
Uniparental Lineages from the Oldest Indigenous Population of Ecuador: The Tsachilas

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection and DNA Extraction
2.2. mtDNA and Y Chromosome Genotyping
2.3. Data Analysis
3. Results
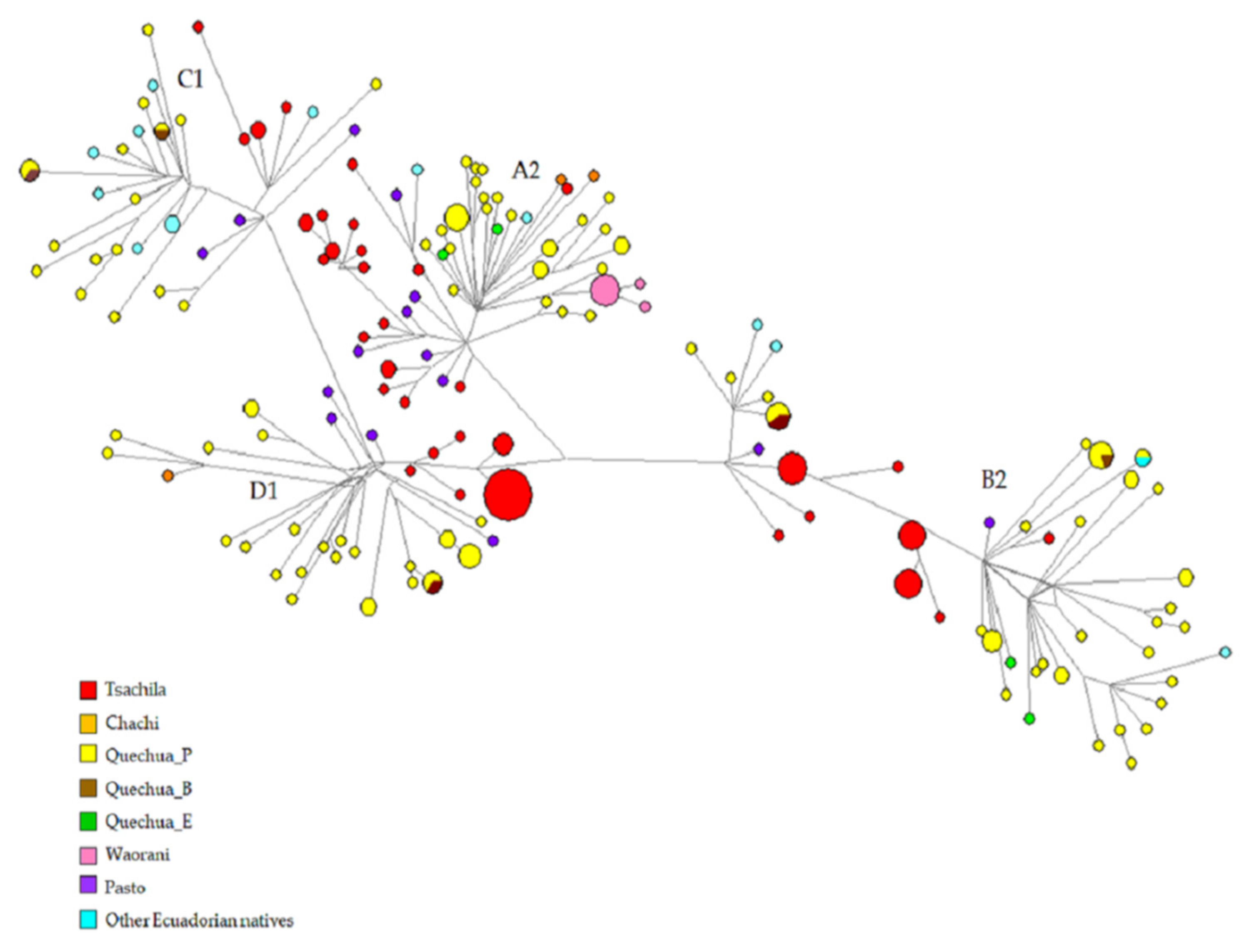
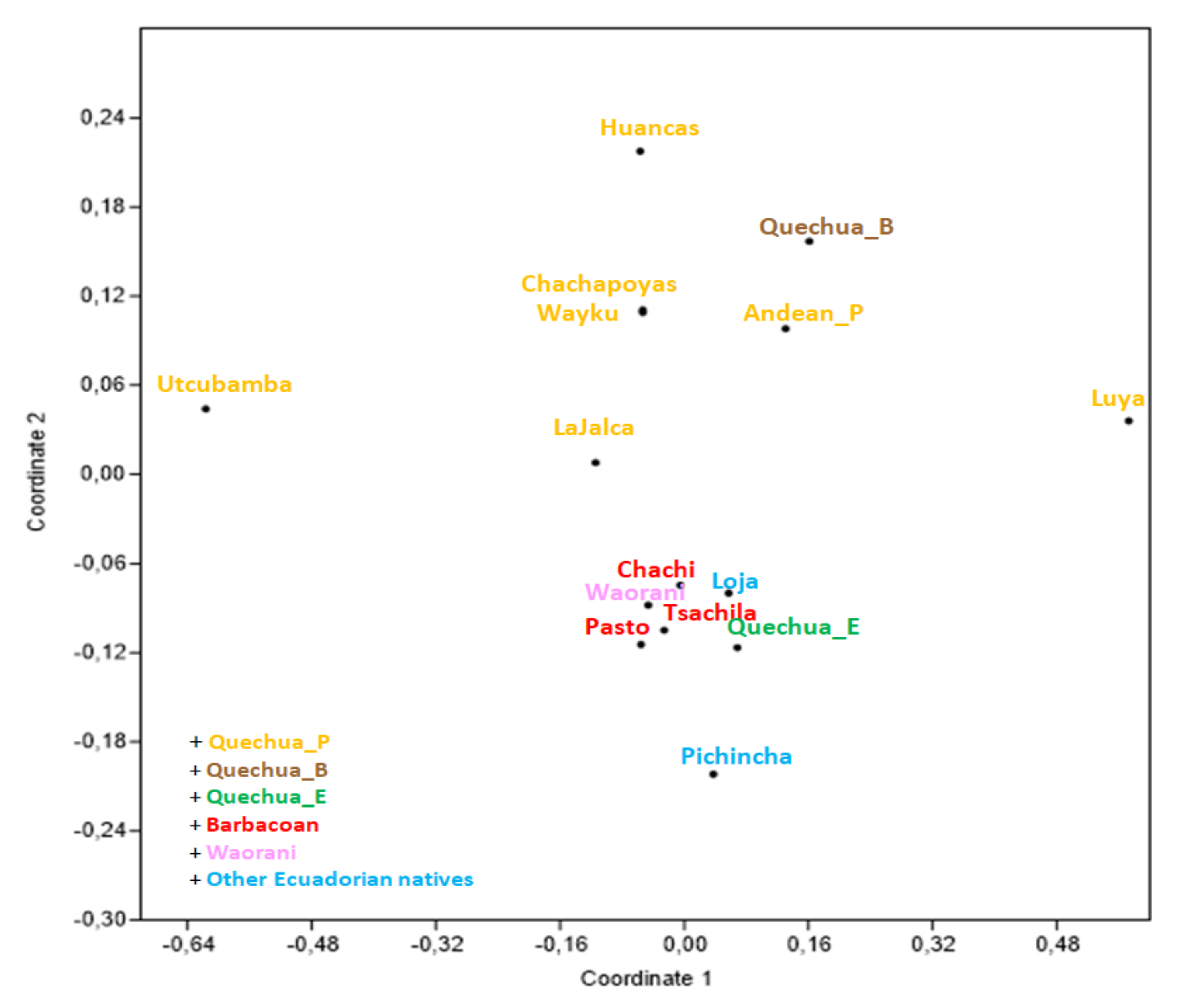
3.1. mtDNA
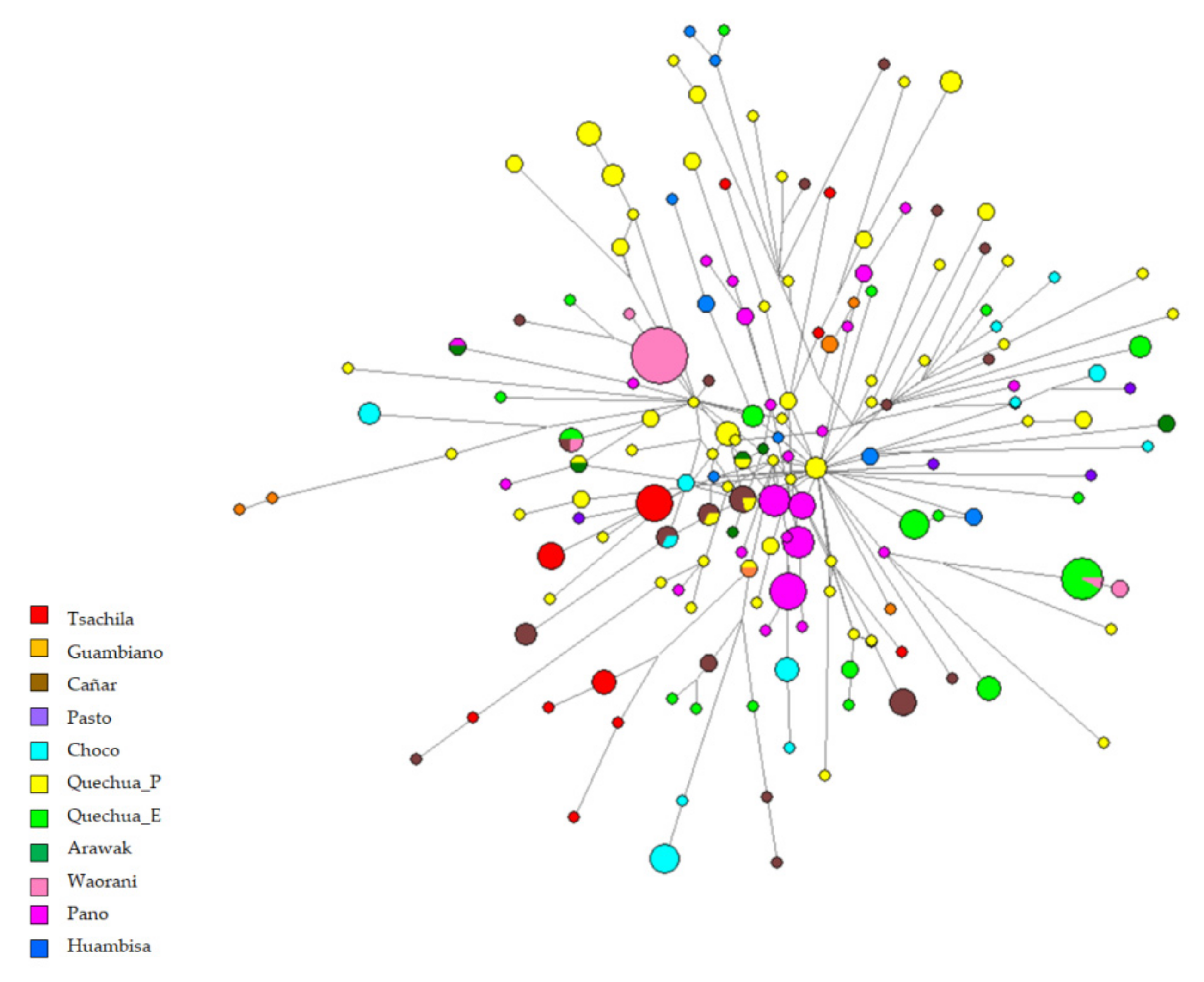
3.2. Y Chromosome
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Llamas, B.; Fehren-Schmitz, L.; Valverde, G.; Soubrier, J.; Mallick, S.; Rohland, N.; Nordenfelt, S.; Valdiosera, C.; Richards, S.M.; Rohrlach, A.; et al. Ancient mitochondrial DNA provides high-resolution time scale of the peopling of the Americas. Sci. Adv. 2016, 2, e1501385. [Google Scholar] [CrossRef] [PubMed]
- Posth, C.; Nakatsuka, N.; Lazaridis, I.; Skoglund, P.; Mallick, S.; Lamnidis, T.C.; Rohland, N.; Nägele, K.; Adamski, N.; Bertolini, E. Reconstructing the deep population history of Central and South America. Cell 2018, 175, 1185–1197.e22. [Google Scholar] [CrossRef]
- Gnecchi-Ruscone, G.A.; Sarno, S.; De Fanti, S.; Gianvincenzo, L.; Giuliani, C.; Boattini, A.; Bortolini, E.; Di Corcia, T.; Sanchez Mellado, C.; Dàvila Francia, T.J.; et al. Dissecting the Pre-Columbian Genomic Ancestry of Native Americans along the Andes-Amazonia divide. Mol. Biol. Evol. 2019, 36, 1254–1269. [Google Scholar] [CrossRef]
- Nakatsuka, N.; Lazaridis, I.; Barbieri, C.; Skoglund, P.; Rohland, N.; Mallick, S.; Posth, C.; Harkins-Kinkaid, K.; Ferry, M.; Harney, É.; et al. A paleogenomic reconstruction of the deep population history of the Andes. Cell 2020, 181, 1131–1145.e21. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, J.R.; Salazar-Granara, A.; Acosta, O.; Castillo-Herrera, W.; Fujita, R.; Pena, S.D.J.; Santos, F.R. Tracing the genomic ancestry of Peruvians reveals a major legacy of pre-Columbian ancestors. J. Hum. Genet. 2013, 58, 627–634. [Google Scholar] [CrossRef]
- Barbieri, C.; Heggarty, P.; Yang Yao, D.; Ferri, G.; De Fanti, S.; Sarno, S.; Ciani, G.; Boattini, A.; Luiselli, D.; Pettener, D. Between Andes and Amazon: The genetic profile of the Arawak-speaking Yanesha. Am. J. Phys. Anthropol. 2014, 155, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, J.R.; Lacerda, D.R.; Jota, M.M.; Robles-Ruiz, P.; Danos, P.; Paz-y-Miño, C.; Wells, S.; Santos, F.R.; Fujita, R. Tracing the genetic history of the ‘Cañaris’ from Ecuador and Peru using uniparental DNA markers. BMC Genom. 2020, 21, 1–10. [Google Scholar] [CrossRef]
- Scliar, M.O.; Gouveia, M.H.; Benazzo, A.; Ghirotto, S.; Fagundes, N.J.; Leal, T.P.; Magalhães, W.C.; Pereira, L.; Rodrigues, M.R.; Soares-Souza, G.B.; et al. Bayesian inferences suggest that Amazon Yunga Natives diverged from Andeans less than 5000 ybp: Implications for South American prehistory. BMC Evol. Biol. 2014, 14, 174. [Google Scholar] [CrossRef]
- Di Corcia, T.; Sanchez Mellado, C.; Davila Francia, T.J.; Ferri, G.; Sarno, S.; Luiselli, D.; Rickards, O. East of the Andes: The genetic profile of the Peruvian Amazon populations. Am. J. Phys. Anthropol. 2017, 163, 328–338. [Google Scholar] [CrossRef]
- Costales Samaniego, A.; CostalesPeñaherrera, D. Etnografía, Lingüística e Historia Antigua de los Caras o YumbosColorados, 1534–1978; Abya-Yala, E., Ed.; University of Michigan: Ann Arbor, MI, USA, 2002; ISBN 978-9-9782-2242-3. [Google Scholar]
- Lippi, R.D. Tropical Forest Archaeology in Western Pichincha, Ecuador. In Case Studies in Archaeology; Quilter, J., Ed.; Wadsworth Publishing: Belmont, CA, USA, 2004; p. 168. ISBN 978-0-5346-1294-8. [Google Scholar]
- Barriga Lopez, F. EtnologiaEcuadoriana; Instituto Ecuatoriano de CreditoEducativo y Becas: Quito, Ecuador, 1987; ISBN 978-9-9786-2888-1. [Google Scholar]
- Greenberg, J.H. Language in the Americas; Stanford University Press: Stanford, CA, USA, 1987; ISBN 978-0-8047-1315-3. [Google Scholar]
- Ruhlen, M. A Guide to the World’s Languages; Stanford University Press: Stanford, CA, USA, 1998; ISBN 978-0-8047-1250-7. [Google Scholar]
- Kaufman, T. Language History in South America: What We Know and How to Know More. In Amazonian Linguistics: Studies in Lowland South American Languages; Payne, D.L., Ed.; University of Texas Press: Austin, TX, USA, 1990; pp. 13–73. ISBN 978-0-2927-2364-1. [Google Scholar]
- Constenla-Umaña, A. Las Lenguas del Area Intermedia: Introduccion a Suestudio Areal; Editorial de la Universidad de Costa Rica: San José, Costa Rica, 1991; ISBN 978-9-9776-7158-1. [Google Scholar]
- Stark, L. Indigenous Languages of Lowland Ecuador: History and Current Status. In South American Indian Languages: Retrospect and Prospect; Manelis Klein, H.E., Stark, L.R., Eds.; University of Texas Press: Austin, TX, USA, 1985; pp. 157–193. ISBN 978-0-2927-3732-7. [Google Scholar]
- Curnow, T.; Liddicoat, A. The Barbacoan Languages of Colombia and Ecuador. Anthropol. Linguist. 1998, 40, 384–408. [Google Scholar]
- Curnow, T. Why Paez is not a Barbacoan Language: The Nonexistence of “Moguex” and the Use of Early Sources. Int. J. Am. Linguist. 1998, 64, 338–351. [Google Scholar] [CrossRef]
- Lippi, R.D.; Gudiño, A.M. Inkas and Yumbos at Palmitopamba in Northwestern Ecuador. In Distant Provinces in the Inka Empire: Toward a Deeper Understanding of Inka Imperialism; Malpass, M., Alconini, S., Eds.; University of Iowa Press: Iowa City, IA, USA, 2010; pp. 260–278. ISBN 978-1-5872-9869-1. [Google Scholar]
- Lippi, R.D. La Expansión de Las Poblaciones Barbacoas en el Noroeste de Ecuador. In Revista de Arqueología del Área Intermedia; Instituto Colombiano de Antropología e Historia y Sociedad Colombiana de Arqueología: Bogotá, Colombia, 2004; Volume 6, pp. 249–276. [Google Scholar]
- CONAIE—Confederación de Nacionalidades Indígenas del Ecuador. Available online: http://conaie.org (accessed on 25 January 2021).
- Von Hagen, W. Los Indios Tsachela del Oeste Ecuatoriano. In Los Clasicos De La Etnografıa Sobre Los Colorados; Juncosa, J., Ed.; Ediciones ABYA-YALA: Quito, Ecuador, 1988; ISBN 978-9978-67-325-6. [Google Scholar]
- Salomon, F. Los Yumbos, Niguas y Tsáchila o «Colorados» Durante la Colonia Española: Etnohistoria del Noroccidente de Pichincha; Ediciones ABYA-YALA: Quito, Ecuador, 1997; ISBN 978-2-8218-4431-5. [Google Scholar]
- Lewis, C.M.; Lizarraga, B.; Tito, R.Y.; Lopez, P.W.; Iannacone, G.C.; Medina, A.; Stone, A.C. Mitochondrial DNA and the peopling of South America. Hum. Biol. 2007, 79, 159–178. [Google Scholar] [CrossRef]
- Bisso-Machado, R.; Bortolini, M.C.; Salzano, F.M. Uniparental genetic markers in South Amerindians. Genet. Mol. Biol. 2012, 35, 365–387. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, S.; Alfonso-Sánchez, M.A.; Valverde, L.; Sánchez, D.; Zarrabeitia, M.T.; Odriozola, A.; Martínez-Jarreta, B.; de Pancorbo, M.M. Genetic uniqueness of the Waorani tribe from the Ecuadorian Amazon. Heredity 2012, 108, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Brandini, S.; Bergamaschi, P.; Cerna, M.F.; Gandini, F.; Bastaroli, F.; Bertolini, E.; Cereda, C.; Ferretti, L.; Gómez-Carballa, A.; Battaglia, V.; et al. The Paleo-Indian Entry into South America According to Mitogenomes. Mol. Biol. Evol. 2018, 35, 299–311. [Google Scholar] [CrossRef]
- Zegura, S.L.; Karafet, T.M.; Zhivotovsky, L.A.; Hammer, M.F. High-resolution SNPs and microsatellite haplotypes point to a single recent entry of Native American Y chromosomes into the Americas. Mol. Biol. Evol. 2004, 21, 164–175. [Google Scholar] [CrossRef]
- Roewer, L.; Nothnagel, M.; Gusmão, L.; Gomes, V.; González, M.; Corach, D.; Sala, A.; Alechine, E.; Palha, T.; Santos, N.; et al. Continent-wide decoupling of Y-chromosomal genetic variation from language and geography in native South Americans. PLoS Genet. 2013, 9, e1003460. [Google Scholar] [CrossRef] [PubMed]
- Guevara, E.K.; Palo, J.U.; Guillén, S.; Sajantila, A. MtDNA and Y-chromosomal diversity in the Chachapoya, a population from the northeast Peruvian Andes-Amazon divide. Am. J. Hum. Biol. 2016, 28, 857–867. [Google Scholar] [CrossRef]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting outprocedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef]
- Torroni, A.; Rengo, C.; Guida, V.; Cruciani, F.; Sellitto, D.; Coppa, A.; Calderon, F.L.; Simionati, B.; Valle, G.; Richards, M.; et al. Do the Four Clades of the mtDNA Haplogroup L2 Evolve at Different Rates? Am. J. Hum. Genet. 2001, 69, 1348–1356. [Google Scholar] [CrossRef]
- Andrews, R.M.; Kubacka, I.; Chinnery, P.F.; Lightowlers, R.N.; Turnbull, D.M.; Howell, N. Reanalysis and revision of the Cambridge reference sequence for human mitochondrial DNA. Nat. Genet. 1999, 23, 147. [Google Scholar] [CrossRef] [PubMed]
- Weissensteiner, H.; Pacher, D.; Kloss-Brandstätter, A.; Forer, L.; Specht, G.; Bandelt, H.-J.; Kronenberg, F.; Salas, A.; Schönherr, S. HaploGrep 2: Mitochondrial haplogroup classification in the era of high-throughput sequencing. Nucleic Acids Res. 2016, 44, W58–W63. [Google Scholar] [CrossRef]
- Van Oven, M.; Kayser, M. Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation. Hum. Mutat. 2009, 30, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Whit Athey. Y Haplogroup Prediction from Y-STR Values. Available online: http://www.hprg.com/hapest5 (accessed on 25 April 2020).
- Sevini, F.; Yao, D.Y.; Lomartire, L.; Barbieri, A.; Vianello, D.; Ferri, G.; Franceschi, Z.A. Analysis of population substructure in two sympatric populations of Gran Chaco Argentina. PLoS ONE 2013, 8, e64054. [Google Scholar] [CrossRef] [PubMed]
- Jota, M.S.; Lacerda, D.R.; Sandoval, J.R.; Vieira, P.P.; Ohasi, D.; Santos-Júnior, J.E.; Acosta, O.; Cuellar, C.; Revollo, S.; Paz-Y-Miño, C.; et al. New native South American Y chromosome lineages. J. Hum. Genet. 2016, 61, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, C.; Sandoval, J.R.; Valqui, J.; Shimelman, A.; Ziemendorff, S.; Schroder, R.; Geppert, M.; Roewer, L.; Gray, R.; Stoneking, M.; et al. Enclaves of genetic diversity resisted Inca impacts on population history. Sci. Rep. 2017, 7, 17411. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, J.R.; Lacerda, D.R.; Acosta, O.; Jota, M.S.; Robles-Ruiz, P.; Salazar-Granara, A.; Vieira, P.P.; Paz-Y-Miño, C.; Fujita, R.; Santos, F.R.; et al. The Genetic History of Peruvian Quechua-Lamistas and Chankas: Uniparental DNA Patterns among Autochthonous Amazonian and Andean Populations. Ann. Hum. Genet. 2016, 80, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Patterson, N.; Price, A.L.; Reich, D. Population structure and eigen analysis. PLoS Genet. 2006, 2, e190. [Google Scholar] [CrossRef]
- Mazières, S.; Guitard, E.; Crubézy, E.; Dugoujon, J.M.; Bortolini, M.C.; Bonatto, S.L.; Hutz, M.H.; Bois, E.; Tiouka, F.; Larrouy, G.; et al. Uniparental (mtDNA, Y-chromosome) polymorphisms in French Guiana and two related populations-implications for the region’s colonization. Ann. Hum. Genet. 2008, 72, 145–156. [Google Scholar] [CrossRef]
- Melton, P.E.; Briceno, I.; Gomez, A.; Devor, E.J.; Bernal, J.E.; Crawford, M.H. Biological relationship between Central and South American Chibchan speaking populations: Evidence from mtDNA. Am. J. Phys. Anthropol. 2007, 133, 753–770. [Google Scholar] [CrossRef]
- Barbieri, C.; Heggarty, P.; Castrì, L.; Luiselli, D.; Pettener, D. Mitochondrial DNA variability in the Titicaca basin: Matches and mismatches with linguistics and ethnohistory. Am. J. Hum. Biol. 2011, 23, 89–99. [Google Scholar] [CrossRef]
- Rickards, O.; Martinez-Lambarga, C.; Lum, J.K.; De Stefano, G.F.; Cann, R.L. mtDNA history of the Cayapa Amerinds of Ecuador: Detection of additional founding lineages for Native American populations. Am. J. Hum. Genet. 1999, 65, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Kolman, C.J.; Bermingham, E. Mitochondrial and nuclearDNA diversity in the Choco and Chibcha Amerinds of Panama. Genetics 1997, 147, 1289–1302. [Google Scholar] [CrossRef]
- Ramallo, V.; Bisso-Machado, R.; Bravi, C.; Coble, M.D.; Salzano, F.M.; Hünemeier, T.; Bortolini, M.C. Demographic expansions in South America: Enlightening a complex scenario with genetic and linguistic data. Am. J. Phys. Anthrop. 2013, 150, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.N.; Mazières, S.; Bravi, C.; Ray, N.; Wang, S.; Burley, M.-W.; Ruiz-Linares, A. Contrasting patterns of nuclear and mtDNA diversity in Native American populations. Ann. Hum. Genet. 2010, 74, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.H.; Salzano, F.M.; Bonatto, S.L.; Hutz, M.H.; Coimbra, C.; Santos, R.V. Mitochondrial DNA polymorphism in three Brazilian Indian tribes. Am. J. Hum. Biol. 1996, 8, 317–323. [Google Scholar] [CrossRef]
- Gómez-Carballa, A.; Ignacio-Veiga, A.; Alvarez-Iglesias, V.; Pastoriza-Mourelle, A.; Ruíz, Y.; Pineda, L.; Carracedo, A.; Salas, A. A melting pot of multicontinental mtDNA lineages in admixed Venezuelans. Am. J. Phys. Anthropol. 2012, 147, 78–87. [Google Scholar] [CrossRef]
- Simão, F.; Ferreira, A.P.; de Carvalho, E.F.; Parson, W.; Gusmão, L. Defining mtDNA origins and population stratification in Rio de Janeiro. Forensic Sci. Int. Genet. 2018, 34, 97–104. [Google Scholar] [CrossRef]
- Arias, L.; Barbieri, C.; Barreto, G.; Stoneking, M.; Pakendorf, B. High-resolution mitochondrial DNA analysis sheds light on human diversity, cultural interactions, and population mobility in Northwestern Amazonia. Am. J. Phys. Anthropol. 2018, 165, 238–255. [Google Scholar] [CrossRef]
- Sandoval, J.R.; Lacerda, D.R.; Jota, M.S.; Elward, R.; Acosta, O.; Pinedo, D.; Danos, P.; Cuellar, C.; Revollo, S.; Santos, F.R.; et al. Genetic ancestry of families of putative Inka descent. Mol. Genet. Genom. 2018, 293, 873–881. [Google Scholar] [CrossRef]
- Tamm, E.; Kivisild, T.; Reidla, M.; Metspalu, M.; Smith, D.G.; Mulligan, C.J.; Bravi, C.M.; Rickards, O.; Martinez-Labarga, C.; Khusnutdinova, E.K. Beringian standstill and spread of Native American founders. PLoS ONE 2007, 2, e829. [Google Scholar] [CrossRef]
- Fagundes, N.J.; Kanitz, R.; Eckert, R.; Valls, A.C.; Bogo, M.R.; Salzano, F.M.; Smith, D.G.; Silva, W.A.; Zago, M.A., Jr.; Ribeiro-dos-Santos, A.K.; et al. Mitochondrial population genomics supports a single pre-Clovis origin with a coastal route for the peopling of the Americas. Am. J. Hum. Genet. 2008, 82, 583–592. [Google Scholar] [CrossRef]
- Perego, U.A.; Achilli, A.; Angerhofer, N.; Accetturo, M.; Pala, M.; Olivieri, A.; HooshiarKashani, B.; Ritchie, K.H.; Scozzari, R.; Kong, Q.P.; et al. Distinctive Paleo-Indian migration routes from Beringia marked by two rare mtDNA haplogroups. Curr. Biol. 2009, 19, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø. PAST—Paleontological STatistics Reference Manual Natural History; Natural History Museum University of Oslo: Oslo, Norway, 2011. [Google Scholar]
- Meyer, S.; Weiss, G.; von Haeseler, A. Pattern of nucleotide substitution and rate heterogeneity in the hypervariable regions I and II of human mtDNA. Genetics 1999, 152, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Balaresque, P.; Poulet, N.; Cussat-Blanc, S.; Gerard, P.; Quintana-Murci, L.; Heyer, E.; Jobling, M.A. Y-chromosome descent clusters and male differential reproductive success: Young lineage expansions dominate Asian pastoral nomadic populations. Eur. J. Hum. Genet. 2015, 23, 1413–1422. [Google Scholar] [CrossRef] [PubMed]
- Wilson, I.J.; Weale, M.E.; Balding, D.J. Inferences from DNA data: Population histories evolutionary processes and forensic match probabilities. J. R. Stat. Soc. 2003, 166, 155–188. [Google Scholar] [CrossRef]
- Fenner, J.N. Cross-cultural estimation of the human generation interval for use in genetics-based population divergence studies. Am. J. Phys. Anthropol. 2005, 128, 415–423. [Google Scholar] [CrossRef]
- Kayser, M.; Roewer, L.; Hedman, M.; Henke, L.; Henke, J.; Brauer, S.; Krüger, C.; Krawczak, M.; Nagy, M.; Dobosz, T.; et al. Characteristics and frequency of germline mutations at microsatellite loci from the human Y chromosome, as revealed by direct observation in father/son pairs. Am. J. Hum. Genet. 2000, 66, 1580–1588. [Google Scholar] [CrossRef]
- Babalini, C.; Tarsi, T.; Martínez-Labarga, C.; Scano, G.; Pepe, G.; De Stefano, G.F.; Rickards, O. COL1A2 (type I collagen) polymorphisms in the Colorado Indians of Ecuador. Ann. Hum. Biol. 2005, 32, 666–678. [Google Scholar] [CrossRef]
- Battaglia, V.; Grugni, V.; Perego, U.A.; Angerhofer, N.; Gomez-Palmieri, J.E.; Woodward, S.R.; Achilli, A.; Myres, N.; Torroni, A.; Semino, O. The first peopling of South America: New evidence from Y-chromosome haplogroup Q. PLoS ONE 2013, 8, e71390. [Google Scholar] [CrossRef] [PubMed]
- Toscanini, U.; Gusmão, L.; Berardi, G.; Gomes, V.; Amorim, A.; Salas, A.; Raimondi, E. Male lineages in South American native groups: Evidence of M19 traveling south. Am. J. Phys. Anthropol. 2011, 146, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Bodner, M.; Perego, U.A.; Huber, G.; Fendt, L.; RÖck, A.W.; Zimmermann, B.; Parson, W. Rapid coastal spread of First Americans: Novel insights from South America’s Southern Cone mitochondrial genomes. Genome Res. 2012, 22, 811–820. [Google Scholar] [CrossRef]
- de Saint Pierre, M.; Bravi, C.M.; Motti, J.M.; Fuku, N.; Tanaka, M.; Llop, E.; Bonatto, S.L.; Moraga, M. An alternative model for the early peopling of southern South America revealed by analyses of three mitochondrial DNA haplogroups. PLoS ONE 2012, 7, e43486. [Google Scholar] [CrossRef]
- Loukotka, C. Classification of South American Indian Languages; University of California: Los Angeles, CA, USA, 1968; ISBN 978-3-1102-5803-5. [Google Scholar]
- Santiana, A. The Colorado Indians Tsachila; Imprenta de la Universidad: Quito, Ecuador, 1951. [Google Scholar]
- Adelaar, W.F.H. The Languages of the Andes; Cambridge University Press: Cambridge, UK, 2004; ISBN 978-0-5114-8685-2. [Google Scholar]
- Constenla, A. Comparative Chibchan Phonology. Master’s Thesis, University of Pennsylvania, Philadelphia, PA, USA, 1981. [Google Scholar]
- Guevara, E.K.; Palo, J.U.; Översti, S.; King, J.L.; Seidel, M.; Stoljarova, M.; Wendt, F.R.; Bus, M.M.; Guengerich, A.; Church, W.B.; et al. Genetic assessment reveals no population substructure and divergent regional and sex-specific histories in the Chachapoyas from northeast Peru. PLoS ONE 2020, 15, e0244497. [Google Scholar] [CrossRef]
- Gayà-Vidal, M.; Moral, P.; Saenz-Ruales, N.; Gerbault, P.; Tonasso, L.; Villena, M.; Vasquez, R.; Bravi, C.M.; Dugoujon, J.M. mtDNA and Y-chromosome diversity in Aymaras and Quechuas from Bolivia: Different stories and special genetic traits of the Andean Altiplano populations. Am. J. Phys. Anthropol. 2011, 145, 215–230. [Google Scholar] [CrossRef] [PubMed]
- EstupiñanViteri, T. Tras las Huellas de Rumiñahui; Auspicio del Banco General Rumiñahui: Quito, Ecuador, 2003; ISBN 978-9-9783-0004-6. [Google Scholar]
- Bray, T. Una ExploraciónArqueológica del Pichincha Occidental, Ecuador. Ronald D. Lippi. Museo Jacinto Jijón y Caamaño, Pontificia Universidad Católica del Ecuador, Quito, 1998. Lat. Am. Antiq. 2000, 11, 107–108. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | 18 |
| Countries | Ecuador, Peru |
| Populations | Tsachila, Charchi, Huambisa, Chachapoya, InkaD |
| Max frequency | 83% (Ecuador) |
| Max variance | 0.095 (Peru) |
| TMRCA (95% CI) | 571 (90–3100) |
| Alpha | 0.0217 |
| Period | 1450 CE |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Corcia, T.; Scano, G.; Martínez-Labarga, C.; Sarno, S.; De Fanti, S.; Luiselli, D.; Rickards, O. Uniparental Lineages from the Oldest Indigenous Population of Ecuador: The Tsachilas. Genes 2021, 12, 1273. https://doi.org/10.3390/genes12081273
Di Corcia T, Scano G, Martínez-Labarga C, Sarno S, De Fanti S, Luiselli D, Rickards O. Uniparental Lineages from the Oldest Indigenous Population of Ecuador: The Tsachilas. Genes. 2021; 12(8):1273. https://doi.org/10.3390/genes12081273
Chicago/Turabian StyleDi Corcia, Tullia, Giuseppina Scano, Cristina Martínez-Labarga, Stefania Sarno, Sara De Fanti, Donata Luiselli, and Olga Rickards. 2021. "Uniparental Lineages from the Oldest Indigenous Population of Ecuador: The Tsachilas" Genes 12, no. 8: 1273. https://doi.org/10.3390/genes12081273
APA StyleDi Corcia, T., Scano, G., Martínez-Labarga, C., Sarno, S., De Fanti, S., Luiselli, D., & Rickards, O. (2021). Uniparental Lineages from the Oldest Indigenous Population of Ecuador: The Tsachilas. Genes, 12(8), 1273. https://doi.org/10.3390/genes12081273