
Genomic Characterization of Fluoroquinolone-Resistant Thermophilic Campylobacter Strains Isolated from Layer Chicken Feces in Gangneung, South Korea by Whole-Genome Sequencing

 ,
,  , , , and
, , , and 
Abstract
1. Introduction
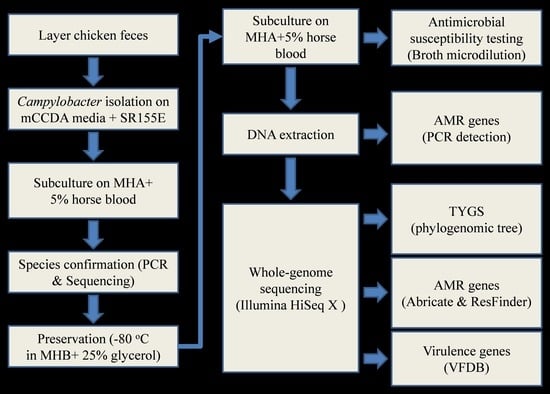
2. Materials and Methods
2.1. Campylobacter Strains and Culture Conditions
2.2. Antimicrobial Susceptibility Testing
2.3. DNA Extraction, Species Confirmation, and Antimicrobial Resistance (AMR) Genes Detection
2.4. Whole-Genome Sequencing
2.5. Construction of Phylogenetic Tree
2.6. Data Analysis
3. Results
3.1. Antimicrobial Resistance Profiles
3.2. Whole-Genome Sequencing Data
3.3. Virulence Genes
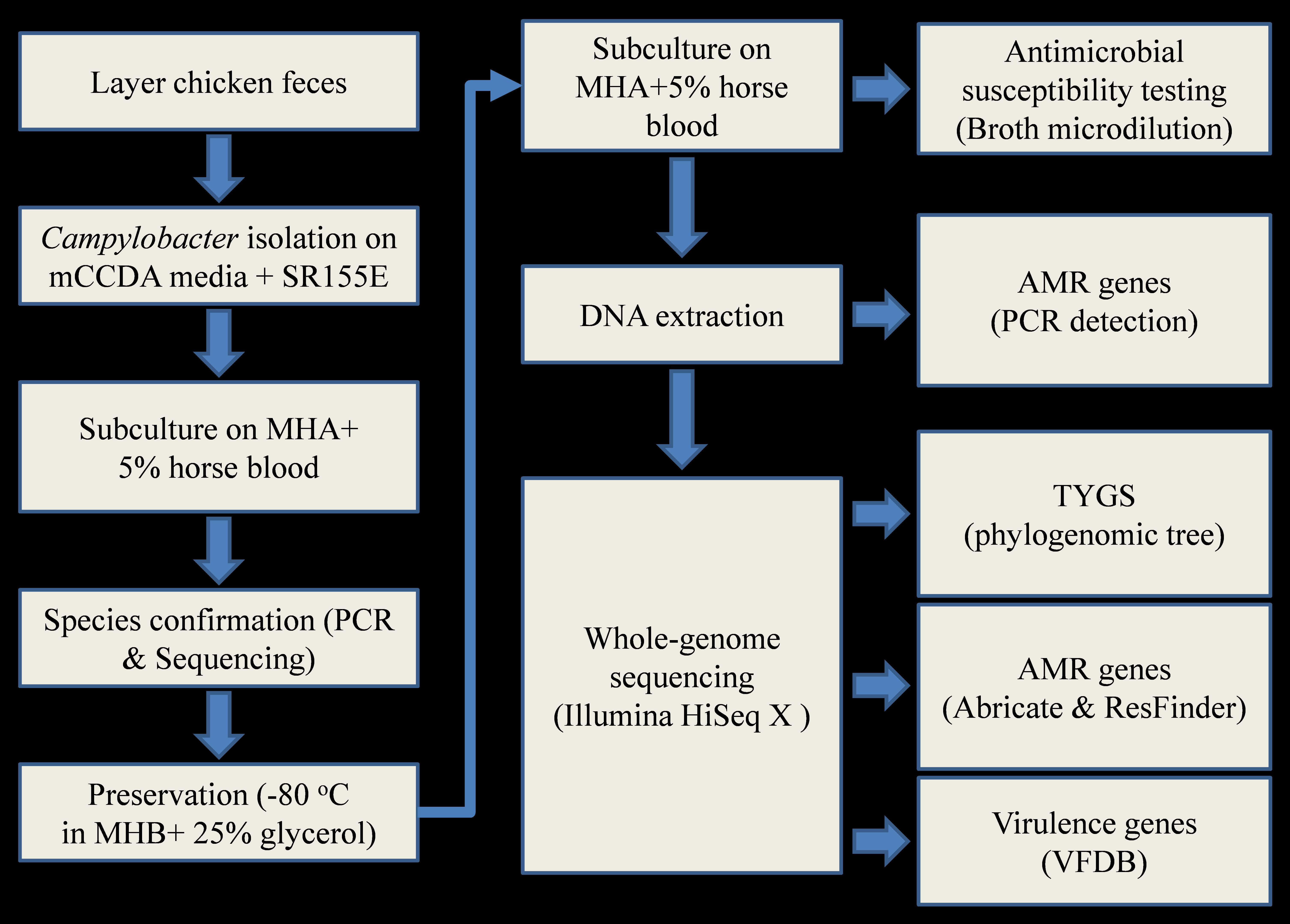
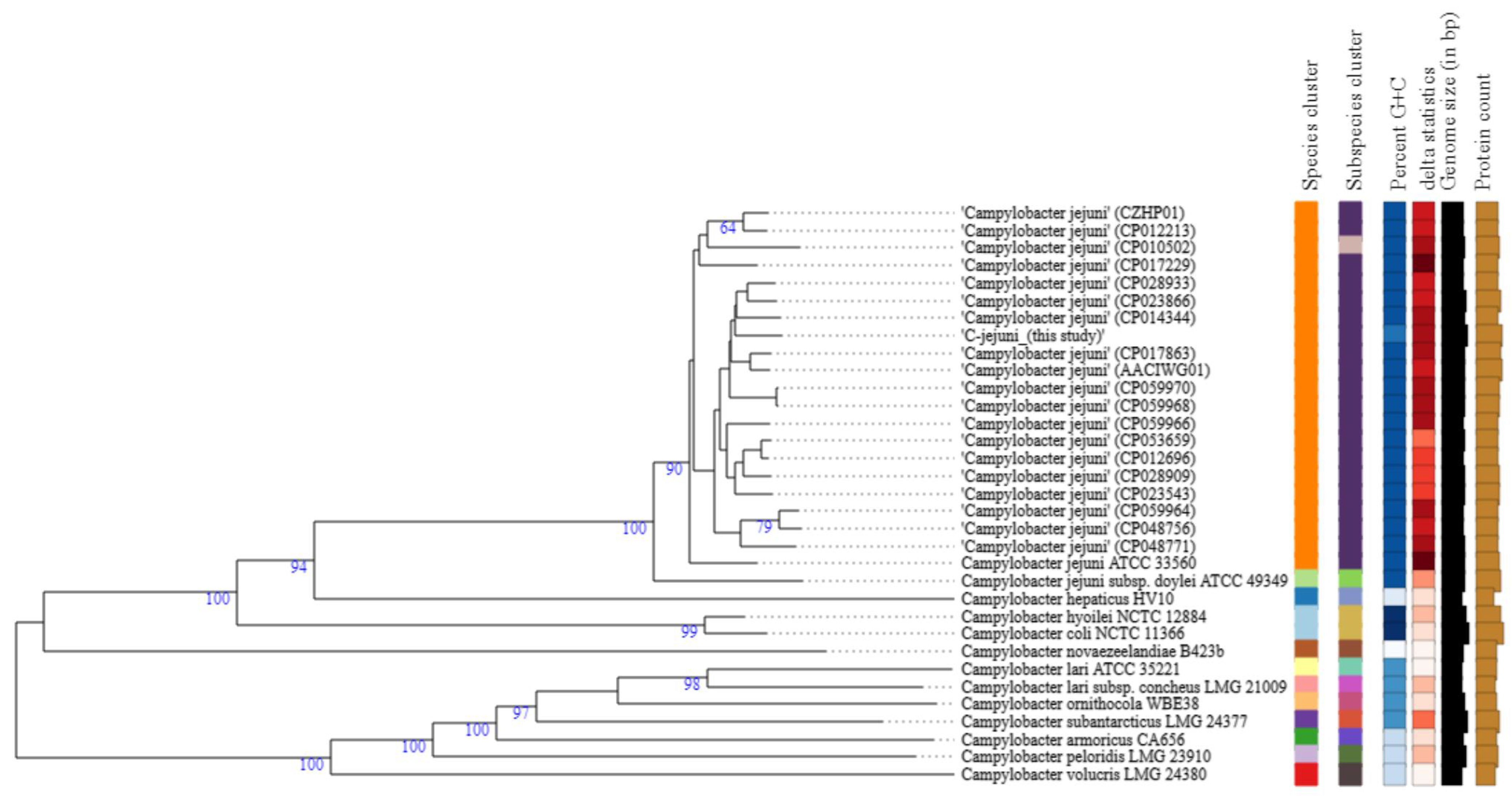
3.4. Phylogenetic Analysis
3.5. Multilocus Sequence Typing (MLST)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef]
- Ghatak, S.; He, Y.; Reed, S.; Irwin, P. Comparative Genomic Analysis of a Multidrug-Resistant Campylobacter jejuni Strain YH002 Isolated from Retail Beef Liver. Foodborne Pathog. Dis. 2020, 17, 576–584. [Google Scholar] [CrossRef]
- Elhadidy, M.; Miller, W.G.; Arguello, H.; Alvarez-Ordóñez, A.; Dierick, K.; Botteldoorn, N. Molecular epidemiology and antimicrobial resistance mechanisms of Campylobacter coli from diarrhoeal patients and broiler carcasses in Belgium. Transbound. Emerg. Dis. 2019, 66, 463–475. [Google Scholar] [CrossRef]
- Humphrey, T.; O’Brien, S.J.; Madsen, M. Campylobacters as zoonotic pathogens: A food production perspective. Int. J. Food Microbiol. 2007, 117, 237–257. [Google Scholar] [CrossRef] [PubMed]
- Shyaka, A.; Kusumoto, A.; Asakura, H.; Kawamoto, K. Whole-Genome Sequences of Eight Campylobacter jejuni Isolates from Wild Birds. Genome Announc. 2015, 3, e00315-15. [Google Scholar] [CrossRef]
- Epping, L.; Golz, J.C.; Knüver, M.-T.; Huber, C.; Thürmer, A.; Wieler, L.H.; Stingl, K.; Semmler, T. Comparison of different technologies for the decipherment of the whole genome sequence of Campylobacter jejuni BfR-CA-14430. Gut Pathog. 2019, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kassem, I.I.; Kehinde, O.; Kumar, A.; Rajashekara, G. Antimicrobial-Resistant Campylobacter in Organically and Conventionally Raised Layer Chickens. Foodborne Pathog. Dis. 2017, 14, 29–34. [Google Scholar] [CrossRef]
- Gahamanyi, N.; Song, D.-G.; Cha, K.H.; Yoon, K.-Y.; Mboera, L.E.; Matee, M.I.; Mutangana, D.; Amachawadi, R.G.; Komba, E.V.; Pan, C.-H. Susceptibility of Campylobacter Strains to Selected Natural Products and Frontline Antibiotics. Antibiotics 2020, 9, 790. [Google Scholar] [CrossRef]
- Sproston, E.L.; Wimalarathna, H.M.L.; Sheppard, S.K. Trends in fluoroquinolone resistance in Campylobacter. Microb. Genom. 2018, 4, e000198. [Google Scholar] [CrossRef] [PubMed]
- Cha, W.; Emosci, R.; Wengert, S.L.; Singh, P.; Newton, D.W.; Esalimnia, H.; Lephart, P.; Ekhalife, W.; Mansfield, L.S.; Rudrik, J.T.; et al. Antimicrobial Susceptibility Profiles of Human Campylobacter jejuni Isolates and Association with Phylogenetic Lineages. Front. Microbiol. 2016, 7, 589. [Google Scholar] [CrossRef] [PubMed]
- Kovács, J.K.; Cox, A.; Schweitzer, B.; Maróti, G.; Kovács, T.; Fenyvesi, H.; Emődy, L.; Schneider, G. Virulence Traits of Inpatient Campylobacter jejuni Isolates, and a Transcriptomic Approach to Identify Potential Genes Maintaining Intracellular Survival. Microorganisms 2020, 8, 531. [Google Scholar] [CrossRef] [PubMed]
- Rokney, A.; Valinsky, L.; Moran-Gilad, J.; Vranckx, K.; Agmon, V.; Weinberger, M. Genomic Epidemiology of Campylobacter jejuni Transmission in Israel. Front. Microbiol. 2018, 9, 2432. [Google Scholar] [CrossRef] [PubMed]
- Kreling, V.; Falcone, F.H.; Kehrenberg, C.; Hensel, A. Campylobacter sp.: Pathogenicity factors and prevention methods—new molecular targets for innovative antivirulence drugs? Appl. Microbiol. Biotechnol. 2020, 104, 10409–10436. [Google Scholar] [CrossRef]
- Nguyen, T.N.M.; Hotzel, H.; El-Adawy, H.; Tran, H.T.; Le, M.T.H.; Tomaso, H.; Neubauer, H.; Hafez, H.M. Genotyping and antibiotic resistance of thermophilic Campylobacter isolated from chicken and pig meat in Vietnam. Gut Pathog. 2016, 8, 1–11. [Google Scholar] [CrossRef]
- Bolton, D.J. Campylobacter virulence and survival factors. Food Microbiol. 2015, 48, 99–108. [Google Scholar] [CrossRef]
- Yamasaki, S.; Asakura, M.; Tsukamoto, T.; Faruque, S.M.; Deb, R.; Ramamurthy, T. Cytolethal Distending Toxin (CDT): Genetic Diversity, Structure and Role In Diarrheal Disease. Toxin Rev. 2006, 25, 61–88. [Google Scholar] [CrossRef]
- Ramires, T.; de Oliveira, M.G.; Kleinubing, N.R.; Würfel, S.D.F.R.; Mata, M.M.; Iglesias, M.A.; Lopes, G.V.; Dellagostin, O.A.; da Silva, W.P. Genetic diversity, antimicrobial resistance, and virulence genes of thermophilic Campylobacter isolated from broiler production chain. Braz. J. Microbiol. 2020, 51, 2021–2032. [Google Scholar] [CrossRef] [PubMed]
- Harrington, C.; Moran, L.; Ridley, A.; Newell, D.; Madden, R. Inter-laboratory evaluation of three flagellin PCR/RFLP methods for typing Campylobacter jejuni and C. coli: The CAMPYNET experience. J. Appl. Microbiol. 2003, 95, 1321–1333. [Google Scholar] [CrossRef]
- Elhadidy, M.; Miller, W.G.; Arguello, H.; Alvarez-Ordóñez, A.; Duarte, A.; Dierick, K.; Botteldoorn, N. Genetic Basis and Clonal Population Structure of Antibiotic Resistance in Campylobacter jejuni Isolated From Broiler Carcasses in Belgium. Front. Microbiol. 2018, 9, 1014. [Google Scholar] [CrossRef] [PubMed]
- Fiedoruk, K.; Daniluk, T.; Rozkiewicz, D.; Oldak, E.; Prasad, S.; Swiecicka, I. Whole-genome comparative analysis of Campylobacter jejuni strains isolated from patients with diarrhea in northeastern Poland. Gut Pathog. 2019, 11, 1–10. [Google Scholar] [CrossRef]
- Iraola, G.; Pérez, R.; Naya, H.; Paolicchi, F.; Pastor, E.; Valenzuela, S.; Calleros, L.; Velilla, A.; Hernández, M.; Morsella, C. Genomic Evidence for the Emergence and Evolution of Pathogenicity and Niche Preferences in the Genus Campylobacter. Genome Biol. Evol. 2014, 6, 2392–2405. [Google Scholar] [CrossRef]
- Golz, J.C.; Epping, L.; Knüver, M.-T.; Borowiak, M.; Hartkopf, F.; Deneke, C.; Malorny, B.; Semmler, T.; Stingl, K. Whole genome sequencing reveals extended natural transformation in Campylobacter impacting diagnostics and the pathogens adaptive potential. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Llarena, A.-K.; Taboada, E.; Rossi, M. Whole-Genome Sequencing in Epidemiology of Campylobacter jejuni Infections. J. Clin. Microbiol. 2017, 55, 1269–1275. [Google Scholar] [CrossRef]
- Maiden, M.C.J.; Van Rensburg, M.J.J.; Bray, J.; Earle, S.G.; Ford, S.A.; Jolley, K.; McCarthy, N.D. MLST revisited: The gene-by-gene approach to bacterial genomics. Nat. Rev. Genet. 2013, 11, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Bogaerts, B.; Winand, R.; Fu, Q.; Van Braekel, J.; Ceyssens, P.-J.; Mattheus, W.; Bertrand, S.; De Keersmaecker, S.C.J.; Roosens, N.H.C.; Vanneste, K. Validation of a Bioinformatics Workflow for Routine Analysis of Whole-Genome Sequencing Data and Related Challenges for Pathogen Typing in a European National Reference Center: Neisseria meningitidis as a Proof-of-Concept. Front. Microbiol. 2019, 10, 362. [Google Scholar] [CrossRef] [PubMed]
- Köser, C.U.; Ellington, M.J.; Peacock, S.J. Whole-genome sequencing to control antimicrobial resistance. Trends Genet. 2014, 30, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Oakeson, K.F.; Wagner, J.M.; Rohrwasser, A.; Atkinson-Dunn, R. Whole-Genome Sequencing and Bioinformatic Analysis of Isolates from Foodborne Illness Outbreaks of Campylobacter jejuni and Salmonella enterica. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [PubMed]
- Quail, M.A.; Smith, M.E.; Coupland, P.; Otto, T.D.; Harris, S.R.; Connor, T.R.; Bertoni, A.; Swerdlow, H.P.; Gu, Y. A tale of three next generation sequencing platforms: Comparison of Ion Torrent, Pacific Biosciences and Illumina MiSeq sequencers. BMC Genom. 2012, 13, 341. [Google Scholar] [CrossRef]
- Fricke, W.F.; Rasko, D.A. Bacterial genome sequencing in the clinic: Bioinformatic challenges and solutions. Nat. Rev. Genet. 2014, 15, 49–55. [Google Scholar] [CrossRef]
- García-Sánchez, L.; Melero, B.; Jaime, I.; Rossi, M.; Ortega, I.; Rovira, J. Biofilm formation, virulence and antimicrobial resistance of different Campylobacter jejuni isolates from a poultry slaughterhouse. Food Microbiol. 2019, 83, 193–199. [Google Scholar] [CrossRef]
- Agunos, A.; Waddell, L.; Léger, D.; Taboada, E. A Systematic Review Characterizing On-Farm Sources of Campylobacter spp. for Broiler Chickens. PLoS ONE 2014, 9, e104905. [Google Scholar] [CrossRef]
- Rama, E.N.; Bailey, M.; Jones, D.R.; Gast, R.; Anderson, K.; Brar, J.; Taylor, R.; Oliver, H.F.; Singh, M. Prevalence, Persistence, and Antimicrobial Resistance of Campylobacter spp. from Eggs and Laying Hens Housed in Five Commercial Housing Systems. Foodborne Pathog. Dis. 2018, 15, 506–516. [Google Scholar] [CrossRef]
- Kurekci, C.; Bishop-Hurley, S.L.; Vercoe, P.; Durmic, Z.; Al Jassim, R.A.M.; McSweeney, C.S. Screening of Australian Plants for Antimicrobial Activity against Campylobacter jejuni. Phytother. Res. 2011, 26, 186–190. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef]
- Ondov, B.D.; Treangen, T.J.; Melsted, P.; Mallonee, A.B.; Bergman, N.H.; Koren, S.; Phillippy, A.M. Mash: Fast genome and metagenome distance estimation using MinHash. Genome Biol. 2016, 17, 132. [Google Scholar] [CrossRef]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Staerfeldt, H.-H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.S.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A Comprehensive, Accurate, and Fast Distance-Based Phylogeny Inference Program: Table 1. Mol. Biol. Evol. 2015, 32, 2798–2800. [Google Scholar] [CrossRef]
- Farris, J.S. Estimating Phylogenetic Trees from Distance Matrices. Am. Nat. 1972, 106, 645–668. [Google Scholar] [CrossRef]
- Kreft, Ł.; Botzki, A.; Coppens, F.; Vandepoele, K.; Van Bel, M. PhyD3: A phylogenetic tree viewer with extended phyloXML support for functional genomics data visualization. Bioinformatics 2017, 33, 2946–2947. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Hahnke, R.L.; Petersen, J.; Scheuner, C.; Michael, V.; Fiebig, A.; Rohde, C.; Rohde, M.; Fartmann, B.; Goodwin, L.A.; et al. Complete genome sequence of DSM 30083T, the type strain (U5/41T) of Escherichia coli, and a proposal for delineating subspecies in microbial taxonomy. Stand. Genom. Sci. 2014, 9, 1–19. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed]
- Tarasov, A.; Vilella, A.J.; Cuppen, E.; Nijman, I.J.; Prins, P. Sambamba: Fast processing of NGS alignment formats. Bioinformatics 2015, 31, 2032–2034. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 genome project data processing subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Souvorov, A.; Agarwala, R.; Lipman, D.J. SKESA: Strategic k-mer extension for scrupulous assemblies. Genome Biol. 2018, 19, 1–13. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Feldgarden, M. Using the NCBI AMRFinder Tool to Determine Antimicrobial Resistance Genotype-Phenotype Correlations Within a Collection of NARMS Isolates. bioRxiv 2019, 550707. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Sproston, E.; Ogden, I.; Macrae, M.; Forbes, K.; Dallas, J.; Sheppard, S.; Cody, A.; Colles, F.; Wilson, M.; Strachan, N. Multi-locus sequence types of Campylobacter carried by flies and slugs acquired from local ruminant faeces. J. Appl. Microbiol. 2010, 109, 829–838. [Google Scholar] [CrossRef]
- Wei, B.; Cha, S.-Y.; Kang, M.; Roh, J.-H.; Seo, H.-S.; Yoon, R.-H.; Jang, H.-K. Antimicrobial Susceptibility Profiles and Molecular Typing of Campylobacter jejuni and Campylobacter coli Isolates from Ducks in South Korea. Appl. Environ. Microbiol. 2014, 80, 7604–7610. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.-Y.; Kwon, Y.-K.; Wei, B.; Jang, H.-K.; Lim, S.-K.; Kim, C.-H.; Jung, S.-C.; Kang, M.-S. Epidemiological relationships of Campylobacter jejuni strains isolated from humans and chickens in South Korea. J. Microbiol. 2016, 55, 13–20. [Google Scholar] [CrossRef]
- Tang, Y.; Fang, L.; Xu, C.; Zhang, Q. Antibiotic resistance trends and mechanisms in the foodborne pathogen, Campylobacter. Anim. Health Res. Rev. 2017, 18, 87–98. [Google Scholar] [CrossRef]
- Abraham, S.; Sahibzada, S.; Hewson, K.; Laird, T.; Abraham, R.; Pavic, A.; Truswell, A.; Lee, T.; O’Dea, M.; Jordan, D. Emergence of Fluoroquinolone-Resistant Campylobacter jejuni and Campylobacter coli among Australian Chickens in the Absence of Fluoroquinolone Use. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef] [PubMed]
- Lynch, C.T.; Lynch, H.; Burke, S.; Hawkins, K.; Buttimer, C.; Mc Carthy, C.; Egan, J.; Whyte, P.; Bolton, D.; Coffey, A.; et al. Antimicrobial Resistance Determinants Circulating among Thermophilic Campylobacter Isolates Recovered from Broilers in Ireland Over a One-Year Period. Antibiotics 2020, 9, 308. [Google Scholar] [CrossRef] [PubMed]
- Seo, K.-S.; Jeong, H.-J.; Yun, K.-W. Antimicrobial activity and chemical components of two plants, Artemisia capillaris and Artemisia iwayomogi, used as Korean herbal Injin. J. Ecol. Environ. 2010, 33, 141–147. [Google Scholar] [CrossRef]
- Frazão, M.R.; Cao, G.; Medeiros, M.I.C.; Duque, S.D.S.; Allard, M.W.; Falcão, J.P. Antimicrobial Resistance Profiles and Phylogenetic Analysis of Campylobacter jejuni Strains Isolated in Brazil by Whole Genome Sequencing. Microb. Drug Resist. 2021, 27, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Park, H.; Kim, J.; Kim, J.H.; Jung, J.I.; Cho, S.; Ryu, S.; Jeon, B. Comparative Analysis of Aerotolerance, Antibiotic Resistance, and Virulence Gene Prevalence in Campylobacter jejuni Isolates from Retail Raw Chicken and Duck Meat in South Korea. Microorganisms 2019, 7, 433. [Google Scholar] [CrossRef] [PubMed]
- Hlashwayo, D.F.; Barbosa, F.; Langa, S.; Sigaúque, B.; Bila, C.G. A Systematic Review of In Vitro Activity of Medicinal Plants from Sub-Saharan Africa against Campylobacter Spp. Evid.-Based Complementary Altern. Med. 2020, 1–13. [Google Scholar] [CrossRef]
- Gahamanyi, N.; Mboera, L.E.G.; Matee, M.I.; Mutangana, D.; Komba, E.V.G. Prevalence, Risk Factors, and Antimicrobial Resistance Profiles of Thermophilic Campylobacter Species in Humans and Animals in Sub-Saharan Africa: A Systematic Review. Int. J. Microbiol. 2020, 1–12. [Google Scholar] [CrossRef]
- Iovine, N.M. Resistance mechanisms in Campylobacter jejuni. Virulence 2013, 4, 230–240. [Google Scholar] [CrossRef]
- Kashoma, I.P.; Kassem, I.; John, J.; Kessy, B.M.; Gebreyes, W.; Kazwala, R.R.; Rajashekara, G. Prevalence and Antimicrobial Resistance of Campylobacter Isolated from Dressed Beef Carcasses and Raw Milk in Tanzania. Microb. Drug Resist. 2016, 22, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Asakura, H.; Sakata, J.; Nakamura, H.; Yamamoto, S.; Murakami, S. Phylogenetic Diversity and Antimicrobial Resistance of Campylobacter coli from Humans and Animals in Japan. Microbes Environ. 2019, 34, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Hlashwayo, D.F.; Sigaúque, B.; Noormahomed, E.V.; Afonso, S.M.S.; Mandomando, I.M.; Bila, C.G. A systematic review and meta-analysis reveal that Campylobacter spp. and antibiotic resistance are widespread in humans in sub-Saharan Africa. PLoS ONE 2021, 16, e0245951. [Google Scholar] [CrossRef]
- Kabir, S.L.; Asakura, M.; Shiramaru, S.; Pal, A.; Hinenoya, A.; Yamasaki, S. Molecular identification and antimicrobial resistance profiles of Campylobacter strains of poultry origin in India with special emphasis on fluoroquinolone resistance. Asian J. Med. Biol. Res. 2015, 1, 1–8. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Y.; Shao, Y.; Hu, Y.; Lou, H.; Chen, X.; Wu, Y.; Mei, L.; Zhou, B.; Zhang, X.; et al. Molecular Characterization and Antibiotic Resistant Profiles of Campylobacter Species Isolated From Poultry and Diarrheal Patients in Southeastern China 2017–2019. Front. Microbiol. 2020, 11, 1244. [Google Scholar] [CrossRef]
- Jonker, A.; Picard, J. Antimicrobial susceptibility in thermophilic Campylobacter species isolated from pigs and chickens in South Africa. J. S. Afr. Vet. Assoc. 2010, 81, 228–236. [Google Scholar] [CrossRef]
- Cuevas, E.; Guirado, P.; Miro, E.; Iglesias-Torrens, Y.; Navarro, F.; Alioto, T.; Gómez-Garrido, J.; Madrid, C.; Balsalobre, C. Tetracycline resistance transmission in Campylobacter is promoted at temperatures resembling the avian reservoir. Vet. Microbiol. 2020, 244, 108652. [Google Scholar] [CrossRef]
- De Vries, S.P.W.; Vurayai, M.; Holmes, M.; Gupta, S.; Bateman, M.; Goldfarb, D.; Maskell, D.J.; Matsheka, M.I.; Grant, A.J. Phylogenetic analyses and antimicrobial resistance profiles of Campylobacter spp. from diarrhoeal patients and chickens in Botswana. PLoS ONE 2018, 13, e0194481. [Google Scholar] [CrossRef]
- Mäesaar, M.; Kramarenko, T.; Meremäe, K.; Sõgel, J.; Lillenberg, M.; Häkkinen, L.; Ivanova, M.; Kovalenko, K.; Hörman, A.; Hanninen, M.-L.; et al. Antimicrobial Resistance Profiles of Campylobacter spp. Isolated from Broiler Chicken Meat of Estonian, Latvian and Lithuanian Origin at Estonian Retail Level and from Patients with Severe Enteric Infections in Estonia. Zoonoses Public Health 2016, 63, 89–96. [Google Scholar] [CrossRef]
- Cantero, G.; Correa-Fiz, F.; Ronco, T.; Strube, M.L.; Cerdà-Cuéllar, M.; Pedersen, K. Characterization of Campylobacter jejuni and Campylobacter coli Broiler Isolates by Whole-Genome Sequencing. Foodborne Pathog. Dis. 2018, 15, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Pillay, S.; Amoako, D.G.; Abia, A.L.K.; Somboro, A.M.; Shobo, C.O.; Perrett, K.; Bester, L.A.; Essack, S.Y. Characterisation of Campylobacter spp. Isolated from Poultry in KwaZulu-Natal, South Africa. Antibiotics 2020, 9, 42. [Google Scholar] [CrossRef]
- Luangtongkum, T.; Jeon, B.; Han, J.; Plummer, P.; Logue, C.M.; Zhang, Q. Antibiotic resistance in Campylobacter: Emergence, transmission and persistence. Futur. Microbiol. 2009, 4, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Long, C.; Xu, J.; Guo, Q.; Zhang, W.; Zhang, Z.; Bater. Are dominant plant species more susceptible to leaf-mining insects? A case study at Saihanwula Nature Reserve, China. Ecol. Evol. 2018, 8, 7633–7648. [Google Scholar] [CrossRef] [PubMed]
- Lapierre, L.; Gatica, M.A.; Riquelme, V.; Vergara, C.; Yañez, J.M.; Martín, B.S.; Sáenz, L.; Vidal, M.; Martínez, M.C.; Araya, P.; et al. Characterization of Antimicrobial Susceptibility and Its Association with Virulence Genes Related to Adherence, Invasion, and Cytotoxicity in Campylobacter jejuni and Campylobacter coli Isolates from Animals, Meat, and Humans. Microb. Drug Resist. 2016, 22, 432–444. [Google Scholar] [CrossRef] [PubMed]
- Bang, D.D.; Scheutz, F.; Pedersen, K.; Handberg, K.; Madsen, M. PCR detection of seven virulence and toxin genes of Campylobacter jejuni and Campylobacter coli isolates from Danish pigs and cattle and cytolethal distending toxin production of the isolates. J. Appl. Microbiol. 2003, 94, 1003–1014. [Google Scholar] [CrossRef]
- Wysok, B.; Wojtacka, J.; Kivistö, R. Pathogenicity of Campylobacter strains of poultry and human origin from Poland. Int. J. Food Microbiol. 2020, 334, 108830. [Google Scholar] [CrossRef]
- Konkel, M.E.; Klena, J.D.; Rivera-Amill, V.; Monteville, M.R.; Biswas, D.; Raphael, B.; Mickelson, J. Secretion of Virulence Proteins from Campylobacter jejuni Is Dependent on a Functional Flagellar Export Apparatus. J. Bacteriol. 2004, 186, 3296–3303. [Google Scholar] [CrossRef] [PubMed]
- Guerry, P.; Ewing, C.P.; Schirm, M.; Lorenzo, M.; Kelly, J.; Pattarini, D.; Majam, G.; Thibault, P.; Logan, S. Changes in flagellin glycosylation affect Campylobacter autoagglutination and virulence. Mol. Microbiol. 2006, 60, 299–311. [Google Scholar] [CrossRef]
- Hermansen, G.; Boysen, A.; Krogh, T.; Nawrocki, A.; Jelsbak, L.; Møller-Jensen, J. HldE Is Important for Virulence Phenotypes in Enterotoxigenic Escherichia coli. Front. Cell. Infect. Microbiol. 2018, 8, 253. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Elbediwi, M.; Zhou, X.; Shuai, H.; Lou, X.; Wang, H.; Li, Y.; Yue, M. Epidemiological and Genomic Characterization of Campylobacter jejuni Isolates from a Foodborne Outbreak at Hangzhou, China. Int. J. Mol. Sci. 2020, 21, 3001. [Google Scholar] [CrossRef]
- Sheppard, S.K.; McCarthy, N.D.; Jolley, K.; Maiden, M. Introgression in the genus Campylobacter: Generation and spread of mosaic alleles. Microbiology 2011, 157, 1066–1074. [Google Scholar] [CrossRef]
- Okamura, M.; Kaneko, M.; Ojima, S.; Sano, H.; Shindo, J.; Shirafuji, H.; Yamamoto, S.; Tanabe, T.; Yoshikawa, Y.; Hu, D.-L. Differential Distribution of Salmonella Serovars and Campylobacter spp. Isolates in Free-Living Crows and Broiler Chickens in Aomori, Japan. Microbes Environ. 2018, 33, 77–82. [Google Scholar] [CrossRef] [PubMed]
- El-Hamid, M.I.A.; El-Aziz, N.K.A.; Samir, M.; El-Naenaeey, E.-S.Y.; Remela, E.M.A.; Mosbah, R.A.; Bendary, M.M. Genetic Diversity of Campylobacter jejuni Isolated From Avian and Human Sources in Egypt. Front. Microbiol. 2019, 10, 2353. [Google Scholar] [CrossRef] [PubMed]
- Carrique-Mas, J.J.; Bryant, J.; Cuong, N.V.; Hoang, N.V.M.; Campbell, J.; Dung, T.T.N.; Duy, D.; Hoa, N.T.; Thompson, C.; Hien, V.V.; et al. An epidemiological investigation of Campylobacter in pig and poultry farms in the Mekong delta of Vietnam. Epidemiol. Infect. 2013, 142, 1425–1436. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Zhang, X.; Xue, F.; Wang, Y.; Jiang, L.; Jiang, Y. Phenotypic Characters and Molecular Epidemiology of Campylobacter Jejuniin East China. J. Food Sci. 2015, 81, M106–M113. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Target Gene | Direction | Sequence (5′-3′) | Amplicon Size | Annealing Temperature (°C) | Reference |
|---|---|---|---|---|---|
| 16S rRNA | Forward | GGATGACACTTTTCGGAGC | 816 | 55 | [8] |
| Reverse | CATTGTAGCACGTGTGTC | ||||
| cj0414 | Forward | CAAATAAAGTTAGAGGTAGAATGT | 161 | ||
| Reverse | CCATAAGCACTAGCTAGCTGAT | ||||
| ask | Forward | GGTATGATTTCTACAAAGCGAG | 502 | ||
| Reverse | ATAAAAGACTATCGTCGCGTG | ||||
| tet(O) | Forward | GCGTTTTGTTTATGTGCG | 559 | 55 | [8] |
| Reverse | ATGGACAACCCGACAGAAG | ||||
| cjgyrA | Forward | GCCTGACGCAAGAGATGGTTTA | 454 | ||
| Reverse | TATGAGGCGGGATGTTTGTCG | ||||
| cmeB | Forward | TCCTAGCAGCACAATATG | 241 | ||
| Reverse | AGCTTCGATAGCTGCATC |
| No | Strain number | Country/Region | Sample Type | Host | Isolation Source | Disease Association |
|---|---|---|---|---|---|---|
| 1 | C. jejuni (CP059968) | China/Henan | Mixed culture | Chicken | Cloacal swab | NA |
| 2 | C. jejuni (CP012696) | USA/Albany CA | NA | Chicken | Chicken breast from retail | NA |
| 3 | C. jejuni (CP048756) | China/Zhejiang | Cell culture | Duck | Meat | NA |
| 4 | C. jejuni (AACIWG01) | USA:TX | NA | Chicken | Feces | NA |
| 5 | C. jejuni (CP012213) | Finland | NA | Human | Feces | Invasive |
| 6 | C. jejuni (CP023866) | USA:VA | NA | Chicken | Carcass | NA |
| 7 | C. jejuni (CP028909) | United Kingdom: London | Mono isolate | Chicken | NA | NA |
| 8 | C. jejuni (CP023543) | USA:CA | NA | Chicken | Chicken breast | Missing |
| 9 | C. jejuni (CP017863) | USA: Tulsa | NA | Chicken | Liver | NA |
| 10 | C. jejuni (CP014344) | South Africa: Cape Town | NA | Human | NA | Enteritis |
| 11 | C. jejuni (CP053659) | Italy:Lozzo Atesino | Mono isolate | Chicken | Feces | NA |
| 12 | C. jejuni (CP028933) | South Korea | NA | Chicken | Meat | NA |
| 13 | C. jejuni (CP059966) | China/Henan | Mixed culture | Chicken | Cloacal swab | NA |
| 14 | C. jejuni (CP048771) | China/Zhejiang | Cell culture | Duck | Meat | NA |
| 15 | C. jejuni (CZHP01) | Spain/Madrid | NA | Chicken | Meat | NA |
| 16 | C. jejuni (CP059970) | China/Henan | Mixed culture | Chicken | Cloacal swab | NA |
| 17 | C. jejuni (CP059964) | China/Henan | Mixed culture | Chicken | Cloacal swab | NA |
| 18 | C. jejuni (CP010502) | Finland | Multi-isolate | Human | Blood | Yes |
| 19 | C. jejuni (CP017229) | South Korea: Seoul | NA | Human | Stool | Food poisoning |
| C. coli | ||||||
| 1 | C. coli(CP061537) | USA: Pennsylvania | NA | Chicken | NA | NA |
| 2 | C. coli(CP023545) | USA:CA | NA | Chicken | Chicken breast | NA |
| 3 | C. coli(CP019977) | United Kingdom: Lincolnshire | Organic chicken farm | |||
| 4 | C. coli(CP027634) | Germany: Berlin | NA | Turkey | Meat | Colonization |
| 5 | C. coli(CP046317) | USA: VA | NA | Human | Gastrointestinal tract | Unknown |
| 6 | C. coli(CP017868) | USA: Tulsa | NA | Chicken | Chicken liver from retail | NA |
| 7 | C. coli(CP017873) | USA: Tulsa | NA | Chicken | Chicken liver from retail | NA |
| 8 | C. coli(CP027638) | Germany: Berlin | NA | Turkey | Meat | Colonization |
| 9 | C. coli(CP011015) | United Kingdom: Cambridge | NA | Human | Feces | NA |
| 10 | C. coli(CP035927) | USA | NA | Chicken | Carcass | NA |
| 11 | C. coli(CP018900) | USA: Wyndmoor, Pennsylvania | NA | Chicken | Carcass/Retail | NA |
| 12 | C. coli(CP058340) | USA | Cell culture | NA | Environmental | NA |
| 13 | C. coli(CP040239) | United Kingdom: Sutton Bonington | NA | Cattle | Slurry | NA |
| 14 | C. coli(CP006702) | United Kingdom | Monoisolate | Human | NA | Gastroenteritis |
| 15 | C. coli(CP038868) | China: Shanghai | NA | Chicken | Cecum | NA |
| 16 | C. coli(CP028187) | Denmark | NA | NA | Missing | NA |
| 17 | C. coli(CP017875) | USA: Tulsa | NA | Pig | Pork | NA |
| 18 | C. coli(CP007181) | Missing | NA | Turkey | Missing | Missing |
| Strain | SRA Accession No. | Reference Length | Mapped Site | Total Read | Mapped Read | Variant | GC (%) | Q30 (%) |
|---|---|---|---|---|---|---|---|---|
| C. jejuni strain 200605 | SAMN17525986 | 1,641,464 | 1,596,540 | 9,800,132 | 8,152,436 | 22,266 | 30.12 | 97.48 |
| C. coli strain 200606 | SAMN17525987 | 1,938,580 | 1,584,482 | 9,922,508 | 8,616,294 | 46,102 | 31.19 | 97.27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gahamanyi, N.; Song, D.-G.; Yoon, K.-Y.; Mboera, L.E.G.; Matee, M.I.; Mutangana, D.; Komba, E.V.G.; Pan, C.-H.; Amachawadi, R.G. Genomic Characterization of Fluoroquinolone-Resistant Thermophilic Campylobacter Strains Isolated from Layer Chicken Feces in Gangneung, South Korea by Whole-Genome Sequencing. Genes 2021, 12, 1131. https://doi.org/10.3390/genes12081131
Gahamanyi N, Song D-G, Yoon K-Y, Mboera LEG, Matee MI, Mutangana D, Komba EVG, Pan C-H, Amachawadi RG. Genomic Characterization of Fluoroquinolone-Resistant Thermophilic Campylobacter Strains Isolated from Layer Chicken Feces in Gangneung, South Korea by Whole-Genome Sequencing. Genes. 2021; 12(8):1131. https://doi.org/10.3390/genes12081131
Chicago/Turabian StyleGahamanyi, Noel, Dae-Geun Song, Kye-Yoon Yoon, Leonard E. G. Mboera, Mecky I. Matee, Dieudonné Mutangana, Erick V. G. Komba, Cheol-Ho Pan, and Raghavendra G. Amachawadi. 2021. "Genomic Characterization of Fluoroquinolone-Resistant Thermophilic Campylobacter Strains Isolated from Layer Chicken Feces in Gangneung, South Korea by Whole-Genome Sequencing" Genes 12, no. 8: 1131. https://doi.org/10.3390/genes12081131
APA StyleGahamanyi, N., Song, D.-G., Yoon, K.-Y., Mboera, L. E. G., Matee, M. I., Mutangana, D., Komba, E. V. G., Pan, C.-H., & Amachawadi, R. G. (2021). Genomic Characterization of Fluoroquinolone-Resistant Thermophilic Campylobacter Strains Isolated from Layer Chicken Feces in Gangneung, South Korea by Whole-Genome Sequencing. Genes, 12(8), 1131. https://doi.org/10.3390/genes12081131