rep. 2017, 7, 12854. [Google Scholar] [CrossRef]
- Nakada-Tsukui, K.; Saito-Nakano, Y.; Husain, A.; Nozaki, T. Conservation and function of Rab small GTPases in Entamoeba: Annotation of E. invadens Rab and its use for the understanding of Entamoeba biology. Exp. Parasitol. 2010, 126, 337–347. [Google Scholar] [CrossRef]
- Hanadate, Y.; Saito-Nakano, Y.; Nakada-Tsukui, K.; Nozaki, T. Endoplasmic reticulum-resident Rab8A GTPase is involved in phagocytosis in the protozoan parasite Entamoeba histolytica. Cell Microbiol. 2016, 18, 1358–1373. [Google Scholar] [CrossRef]
- Hanadate, Y.; Saito-Nakano, Y.; Nakada-Tsukui, K.; Nozaki, T. Identification and characterization of the entamoeba histolytica rab8a binding protein: A Cdc50 homolog. Int. J. Mol. Sci. 2018, 19, 3831. [Google Scholar] [CrossRef]
- Bansal, D.; Ave, P.; Kerneis, S.; Frileux, P.; Boché, O.; Baglin, A.C.; Dubost, G.; Leguern, A.S.; Prevost, M.C.; Bracha, R.; et al. An ex-vivo human intestinal model to study Entamoeba histolytica pathogenesis. PLoS Negl. Trop. Dis. 2009, 3, 551. [Google Scholar] [CrossRef]
- Que, X.; Reed, S.L. Cysteine proteinases and the pathogenesis of amebiasis. Clin. Microbiol. Rev. 2000, 13, 196–206. [Google Scholar] [CrossRef]
- Temesvari, L.A.; Harris, E.N.; Stanley, S.L.; Cardelli, J.A. Early and late endosomal compartments of Entamoeba histolytica are enriched in cysteine proteases, acid phosphatase and several Ras-related Rab GTPases. Mol. Biochem. Parasitol. 1999, 103, 225–241. [Google Scholar] [CrossRef][Green Version]
- Freitas, M.A.R.; Fernandes, H.C.; Calixto, V.C.; Martins, A.S.; Silva, E.F.; Pesquero, J.L.; Gomes, M.A. Entamoeba histolytica: Cysteine proteinase activity and virulence. Focus on cysteine proteinase 5 expression levels. Exp. Parasitol. 2009, 122, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Siqueira-Neto, J.L.; Debnath, A.; McCall, L.I.; Bernatchez, J.A.; Ndao, M.; Reed, S.L.; Rosenthal, P.J. Cysteine proteases in protozoan parasites. PLoS Negl. Trop. Dis. 2018, 12, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Matthiesen, J.; Bär, A.K.; Bartels, A.K.; Marien, D.; Ofori, S.; Biller, L.; Tannich, E.; Lotter, H.; Bruchhaus, I. Overexpression of specific cysteine peptidases confers pathogenicity to a nonpathogenic Entamoeba histolytica clone. MBio 2013, 4, 00072-13. [Google Scholar] [CrossRef] [PubMed]
- Mitra, B.N.; Yasuda, T.; Kobayashi, S.; Saito-Nakano, Y.; Nozaki, T. Differences in morphology of phagosomes and kinetics of acidification and degradation in phagosomes between the pathogenic Entamoeba histolytica and the non-pathogenic Entamoeba dispar. Cell Motil. Cytoskelet. 2005, 62, 84–99. [Google Scholar] [CrossRef]
- Mitra, B.N.; Kobayashi, S.; Saito-Nakano, Y.; Nozaki, T. Entamoeba histolytica: Differences in phagosome acidification and degradation between attenuated and virulent strains. Exp. Parasitol. 2006, 114, 57–61. [Google Scholar] [CrossRef]
- Ragland, S.A.; Criss, A.K. From bacterial killing to immune modulation: Recent insights into the functions of lysozyme. PLoS Pathog. 2017, 13, 1–22. [Google Scholar] [CrossRef]
- Fleming, D.; Chahin, L.; Rumbaugh, K. Glycoside hydrolases degrade polymicrobial bacterial biofilms in wounds. Antimicrob. Agents Chemother. 2017, 61, 1–9. [Google Scholar] [CrossRef]
- Koo, I.C.; Ohol, Y.M.; Wu, P.; Morisaki, J.H.; Cox, J.S.; Brown, E.J. Role for lysosomal enzyme β-hexosaminidase in the control of mycobacteria infection. Proc. Natl. Acad. Sci. USA 2008, 105, 710–715. [Google Scholar] [CrossRef]
- Marumo, K.; Nakada-Tsukui, K.; Tomii, K.; Nozaki, T. Ligand heterogeneity of the cysteine protease binding protein family in the parasitic protist Entamoeba histolytica. Int. J. Parasitol. 2014, 44, 625–635. [Google Scholar] [CrossRef]
- Furukawa, A.; Nakada-Tsukui, K.; Nozaki, T. Novel transmembrane receptor involved in phagosome transport of lysozymes and β-hexosaminidase in the enteric protozoan Entamoeba histolytica. PLoS Pathog. 2012, 8, e1002539. [Google Scholar] [CrossRef]
- Isnard, A.; Shio, M.T.; Olivier, M. Impact of Leishmania metalloprotease GP63 on macrophage signaling. Front. Cell Infect. Microbiol. 2012, 2, 72. [Google Scholar] [CrossRef]
- da Silva Lira Filho, A.; Fajardo, E.F.; Chang, K.P.; Clément, P.; Olivier, M. Leishmania Exosomes/Extracellular Vesicles Containing GP63 Are Essential for Enhance Cutaneous Leishmaniasis Development Upon Co-Inoculation of Leishmania amazonensis and Its Exosomes. Front. Cell Infect. Microbiol. 2022, 11, 1–14. [Google Scholar] [CrossRef]
- Castro Neto, A.L.; Brito, A.N.A.L.M.; Rezende, A.M.; Magalhães, F.B.; De Melo Neto, O.P. In silico characterization of multiple genes encoding the GP63 virulence protein from Leishmania braziliensis: Identification of sources of variation and putative roles in immune evasion. BMC Genomics 2019, 20, 118. [Google Scholar] [CrossRef]
- Lieke, T.; Nylén, S.; Eidsmo, L.; McMaster, W.R.; Mohammadi, A.M.; Khamesipour, A.; Berg, L.; Akuffo, H. Leishmania surface protein gp63 binds directly to human natural killer cells and inhibits proliferation. Clin. Exp. Immunol. 2008, 153, 221–230. [Google Scholar] [CrossRef]
- Hassani, K.; Shio, M.T.; Martel, C.; Faubert, D.; Olivier, M. Absence of metalloprotease GP63 alters the protein content of leishmania exosomes. PLoS One 2014, 9, 95007. [Google Scholar] [CrossRef]
- Sharma, M.; Morgado, P.; Zhang, H.; Ehrenkaufer, G.; Manna, D.; Singh, U. Characterization of Extracellular Vesicles from Entamoeba histolytica Identifies Roles in Intracellular Communication That Regulates Parasite Growth and Development. Infect. Immun. 2020, 88, e00349-20. [Google Scholar] [CrossRef]
- Saharan, O.; Mehendale, N.; Kamat, S.S. Phagocytosis: A (Sphingo) Lipid Story. Curr. Res. Chem. Biol. 2022, 2, 100030. [Google Scholar] [CrossRef]
- Williams, E.T.; Chen, X.; Moore, D.J. VPS35, the retromer complex and Parkinson’s disease. J. Parkinsons. Dis. 2017, 7, 219–233. [Google Scholar] [CrossRef]
- Welter, B.H.; Temesvari, L.A. A unique Rab GTPase, EhRabA, of Entamoeba histolytica, localizes to the leading edge of motile cells. Mol. Biochem. Parasitol. 2004, 135, 185–195. [Google Scholar] [CrossRef]
- Welter, B.H.; Temesvari, L.A. Overexpression of a mutant form of EhRabA, a unique rab GTPase of entamoeba histolytica, alters endoplasmic reticulum morphology and localization of the Gal/GalNAc adherence lectin. Eukaryot. Cell 2009, 8, 1014–1026. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.C.; Griffiths, G.; Wessling-Resnick, M.; Fransen, J.A.M.; Bennett, H.; Jones, A.T. A role for the small GTPase Rab21 in the early endocytic pathway. J. Cell Sci. 2004, 117, 6297–6311. [Google Scholar] [CrossRef] [PubMed]
- Levin-Konigsberg, R.; Grinstein, S. Phagosome-endoplasmic reticulum contacts: Kissing and not running. Traffic 2020, 21, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Jo, E.K.; Yuk, J.M.; Shin, D.M.; Sasakawa, C. Roles of autophagy in elimination of intracellular bacterial pathogens. Front. Immunol. 2013, 4, 1–9. [Google Scholar] [CrossRef]
- Zindrou, S.; Orozco, E.; Linder, E.; Téllez, A.; Björkman, A. Specific detection of Entamoeba histolytica DNA by hemolysin gene targeted PCR. Acta Trop. 2001, 78, 117–125. [Google Scholar] [CrossRef]
- Rath, P.P.; Gourinath, S. The actin cytoskeleton orchestra in Entamoeba histolytica. Proteins Struct. Funct. Bioinforma. 2020, 88, 1361–1375. [Google Scholar] [CrossRef]
- Bharadwaj, R.; Sharma, S.; Arya, R.; Bhattacharya, S.; Bhattacharya, A. EhRho1 regulates phagocytosis by modulating actin dynamics through EhFormin1 and EhProfilin1 in Entamoeba histolytica. Cell Microbiol. 2018, 20, e12851. [Google Scholar] [CrossRef]
- Dos Remedios, C.G.; Chhabra, D.; Kekic, M.; Dedova, I.V.; Tsubakihara, M.; Berry, D.A.; Nosworthy, N.J. Actin binding proteins: Regulation of cytoskeletal microfilaments. Physiol. Rev. 2003, 83, 433–473. [Google Scholar] [CrossRef]


{kind=link}
| Number | Journal | Year | Method | Detected Proteins | Reference |
|---|---|---|---|---|---|
| 1 | Mol. Biochem. Parasitol. | 2006 | Density-gradient | 155 | [2] |
| 2 | Arch. Med. Res. | 2006 | Density-gradient | 151 | [1] |
| 3 | Infect. Immun. | 2013 | Density-gradient | 375 | [6] |
| 4 | Cell. Microbiol. | 2019 | Magnetic beads (serum coated; cross linking) | 884 | [4] |
| 5 | Front. Cell. Infect. Microbiol. | 2022 | Magnetic beads (serum coated; cross-linking) | 738 | [5] |
| AmoebaDB Annotation | EHI Number | Common Name | Phagosome Isolation Method | Proteome Number | General Function |
|---|---|---|---|---|---|
| Rab family GTPase | EHI_108610 | Rab1A | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Membrane traffic |
| Small GTPase Rab7A | EHI_192810 | Rab7A | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Early phagosome maturation |
| EhRab7B protein | EHI_081330 | Rab7B | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Membrane traffic |
| EhRab7D protein | EHI_082070 | Rab7D | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Phagosome maturation |
| Small GTPase Rab11B | EHI_107250 | Rab11B | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Phagosome maturation (in Eh)/CP secretion (in Eh)/endosome recycling |
| Rab family GTPase | EHI_161030 | Rab11C | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Endosome recycling |
| Rab family GTPase | EHI_153690 | RabC1 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Membrane traffic |
| Rab family GTPase | EHI_143650 | RabC3 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Membrane traffic |
| Rho family GTPase | EHI_070730 | Rho1 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Cell motility |
| Rho family GTPase | EHI_129750 | Rho7 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Cell motility |
| Ras family GTPase | EHI_058090 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Signal transduction | |
| Cell surface protease gp63, putative | EHI_200230 | Gp63 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme |
| Lysozyme, putative | EHI_199110 | Lysozyme I | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme |
| Lysozyme, putative | EHI_096570 | Lysozyme II | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme |
| Cysteine proteinase 1 | EHI_074180 | CP-A1 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme |
| Histolysain | EHI_033710 | CP-A2 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme |
| Cysteine proteinase, putative | EHI_050570 | CP-A4 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme |
| Cysteine proteinase | EHI_168240 | CP-A5 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme |
| Dipeptidyl-peptidase | EHI_136440 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme | |
| α-amylase family protein | EHI_023360 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme | |
| β-hexosaminidase | EHI_007330 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme | |
| Serine carboxypeptidase (S28) family protein | EHI_054530 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme | |
| Rab family GTPase | EHI_168600 | RabA | Magnetic beads | 4,5 | Membrane traffic |
| Rab family GTPase | EHI_079890 | RabC7 | Magnetic beads | 4,5 | Membrane traffic |
| Rab family GTPase | EHI_122730 | RabC5 | Magnetic beads | 4,5 | Membrane traffic |
| Rab family GTPase | EHI_053420 | RabI2 | Magnetic beads | 4,5 | Membrane traffic |
| Rab family GTPase | EHI_024680 | RabK1 | Magnetic beads | 4,5 | Membrane traffic |
| Rab family GTPase | EHI_117890 | RabP2 | Magnetic beads | 4,5 | Membrane traffic |
| Ras-related protein | EHI_129330 | Rab21 | Magnetic beads | 4,5 | Membrane traffic |
| Rho family GTPase | EHI_190440 | Rho10 | Magnetic beads | 4,5 | Cell motility |
| Rho family GTPase | EHI_135450 | Rho13 | Magnetic beads | 4,5 | Cell motility |
| Ras family GTPase | EHI_198330 | Not assigned | Magnetic beads | 4,5 | Signal transduction |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watanabe, N.; Nakada-Tsukui, K.; Nozaki, T. Molecular Dissection of Phagocytosis by Proteomic Analysis in Entamoeba histolytica. Genes 2023, 14, 379. https://doi.org/10.3390/genes14020379
Watanabe N, Nakada-Tsukui K, Nozaki T. Molecular Dissection of Phagocytosis by Proteomic Analysis in Entamoeba histolytica. Genes. 2023; 14(2):379. https://doi.org/10.3390/genes14020379
Chicago/Turabian StyleWatanabe, Natsuki, Kumiko Nakada-Tsukui, and Tomoyoshi Nozaki. 2023. "Molecular Dissection of Phagocytosis by Proteomic Analysis in Entamoeba histolytica" Genes 14, no. 2: 379. https://doi.org/10.3390/genes14020379
APA StyleWatanabe, N., Nakada-Tsukui, K., & Nozaki, T. (2023). Molecular Dissection of Phagocytosis by Proteomic Analysis in Entamoeba histolytica. Genes, 14(2), 379. https://doi.org/10.3390/genes14020379