
Endopolyploidy Variation in Wild Barley Seeds across Environmental Gradients in Israel

 , , , , , and
, , , , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Analysis of Nuclear DNA Content and Calculation of the Super Cycle Value (SCV)
2.3. Determination of Seed Morphology Parameters
2.4. Cell Death Assay by Evans Blue Staining
2.5. Statistical Analysis
3. Results
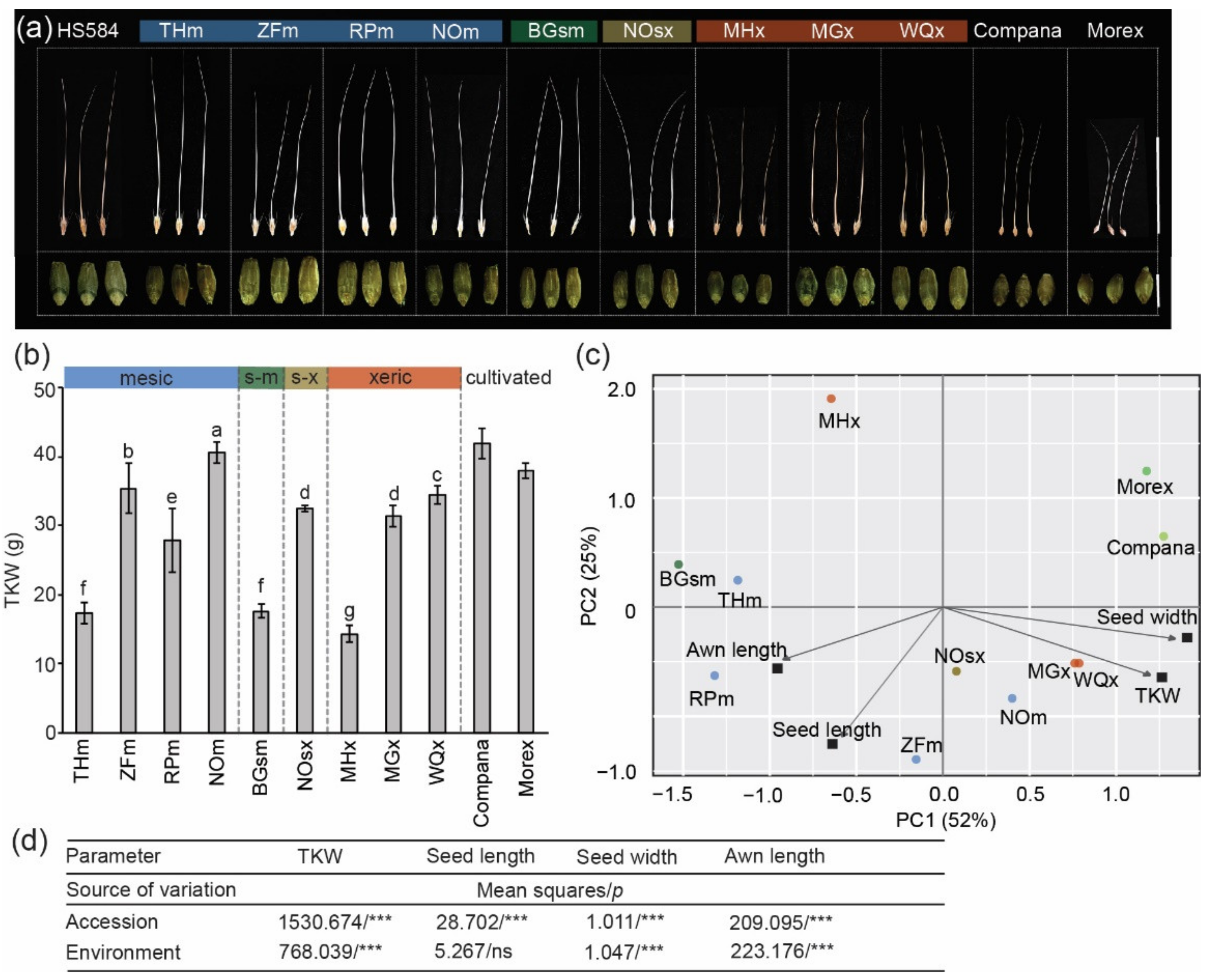
3.1. Variation in Mature Dry Seed Morphology of Wild Barley
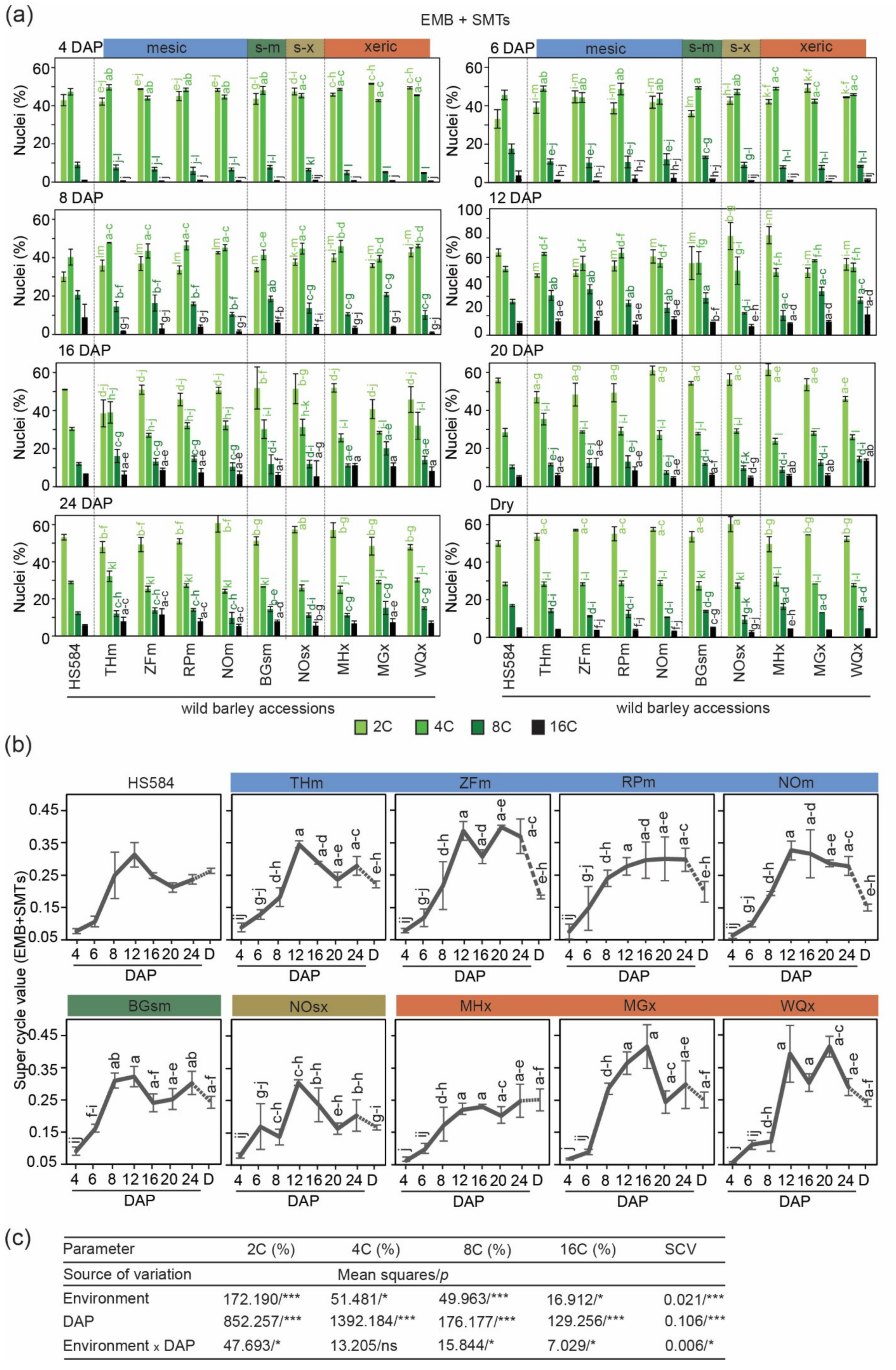
3.2. Variation in Endoreduplication Dynamics in Developing Wild Barley Seeds
3.2.1. Diploid Seed Tissues
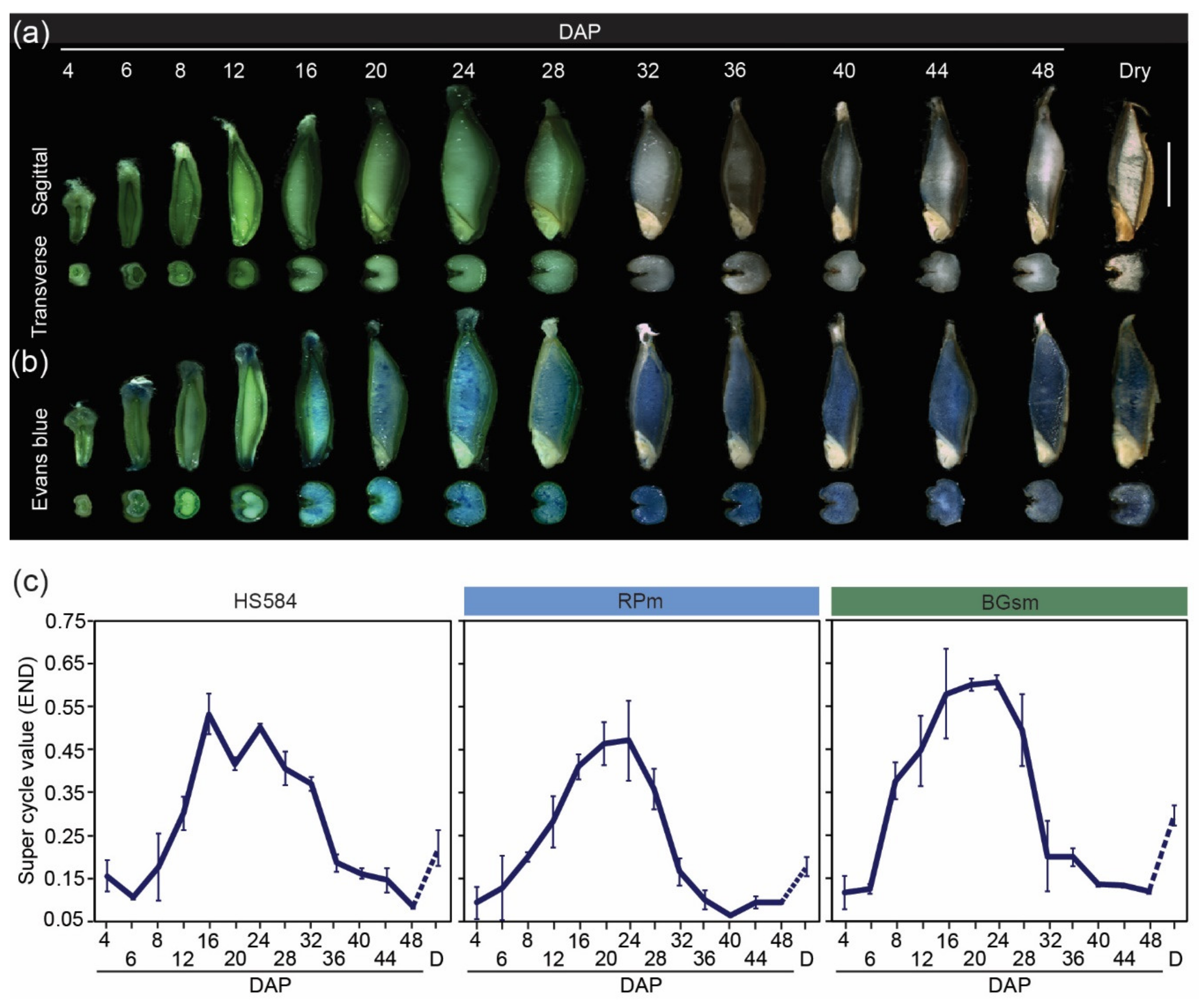
3.2.2. Triploid Endosperm Tissues
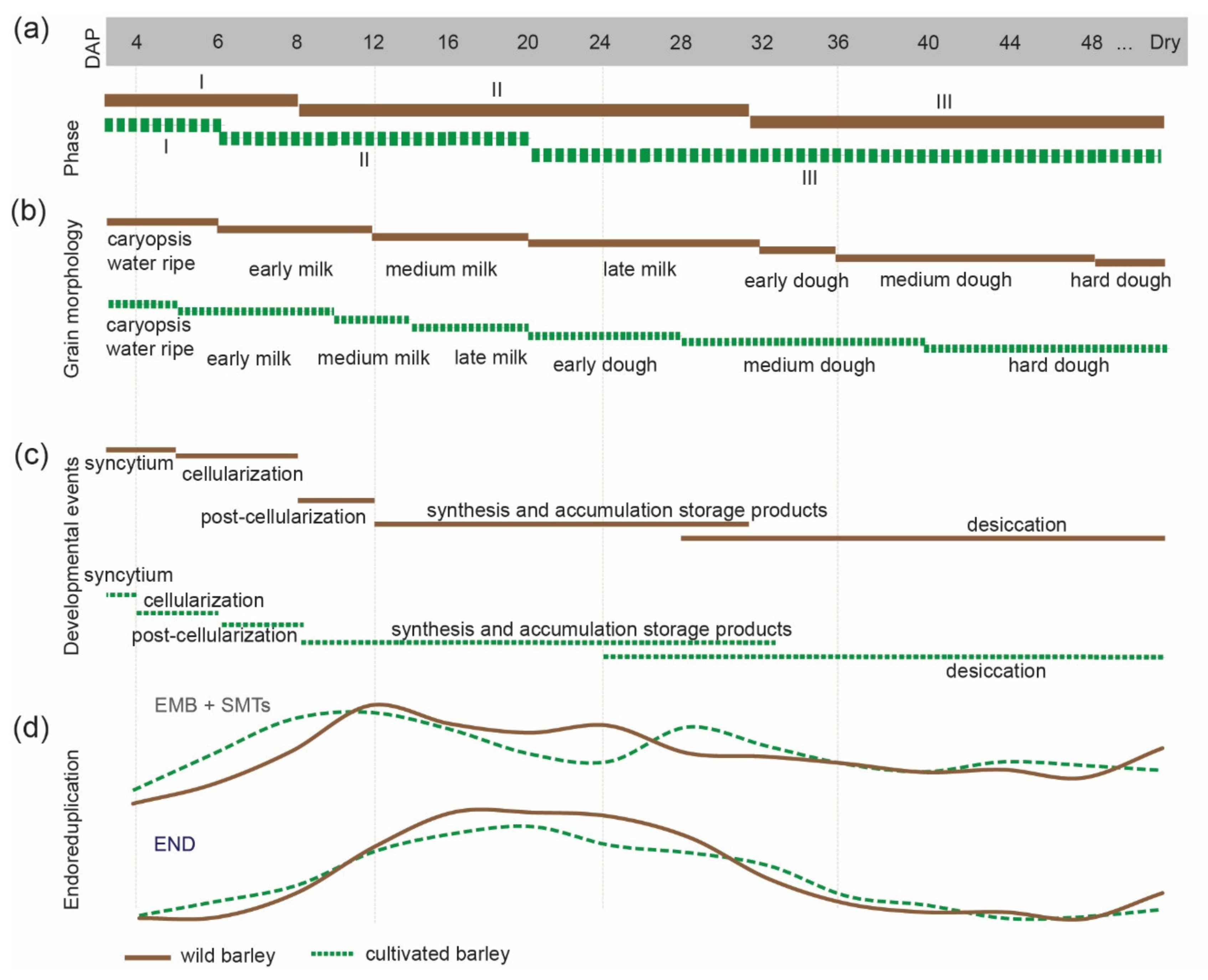
3.3. Comparison of Endoreduplication Dynamics in Developing Seeds of Wild and Cultivated Barley
3.4. Morphological and Cellular Changes during 48 Days of Wild Barley Seed Development
4. Discussion
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Badr, A.; Müller, K.; Schäfer-Pregl, R.; El Rabey, H.; Effgen, S.; Ibrahim, H.H.; Pozzi, C.; Rohde, W.; Salamini, F. On the origin and domestication history of barley (Hordeum vulgare). Mol. Biol. Evol. 2000, 17, 499–510. [Google Scholar] [CrossRef]
- Mascher, M.; Schuenemann, V.J.; Davidovich, U.; Marom, N.; Himmelbach, A.; Hübner, S.; Korol, A.; David, M.; Reiter, E.; Riehl, S.; et al. Genomic analysis of 6,000-year-old cultivated grain illuminates the domestication history of barley. Nat. Genet. 2016, 48, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Jakob, S.S.; Rödder, D.; Engler, J.O.; Shaaf, S.; Özkan, H.; Blattner, F.R.; Kilian, B. Evolutionary history of wild barley (Hordeum vulgare subsp. spontaneum) analyzed using multilocus sequence data and paleodistribution modeling. Genome Biol. Evol. 2014, 6, 685–702. [Google Scholar] [CrossRef] [PubMed]
- Krugman, T.; Nevo, E.; Beharav, A.; Sela, H.; Fahima, T. The Institute of Evolution Wild Cereal Gene Bank at the University of Haifa. Isr. J. Plant. Sci. 2019, 65, 129–146. [Google Scholar] [CrossRef]
- Von Korff, M.; Wang, H.; Léon, J.; Pillen, K. Development of candidate introgression lines using an exotic barley accession (Hordeum vulgare ssp. spontaneum) as donor. Theor. Appl. Genet. 2004, 109, 1736–1745. [Google Scholar] [CrossRef]
- Nevo, E. Origin, evolution, population genetics and resources for breeding of wild barley, Hordeum spontaneum, in the Fertile Crescent. In Barley: Genetics, Biochemistry, Molecular Biology and Biotechnology; Shewry, P., Ed.; Centre for Agriculture and Bioscience International: Wallingford, UK, 1992; pp. 19–43. ISBN 0-85198-725-7. [Google Scholar]
- Fetch, T.G.; Steffenson, B.J.; Nevo, E. Diversity and sources of multiple disease resistance in Hordeum spontaneum. Plant. Dis. 2003, 87, 1439–1448. [Google Scholar] [CrossRef]
- Grossi, M.; Giorni, E.; Rizza, F.; Stanca, A.M.; Cattivelli, L. Wild and cultivated barleys show differences in the expression pattern of a cold-regulated gene family under different light and temperature conditions. Plant. Mol. Biol. 1998, 38, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.; Chen, X.; Han, Z.; Wu, X.; Zhang, S.; Li, Q.; Nazir, M.M.; Zhang, G.; Zeng, F. Screening of worldwide barley collection for drought tolerance: The assessment of various physiological measures as the selection criteria. Front. Plant. Sci. 2020, 11, 1159. [Google Scholar] [CrossRef]
- Nevo, E.; Krugman, T.; Beiles, A. Genetic resources for salt tolerance in the wild progenitors of wheat (Triticum dicoccoides) and barley (Hordeum spontaneum) in Israel. Plant. Breed. 1993, 110, 338–341. [Google Scholar] [CrossRef]
- Nowicka, A.; Kovacik, M.; Tokarz, B.; Vrána, J.; Zhang, Y.; Weigt, D.; Doležel, J.; Pecinka, A. Dynamics of endoreduplication in developing barley seeds. J. Exp. Bot. 2021, 72, 268–282. [Google Scholar] [CrossRef]
- Tottman, D.R.; Makepeace, R.J.; Broad, H. An explanation of the decimal code for the growth stages of cereals, with illustrations. Ann. Appl. Biol. 1979, 93, 221–234. [Google Scholar] [CrossRef]
- Evers, T.; Millar, S. Cereal grain structure and development: Some implications for quality. J. Cereal Sci. 2002, 36, 261–284. [Google Scholar] [CrossRef]
- Radchuk, V.; Borisjuk, L. Physical, metabolic and developmental functions of the seed coat. Front. Plant. Sci. 2014, 5, 510. [Google Scholar] [CrossRef]
- Evers, A.D.; Blakeney, A.; O’Brien, L. Cereal structure and composition. Aust. J. Agric. Res. 1999, 50, 629–650. [Google Scholar] [CrossRef]
- Olsen, O.-A. Endosperm development: Cellularization and cell fate specification. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 2001, 52, 233–267. [Google Scholar] [CrossRef]
- Rodríguez, M.V.; Barrero, J.M.; Corbineau, F.; Gubler, F.; Benech-Arnold, R.L. Dormancy in cereals (not too much, not so little): About the mechanisms behind this trait. Seed Sci. Res. 2015, 25, 99–119. [Google Scholar] [CrossRef]
- D’amato, F. Endopolyploidy as a factor in plant tissue development. Caryologia 1964, 17, 41–52. [Google Scholar] [CrossRef][Green Version]
- Nagl, W. DNA endoreduplication and polyteny understood as evolutionary strategies. Nature 1976, 261, 614–615. [Google Scholar] [CrossRef]
- Sabelli, P.A.; Larkins, B.A. The development of endosperm in grasses. Plant. Physiol. 2009, 149, 14–26. [Google Scholar] [CrossRef]
- De Veylder, L.; Larkin, J.C.; Schnittger, A. Molecular control and function of endoreplication in development and physiology. Trends Plant. Sci. 2011, 16, 624–634. [Google Scholar] [CrossRef]
- Scholes, D.R.; Paige, K.N. Plasticity in ploidy: A generalized response to stress. Trends Plant. Sci. 2015, 20, 165–175. [Google Scholar] [CrossRef]
- Gendreau, E.; Traas, J.; Desnos, T.; Grandjean, O.; Caboche, M.; Hofte, H. Cellular basis of hypocotyl growth in Arabidopsis thaliana. Plant. Physiol. 1997, 114, 295–305. [Google Scholar] [CrossRef]
- Bhosale, R.; Boudolf, V.; Cuevas, F.; Lu, R.; Eekhout, T.; Hu, Z.; Van Isterdael, G.; Lambert, G.M.; Xu, F.; Nowack, M.K.; et al. A spatiotemporal dna endoploidy map of the Arabidopsis root reveals roles for the endocycle in root development and stress adaptation. Plant. Cell 2018, 30, 2330–2351. [Google Scholar] [CrossRef]
- Lingua, G.; Fusconi, A.; Berta, G. The nucleus of differentiated root plant cells: Modifications induced by arbuscular mycorrhizal fungi. Eur. J. Histochem. 2001, 45, 9–20. [Google Scholar] [CrossRef]
- Chandran, D.; Inada, N.; Hather, G.; Kleindt, C.K.; Wildermuth, M.C. Laser microdissection of Arabidopsis cells at the powdery mildew infection site reveals site-specific processes and regulators. Proc. Natl. Acad. Sci. USA 2010, 107, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Cookson, S.J.; Radziejwoski, A.; Granier, C. Cell and leaf size plasticity in Arabidopsis: What is the role of endoreduplication? Plant. Cell Environ. 2006, 29, 1273–1283. [Google Scholar] [CrossRef] [PubMed]
- Tran, V.; Weier, D.; Radchuk, R.; Thiel, J.; Radchuk, V. Caspase-like activities accompany programmed cell death events in developing barley grains. PLoS ONE 2014, 9, e109426. [Google Scholar] [CrossRef] [PubMed]
- Radchuk, V.; Tran, V.; Radchuk, R.; Diaz-Mendoza, M.; Weier, D.; Fuchs, J.; Riewe, D.; Hensel, G.; Kumlehn, J.; Munz, E.; et al. Vacuolar processing enzyme 4 contributes to maternal control of grain size in barley by executing programmed cell death in the pericarp. New Phytol. 2018, 218, 1127–1142. [Google Scholar] [CrossRef] [PubMed]
- Young, T.E.; Gallie, D.R. Programmed cell death during endosperm development. Plant. Mol. Biol. 2000, 44, 283–301. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, J.; Li, D.; Liu, C.M. Rice caryopsis development II: Dynamic changes in the endosperm. J. Integr. Plant. Biol. 2016, 58, 786–798. [Google Scholar] [CrossRef]
- Nevo, E. Evolution of wild barley at “Evolution Canyon”: Adaptation, speciation, pre-agricultural collection, and barley improvement. Isr. J. Plant. Sci. 2015, 62, 22–32. [Google Scholar] [CrossRef]
- Waddington, S.R.; Cartwright, P.M.; Wall, P.C. A quantitative scale of spike initial and pistil development in barley and wheat. Ann. Bot. 1983, 51, 119–130. [Google Scholar] [CrossRef]
- Kovacik, M.; Nowicka, A.; Pecinka, A. Isolation of high purity tissues from developing barley seeds. J. Vis. Exp. 2020, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Nevo, E.; Zohary, D.; Brown, A.H.D.; Haber, M. Genetic diversity and environmental associations of wild barley, Hordeum spontaneum, in Israel. Evolution 1979, 33, 815–833. [Google Scholar] [CrossRef]
- Rewers, M.; Sliwinska, E. Endoreduplication intensity as a marker of seed developmental stage in the Fabaceae. Cytom. Part A 2012, 81A, 1067–1075. [Google Scholar] [CrossRef]
- Nevo, E.; Beiles, A.; Kaplan, D.; Storch, N.; Zohary, D. Genetic diversity and environmental associations of wild barley, Hordeum spontaneum (Poaceae), in Iran. Plant. Syst. Evol. 1986, 153, 141–164. [Google Scholar] [CrossRef]
- Hosseini, M.; Ghorbani, R.; Rashed Mohassel, M.H.; Yassaie, M. Correlation of environmental factors and phenotypic diversity of iranian wild barley (Hordeum Spontaneum Koch) populations. Acta Oecologica 2019, 95, 19–25. [Google Scholar] [CrossRef]
- Shakhatreh, Y.; Haddad, N.; Alrababah, M.; Grando, S.; Ceccarelli, S. Phenotypic diversity in wild barley (Hordeum vulgare L. ssp. spontaneum (C. Koch) Thell.) accessions collected in Jordan. Genet. Resour. Crop. Evol. 2010, 57, 131–146. [Google Scholar] [CrossRef]
- Volis, S.; Mendlinger, S.; Ward, D. Adaptive traits of wild barley plants of Mediterranean and desert origin. Oecologia 2002, 133, 131–138. [Google Scholar] [CrossRef]
- Dante, R.A.; Larkins, B.A.; Sabelli, P.A. Cell cycle control and seed development. Front. Plant. Sci. 2014, 5, 493. [Google Scholar] [CrossRef]
- Sreenivasulu, N.; Borisjuk, L.; Junker, B.H.; Mock, H.-P.; Rolletschek, H.; Seiffert, U.; Weschke, W.; Wobus, U. Barley grain development: Toward an integrative view. In International Review of Cell and Molecular Biology; Jeon, K., Ed.; Elsevier: Amsterdam, The Netherlands, 2010; Volume 281, pp. 49–89. [Google Scholar]
- Rewers, M.; Sadowski, J.; Sliwinska, E. Endoreduplication in cucumber (Cucumis sativus) seeds during development, after processing and storage, and during germination. Ann. Appl. Biol. 2009, 155, 431–438. [Google Scholar] [CrossRef]
- Barow, M.; Meister, A. Endopolyploidy in seed plants is differently correlated to systematics, organ, life strategy and genome size. Plant. Cell Environ. 2003, 26, 571–584. [Google Scholar] [CrossRef]
- Sliwinska, E.; Lukaszewska, E. Polysomaty in growing in vitro sugar-beet (Beta vulgaris L.) seedlings of different ploidy level. Plant. Sci. 2005, 168, 1067–1074. [Google Scholar] [CrossRef]
- Chojecki, A.; Gale, M.; Bayliss, M. The number and sizes of starch granules in the wheat endosperm, and their association with grain weight. Ann. Bot. 1986, 58, 819–831. [Google Scholar] [CrossRef]
- Kobayashi, H. Variations of endoreduplication and its potential contribution to endosperm development in rice (Oryza sativa L.). Plant. Prod. Sci. 2019, 22, 227–241. [Google Scholar] [CrossRef]
- Kladnik, A.; Chourey, P.S.; Pring, D.R.; Dermastia, M. Development of the endosperm of Sorghum bicolor during the endoreduplication-associated growth phase. J. Cereal Sci. 2006, 43, 209–215. [Google Scholar] [CrossRef]
- Sabelli, P.A.; Larkins, B.A. Regulation and function of retinoblastoma-related plant genes. Plant. Sci. 2009, 177, 540–548. [Google Scholar] [CrossRef]
- Keown, A.C.; Taiz, L.; Jones, R.L. The nuclear DNA content of developing barley aleurone cells. Am. J. Bot. 1977, 64, 1248–1253. [Google Scholar] [CrossRef]
- Sreenivasulu, N.; Usadel, B.; Winter, A.; Radchuk, V.; Scholz, U.; Stein, N.; Weschke, W.; Strickert, M.; Close, T.J.; Stitt, M.; et al. Barley grain maturation and germination: Metabolic pathway and regulatory network commonalities and differences highlighted by new MapMan/PageMan profiling tools. Plant. Physiol. 2008, 146, 1738–1758. [Google Scholar] [CrossRef]
- Burešová, V.; Kopecký, D.; Bartoš, J.; Martinek, P.; Watanabe, N.; Vyhnánek, T.; Doležel, J. Variation in genome composition of blue-aleurone wheat. Theor. Appl. Genet. 2015, 128, 273–282. [Google Scholar] [CrossRef]
- Jeewani, D.C.; Hua, W.Z. Grain color development and segregation of blue wheat. Res. J. Biotechnol. 2017, 12, 40–45. [Google Scholar]
- Chevalier, C.; Nafati, M.; Mathieu-Rivet, E.; Bourdon, M.; Frangne, N.; Cheniclet, C.; Renaudin, J.P.; Gvaudant, F.; Hernould, M. Elucidating the functional role of endoreduplication in tomato fruit development. Ann. Bot. 2011, 107, 1159–1169. [Google Scholar] [CrossRef] [PubMed]
- Sterken, R.; Kiekens, R.; Boruc, J.; Zhang, F.; Vercauteren, A.; Vercauteren, I.; De Smet, L.; Dhondt, S.; Inzé, D.; De Veylder, L.; et al. Combined linkage and association mapping reveals CYCD5;1 as a quantitative trait gene for endoreduplication in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 4678–4683. [Google Scholar] [CrossRef]
- Hübner, S.; Höffken, M.; Oren, E.; Haseneyer, G.; Stein, N.; Graner, A.; Schmid, K.; Fridman, E. Strong correlation of wild barley (Hordeum spontaneum) population structure with temperature and precipitation variation. Mol. Ecol. 2009, 18, 1523–1536. [Google Scholar] [CrossRef]
- Barow, M. Endopolyploidy in seed plants. BioEssays 2006, 28, 271–281. [Google Scholar] [CrossRef]
- Siddappaji, M.H.; Scholes, D.R.; Bohn, M.; Paige, K.N. Overcompensation in response to herbivory in Arabidopsis thaliana: The role of glucose-6-phosphate dehydrogenase and the oxidative pentose-phosphate pathway. Genetics 2013, 195, 589–598. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality/Name (All within Israel) | Genebank | Acronym | Type of Environment | Longitude (N) | Latitude (E) | Altitude (a.s.l.) | Maximum Temperature in April (°C) | Rainfall in April (mm) | Annual Rainfall (mm) | Average Annual Humidity at 14:00 Mean ± SD | Annual Evaporation (cm) | Soil Type |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bar Giyyora | ICGB | BGsm | Semi-mesic | 35.083333 | 31.716667 | 760 | 22 | 18 | 535 | 47.1 ± 10.8 | 215 | T |
| HS584 | IPK | HS584 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. |
| Machtesh Gadol | ICGB | MGx | Xeric | 35.000000 | 30.950000 | n.a. | 25 | 3 | 70 | n.a. | n.a. | n.a. |
| Mehola | ICGB | MHx | Xeric | 35.533333 | 32.350000 | −150 | 30 | 6 | <200 | 37.5 | 240 | A |
| Nahal Oren | ICGB | NOm | Mesic | 34.966667 | 32.716667 | n. a. | 24 | 13 | 584 | n.a. | n.a. | n.a. |
| Nahal Oren | ICGB | NOsx | Semi- xeric | 34.966667 | 32.716667 | n. a. | 24 | 13 | 584 | n.a. | n.a. | n.a. |
| Rosh Pinna | ICGB | RPm | Mesic | 35.550000 | 32.983333 | 700 | 25 | 20 | 535 | 43.6 ± 10.5 | 220 | T |
| Tel Hai | ICGB | THm | Mesic | 35.573979 | 33.234719 | 400 | 26 | 23 | 768 | 46.9 ± 7.6 | 220 | T |
| Wadi Qilt | ICGB | WQx | Xeric | 35.44565 | 31.859 | 50 | 30 | 6 | <200 | 34.7 ± 9.3 | 330 | A |
| Zefat | ICGB | ZFm | Mesic | 35.496001 | 32.969206 | 800 | 20 | 27 | 670 | 50.4 ± 13.1 | 220 | R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowicka, A.; Sahu, P.P.; Kovacik, M.; Weigt, D.; Tokarz, B.; Krugman, T.; Pecinka, A. Endopolyploidy Variation in Wild Barley Seeds across Environmental Gradients in Israel. Genes 2021, 12, 711. https://doi.org/10.3390/genes12050711
Nowicka A, Sahu PP, Kovacik M, Weigt D, Tokarz B, Krugman T, Pecinka A. Endopolyploidy Variation in Wild Barley Seeds across Environmental Gradients in Israel. Genes. 2021; 12(5):711. https://doi.org/10.3390/genes12050711
Chicago/Turabian StyleNowicka, Anna, Pranav Pankaj Sahu, Martin Kovacik, Dorota Weigt, Barbara Tokarz, Tamar Krugman, and Ales Pecinka. 2021. "Endopolyploidy Variation in Wild Barley Seeds across Environmental Gradients in Israel" Genes 12, no. 5: 711. https://doi.org/10.3390/genes12050711
APA StyleNowicka, A., Sahu, P. P., Kovacik, M., Weigt, D., Tokarz, B., Krugman, T., & Pecinka, A. (2021). Endopolyploidy Variation in Wild Barley Seeds across Environmental Gradients in Israel. Genes, 12(5), 711. https://doi.org/10.3390/genes12050711