
TERT Promoter Mutation Analysis of Whole-Organ Mapping Bladder Cancers


 ,
, 
 ,
, 


Abstract
1. Introduction
2. Materials and Methods
2.1. Whole-Organ Mapping Bladder Tumor Specimens and Strategy
2.2. Immunohistochemical Analysis of Tumor Associated Normal Urothelium and Non-Invasive Urothelial Lesions from Whole-Organ Mapping Bladder Cancer Specimens
2.3. DNA Isolation
2.4. TERT Promoter Mutation Analysis
2.5. Telomere Length Determination
2.6. Statistical Analysis
3. Results
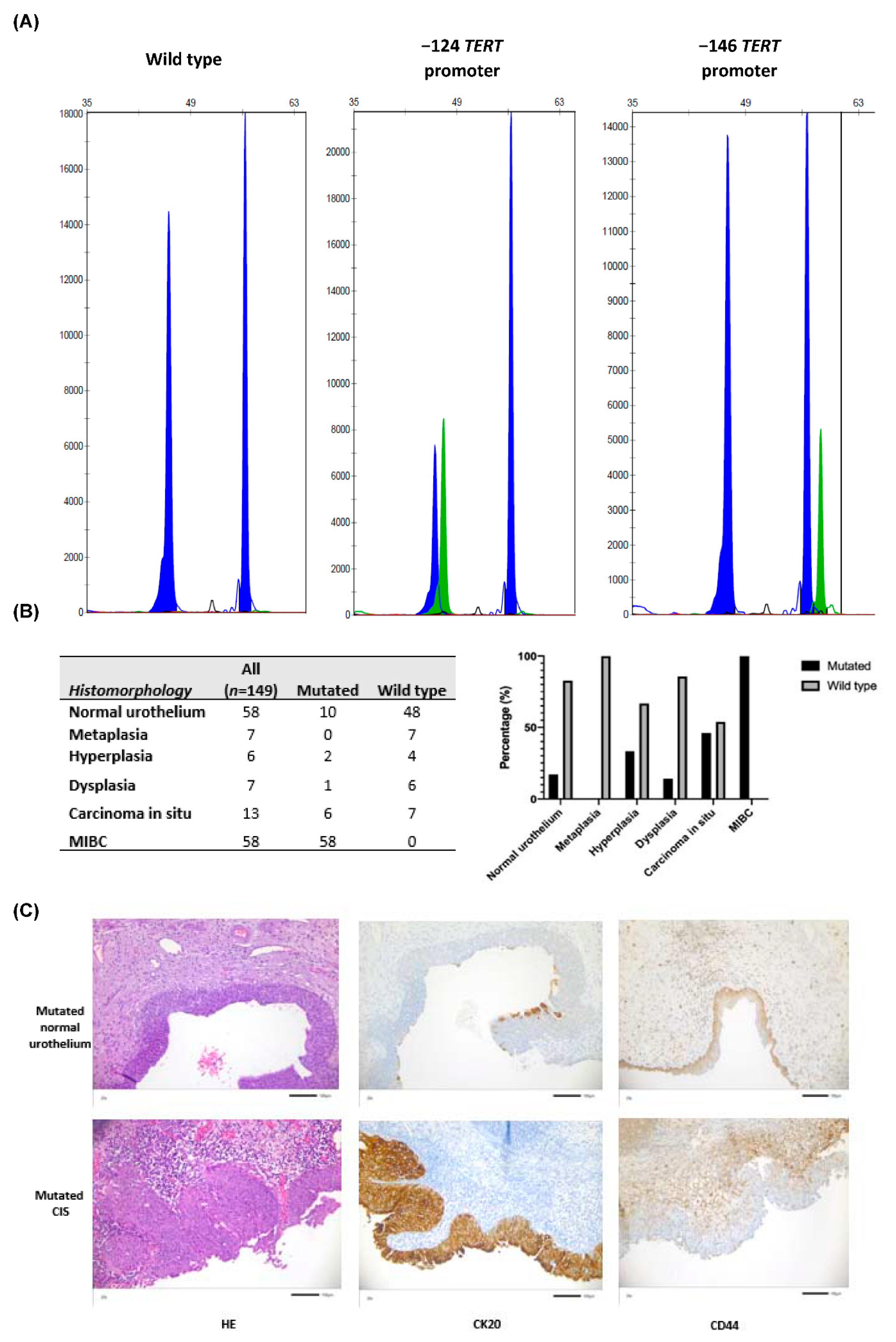
3.1. TERT Promoter Mutations Were Identified within Tumor Associated Normal Urothelium, Non-Invasive Urothelial Lesions, CIS and MIBC from Whole-Organ Mapping Bladder Cancer Specimens
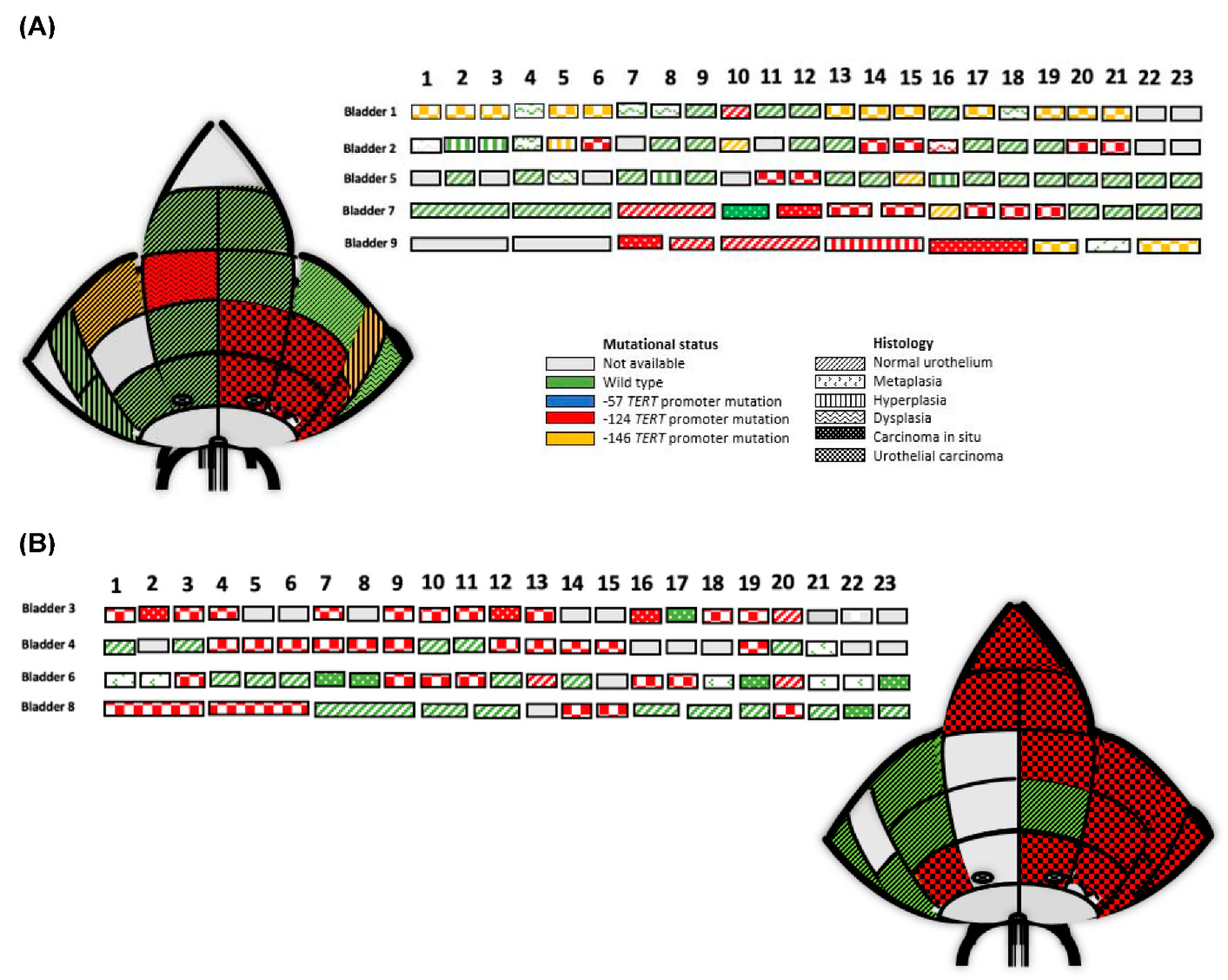
3.2. Clonality and TERT Promoter Mutations
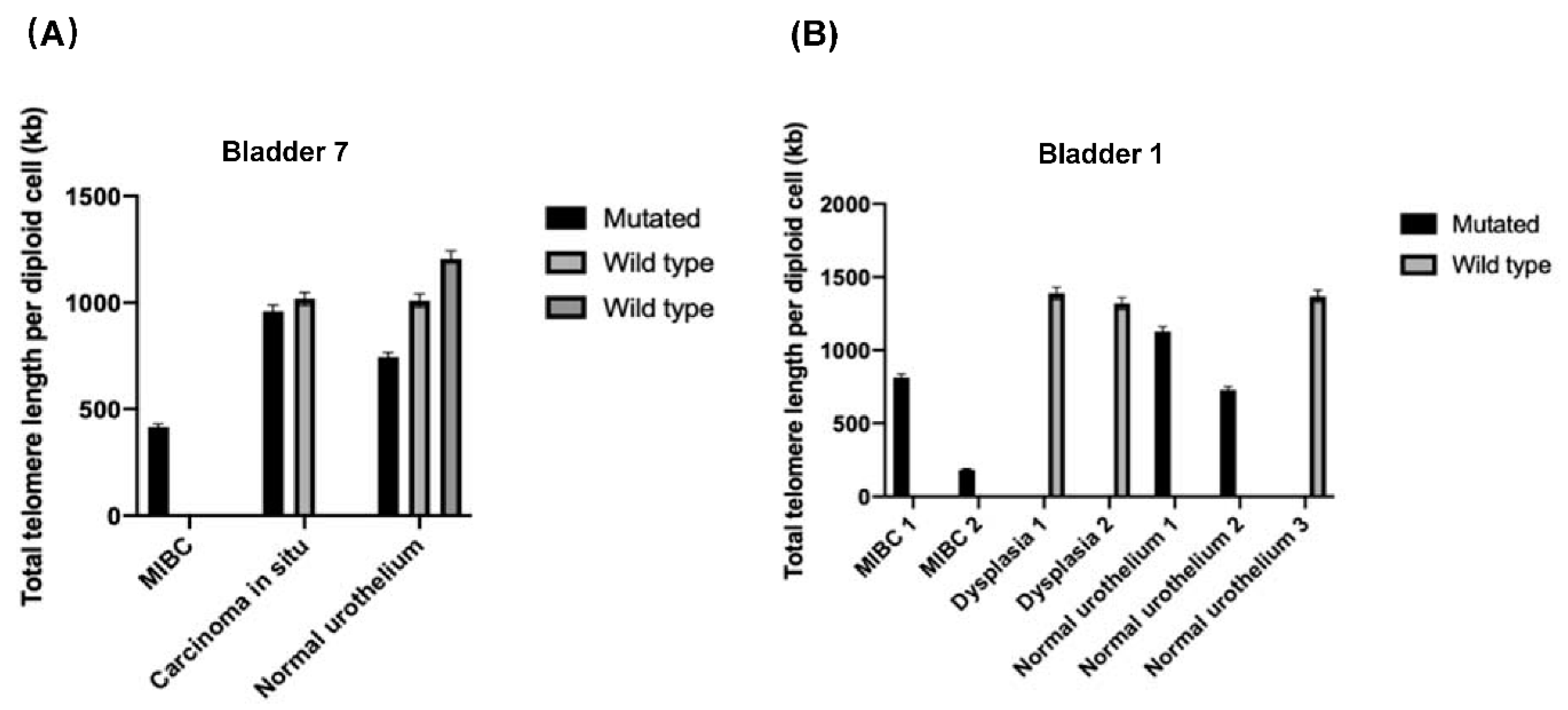
3.3. Telomere Length Analysis within the Whole-Organ Mapping Bladder Specimens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allory, Y.; Beukers, W.; Sagrera, A.; Flandez, M.; Marques, M.; Marquez, M.; van der Keur, K.A.; Dyrskjot, L.; Lurkin, I.; Vermeij, M.; et al. Telomerase reverse transcriptase promoter mutations in bladder cancer: High frequency across stages, detection in urine, and lack of association with outcome. Eur. Urol. 2014, 65, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Chakravarti, D.; LaBella, K.A.; DePinho, R.A. Telomeres: History, health, and hallmarks of aging. Cell 2021, 184, 306–322. [Google Scholar] [CrossRef] [PubMed]
- Gunes, C.; Wezel, F.; Southgate, J.; Bolenz, C. Implications of TERT promoter mutations and telomerase activity in urothelial carcinogenesis. Nat. Rev. Urol. 2018, 15, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Stasik, S.; Salomo, K.; Heberling, U.; Froehner, M.; Sommer, U.; Baretton, G.B.; Ehninger, G.; Wirth, M.P.; Thiede, C.; Fuessel, S. Evaluation of TERT promoter mutations in urinary cell-free DNA and sediment DNA for detection of bladder cancer. Clin. Biochem. 2019, 64, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Hafner, C.; Knuechel, R.; Stoehr, R.; Hartmann, A. Clonality of multifocal urothelial carcinomas: 10 years of molecular genetic studies. Int. J. Cancer 2002, 101, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wullweber, A.; Strick, R.; Lange, F.; Sikic, D.; Taubert, H.; Wach, S.; Wullich, B.; Bertz, S.; Weyerer, V.; Stoehr, R.; et al. Bladder tumor subtype commitment occurs in carcinoma in-situ driven by key signaling pathways including ECM remodeling. Cancer Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Czerniak, B.; Dinney, C.; McConkey, D. Origins of Bladder Cancer. Annu. Rev. Pathol. 2016, 11, 149–174. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, P.A.; Moch, H.; Cubilla, A.L.; Ulbright, T.M.; Reuter, V.E. The 2016 WHO Classification of Tumours of the Urinary System and Male Genital Organs-Part B: Prostate and Bladder Tumours. Eur. Urol. 2016, 70, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Hurst, C.D.; Platt, F.M.; Knowles, M.A. Comprehensive mutation analysis of the TERT promoter in bladder cancer and detection of mutations in voided urine. Eur. Urol. 2014, 65, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.J.; Werner, B.; Barnes, C.P.; Graham, T.A.; Sottoriva, A. Identification of neutral tumor evolution across cancer types. Nat. Genet. 2016, 48, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Heide, T.; Maurer, A.; Eipel, M.; Knoll, K.; Geelvink, M.; Veeck, J.; Knuechel, R.; van Essen, J.; Stoehr, R.; Hartmann, A.; et al. Multiregion human bladder cancer sequencing reveals tumour evolution, bladder cancer phenotypes and implications for targeted therapy. J. Pathol. 2019, 248, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Burger, M.; Catto, J.W.; Dalbagni, G.; Grossman, H.B.; Herr, H.; Karakiewicz, P.; Kassouf, W.; Kiemeney, L.A.; La Vecchia, C.; Shariat, S.; et al. Epidemiology and risk factors of urothelial bladder cancer. Eur. Urol. 2013, 63, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Fujita, K.; Nojima, S.; Tomiyama, E.; Matsushita, M.; Koh, Y.; Nakano, K.; Wang, C.; Ishizuya, Y.; Kato, T.; et al. TERT C228T mutation in non-malignant bladder urothelium is associated with intravesical recurrence for patients with non-muscle invasive bladder cancer. Mol. Oncol. 2020, 14, 2375–2383. [Google Scholar] [CrossRef] [PubMed]
- Bertz, S.; Stohr, R.; Gaisa, N.T.; Wullich, B.; Hartmann, A.; Agaimy, A. TERT promoter mutation analysis as a surrogate to morphology and immunohistochemistry in problematic spindle cell lesions of the urinary bladder. Histopathology 2020, 77, 949–962. [Google Scholar] [CrossRef] [PubMed]
- Weyerer, V.; Eckstein, M.; Comperat, E.; Juette, H.; Gaisa, N.T.; Allory, Y.; Stohr, R.; Wullich, B.; Roupret, M.; Hartmann, A.; et al. Pure Large Nested Variant of Urothelial Carcinoma (LNUC) Is the Prototype of an FGFR3 Mutated Aggressive Urothelial Carcinoma with Luminal-Papillary Phenotype. Cancers 2020, 12, 763. [Google Scholar] [CrossRef] [PubMed]
- Kilian, A.; Bowtell, D.D.; Abud, H.E.; Hime, G.R.; Venter, D.J.; Keese, P.K.; Duncan, E.L.; Reddel, R.R.; Jefferson, R.A. Isolation of a candidate human telomerase catalytic subunit gene, which reveals complex splicing patterns in different cell types. Hum. Mol. Genet. 1997, 6, 2011–2019. [Google Scholar] [CrossRef] [PubMed]
- Borah, S.; Xi, L.; Zaug, A.J.; Powell, N.M.; Dancik, G.M.; Cohen, S.B.; Costello, J.C.; Theodorescu, D.; Cech, T.R. Cancer. TERT promoter mutations and telomerase reactivation in urothelial cancer. Science 2015, 347, 1006–1010. [Google Scholar] [CrossRef] [PubMed]
- Dratwa, M.; Wysoczanska, B.; Turlej, E.; Anisiewicz, A.; Maciejewska, M.; Wietrzyk, J.; Bogunia-Kubik, K. Heterogeneity of telomerase reverse transcriptase mutation and expression, telomerase activity and telomere length across human cancer cell lines cultured in vitro. Exp. Cell Res. 2020, 396, 112298. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Mapping Sample | Gender | Stage | L | V | Pn | WHO Grading 2016 | WHO Grading 1973 | Resection Margin | Number of Positions |
|---|---|---|---|---|---|---|---|---|---|
| Bladder 1 | Male | pT3 | L1 | V0 | P1 | High-grade | G3 | R0 | 21 |
| Bladder 2 | Female | pT3 | L1 | V0 | Pn0 | High-grade | G3 | R0 | 19 |
| Bladder 3 | Male | pT4 | L1 | V1 | Pn0 | High-grade | G3 | R0 | 17 |
| Bladder 4 | Female | pT3 | L1 | V1 | Pn1 | High-grade | G3 | R0 | 18 |
| Bladder 5 | Male | pT3 | L0 | V1 | Pn0 | High-grade | G3 | R0 | 19 |
| Bladder 6 | Female | pT3 | L1 | V0 | Pn0 | High-grade | G3 | R0 | 22 |
| Bladder 7 | Male | pT2 | L0 | V0 | Pn1 | High-grade | G3 | R0 | 15 |
| Bladder 8 | Male | pT3 | L1 | V1 | Pn1 | High-grade | G3 | R2 | 14 |
| Bladder 9 | Female | pT4 | L1 | V1 | Pn1 | High-grade | G3 | R1 | 8 |
| Antibody | Company | Clone | Dilution |
|---|---|---|---|
| CD44 | Dako | DF1485 | 1:40 |
| CK20 | Dako | Ks20.8 | 1:50 |
| P53 | Dako | DO-7 | 1:50 |
| Ki-67 | Dako | MIB-1 | 1:100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weyerer, V.; Eckstein, M.; Strissel, P.L.; Wullweber, A.; Lange, F.; Tögel, L.; Geppert, C.I.; Sikic, D.; Taubert, H.; Wach, S.; et al. TERT Promoter Mutation Analysis of Whole-Organ Mapping Bladder Cancers. Genes 2021, 12, 230. https://doi.org/10.3390/genes12020230
Weyerer V, Eckstein M, Strissel PL, Wullweber A, Lange F, Tögel L, Geppert CI, Sikic D, Taubert H, Wach S, et al. TERT Promoter Mutation Analysis of Whole-Organ Mapping Bladder Cancers. Genes. 2021; 12(2):230. https://doi.org/10.3390/genes12020230
Chicago/Turabian StyleWeyerer, Veronika, Markus Eckstein, Pamela L. Strissel, Adrian Wullweber, Fabienne Lange, Lars Tögel, Carol I. Geppert, Danijel Sikic, Helge Taubert, Sven Wach, and et al. 2021. "TERT Promoter Mutation Analysis of Whole-Organ Mapping Bladder Cancers" Genes 12, no. 2: 230. https://doi.org/10.3390/genes12020230
APA StyleWeyerer, V., Eckstein, M., Strissel, P. L., Wullweber, A., Lange, F., Tögel, L., Geppert, C. I., Sikic, D., Taubert, H., Wach, S., Wullich, B., Hartmann, A., Stoehr, R., & Giedl, J. (2021). TERT Promoter Mutation Analysis of Whole-Organ Mapping Bladder Cancers. Genes, 12(2), 230. https://doi.org/10.3390/genes12020230