
Metabolite Genome-Wide Association Study for Indoleamine 2,3-Dioxygenase Activity Associated with Chronic Kidney Disease


1
Department of Medical Sciences, Graduate School, Soonchunhyang University, Asan 31538, Chungnam, Korea
2
Department of Biomedical Laboratory Science, College of Life and Health Sciences, Hoseo University, Asan 31499, Chungnam, Korea
3
Department of Biomedical Laboratory Science, College of Medical Sciences, Soonchunhyang University, Asan 31538, Chungnam, Korea
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Genes 2021, 12(12), 1905; https://doi.org/10.3390/genes12121905
Submission received: 12 November 2021
/
Revised: 25 November 2021
/
Accepted: 26 November 2021
/
Published: 27 November 2021
(This article belongs to the Section Human Genomics and Genetic Diseases)
Abstract
:Chronic kidney disease (CKD) causes progressive damage to kidney function with increased inflammation. This process contributes to complex amino acid changes. Indoleamine 2,3-dioxygenase (IDO) has been proposed as a new biomarker of CKD in previous studies. In our research, we performed a metabolite genome-wide association study (mGWAS) to identify common and rare variants associated with IDO activity in a Korean population. In addition, single-nucleotide polymorphisms (SNPs) selected through mGWAS were further analyzed for associations with the estimated glomerular filtration rate (eGFR) and CKD. A total of seven rare variants achieved the genome-wide significance threshold (p < 1 × 10−8). Among them, four genes (TNFRSF19, LOC105377444, LOC101928535, and FSTL5) associated with IDO activity showed statistically significant associations with eGFR and CKD. Most of these rare variants appeared specifically in an Asian geographic region. Furthermore, 15 common variants associated with IDO activity were detected in this study and five novel genes (RSU1, PDGFD, SNX25, LOC107984031, and UBASH3B) associated with CKD and eGFR were identified. This study discovered several loci for IDO activity via mGWAS and provided insight into the underlying mechanisms of CKD through association analysis with CKD. To the best of our knowledge, this is the first study to suggest a genetic link between IDO activity and CKD through comparative and integrated analysis.

1. Introduction
Chronic kidney disease (CKD) is caused by several factors, including diabetes, high blood pressure, and glomerulonephritis [1]. CKD can lead to end-stage renal failure due to the gradual loss of kidney function and fibrosis caused by inflammation [2,3]. The Centers for Disease Control and Prevention (CDC) have reported that 90% of adults with CKD are unaware that they have CKD [4]. The early stages of CKD have few signs or symptoms. However, the progression of CKD increases cardiovascular morbidity and mortality, making treatment difficult [5]. Therefore, the management of patients with CKD should focus on delaying disease progression by identifying risk factors through early diagnosis [6].
Metabolomics is a field of interest in nephrology because many metabolites, which are small molecules, are freely filtered by the kidneys [7]. In addition, metabolites that play important roles in numerous biological pathways are also known as potential biomarkers of several diseases, including CKD [8,9,10]. Currently, the estimated glomerular filtration rate (eGFR) calculated by creatinine is mainly used for the diagnosis of kidney function [11]. However, previous studies have reported that it is difficult to use creatinine to detect early kidney damage [12,13]. For this reason, new biomarkers that can identify kidney damage at an early stage are needed.
Indoleamine 2,3-dioxygenase (IDO) is an enzyme involved in the metabolism of tryptophan belonging to the kynurenine pathway. It is induced by pro-inflammatory cytokines [14]. It is evaluated as the kynurenine to tryptophan ratio (K/T ratio) [15]. IDO is also known to play an important role in immunological processes such as infection, autoimmunity, and chronic inflammation [16]. Moreover, Mohib et al. [17] have reported that IDO induced by pro-inflammatory cytokines plays an apoptotic role in renal tubular epithelial cells. Since the kidneys are involved in the clearance of tryptophan metabolites, their impairment is associated with increased tryptophan metabolism and IDO activity [18,19,20]. Interestingly, one study has reported that CKD-induced IDO activity is correlated with CKD severity and major inflammatory markers such as high-sensitivity C-reactive protein (hs-CRP) and soluble TNF-receptor-1 (sTNFR-I) [15]. In addition, Lee et al. [21] have demonstrated an association between IDO activity and CKD in Koreans.
Previous studies examining the genetic influence of CKD patients have estimated the heritability of CKD to be 30–75% [22,23]. In this regard, several studies have identified many genetic loci associated with CKD through genome-wide association analysis [24,25]. Although genome-wide association study (GWAS) may indicate risk for disease, it does not clearly explain the underlying cause. Thus, other research groups have performed an integrated analysis of GWAS with other fields such as metabolomics, proteomics, and transcriptomics to overcome the limitations of GWAS [26,27]. To the best of our knowledge, metabolite GWAS (mGWAS) for IDO activity associated with CKD has not been reported yet. Therefore, this study performed mGWAS to identify common and rare variants associated with IDO activity using the Korean Association REsource (KARE) cohort. To find a biological link between genetic loci and CKD, we further analyzed associations of single-nucleotide polymorphisms (SNPs) related to IDO activity with CKD and eGFR.
2. Materials and Methods
2.1. Ethics Statement
This study was approved by the Institutional Review Board (IRB) of the Korea Disease Control and Prevention Agency (KDCA, KBN-2021-003, 26 January 2021) and Soonchunhyang University (202012-BR-086-01, 15 December 2020). Written informed consent was obtained from all participants.
2.2. Study Participants
This study used the Korean Association REsource (KARE) cohort as part of the Korean Genome and Epidemiology Study (KoGES). The KARE cohort is a community-based cohort in Ansung (rural) and Ansan (urban) areas. Follow-up surveys were conducted every two years from 2001 to 2014. Among them, secondary follow-up data (2005–2006) with metabolite information were used as baseline data. A total of 2579 participants aged 43–74 years were included in the present study. The KARE cohort has been previously described in detail [28]. According to the criteria of the Kidney Disease Improving Global Outcome (KDIGO), participants were classified into CKD (cases, n = 264) and non-CKD (controls, n = 1550) groups based on eGFR. CKD was defined as eGFR levels below 60 mL/min/1.72 m2. Non-CKD was defined as an eGFR of more than 60 mL/min/1.72 m2. Participants with a history of hypertension and diabetes as risk factors for kidney disease and those who were taking medications related to these diseases were excluded from the control group.
2.3. General Characteristics
Anthropometric and biochemical information of participants was obtained from the KARE database. General characteristics of participants are listed in Table 1. Height (m) and weight (kg) were measured using an automated measuring instrument (Dong Sahn Jenix Co., Seoul, Korea) three times to obtain average values. Body mass index (BMI; kg/m2) was calculated as weight/squared height. Blood pressure was measured with a mercury sphygmomanometer (Baumanometer; W.A. Baum, Copiague, NY, USA). Biochemical parameters such as serum creatinine and blood urea nitrogen (BUN) levels were analyzed through blood sampling. Creatinine levels were assessed with the Jaffe method using an automatic analyzer (Hitachi, Tokyo, Japan). eGFR was calculated using the CKD-Epidemiology Collaboration formula as follows: GFR = 141 × min (creatinine/κ, 1)α × max (creatinine/κ, 1)−1.209 × 0.993age × gender (for women, gender = 1.018; α = −0.329; κ = 0.7; for men, gender = 1; α = −0.411; κ = 0.9).
2.4. Metabolite Measurements
To quantify kynurenine and tryptophan, serum samples collected from 2579 participants were analyzed using an AbsoluteIDQ p180 kit (BIOCRATES Life Science, Innsbruck, Austria) according to the manufacturer’s instructions. Liquid chromatography/tandem mass spectrometry (LC–MS/MS) was conducted using an API 4000 QTRAP system (Applied Biosystems, Foster City, CA, USA) equipped with an Agilent 1200 HPLC system (Agilent Technologies, Santa Clara, CA, USA) to measure metabolites. The quality control (QC) process for analyzed metabolites has been described in detail elsewhere [29]. Briefly, both kynurenine and tryptophan used in this study met the following criteria: the coefficient of variance for each metabolite in the reference standards < 25%, 50% of the analyzed metabolite concentrations in the reference standards > limit of detection, and 50% of the analyzed metabolite concentrations in the experimental samples > limit of detection. Pooled human normal serums were used as reference standards. IDO activity was estimated as the ratio of kynurenine to tryptophan.
2.5. Genotyping and Imputation
Genotyping of the KARE dataset was performed using an Affymetrix Genome-Wide Human SNP array 5.0 (Affymetrix, Santa Clara, CA, USA). QC criteria of samples and variants have been previously described [30]. Briefly, samples with low call rates (<96%), DNA contamination, gender inconsistency, and serious concomitant illnesses were excluded. Exclusion criteria for variants were: Hardy–Weinberg equilibrium (HWE) p-values < 1 × 10−6, missing call rates > 5%, and minor allele frequency (MAF) < 0.01. After the QC process, imputation analysis of genetic variants was performed using an IMPUTE2 program with 1000 Genomes Phase I data as a reference panel [31]. A total of 6,461,358 SNPs were included in this study. Locations of variants were assigned using National Center for Biotechnology Information (NCBI) Human Genome Build 37 (hg19).
2.6. Statistical Analysis
All statistical analyses were conducted with PLINK version 1.90 β (https://www.cog-genomics.org/plink2 (accessed on 26 July 2021)) [32]. Linear regression was used to assess associations of variants with IDO and eGFR. A case–control study was performed using logistic regression analysis. All regression analyses were based on an additive model and adjusted for age, gender, area, BMI, hemoglobin A1C (HbA1c), drinking, smoking, systolic blood pressure (SBP), and hs-CRP. The cutoff p-value was p < 5 × 10−8 for rare variants and p < 1 × 10−5 for common variants. Statistical significance between two groups was confirmed via Student’s t-test. After performing mGWAS for IDO activity, linkage disequilibrium (LD) among variants was considered through clumping analysis. Variants were clumped through the following criteria: significance threshold of p < 0.05, LD threshold < 0.5, and physical distance threshold < 1000 kb. The variant with the lowest p-value among clumped variants was selected as the index variant. Manhattan plot and LD block were drawn using the Haploview version 4.1 program (Whitehead Institute for Biomedical Research, Cambridge, MA, USA). Geographical distribution maps for variants were generated based on the 1000 genome database via the Geography of Genetic Variants (GGV) browser (http://popgen.uchicago.edu/ggv/ (accessed on 20 August 2021)). LocusZoom browser (http://locuszoom.org/ (accessed on 16 September 2021)) was used to draw regional plots. Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway database (https://www.genome.jp/kegg/ (accessed on 2 September 2021)) was used to investigate biological processes involved in IDO activity and CKD.
3. Results
3.1. Participant Characteristics
The baseline characteristics of the study population are shown in Table 1.
A total of 2579 participants with metabolite and genotype data were included in this study. To investigate the genetic association of IDO activity and CKD, participants were divided into cases with CKD (n = 264) and controls (n = 1550). The mean age of all participants was 57.10 ± 9.05 years, with the case group being older (mean age: 65.72 ± 6.53 years) than the control group (mean age: 54.98 ± 8.64 years). In addition, values of kidney-related traits such as eGFR, creatinine, and BUN showed significant differences (p < 0.001) between cases and controls.
3.2. Associations between Common Variants and IDO Activity Related to CKD
This study performed mGWAS of IDO activity in the KARE dataset and summarized the association between SNPs and IDO activity through a Manhattan plot (Figure 1).
As a result, 15 SNPs with MAF > 0.05 passed the significant threshold of 1 × 10−5 (Table 2). Of these, the strongest association with IDO activity was observed for rs59178336 in the RSU1 (Ras Suppressor Protein 1) gene located on chromosome 10. We further studied associations of variants related to IDO activity with eGFR and CKD. We found that rs59178336, a variant with the lowest p-value for IDO activity, showed a significant association with CKD (p < 0.05). Minor allele carriers of rs59178336 significantly increased both the IDO level (β = 0.26, p = 9.41 × 10−8) and CKD risk (OR = 1.47, 95% CI: 1.02–2.14, p = 0.041). Although rs12226572 in the UBASH3B (Ubiquitin Associated and SH3 Domain Containing B) gene was significantly associated with CKD (p < 0.025), it did not exhibit a strong LD with surrounding SNPs at the 11q24.1 locus. Three SNPs (rs2513735, p = 0.012; rs78259836, p = 0.024; rs7237751, p = 0.043) in the PDGFD (Platelet-Derived Growth Factor D), SNX25 (Sorting Nexin 25), and LOC107984031 genes were also identified as significant variants for eGFR. Regional plots around the RSU1, PDGFD, SNX25, and LOC107984031 genes revealed several SNPs in the LD, with top SNPs involved in IDO activity (Figure S1). On chromosome 11, rs2513735 near the PDGFD gene showed the most significant result for eGFR (β = −1.45, p = 0.012) among common variants. Additionally, the mitogen-activated protein kinase (MAPK) pathway known to regulate various cellular processes was identified through KEGG pathway analysis of the PDGFD gene (Figure S2).
3.3. Associations between Rare Variants and IDO Activity Related to CKD
A total of seven SNPs reached the GWAS threshold (p < 5 × 10−8), all of which were rare, with MAF < 0.05 (Table 3). Among these, rs182145739, located in the LOC105377444 gene on chromosome 4, reached significance for both IDO activity (p < 5 × 10−8) and CKD (p < 0.05). The minor allele T of rs182145739 was positively associated with the IDO level (β = 0.82, p = 8.46 × 10−10). In addition, CKD risk was increased (OR = 2.63, 95% CI 1.15–6.03, p = 0.022) in minor allele carriers. Association analysis for rare variants with eGFR showed that three loci (near TNFRSF19, TNF Receptor Superfamily Member 19; LOC101928535, FSTL5, Follistatin Like 5) passed a significant threshold of 0.05. Figure 2 shows regional association plots for IDO activity, eGFR, and CKD of TNFRSF19 ± 100 kb (13q12.12) with the LD block structure. Five SNPs (rs76318819, rs117150322, rs180794424, rs148054567, and rs143600269) plotted were significantly associated with both IDO activity and eGFR. Among them, rs117150322 and rs180794424, rs148054567 and rs143600269 were included in the same LD block. Rare variants passing the significant threshold of 1 × 10−5 are shown in Table S1.
3.4. Geographical Distribution of Rare Variants
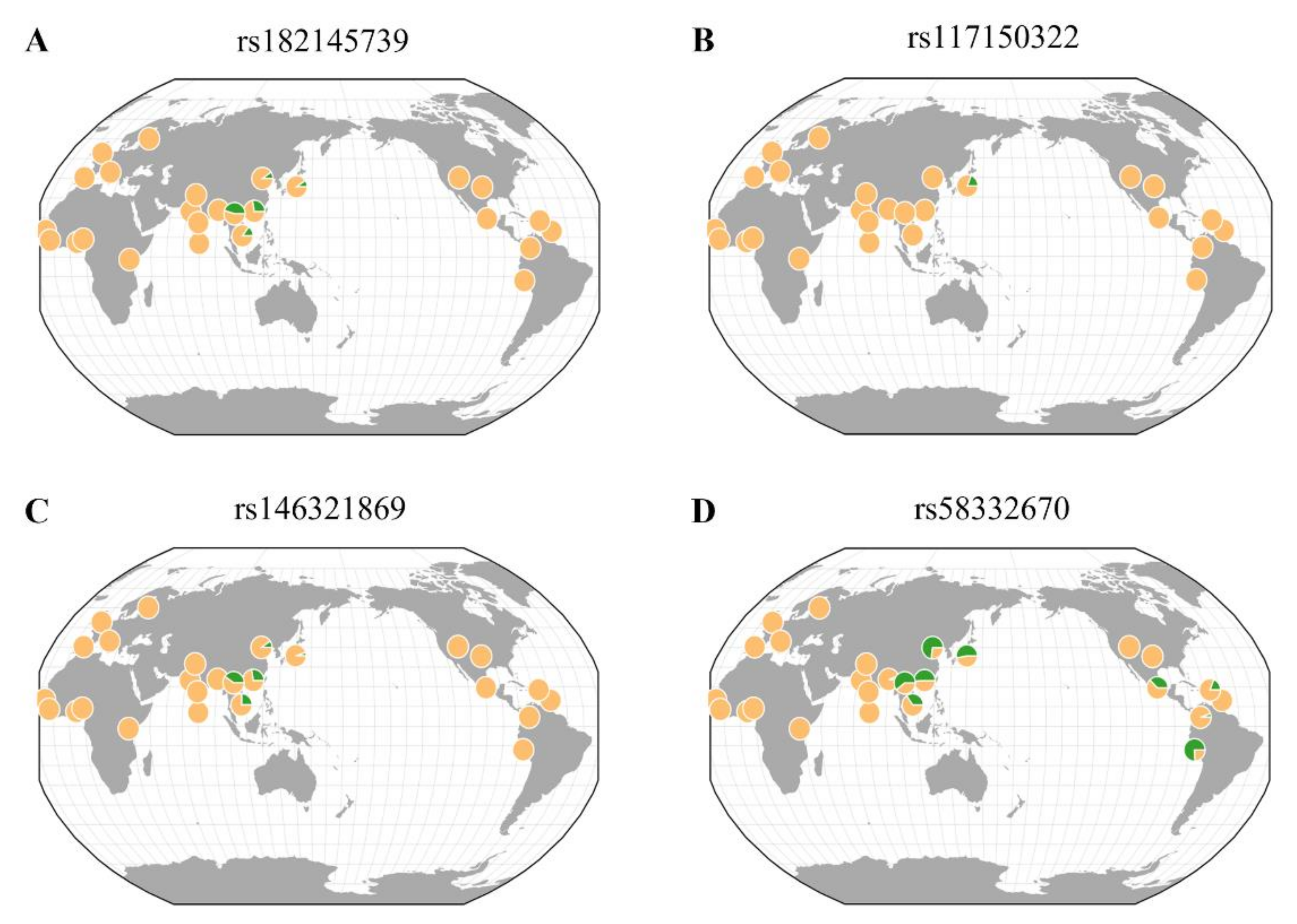
This study further analyzed the geographic distributions of rare variants associated with eGFR and CKD, as well as IDO activity using the GGV browser (Figure 3). Rare variants (rs182145739, rs117150322, and rs146321869) were mostly seen in East Asia. For rs58332670, it was found in East Asia and America. In particular, rs117150322, located near the TNFRSF19 gene, was detected only in a Japanese population (MAF = 0.019). The MAF of rs117150322 in Korea was 0.009, which was lower than that in Japan.
4. Discussion
Several studies have performed metabolite profiling to evaluate physiological pathways for CKD [15,33,34,35]. Interestingly, they have identified an association between CKD and the kynurenine pathway. According to another study, CKD is strongly associated with IDO activity, which degrades tryptophan to kynurenine in the kynurenine pathway [36]. Furthermore, IDO activity was positively correlated with CKD (OR = 12.65, 95% CI: 6.55–24.44) in a Korean population [21]. All of these studies were conducted during epidemiological investigations. Although CKD is a complex disease with high heritability, studies that perform a genetic analysis for the association between IDO activity and CKD have not been reported yet. Therefore, we performed mGWAS to identify genetic variants and potential loci affecting IDO activity associated with CKD in Koreans.
Our results revealed that 15 common variants had significant associations with IDO activity (p < 1 × 10−5) (Table 2) and that seven rare variants reached the GWAS threshold for IDO activity (Table 3). Additionally, SNPs related to IDO activity were analyzed for eGFR and CKD. Among genetic signals associated with eGFR and CKD, common variants were found at the RSU1, PDGFD, SNX25, LOC107984031, and UBASH3B genes and rare variants were identified at the LOC105377444, TNFRSF19, LOC101928535, FSTL5 genes. We focused on the RSU1, PDGFD, SNX25, and TNFRSF19 genes because evidence showing that other genes were associated with CKD was insufficient.
In the case of the RSU1, PDGFD, and TNFRSF19 genes, they were associated with the MAPK pathway [37,38,39]. The SNX25 gene was related to dopamine receptors. MAPK pathways include extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and p38 mitogen-activated protein kinase (p38MAPK) [40]. Although the MAPK pathway is generally known to regulate proliferation, many studies have suggested that it is an intracellular signaling pathway underlying kidney development [40,41,42]. Moreover, Fujigaki et al. reported that IDO activity is related to the MAPK pathways [43]. Dopamine receptors exist as D1-like (D1R and D5R) and D2-like (D2R, D3R, and D4R) subtypes according to their structure and pharmacology [44]. Among them, D1R is widely expressed in the kidney. It plays a central role in regulating blood pressure and sodium balance [45,46].
A recent study has reported that RSU1 is a critical mediator in downregulating ERK signaling through extracellular matrix (ECM) detachment [37]. The ERK pathway responsible for basic cellular processes is the most important signaling cascade of MAPK pathways [47]. It has also been reported that reduced ERK activity can improve antioxidant effects and kidney function [48]. In the present study, our results showed that, among common variants, rs59178336, located in the intron of the RSU1 gene, was significantly associated with IDO activity and CKD. Interestingly, Reznichenko et al. [49] have confirmed that the CUBN gene, located close to the RSU1 gene, is associated with end-stage renal disease. Therefore, the RSU1 gene might be associated with kidney disease.
Our results also identified associations of regions near the PDGFD gene involved in ERK signaling with IDO activity and eGFR (Table 2, Figure S2) [38]. Similar to our results, a previous study has reported that a genetic variant (rs7103465) in the PDGFD gene is associated with the ratio of urine albumin to creatinine (p = 3.7 × 10−7) in Latin Americans [50]. Moreover, it has been reported that PDGFD is overexpressed in hepatic and renal fibrosis [51,52]. Charni et al. demonstrated that PDGFD is regulated by TGF-β, which activates MAPK pathways such as ERK, JNK, and p38MAPK [53].
Furthermore, it has been reported that SNX25 overexpression is associated with increased expression levels and signaling of D1R [54]. Another study has indicated that SNX5 depletion can result in hypertension in normotensive mice [55]. Therefore, the SNX25 gene encoding protein SNX25 might be associated with hypertension, a risk factor for kidney disease. In our data, minor carriers of rs78259836 belonging to the SNX25 gene increased IDO activity but decreased eGFR (Table 2).
Previous studies have reported that an understanding of progenitor cells involved in kidney damage and repair can provide insight into renal pathology and identify novel therapeutic targets [56,57]. Schutgens et al. [58] have demonstrated that TNFRSF19 is a marker gene for epithelial progenitor cells that contributes to adult kidney development in vivo. Moreover, previous studies have reported that overexpression of the TNFRSF19 gene can activate the JNK pathway and that its activation causes damage and fibrosis in the human kidney [39,59]. The JNK pathway involved in inflammation has also been reported as a mechanism regulating IDO [60,61]. Through mGWAS analysis, we discovered several SNPs located near the TNFRSF19 gene associated with IDO activity (Table 3, Figure 2). The results of our study suggest that the TNFRSF19 gene might regulate CKD by inducing IDO through JNK, which belongs to the MAPK family.
In summary, this study performed an mGWAS for IDO activity obtained from 2579 participants in the KARE cohort. A total of 22 novel SNPs (15 common and 7 rare variants) were found and further analyzed for genetic associations with eGFR and CKD. For variants selected based on metabolites, we investigated their associations with CKD compared to other study groups. As a result, four genes (RSU1, PDGFD, SNX25, and TNFRSF19) were associated with CKD by regulating IDO activity. In particular, our data highlight that the RSU1 and PDGFD genes are potential mediators of CKD associated with ERK, which belongs to the MAPK family. The results of this study also suggest that rare variants of the TNFRSF19 gene are associated with CKD, specifically in Asians, through the JNK pathway. The gene–metabolite associations identified in our study provide insight into the underlying mechanisms for CKD. However, functional analyses of mRNA expression levels and proteins should be performed to validate these findings. In addition, modern technological advances have made metabolite analysis possible, but the number of samples is still limited. Therefore, replication studies in other cohorts are needed to confirm the accuracy of this study.
Supplementary Materials
The following are available online athttps://www.mdpi.com/article/10.3390/genes12121905/s1, Figure S1: Regional association plots of IDO activity at 10p13, 1p12, 4q35.1, and 18q21.1 loci. The statistical significances between the SNPs near RSU1 (A), PDGFD (B), SNX25 (C), and LOC107984031 (D) genes and IDO activity are plotted as −log10 p values. The purple diamond represents the SNP most strongly associated with IDO activity. Levels of linkage disequilibrium (r2) of top SNPs and surrounding SNPs are shown in different colors. The regional plots for SNPs were generated via LocusZoom browser (http://locuszoom.org/ (accessed on 16 September 2021)). Figure S2: The KEGG pathway for MAPK signaling pathway. A red box indicates the PDGFD gene found in growth factor (GF) according to KEGG. The image of a pathway was generated via KEGG browser (https://www.genome.jp/kegg/ (accessed on 2 September 2021)). Table S1: Rare variants associated with indoleamine 2,3-dioxygenase activity in Koreans.
Author Contributions
Conceptualization, H.-R.K. and H.-S.J.; methodology, H.-S.J.; software, H.-R.K.; validation, H.-R.K., H.-S.J. and Y.-B.E.; investigation, H.-R.K.; resources, Y.-B.E.; data curation, H.-R.K. and H.-S.J.; writing—original draft preparation, H.-R.K. and H.-S.J.; writing—review and editing, H.-S.J. and Y.-B.E.; supervision, Y.-B.E.; project administration, Y.-B.E.; funding acquisition, Y.-B.E. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Soonchunhyang University research fund and a National Research Foundation of Korea (NRF) grant funded by the Korean government (MSIT) (NRF-2020R1F1A1071977).
Institutional Review Board Statement
This study was approved by the Institutional Review Board (IRB) of the Korea Disease Control and Prevention Agency (KDCA, KBN-2021-003, 26 January 2021) and Soonchunhyang University (202012-BR-086-01, 15 December 2020).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to ethnical concerns.
Conflicts of Interest
All authors declare no conflict of interest.
References
- Parmar, M.S. Chronic renal disease. BMJ 2002, 325, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Levey, A.; Atkins, R.; Coresh, J.; Cohen, E.; Collins, A.; Eckardt, K.-U.; Nahas, M.; Jaber, B.; Jadoul, M.; Levin, A. Chronic kidney disease as a global public health problem: Approaches and initiatives–a position statement from Kidney Disease Improving Global Outcomes. Kidney Int. 2007, 72, 247–259. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.M. Inflammatory mediators and renal fibrosis. Adv. Exp. Med. Biol. 2019, 1165, 381–406. [Google Scholar] [PubMed]
- Center for Disease Control and Prevention (CDC). Chronic Kidney Disease in the United States. 2021. Available online: https://www.cdc.gov/kidneydisease/publications-resources/CKD-national-facts.html (accessed on 20 September 2021).
- Wouters, O.J.; O’donoghue, D.J.; Ritchie, J.; Kanavos, P.G.; Narva, A.S. Early chronic kidney disease: Diagnosis, management and models of care. Nat. Rev. Nephrol. 2015, 11, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krol, E.; Rutkowski, B.; Czarniak, P.; Kraszewska, E.; Lizakowski, S.; Szubert, R.; Czekalski, S.; Sulowicz, W.; Wiecek, A. Early detection of chronic kidney disease: Results of the PolNef study. Am. J. Nephrol. 2009, 29, 264–273. [Google Scholar] [PubMed] [Green Version]
- Köttgen, A.; Raffler, J.; Sekula, P.; Kastenmüller, G. Genome-wide association studies of metabolite concentrations (mGWAS): Relevance for nephrology. Semin. Nephrol. 2018, 38, 151–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, A.; Sun, H.; Yan, G.; Wang, P.; Wang, X. Metabolomics for biomarker discovery: Moving to the clinic. Biomed. Res. Int. 2015, 2015, 354671. [Google Scholar] [CrossRef]
- Graham, S.F.; Chevallier, O.P.; Roberts, D.; Holscher, C.; Elliott, C.T.; Green, B.D. Investigation of the human brain metabolome to identify potential markers for early diagnosis and therapeutic targets of Alzheimer’s disease. Anal. Chem. 2013, 85, 1803–1811. [Google Scholar] [CrossRef]
- Zhao, Y.Y. Metabolomics in chronic kidney disease. Clin. Chim. Acta. 2013, 422, 59–69. [Google Scholar] [CrossRef]
- Levey, A.S.; Becker, C.; Inker, L.A. Glomerular filtration rate and albuminuria for detection and staging of acute and chronic kidney disease in adults: A systematic review. JAMA 2015, 313, 837–846. [Google Scholar] [CrossRef] [Green Version]
- Toffaletti, J.G. Clarifying the Confusion of GFRs, Creatinine, and Cystatin C. 2018. Available online: https://acutecaretesting.org/ (accessed on 9 September 2021).
- Perrone, R.D.; Madias, N.E.; Levey, A.S. Serum creatinine as an index of renal function: New insights into old concepts. Clin. Chem. 1992, 38, 1933–1953. [Google Scholar] [CrossRef]
- Chen, Y.; Guillemin, G.J. Kynurenine pathway metabolites in humans: Disease and healthy States. Int. J. Tryptophan Res. 2009, 2, IJTR.S2097. [Google Scholar] [CrossRef] [Green Version]
- Schefold, J.C.; Zeden, J.P.; Fotopoulou, C.; von Haehling, S.; Pschowski, R.; Hasper, D.; Volk, H.D.; Schuett, C.; Reinke, P. Increased indoleamine 2,3-dioxygenase (IDO) activity and elevated serum levels of tryptophan catabolites in patients with chronic kidney disease: A possible link between chronic inflammation and uraemic symptoms. Nephrol. Dial. Transplant. 2009, 24, 1901–1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mbongue, J.C.; Nicholas, D.A.; Torrez, T.W.; Kim, N.S.; Firek, A.F.; Langridge, W.H. The role of indoleamine 2, 3-dioxygenase in immune suppression and autoimmunity. Vaccines 2015, 3, 703–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohib, K.; Guan, Q.; Diao, H.; Du, C.; Jevnikar, A.M. Proapoptotic activity of indoleamine 2,3-dioxygenase expressed in renal tubular epithelial cells. Am. J. Physiol.-Renal Physiol. 2007, 293, F801–F812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, K.; Fujigaki, S.; Heyes, M.P.; Shibata, K.; Takemura, M.; Fujii, H.; Wada, H.; Noma, A.; Seishima, M. Mechanism of increases in L-kynurenine and quinolinic acid in renal insufficiency. Am. J. Physiol. Renal. Physiol. 2000, 279, F565–F572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawlak, D.; Tankiewicz, A.; Mysliwiec, P.; Buczko, W. Tryptophan metabolism via the kynurenine pathway in experimental chronic renal failure. Nephron 2002, 90, 328–335. [Google Scholar] [CrossRef]
- Aregger, F.; Uehlinger, D.E.; Fusch, G.; Bahonjic, A.; Pschowski, R.; Walter, M.; Schefold, J.C. Increased urinary excretion of kynurenic acid is associated with non-recovery from acute kidney injury in critically ill patients. BMC Nephrol. 2018, 19, 44. [Google Scholar] [CrossRef]
- Lee, H.; Jang, H.B.; Yoo, M.G.; Park, S.I.; Lee, H.J. Amino acid metabolites associated with chronic kidney disease: An Eight-Year Follow-Up Korean Epidemiology Study. Biomedicines 2020, 8, 222. [Google Scholar] [CrossRef]
- Canadas-Garre, M.; Anderson, K.; Cappa, R.; Skelly, R.; Smyth, L.J.; McKnight, A.J.; Maxwell, A.P. Genetic Susceptibility to Chronic Kidney Disease—Some More Pieces for the Heritability Puzzle. Front. Genet. 2019, 10, 453. [Google Scholar] [CrossRef] [PubMed]
- MacCluer, J.W.; Scavini, M.; Shah, V.O.; Cole, S.A.; Laston, S.L.; Voruganti, V.S.; Paine, S.S.; Eaton, A.J.; Comuzzie, A.G.; Tentori, F.; et al. Heritability of measures of kidney disease among Zuni Indians: The Zuni Kidney Project. Am. J. Kidney Dis. 2010, 56, 289–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tin, A.; Kottgen, A. Genome-Wide Association Studies of CKD and Related Traits. Clin. J. Am. Soc. Nephrol. 2020, 15, 1643–1656. [Google Scholar] [CrossRef] [PubMed]
- Boger, C.A.; Heid, I.M. Chronic kidney disease: Novel insights from genome-wide association studies. Kidney Blood Press. Res. 2011, 34, 225–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzoni, C.; Kia, D.A.; Vandrovcova, J.; Hardy, J.; Wood, N.W.; Lewis, P.A.; Ferrari, R. Genome, transcriptome and proteome: The rise of omics data and their integration in biomedical sciences. Brief. Bioinform. 2018, 19, 286–302. [Google Scholar] [CrossRef]
- Liu, L.; Kiryluk, K. Insights into CKD from Metabolite GWAS. J. Am. Soc. Nephrol. 2018, 29, 1349–1351. [Google Scholar] [CrossRef]
- Kim, Y.; Han, B.G.; KoGES Group. Cohort Profile: The Korean Genome and Epidemiology Study (KoGES) Consortium. Int. J. Epidemiol. 2017, 46, 1350. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Xu, T.; Lee, Y.; Kim, N.-H.; Kim, Y.-J.; Kim, J.-M.; Cho, S.Y.; Kim, K.-Y.; Nam, M.; Adamski, J. Identification of putative biomarkers for type 2 diabetes using metabolomics in the Korea Association REsource (KARE) cohort. Metabolomics 2016, 12, 178. [Google Scholar] [CrossRef]
- Cho, Y.S.; Go, M.J.; Kim, Y.J.; Heo, J.Y.; Oh, J.H.; Ban, H.-J.; Yoon, D.; Lee, M.H.; Kim, D.-J.; Park, M. A large-scale genome-wide association study of Asian populations uncovers genetic factors influencing eight quantitative traits. Nat. Genet. 2009, 41, 527–534. [Google Scholar] [CrossRef]
- Howie, B.N.; Donnelly, P.; Marchini, J. A flexible and accurate genotype imputation method for the next generation of genome-wide association studies. PLoS Genet. 2009, 5, e1000529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Mor, A.; Kalaska, B.; Pawlak, D. Kynurenine pathway in chronic kidney disease: What’s old, what’s new, and what’s next? Int. J. Tryptophan Res. 2020, 13, 1178646920954882. [Google Scholar] [CrossRef]
- Pawlak, K.; Kowalewska, A.; Pawlak, D.; Mysliwiec, M. Kynurenine and its metabolites—Kynurenic acid and anthranilic acid are associated with soluble endothelial adhesion molecules and oxidative status in patients with chronic kidney disease. Am. J. Med. Sci. 2009, 338, 293–300. [Google Scholar] [CrossRef]
- Rhee, E.P.; Clish, C.B.; Ghorbani, A.; Larson, M.G.; Elmariah, S.; McCabe, E.; Yang, Q.; Cheng, S.; Pierce, K.; Deik, A.; et al. A combined epidemiologic and metabolomic approach improves CKD prediction. J. Am. Soc. Nephrol. 2013, 24, 1330–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goek, O.N.; Prehn, C.; Sekula, P.; Romisch-Margl, W.; Doring, A.; Gieger, C.; Heier, M.; Koenig, W.; Wang-Sattler, R.; Illig, T.; et al. Metabolites associate with kidney function decline and incident chronic kidney disease in the general population. Nephrol. Dial. Transplant. 2013, 28, 2131–2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Liu, J.; Tu, Y.; Zhao, Z.; Qu, J.; Chen, K.; Chen, Y.; Sun, Y.; Zhao, H.; Deng, Y.; et al. RSU-1 interaction with prohibitin-2 links cell-extracellular matrix detachment to downregulation of ERK signaling. J. Biol. Chem. 2021, 296, 100109. [Google Scholar] [CrossRef]
- Najy, A.J.; Won, J.J.; Movilla, L.S.; Kim, H.-R.C. Differential tumorigenic potential and matriptase activation between PDGF B versus PDGF D in prostate cancer. Mol. Cancer Res. 2012, 10, 1087–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eby, M.T.; Jasmin, A.; Kumar, A.; Sharma, K.; Chaudhary, P.M. TAJ, a novel member of the tumor necrosis factor receptor family, activates the c-Jun N-terminal kinase pathway and mediates caspase-independent cell death. J. Biol. Chem. 2000, 275, 15336–15342. [Google Scholar] [CrossRef] [Green Version]
- Kurtzeborn, K.; Kwon, H.N.; Kuure, S. MAPK/ERK signaling in regulation of renal differentiation. Int. J. Mol. Sci. 2019, 20, 1779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hida, M.; Omori, S.; Awazu, M. ERK and p38 MAP kinase are required for rat renal development. Kidney Int. 2002, 61, 1252–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ihermann-Hella, A.; Lume, M.; Miinalainen, I.J.; Pirttiniemi, A.; Gui, Y.; Peranen, J.; Charron, J.; Saarma, M.; Costantini, F.; Kuure, S. Mitogen-activated protein kinase (MAPK) pathway regulates branching by remodeling epithelial cell adhesion. PLoS Genet. 2014, 10, e1004193. [Google Scholar] [CrossRef] [Green Version]
- Fujigaki, H.; Saito, K.; Fujigaki, S.; Takemura, M.; Sudo, K.; Ishiguro, H.; Seishima, M. The signal transducer and activator of transcription 1alpha and interferon regulatory factor 1 are not essential for the induction of indoleamine 2,3-dioxygenase by lipopolysaccharide: Involvement of p38 mitogen-activated protein kinase and nuclear factor-kappaB pathways, and synergistic effect of several proinflammatory cytokines. J. Biochem. 2006, 139, 655–662. [Google Scholar] [PubMed]
- Beaulieu, J.M.; Espinoza, S.; Gainetdinov, R.R. Dopamine receptors–IUPHAR Review 13. Br. J. Pharmacol. 2015, 172, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Armando, I.; Villar, V.A.; Jose, P.A. Dopamine and renal function and blood pressure regulation. Compr. Physiol. 2011, 1, 1075–1117. [Google Scholar] [PubMed]
- Harris, R.C.; Zhang, M.Z. Dopamine, the kidney, and hypertension. Curr. Hypertens. Rep. 2012, 14, 138–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pat, B.; Yang, T.; Kong, C.; Watters, D.; Johnson, D.W.; Gobe, G. Activation of ERK in renal fibrosis after unilateral ureteral obstruction: Modulation by antioxidants. Kidney Int. 2005, 67, 931–943. [Google Scholar] [CrossRef] [PubMed]
- Reznichenko, A.; Snieder, H.; van den Born, J.; de Borst, M.H.; Damman, J.; van Dijk, M.C.; van Goor, H.; Hepkema, B.G.; Hillebrands, J.-L.; Leuvenink, H.G. CUBN as a novel locus for end-stage renal disease: Insights from renal transplantation. PLoS ONE 2012, 7, e36512. [Google Scholar] [CrossRef] [Green Version]
- Qian, H.; Kowalski, M.H.; Kramer, H.J.; Tao, R.; Lash, J.P.; Stilp, A.M.; Cai, J.; Li, Y.; Franceschini, N. Genome-wide association of kidney traits in hispanics/latinos using dense imputed whole-genome sequencing data: The hispanic community health study/study of latinos. Circ. Genom. Precis. Med. 2020, 13, e002891. [Google Scholar] [CrossRef] [PubMed]
- Ostendorf, T.; van Roeyen, C.R.; Peterson, J.D.; Kunter, U.; Eitner, F.; Hamad, A.J.; Chan, G.; Jia, X.C.; Macaluso, J.; Gazit-Bornstein, G.; et al. A fully human monoclonal antibody (CR002) identifies PDGF-D as a novel mediator of mesangioproliferative glomerulonephritis. J. Am. Soc. Nephrol. 2003, 14, 2237–2247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhanna, N.; Doron, S.; Wald, O.; Horani, A.; Eid, A.; Pappo, O.; Friedman, S.L.; Safadi, R. Activation of hepatic stellate cells after phagocytosis of lymphocytes: A novel pathway of fibrogenesis. Hepatology 2008, 48, 963–977. [Google Scholar] [CrossRef] [Green Version]
- Charni Chaabane, S.; Coomans de Brachene, A.; Essaghir, A.; Velghe, A.; Lo Re, S.; Stockis, J.; Lucas, S.; Khachigian, L.M.; Huaux, F.; Demoulin, J.B. PDGF-D expression is down-regulated by TGFbeta in fibroblasts. PLoS ONE 2014, 9, e108656. [Google Scholar] [CrossRef] [Green Version]
- Free, R.B.; Hazelwood, L.A.; Spalding, H.N.; Cabrera, D.M.; Sibley, D.R. Sorting nexin-25, a novel member of the dopamine receptor signalplex, up-regulates D1 and D2 dopamine receptor expression in HEK293 cells. FASEB J. 2007, 21, A423. [Google Scholar] [CrossRef]
- Yang, J.; Armando, I.; Jones, J.; Zeng, C.; Jose, P.; Villar, V. Sorting nexins: New determinants for the development of hypertension. Ann. Clin. Exp. Hypertens. 2014, 2, 1008. [Google Scholar]
- Huang, J.; Kong, Y.; Xie, C.; Zhou, L. Stem/progenitor cell in kidney: Characteristics, homing, coordination, and maintenance. Stem. Cell Res. Ther. 2021, 12, 197. [Google Scholar] [CrossRef] [PubMed]
- Chou, Y.H.; Pan, S.Y.; Yang, C.H.; Lin, S.L. Stem cells and kidney regeneration. J. Formos. Med. Assoc. 2014, 113, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Schutgens, F.; Rookmaaker, M.B.; Blokzijl, F.; van Boxtel, R.; Vries, R.; Cuppen, E.; Verhaar, M.C.; Clevers, H. Troy/TNFRSF19 marks epithelial progenitor cells during mouse kidney development that continue to contribute to turnover in adult kidney. Proc. Natl. Acad. Sci. USA 2017, 114, E11190–E11198. [Google Scholar] [CrossRef] [Green Version]
- Grynberg, K.; Ma, F.Y.; Nikolic-Paterson, D.J. The JNK signaling pathway in renal fibrosis. Front. Physiol. 2017, 8, 829. [Google Scholar] [CrossRef]
- Mei, J.; Li, M.Q.; Ding, D.; Li, D.J.; Jin, L.P.; Hu, W.G.; Zhu, X.Y. Indoleamine 2,3-dioxygenase-1 (IDO1) enhances survival and invasiveness of endometrial stromal cells via the activation of JNK signaling pathway. Int. J. Clin. Exp. Pathol. 2013, 6, 431–444. [Google Scholar]
- Opitz, C.A.; Litzenburger, U.M.; Opitz, U.; Sahm, F.; Ochs, K.; Lutz, C.; Wick, W.; Platten, M. The indoleamine-2,3-dioxygenase (IDO) inhibitor 1-methyl-D-tryptophan upregulates IDO1 in human cancer cells. PLoS ONE 2011, 6, e19823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Manhattan plot of results from metabolite GWAS analysis of IDO activity. Red line indicates genome-wide significance threshold at p < 1 × 10−8. Blue line represents significance threshold at p < 1 × 10−5. Vertical axis indicates −log10 p values from linear regression adjusted for age, area, gender, BMI, drinking, smoking, SBP, hs-CRP, and HbA1c. Horizontal axis shows chromosomal positions. The Manhattan plot was generated with the Haploview program.
Figure 1.
Manhattan plot of results from metabolite GWAS analysis of IDO activity. Red line indicates genome-wide significance threshold at p < 1 × 10−8. Blue line represents significance threshold at p < 1 × 10−5. Vertical axis indicates −log10 p values from linear regression adjusted for age, area, gender, BMI, drinking, smoking, SBP, hs-CRP, and HbA1c. Horizontal axis shows chromosomal positions. The Manhattan plot was generated with the Haploview program.
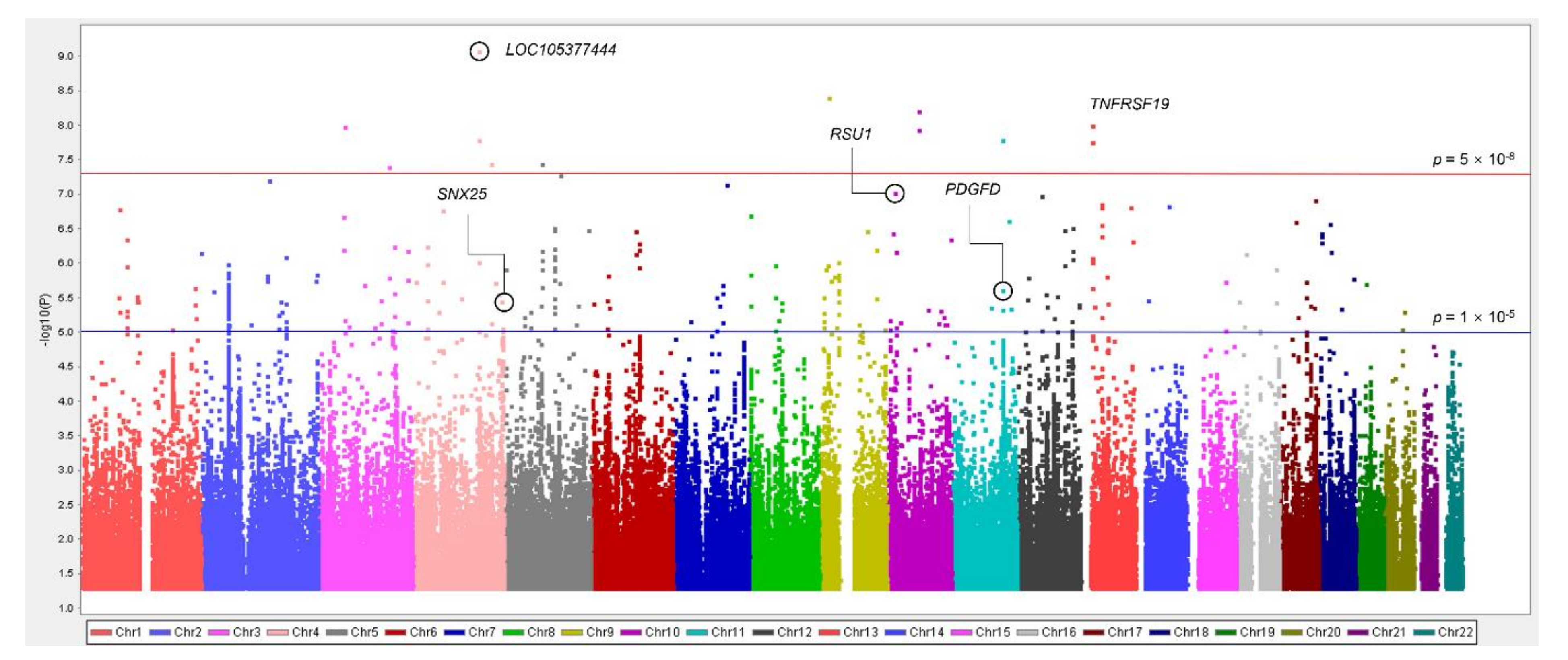
Figure 2.
Plot of p-values of studied variants within TNFRSF19 ± 100 kb regions. Results of association analysis between variants and IDO activity, eGFR, and CKD are plotted. Standard significant p-value threshold (p = 0.05) and GWAS p-value threshold (p = 5 × 10−8) are indicated by blue and red lines, respectively. The bottom panel shows a Haploview of LD (r2) based on genotyping data from KARE data. It was generated using the Haploview program.
Figure 2.
Plot of p-values of studied variants within TNFRSF19 ± 100 kb regions. Results of association analysis between variants and IDO activity, eGFR, and CKD are plotted. Standard significant p-value threshold (p = 0.05) and GWAS p-value threshold (p = 5 × 10−8) are indicated by blue and red lines, respectively. The bottom panel shows a Haploview of LD (r2) based on genotyping data from KARE data. It was generated using the Haploview program.
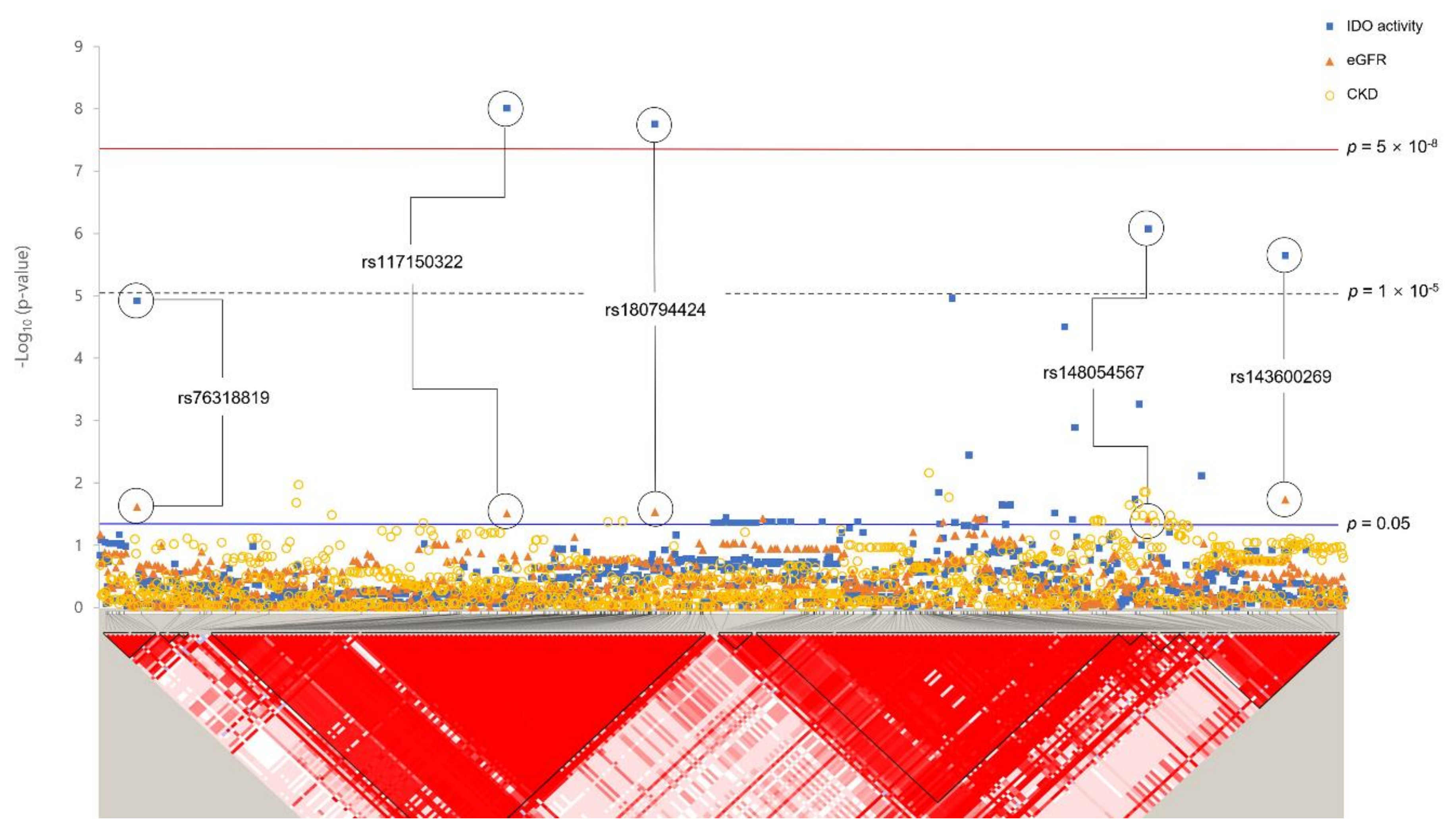
Figure 3.
Geographical distributions of rare variants associated with eGFR and CKD. Each pie chart shows a minor allele frequency based on 1000 genomes (hg 19). The frequency scale of the green pie is the proportion out of 0.1. (A) rs182145739, (B) rs117150322, (C) rs146321869, (D) rs58332670.
Figure 3.
Geographical distributions of rare variants associated with eGFR and CKD. Each pie chart shows a minor allele frequency based on 1000 genomes (hg 19). The frequency scale of the green pie is the proportion out of 0.1. (A) rs182145739, (B) rs117150322, (C) rs146321869, (D) rs58332670.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characteristics of participants in the Korean population.
| Characteristics | Quantitative Trait Analysis | Case–Control Analysis for CKD | ||
|---|---|---|---|---|
| Controls | Cases | p-Value * | ||
| Number of participants | 2579 | 1550 | 264 | |
| Gender [men (%)] | 1218 (47.23) | 789 (50.90) | 81 (30.68) | <0.001 |
| Age (M years ± SD) | 57.10 ± 9.05 | 54.98 ± 8.64 | 65.72 ± 6.53 | <0.001 |
| Height (M cm ± SD) | 159.55 ± 9.16 | 160.55 ± 8.98 | 155.42 ± 8.28 | <0.001 |
| Weight (M kg ± SD) | 62.63 ± 10.36 | 62.30 ± 10.41 | 60.88 ± 9.53 | 0.042 |
| BMI (M kg/m2 ± SD) | 24.56 ± 3.23 | 24.11 ± 3.09 | 25.20 ± 3.47 | <0.001 |
| eGFR (mL/min/1.73 m2) | 75.58 ± 11.92 | 78.68 ± 9.69 | 55.24 ± 9.21 | <0.001 |
| Creatinine (mg/dL) | 0.98 ± 0.20 | 0.96 ± 0.14 | 1.18 ± 0.42 | <0.001 |
| BUN (mg/dL) | 15.69 ± 4.26 | 15.33 ± 3.92 | 17.91 ± 5.45 | <0.001 |
BMI, body mass index; eGFR, estimated glomerular filtration rate; BUN, blood urea nitrogen; CKD, chronic kidney disease; M, mean value; SD, standard deviation. *, Significant differences in characteristics between cases and controls were determined with Student’s t-test.
Table 2.
Common variants associated with indoleamine 2,3-dioxygenase activity in Koreans.
| No. | SNP | Nearest Gene | Chromosome Position | Minor Allele | MAF | Function | IDO Activity | eGFR | CKD | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β ± S.E. | p-Value | β ± S.E | p-Value | OR (95% CI) | p-Value | |||||||
| 1 | rs59178336 | RSU1 | 10:16822091 | C | 0.095 | Intron | 0.26 ± 0.049 | 9.41 × 10−8 | −0.59 ± 0.50 | 0.235 | 1.47 (1.02–2.14) | 0.041 |
| 2 | rs10469937 | CCDC85A | 2:56629317 | C | 0.435 | - | −0.14 ± 0.028 | 1.03 × 10−6 | 0.35 ± 0.29 | 0.228 | 0.91 (0.73–1.14) | 0.407 |
| 3 | rs7588698 | HDAC4 | 2:240041896 | A | 0.055 | Intron | 0.30 ± 0.062 | 1.42 × 10−6 | −0.78 ± 0.64 | 0.225 | 1.43 (0.88–2.32) | 0.145 |
| 4 | rs2513735 | PDGFD | 11:104081184 | T | 0.065 | - | 0.27 ± 0.057 | 2.39 × 10−6 | −1.45 ± 0.58 | 0.012 | 1.08 (0.70–1.67) | 0.730 |
| 5 | rs6730950 | RTN4 | 2:55386276 | C | 0.149 | - | 0.19 ± 0.040 | 2.90 × 10−6 | −0.75 ± 0.41 | 0.065 | 1.06 (0.78–1.43) | 0.721 |
| 6 | rs1094818 | WARS2 | 1:119238523 | G | 0.078 | - | 0.25 ± 0.053 | 3.49 × 10−6 | −0.95 ± 0.54 | 0.080 | 1.34 (0.91–1.96) | 0.140 |
| 7 | rs78549225 | RBFOX1 | 16:6969406 | G | 0.119 | Intron | 0.21 ± 0.044 | 3.50 × 10−6 | −0.66 ± 0.46 | 0.148 | 1.24 (0.88–1.74) | 0.222 |
| 8 | rs78259836 | SNX25 | 4:186261649 | A | 0.105 | Intron | 0.21 ± 0.046 | 3.54 × 10−6 | −1.07 ± 0.47 | 0.024 | 1.12 (0.79–1.57) | 0.522 |
| 9 | rs7237751 | LOC107984031 | 18:47255785 | G | 0.139 | Upstream | 0.19 ± 0.041 | 4.42 × 10−6 | −0.86 ± 0.42 | 0.043 | 1.27 (0.93–1.74) | 0.137 |
| 10 | rs12226572 | UBASH3B | 11:122648650 | A | 0.054 | Intron | 0.29 ± 0.062 | 4.54 × 10−6 | −1.05 ± 0.64 | 0.100 | 1.64 (1.07–2.51) | 0.025 |
| 11 | rs143090547 | PLPP1 | 5:54753484 | T | 0.056 | Intron | 0.28 ± 0.062 | 5.00 × 10−6 | −1.23 ± 0.63 | 0.051 | 1.37 (0.86–2.19) | 0.190 |
| 12 | rs17608925 | ORMDL3 | 17:38082831 | C | 0.066 | Intron | 0.26 ± 0.056 | 5.84 × 10−6 | −0.01 ± 0.58 | 0.989 | 1.36 (0.90–2.05) | 0.147 |
| 13 | rs73192989 | RBM19 | 12:114580187 | T | 0.080 | - | 0.23 ± 0.051 | 6.77 × 10−6 | −0.75 ± 0.53 | 0.155 | 1.00 (0.67–1.50) | 0.989 |
| 14 | rs199564331 | BRINP3 | 1:190127911 | D | 0.113 | Intron | 0.20 ± 0.045 | 9.02 × 10−6 | −0.56 ± 0.47 | 0.226 | 1.27 (0.90–1.80) | 0.174 |
| 15 | rs3773884 | MME | 3:154859650 | G | 0.052 | Intron | 0.28 ± 0.063 | 9.33 × 10−6 | −0.52 ± 0.64 | 0.418 | 1.28 (0.81–2.01) | 0.291 |
SNP, single-nucleotide polymorphism; MAF, minor allele frequency; IDO, indoleamine 2,3-dioxygenase; CKD, chronic kidney disease; eGFR, estimated glomerular filtration rate; β, regression coefficient; S.E., standard error; OR, odds ratio; CI, confidence interval. All analyses were adjusted for age, area, gender, BMI, drinking, smoking, SBP, hs-CRP, and HbA1c. The cutoff p-value was p < 1 × 10−5 for IDO activity and p < 0.05 for eGFR and CKD. Genes stated in the manuscript are indicated in bold.
Table 3.
Rare variants associated with indoleamine 2,3-dioxygenase activity in Koreans.
| No. | SNP | Nearest Gene | Chromosome Position | Minor Allele | MAF | HWE p-Value | Function | IDO Activity | eGFR | CKD | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β ± S.E | p-Value | β ± S.E | p-Value | OR (95% CI) | p-Value | ||||||||
| 1 | rs182145739 | LOC105377444 | 4:138651320 | T | 0.011 | 1 | Intron | 0.82 ± 0.13 | 8.46 × 10−10 | −1.08 ± 1.36 | 0.427 | 2.63 (1.15–6.03) | 0.022 |
| 2 | rs149763281 | SLC24A2 | 9:20104936 | C | 0.011 | 0.277 | Intron | 0.78 ± 0.13 | 3.88 × 10−9 | −1.68 ± 1.36 | 0.218 | 0.95 (0.34–2.72) | 0.936 |
| 3 | rs117150322 | TNFRSF19 | 13:24120841 | A | 0.009 | 0.191 | - | 0.83 ± 0.15 | 9.89 × 10−9 | −3.22 ± 1.49 | 0.031 | 1.01 (0.30–3.38) | 0.994 |
| 4 | rs188289326 | CACNA2D3 | 3:54867936 | A | 0.010 | 1 | Intron | 0.81 ± 0.14 | 1.02 × 10−8 | −2.34 ± 1.45 | 0.107 | 1.20 (0.49–2.96) | 0.690 |
| 5 | rs146321869 | LOC101928535 | 11:106108552 | G | 0.009 | 0.183 | - | 0.83 ± 0.15 | 1.16 × 10−8 | −3.34 ± 1.51 | 0.026 | 1.88 (0.67–5.26) | 0.232 |
| 6 | rs337828 | ARSB | 5:78196735 | G | 0.009 | 1 | Intron | 0.81 ± 0.15 | 3.55 × 10−8 | −1.53 ± 1.51 | 0.310 | 1.50 (0.55–4.14) | 0.431 |
| 7 | rs58332670 | FSTL5 | 4:163207867 | C | 0.035 | 0.549 | - | 0.42 ± 0.08 | 3.58 × 10−8 | −2.24 ± 0.79 | 4.32 × 10−3 | 1.43 (0.80–2.57) | 0.231 |
SNP, single nucleotide polymorphism; MAF, minor allele frequency; HWE, Hardy-Weinberg equilibrium; IDO, indoleamine 2,3-dioxygenase; CKD, chronic kidney disease; eGFR, estimated glomerular filtration rate; β, regression coefficient; S.E, standard error; OR, odds ratio; CI, confidence interval. All analyses were adjusted for age, area, gender, BMI, drinking, smoking, SBP, hs-CRP, and HbA1c. Cutoff p-values were p < 5 × 10−8 for IDO activity and p < 0.05 for eGFR and CKD. Gene highlighted in the manuscript are indicated in bold.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, H.-R.; Jin, H.-S.; Eom, Y.-B. Metabolite Genome-Wide Association Study for Indoleamine 2,3-Dioxygenase Activity Associated with Chronic Kidney Disease. Genes 2021, 12, 1905. https://doi.org/10.3390/genes12121905
AMA Style
Kim H-R, Jin H-S, Eom Y-B. Metabolite Genome-Wide Association Study for Indoleamine 2,3-Dioxygenase Activity Associated with Chronic Kidney Disease. Genes. 2021; 12(12):1905. https://doi.org/10.3390/genes12121905
Chicago/Turabian StyleKim, Hye-Rim, Hyun-Seok Jin, and Yong-Bin Eom. 2021. "Metabolite Genome-Wide Association Study for Indoleamine 2,3-Dioxygenase Activity Associated with Chronic Kidney Disease" Genes 12, no. 12: 1905. https://doi.org/10.3390/genes12121905
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.