
Abnormal Long Non-Coding RNAs Expression Patterns Have the Potential Ability for Predicting Survival and Treatment Response in Breast Cancer

 , ,
, , 
 and
and
Abstract
:1. Introduction
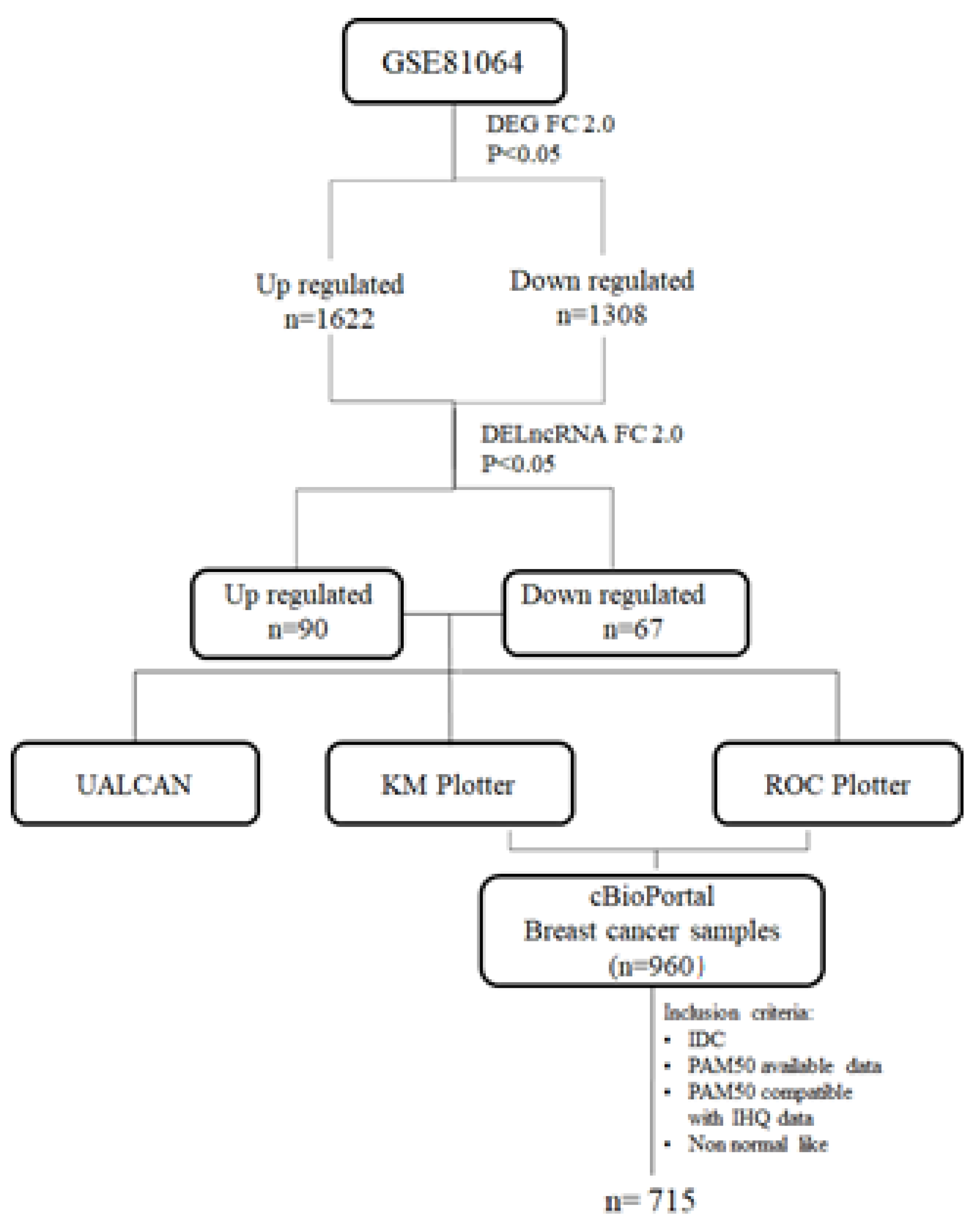
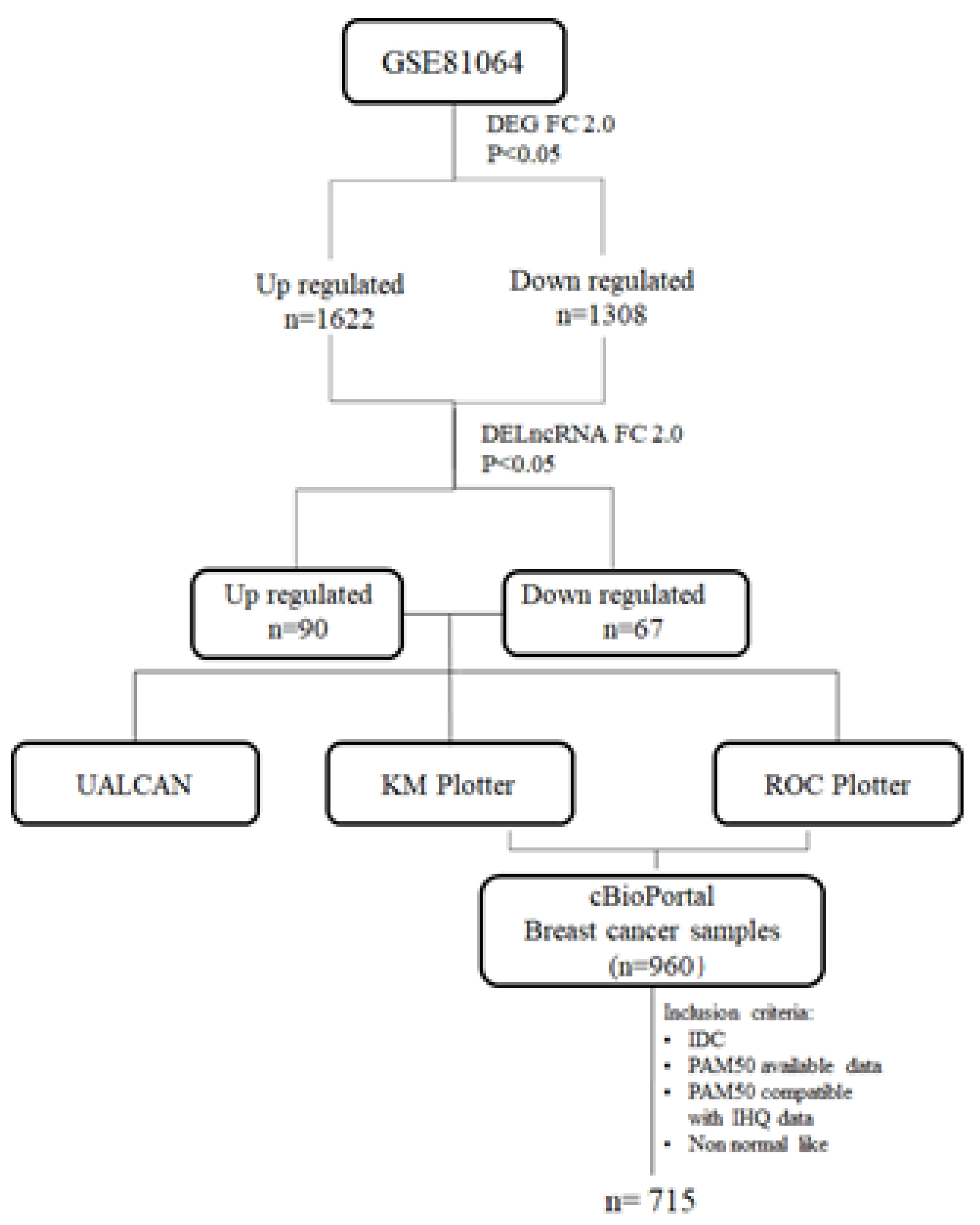
2. Materials and Methods
3. Results
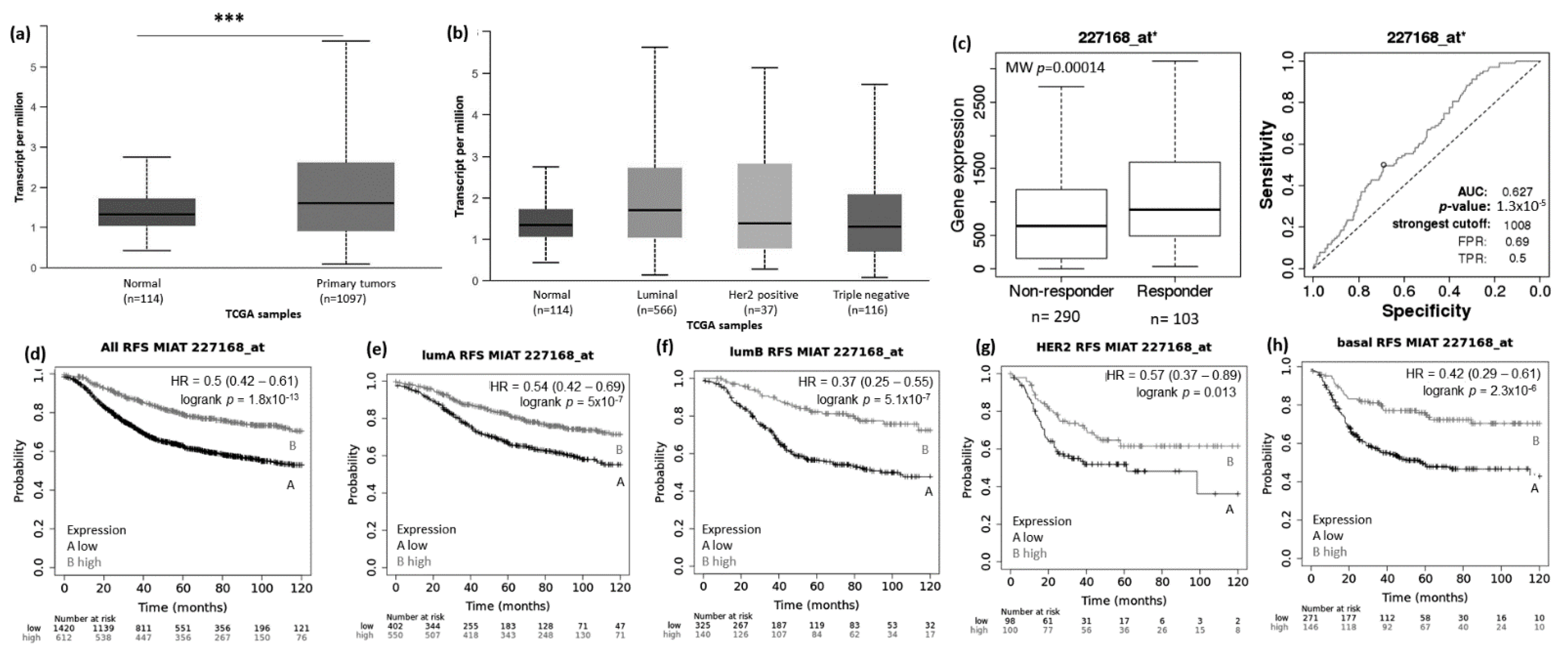
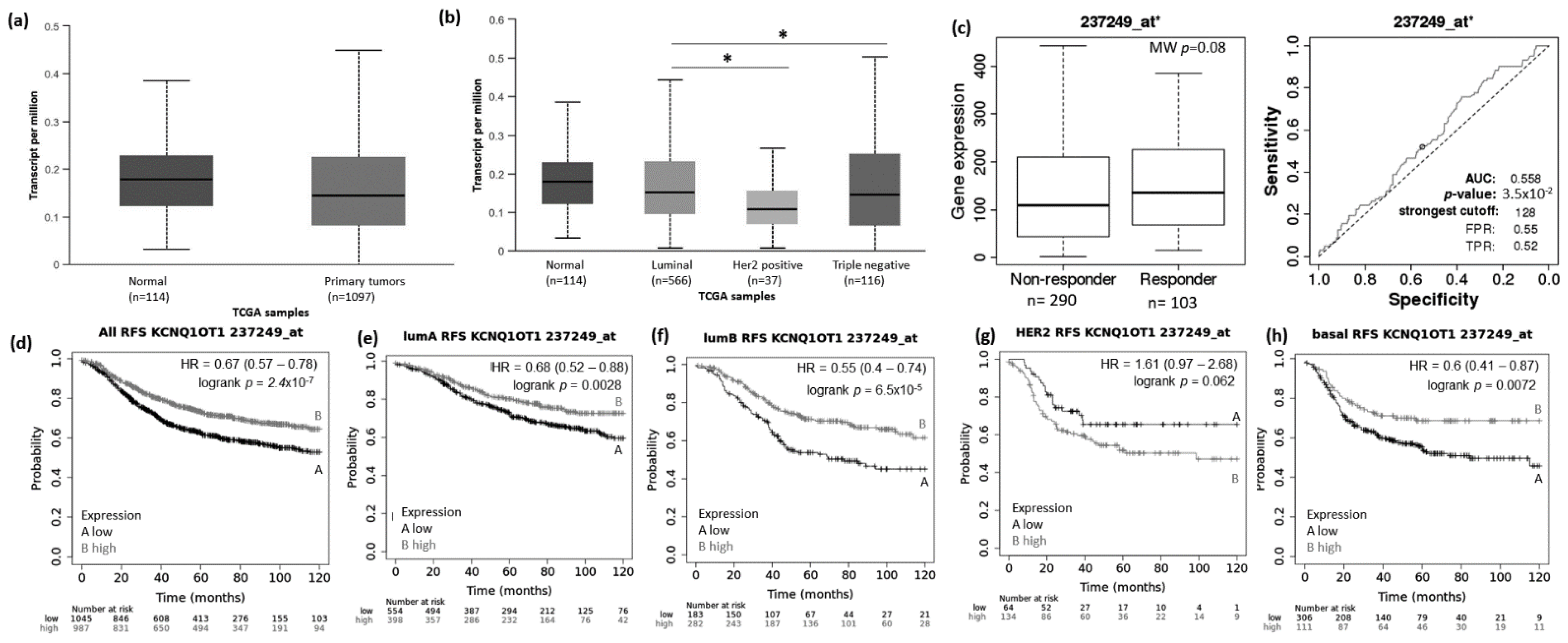
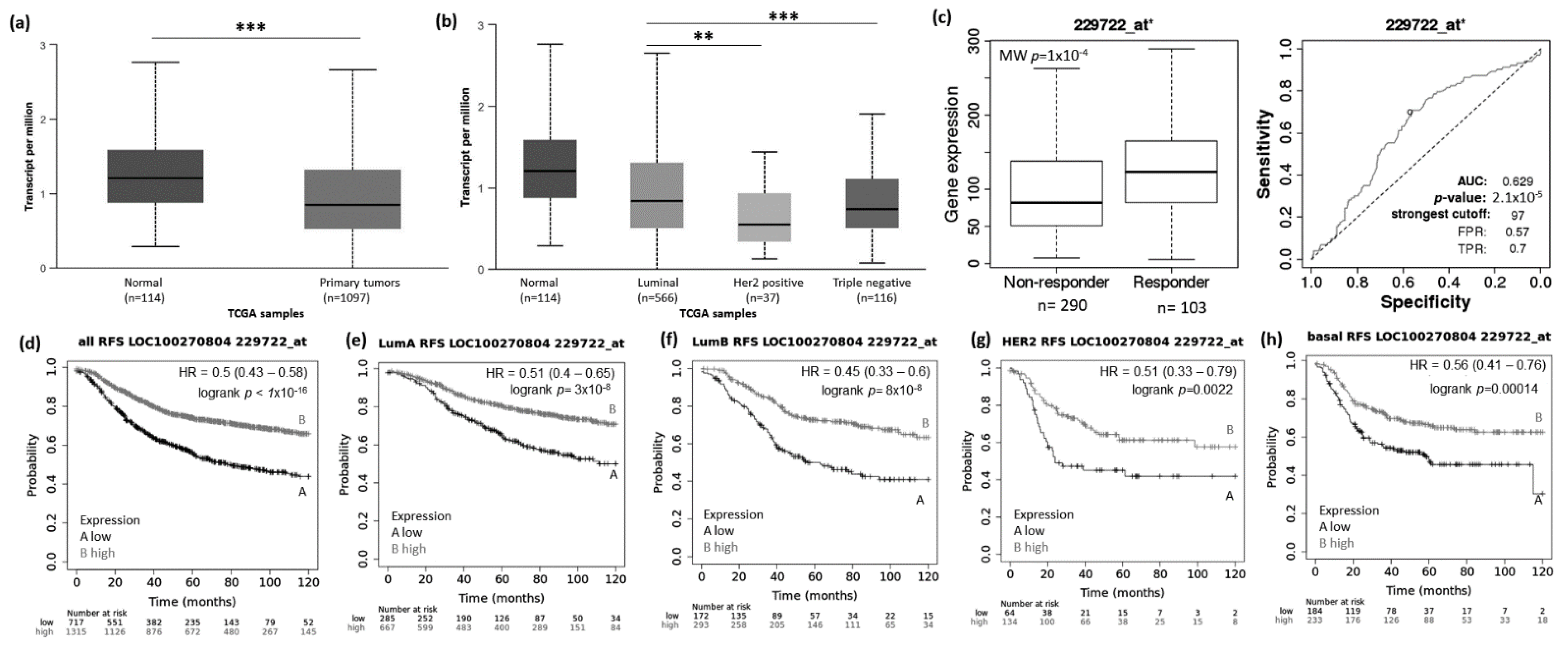
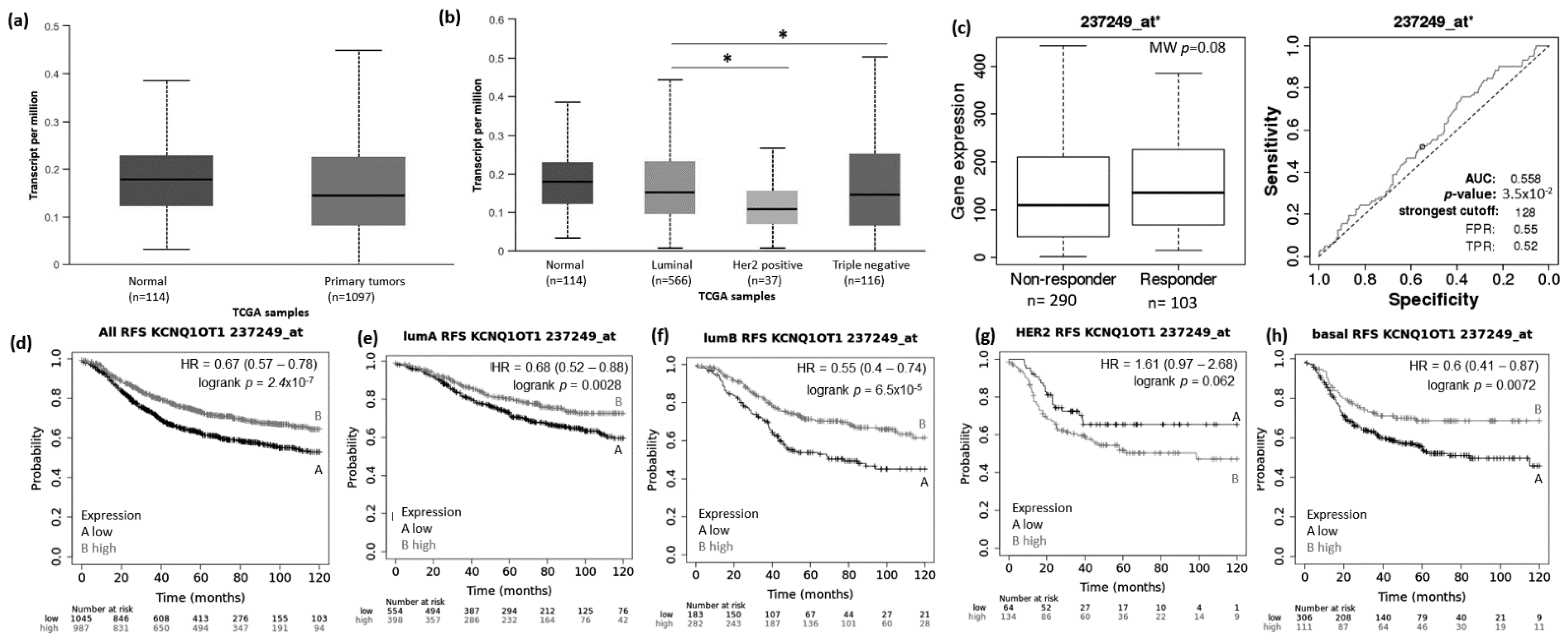
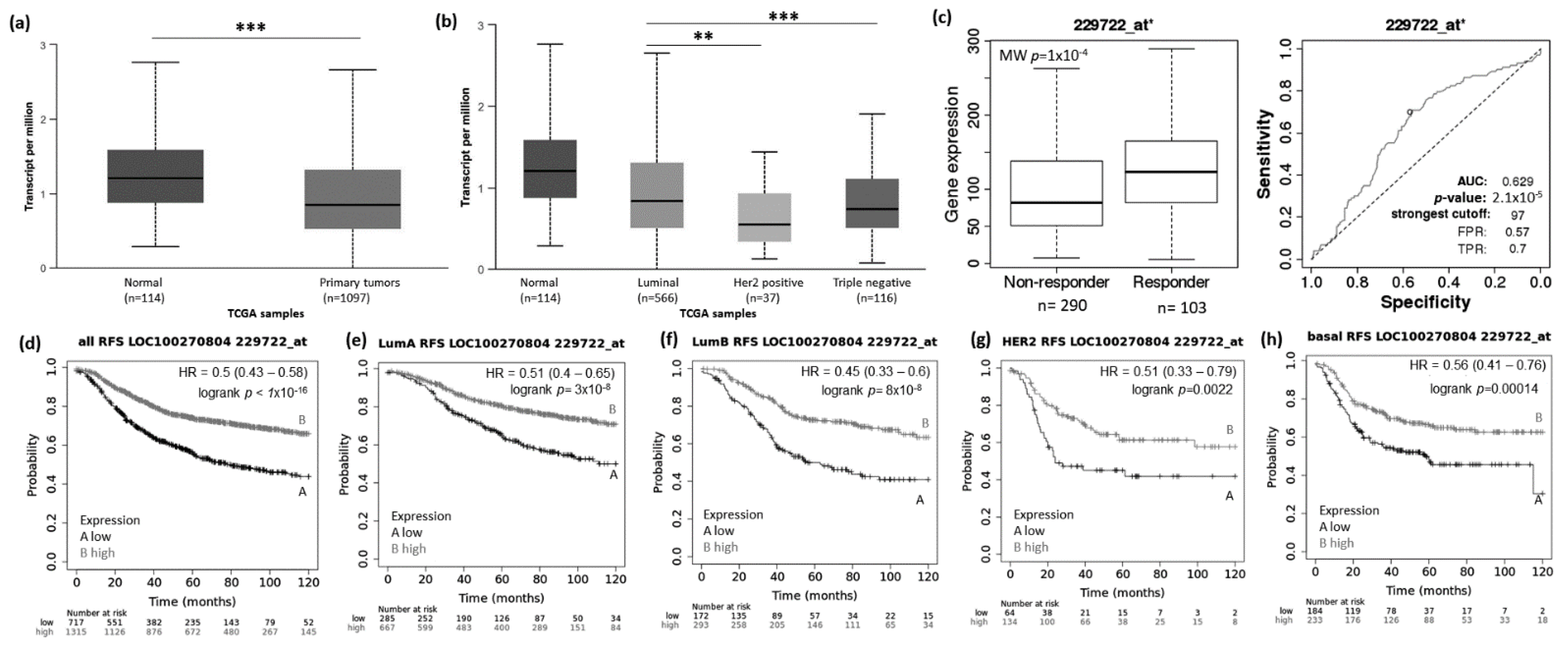
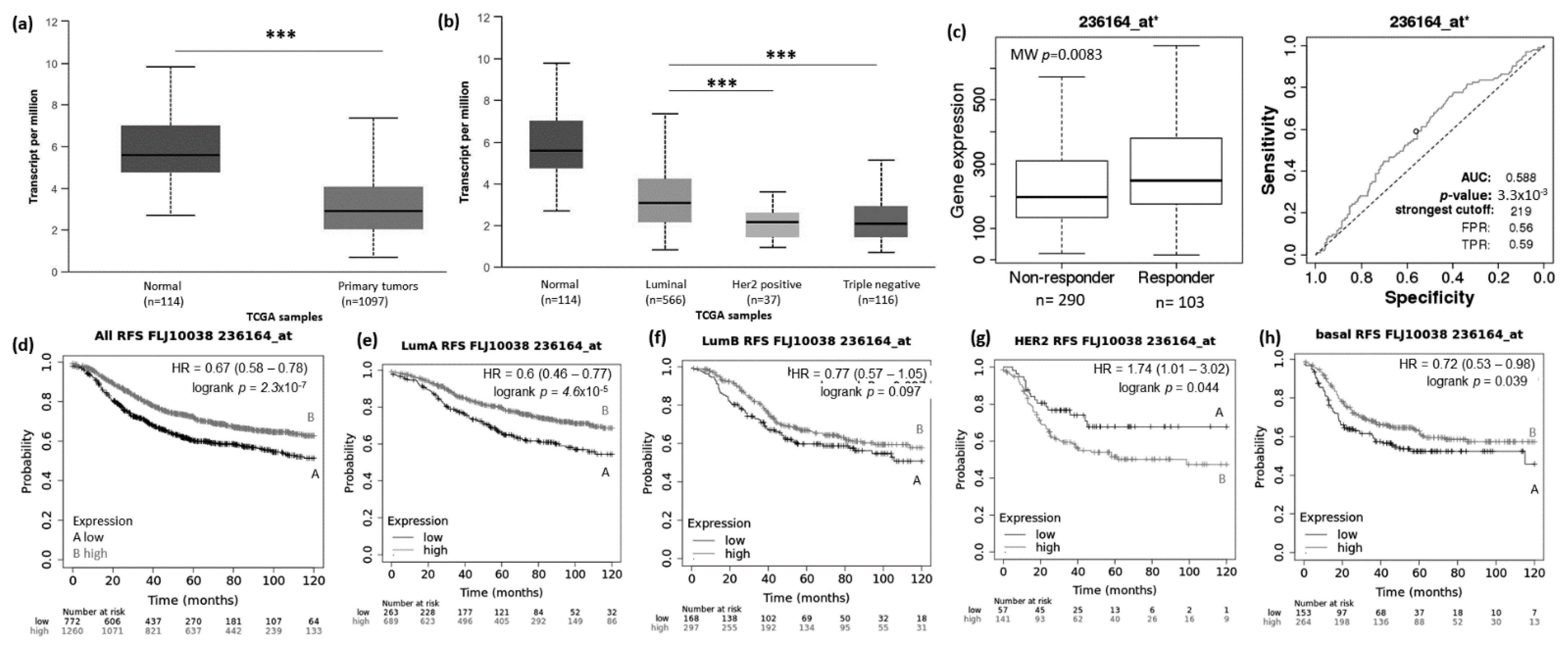
3.1. Abnormal Expression Patterns of Four lncRNAs in BRCA Subtypes
3.2. lncRNAs Expression Patterns Have Impact on Patients Survival
3.3. lncRNAs Expression Patterns Modulate BRCA Response to Taxane Treatment
3.4. Specific lncRNAs Expression Pattern Is Associated with Negative ER and PR BRCA Tumors
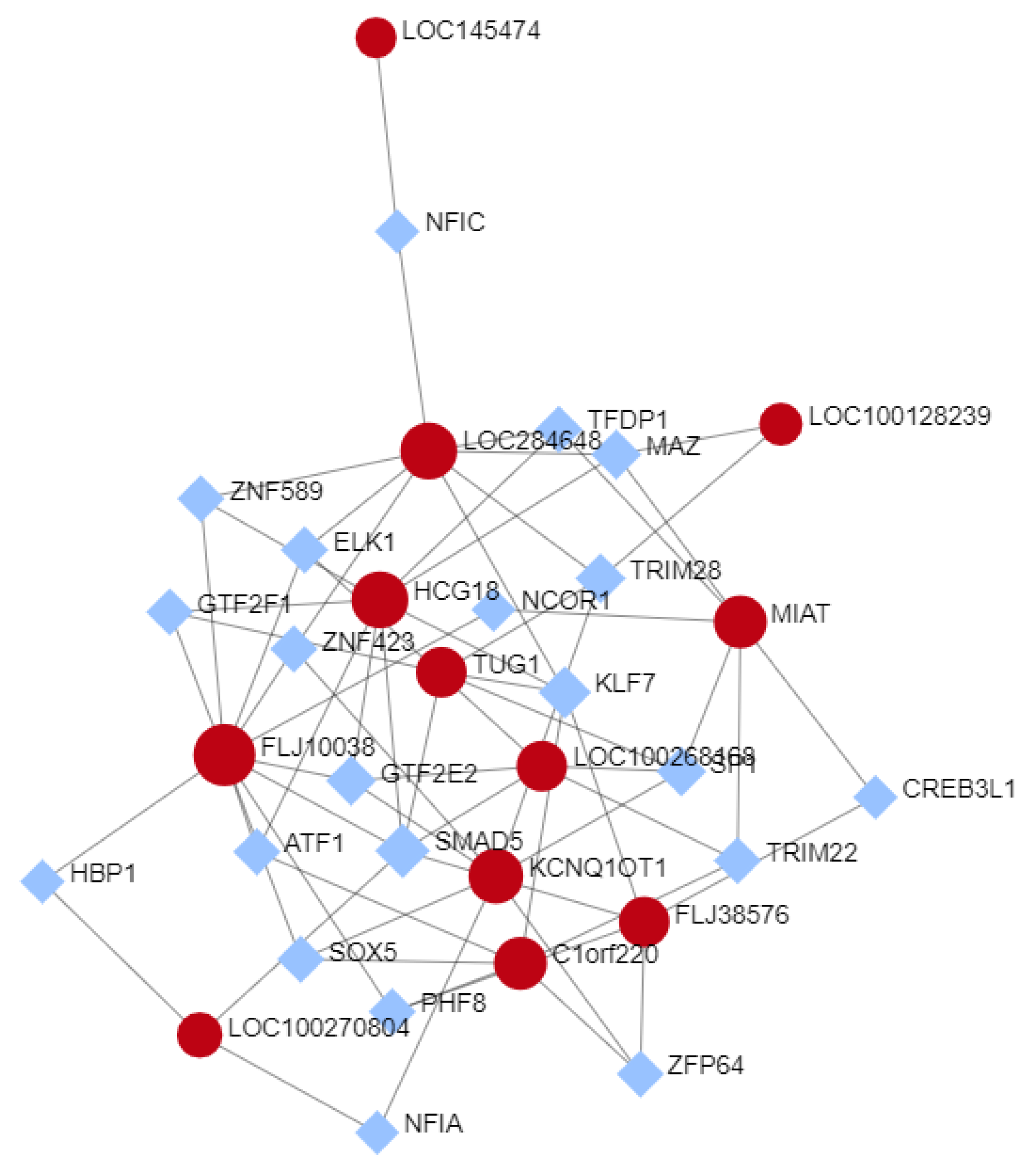
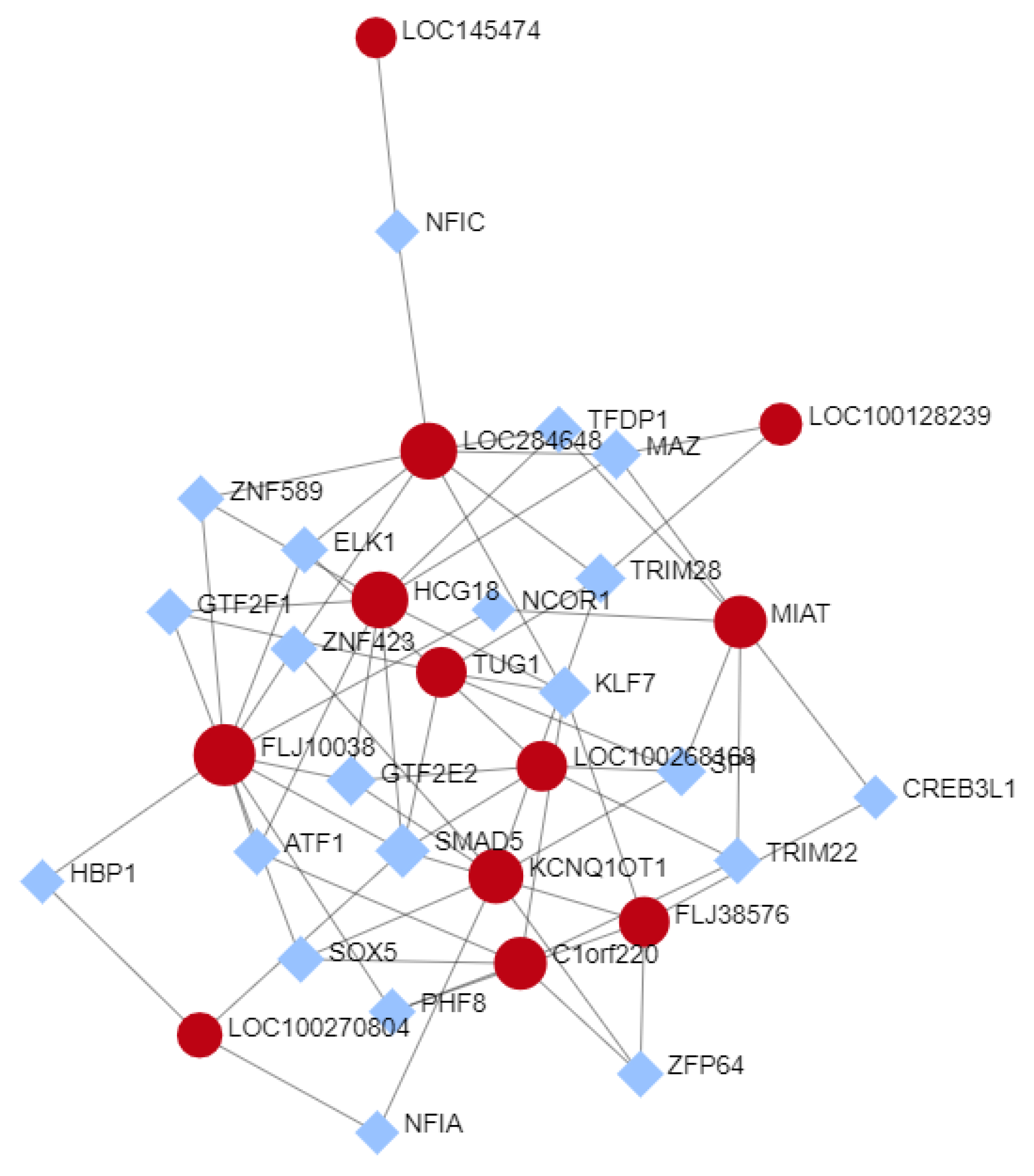
3.5. Selected lncRNAs Expression TF-Interactions Network
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Allemani, C.; Matsuda, T.; Di Carlo, V.; Harewood, R.; Matz, M.; Nikšić, M.; Bonaventure, A.; Valkov, M.; Johnson, C.J.; Estève, J.; et al. Global surveillance of trends in cancer survival 2000-14 (CONCORD-3): Analysis of individual records for 37 513 025 patients diagnosed with one of 18 cancers from 322 population-based registries in 71 countries. Lancet 2018, 391, 1023–1075. [Google Scholar] [CrossRef] [Green Version]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; Van De Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Sørlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef] [Green Version]
- Cech, T.R.; Steitz, J.A. The Noncoding RNA Revolution—Trashing Old Rules to Forge New Ones. Cell 2014, 157, 77–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, G.; Veneziano, D.; Acunzo, M.; Croce, C.M. Small non-coding RNA and cancer. Carcinogenesis 2017, 38, 485–491. [Google Scholar] [CrossRef] [Green Version]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhan, A.; Soleimani, M.; Mandal, S.S. Long Noncoding RNA and Cancer: A New Paradigm. Cancer Res. 2017, 77, 3965–3981. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.; Wang, D.; Wang, J.; Yu, W.; Yang, J. Long Non-Coding RNA in the Pathogenesis of Cancers. Cells 2019, 8, 1015. [Google Scholar] [CrossRef] [Green Version]
- Perry, R.B.; Ulitsky, I. The functions of long noncoding RNAs in development and stem cells. Development 2016, 143, 3882–3894. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Ye, C.; Xiong, H.; Shen, Y.; Lu, Y.; Zhou, J.; Wang, L. Dysregulation of long non-coding RNA in breast cancer: An overview of mechanism and clinical implication. Oncotarget 2017, 8, 5508–5522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barsyte-Lovejoy, D.; Lau, S.K.; Boutros, P.C.; Khosravi, F.; Jurisica, I.; Andrulis, I.L.; Tsao, M.S.; Penn, L.Z. The c-Myc oncogene directly induces the H19 noncoding RNA by allele-specific binding to potentiate tumorigenesis. Cancer Res. 2006, 66, 5330–5337. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zhou, N.; Watabe, K.; Lu, Z.; Wu, F.; Xu, M.; Mo, Y.Y. Long non-coding RNA UCA1 promotes breast tumor growth by suppression of p27 (Kip1). Cell Death Dis. 2014, 5, e1008. [Google Scholar] [CrossRef]
- Hajjari, M.; Salavaty, A. HOTAIR: An oncogenic long non-coding RNA in different cancers. Cancer Biol. Med. 2015, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chi, Y.; Huang, S.; Yuan, L.; Liu, M.; Huang, N.; Zhou, S.; Zhou, B.; Wu, J. Role of BC040587 as a predictor of poor outcome in breast cancer. Cancer Cell Int. 2014, 14, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickard, M.R.; Williams, G.T. The hormone response element mimic sequence of GAS5 lncRNA is sufficient to induce apoptosis in breast cancer cells. Oncotarget 2016, 7, 10104–10116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Wang, P.; You, Z.; Meng, H.; Mu, G.; Bai, X.; Zhang, G.; Zhang, J.; Pang, D. The long non-coding RNA EPB41L4A-AS2 inhibits tumor proliferation and is associated with favorable prognoses in breast cancer and other solid tumors. Oncotarget 2016, 7, 20704–20717. [Google Scholar] [CrossRef] [Green Version]
- Milani, A.; Geuna, E.; Mittica, G.; Valabrega, G. Overcoming endocrine resistance in metastatic breast cancer: Current evidence and future directions. World J. Clin. Oncol. 2014, 5, 990–1001. [Google Scholar] [CrossRef] [Green Version]
- Corrà, F.; Agnoletto, C.; Minotti, L.; Baldassari, F.; Volinia, S. The Network of Non-coding RNAs in Cancer Drug Resistance. Front. Oncol. 2018, 8, 327. [Google Scholar] [CrossRef] [Green Version]
- Si, X.; Zang, R.; Zhang, E.; Liu, Y.; Shi, X.; Shao, L.; Li, A.; Yang, N.; Han, X.; Pan, B.; et al. LncRNA H19 confers chemoresistance in ERα-positive breast cancer through epigenetic silencing of the pro-apoptotic gene BIK. Oncotarget 2016, 7, 81452–81462. [Google Scholar] [CrossRef] [Green Version]
- de Bessa Garcia, S.A.; Pavanelli, A.C.; Cruz E Melo, N.; Nagai, M.A. Prostate apoptosis response 4 (PAR4) expression modulates WNT signaling pathways in MCF7 breast cancer cells: A possible mechanism underlying PAR4-mediated docetaxel chemosensitivity. Int. J. Mol. Med. 2017, 39, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- Nagy, Á.; Lánczky, A.; Menyhárt, O.; Győrffy, B. Validation of miRNA prognostic power in hepatocellular carcinoma using expression data of independent datasets. Sci. Rep. 2018, 8, 9227. [Google Scholar] [CrossRef]
- Fekete, J.T.; Győrffy, B. ROCplot.org: Validating predictive biomarkers of chemotherapy/hormonal therapy/anti-HER2 therapy using transcriptomic data of 3,104 breast cancer patients. Int. J. Cancer 2019, 145, 3140–3151. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Soufan, O.; Ewald, J.; Hancock, R.E.W.; Basu, N.; Xia, J. NetworkAnalyst 3.0: A visual analytics platform for comprehensive gene expression profiling and meta-analysis. Nucleic Acids Res. 2019, 47, W234–W241. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repečka, K.; McDade, F.; Kamath, A.; Banerjee, A.; Luo, Y.; Rogers, D.; Brooks, A.N.; et al. Visualizing and interpreting cancer genomics data via the Xena platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef]
- Moore, J.E.; Purcaro, M.J.; Pratt, H.E.; Epstein, C.B.; Shoresh, N.; Adrian, J.; Kawli, T.; Davis, C.A.; Dobin, A.; Kaul, R.; et al. Expanded encyclopaedias of DNA elements in the human and mouse genomes. Nature 2020, 583, 699–710. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Richard, J.L.C.; Eichhorn, P.J.A. Deciphering the roles of lncRNAs in breast development and disease. Oncotarget 2018, 9, 20179–20212. [Google Scholar] [CrossRef]
- Zhou, S.; He, Y.; Yang, S.; Hu, J.; Zhang, Q.; Chen, W.; Xu, H.; Zhang, H.; Zhong, S.; Zhao, J.; et al. The regulatory roles of lncRNAs in the process of breast cancer invasion and metastasis. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [Green Version]
- Ishii, N.; Ozaki, K.; Sato, H.; Mizuno, H.; Susumu, S.; Takahashi, A.; Miyamoto, Y.; Ikegawa, S.; Kamatani, N.; Hori, M.; et al. Identification of a novel non-coding RNA, MIAT, that confers risk of myocardial infarction. J. Hum. Genet. 2006, 51, 1087–1099. [Google Scholar] [CrossRef] [Green Version]
- Luan, T.; Zhang, X.; Wang, S.; Song, Y.; Zhou, S.; Lin, J.; An, W.; Yuan, W.; Yang, Y.; Cai, H.; et al. Long non-coding RNA MIAT promotes breast cancer progression and functions as ceRNA to regulate DUSP7 expression by sponging miR-155-5p. Oncotarget 2017, 8, 76153–76164. [Google Scholar] [CrossRef]
- Alipoor, F.J.; Asadi, M.H.; Torkzadeh-Mahani, M. MIAT lncRNA is overexpressed in breast cancer and its inhibition triggers senescence and G1 arrest in MCF7 cell line. J. Cell Biochem. 2018, 119, 6470–6481. [Google Scholar] [CrossRef]
- Almnaseer, Z.A.; Mourtada-Maarabouni, M. Long noncoding RNA MIAT regulates apoptosis and the apoptotic response to chemotherapeutic agents in breast cancer cell lines. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Hu, X.; Yu, S.; Deng, S.; Yan, M.; Sun, F.; Song, J.; Tang, L. Silence of lncRNA MIAT-mediated inhibition of DLG3 promoter methylation suppresses breast cancer progression via the Hippo signaling pathway. Cell Signal. 2020, 73, 109697. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, B.; Wu, X.; Huang, Q.; Chen, W.; Zhu, H.; Qu, X.; Xie, L.; Ma, X.; Huang, G. Long non-coding RNA MIAT is estrogen-responsive and promotes estrogen-induced proliferation in ER-positive breast cancer cells. Biochem. Biophys. Res. Commun. 2018, 503, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Safe, S. Transcriptional activation of genes by 17 β-estradiol through estrogen receptor-Sp1 interactions. Vitam. Horm. 2001, 62, 231–252. [Google Scholar] [CrossRef]
- Green, K.A.; Carroll, J.S. Oestrogen-receptor-mediated transcription and the influence of co-factors and chromatin state. Nat. Rev. Cancer 2007, 7, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Bartella, V.; Rizza, P.; Barone, I.; Zito, D.; Giordano, F.; Giordano, C.; Catalano, S.; Mauro, L.; Sisci, D.; Panno, M.L.; et al. Estrogen receptor β binds Sp1 and recruits a corepressor complex to the estrogen receptor α gene promoter. Breast Cancer Res. Treat. 2012, 134, 569–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammad, F.; Pandey, R.R.; Nagano, T.; Chakalova, L.; Mondal, T.; Fraser, P.; Kanduri, C. Kcnq1ot1/Lit1 noncoding RNA mediates transcriptional silencing by targeting to the perinucleolar region. Mol. Cell. Biol. 2008, 28, 3713–3728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Yang, P.; Qiu, X.; Liang, S.; Guan, B.; Yang, H.; Li, F.; Sun, L.; Liu, H.; Zou, G.; et al. KCNQ1OT1 facilitates progression of non-small-cell lung carcinoma via modulating miRNA-27b-3p/HSP90AA1 axis. J. Cell Physiol. 2019, 234, 11304–11314. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Qin, S.; Zayas, J.; Ingle, J.N.; Liu, M.; Weinshilboum, R.M.; Shen, K.; Wang, L. 4-Hydroxytamoxifen enhances sensitivity of estrogen receptor α-positive breast cancer to docetaxel in an estrogen and ZNF423 SNP-dependent fashion. Breast Cancer Res. Treat. 2019, 175, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Wang, C.; Liang, C.; Yang, H.; Chen, D.; Yu, X.; Zhao, W.; Geng, D.; Li, S.; Chen, Z.; et al. The Dysregulated Expression of KCNQ1OT1 and Its Interaction with Downstream Factors miR-145/CCNE2 in Breast Cancer Cells. Cell Physiol. Biochem. 2018, 49, 432–446. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Li, Y.; Ye, Q.; Qin, Y. YY1-mediated long non-coding RNA Kcnq1ot1 promotes the tumor progression by regulating PTEN via DNMT1 in triple negative breast cancer. Cancer Gene Ther. 2020, 1–14. [Google Scholar] [CrossRef]
- Rodriguez, B.A.; Weng, Y.I.; Liu, T.M.; Zuo, T.; Hsu, P.Y.; Lin, C.H.; Cheng, A.L.; Cui, H.; Yan, P.S.; Huang, T.H. Estrogen-mediated epigenetic repression of the imprinted gene cyclin-dependent kinase inhibitor 1C in breast cancer cells. Carcinogenesis 2011, 32, 812–821. [Google Scholar] [CrossRef] [Green Version]
- Ingle, J.N.; Liu, M.; Wickerham, D.L.; Schaid, D.J.; Wang, L.; Mushiroda, T.; Kubo, M.; Costantino, J.P.; Vogel, V.G.; Paik, S.; et al. Selective estrogen receptor modulators and pharmacogenomic variation in ZNF423 regulation of BRCA1 expression: Individualized breast cancer prevention. Cancer Discov. 2013, 3, 812–825. [Google Scholar] [CrossRef] [Green Version]
- Bond, H.M.; Scicchitano, S.; Chiarella, E.; Amodio, N.; Lucchino, V.; Aloisio, A.; Montalcini, Y.; Mesuraca, M.; Morrone, G. ZNF423: A New Player in Estrogen Receptor-Positive Breast Cancer. Front. Endocrinol. 2018, 9, 255. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Xie, G.; Chen, Q.; Xu, Z.; Bai, W.; Chen, M. Identification of a novel lncRNA GABPB1-IT1 that is downregulated and predicts a poor prognosis in non-small cell lung cancer. Oncol. Lett. 2019, 18, 838–845. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Category | MIAT | KCNQ1OT1 | FLJ10038 | LOC100270804 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Low n (%) | High n (%) | p-Value | Low n (%) | High n (%) | p-Value | Low n (%) | High n (%) | p-value | Low n (%) | High n (%) | p-Value | ||
| Age (years) | <50 y | 42 (6.7) | 144 (23.1) | 0.697 | 94 (15.1) | 92 (14.8) | 0.644 | 93 (14.9) | 93 (14.9) | 0.814 | 31 (5.0) | 155 (24.9) | 0.534 |
| ≥50 y | 105 (19.9) | 332 (53.3) | 212 (34.0) | 225 (36.1) | 214 (34.3) | 223 (35.8) | 82 (13.2) | 355 (57.0) | |||||
| Tumor Size | Early | 125 (20.1) | 412 (66.2) | 0.599 | 270 (43.4) | 267 (42.9) | 0.174 | 265 (42.6) | 272 (43.7) | 0.849 | 96 (15.4) | 441 (70.9) | 0.637 |
| late | 22 (3.5) | 63 (10.1) | 36 (5.8) | 49 (7.9) | 41 (6.6) | 44 (7.1) | 17 (2.7) | 68 (10.9) | |||||
| Nodal status | pN | 67 (11.0) | 216 (35.4) | 0.900 | 136 (22.3) | 147 (24.1) | 0.606 | 143 (23.4) | 140 (23.0) | 0.639 | 53 (8.7) | 230 (37.7) | 0.752 |
| pN+ | 76 (12.5) | 251 (41.1) | 164 (26.9) | 163 (26.7) | 159 (26.1) | 168 (27.5) | 58 (9.5) | 269 (44.1) | |||||
| Presence of Metastasis | pM | 133 (23.3) | 424 (74.4) | 1.000 | 267 (46.8) | 290 (50.9) | 0.076 | 278 (48.8) | 279 (48.9) | 0.779 | 101 (17.7) | 456 (80.0) | 0.332 |
| pM+ | 3 (0.5) | 10 (1.8) | 3 (0.5) | 10 (1.8) | 7 (1.2) | 6 (1.1) | 1 (0.2) | 12 (2.1) | |||||
| Tumor Stage | Early | 108 (17.8) | 361 (59.6) | 0.541 | 224 (37.5) | 245 (40.4) | 0.323 | 225 (37.1) | 244 (40.3) | 0.274 | 84 (13.9) | 385 (63.5) | 0.632 |
| late | 35 (5.8) | 102 (16.8) | 72 (11.9) | 65 (10.7) | 73 (12.0) | 64 (10.6) | 27 (4.5) | 110 (18.2) | |||||
| ER status | negative | 41 (6.9) | 99 (16.7) | 0.093 | 67 (11.3) | 73 (12.3) | 0.691 | 91 (15.4) | 49 (8.3) | <0.0001 | 32 (5.4) | 108 (18.2) | 0.154 |
| positive | 101 (17.1) | 351 (59.3) | 225 (38.0) | 227 (38.3) | 199 (33.6) | 253 (42.7) | 79 (13.3) | 373 (63.0) | |||||
| PR status | negative | 56 (9.5) | 144 (24.4) | 0.081 | 90 (15.3) | 110 (18.6) | 0.118 | 119 (20.2) | 81 (13.7) | <0.0001 | 43 (7.3) | 157 (26.6) | 0.232 |
| positive | 84 (14.2) | 306 (51.9) | 202 (34.2) | 188 (31.9) | 170 (28.8) | 220 (37.3) | 68 (11.5) | 322 (54.6) | |||||
| Menopausal Status | pre | 28 (5.3) | 105 (19.7) | 0.486 | 67 (12.6) | 66 (12.4) | 0.708 | 62 (11.6) | 71 (13.3) | 0.706 | 23 (4.3) | 110 (20.6) | 0.905 |
| post | 96 (18.0) | 304 (57.0) | 194 (36.4) | 206 (28.6) | 194 (36.4) | 206 (38.6) | 71 (13.3) | 329 (61.7) | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavanelli, A.C.; Mangone, F.R.; Barros, L.R.C.; Machado-Rugolo, J.; Capelozzi, V.L.; Nagai, M.A. Abnormal Long Non-Coding RNAs Expression Patterns Have the Potential Ability for Predicting Survival and Treatment Response in Breast Cancer. Genes 2021, 12, 996. https://doi.org/10.3390/genes12070996
Pavanelli AC, Mangone FR, Barros LRC, Machado-Rugolo J, Capelozzi VL, Nagai MA. Abnormal Long Non-Coding RNAs Expression Patterns Have the Potential Ability for Predicting Survival and Treatment Response in Breast Cancer. Genes. 2021; 12(7):996. https://doi.org/10.3390/genes12070996
Chicago/Turabian StylePavanelli, Ana Carolina, Flavia Rotea Mangone, Luciana R. C. Barros, Juliana Machado-Rugolo, Vera L. Capelozzi, and Maria A. Nagai. 2021. "Abnormal Long Non-Coding RNAs Expression Patterns Have the Potential Ability for Predicting Survival and Treatment Response in Breast Cancer" Genes 12, no. 7: 996. https://doi.org/10.3390/genes12070996
APA StylePavanelli, A. C., Mangone, F. R., Barros, L. R. C., Machado-Rugolo, J., Capelozzi, V. L., & Nagai, M. A. (2021). Abnormal Long Non-Coding RNAs Expression Patterns Have the Potential Ability for Predicting Survival and Treatment Response in Breast Cancer. Genes, 12(7), 996. https://doi.org/10.3390/genes12070996