
The Persistence of Staphylococcus aureus in Pressure Ulcers: A Colonising Role

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Whole Genome Sequencing
2.3. In Vivo Caenorhabditis elegans Killing Model
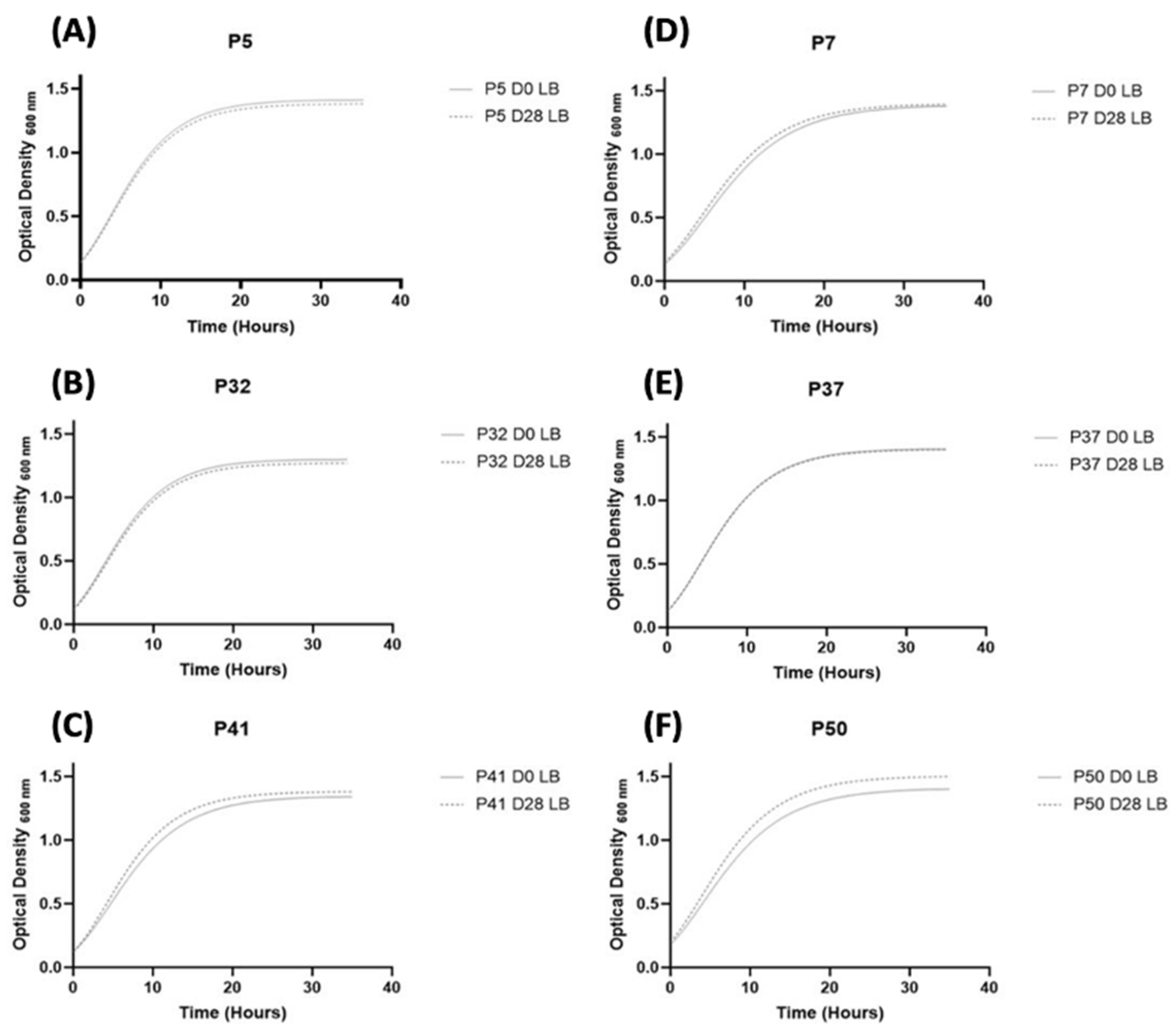
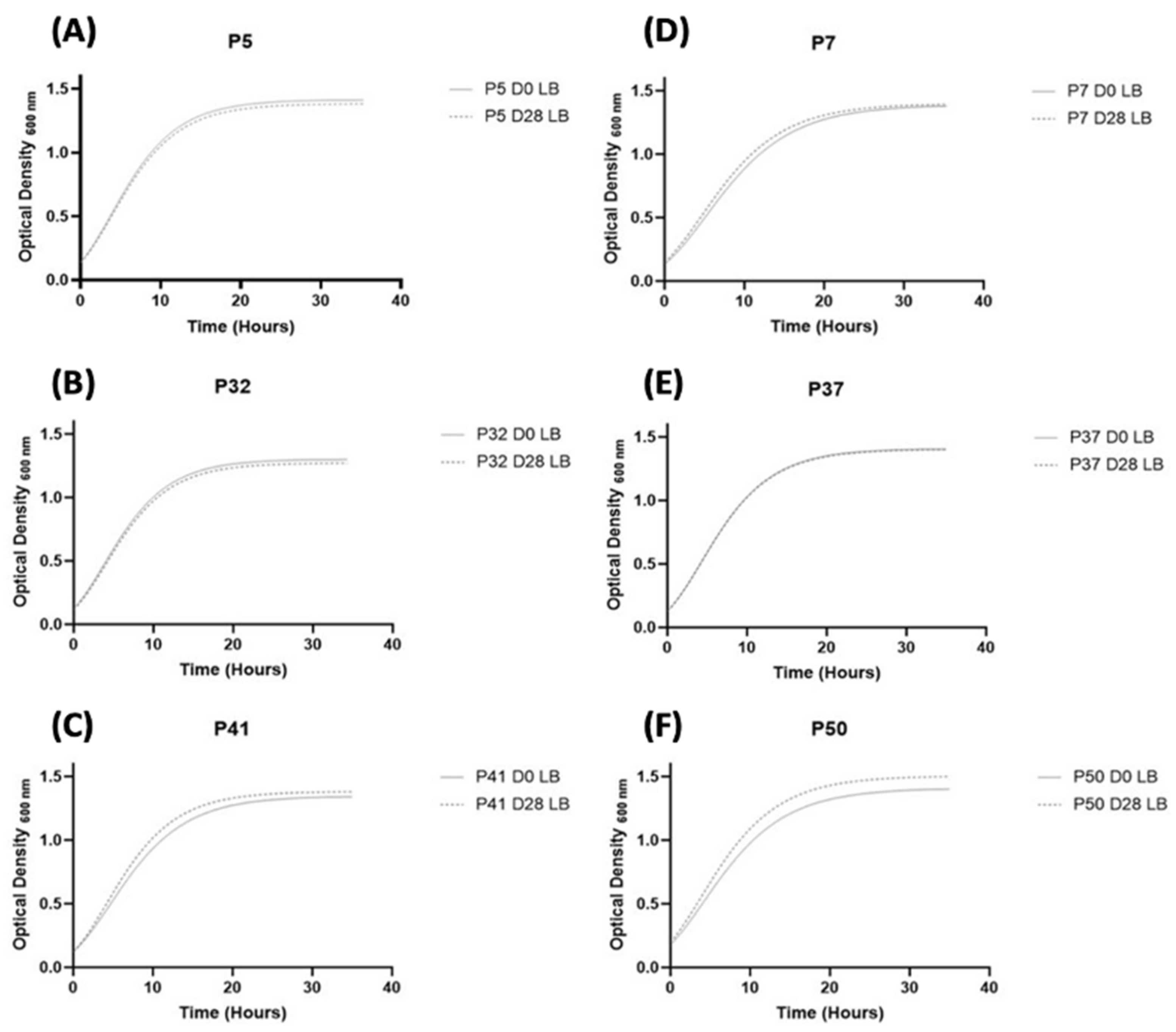
2.4. Growth Curves Evaluation
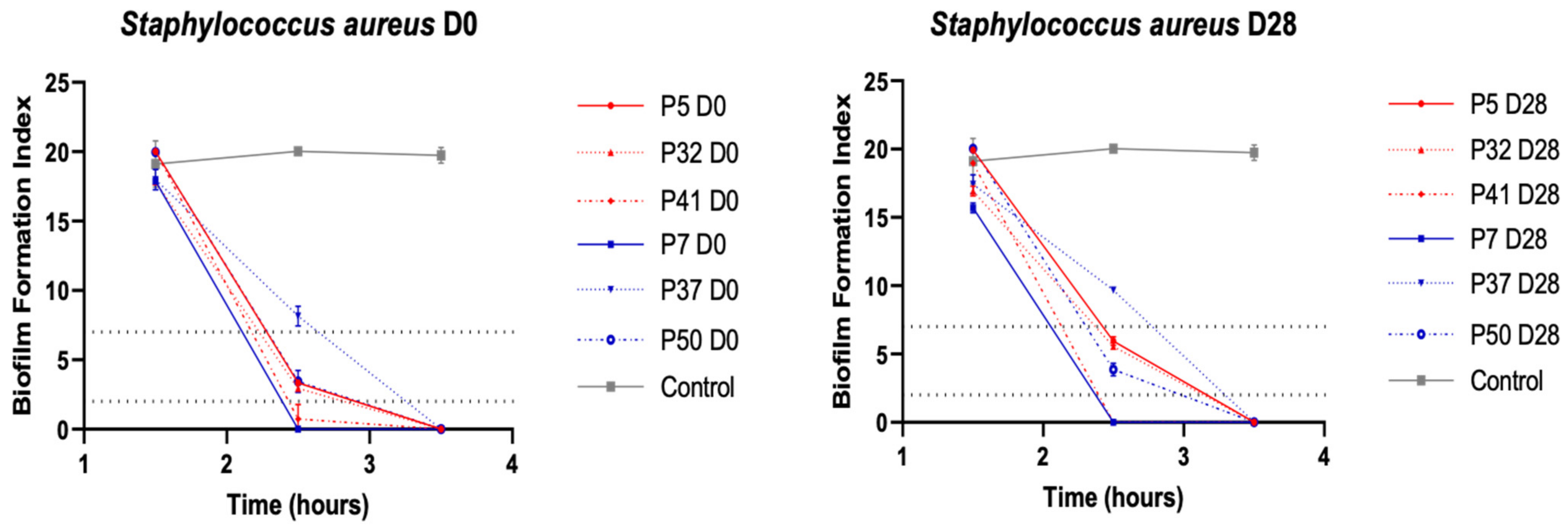
2.5. Determination of Early Biofilm Formation
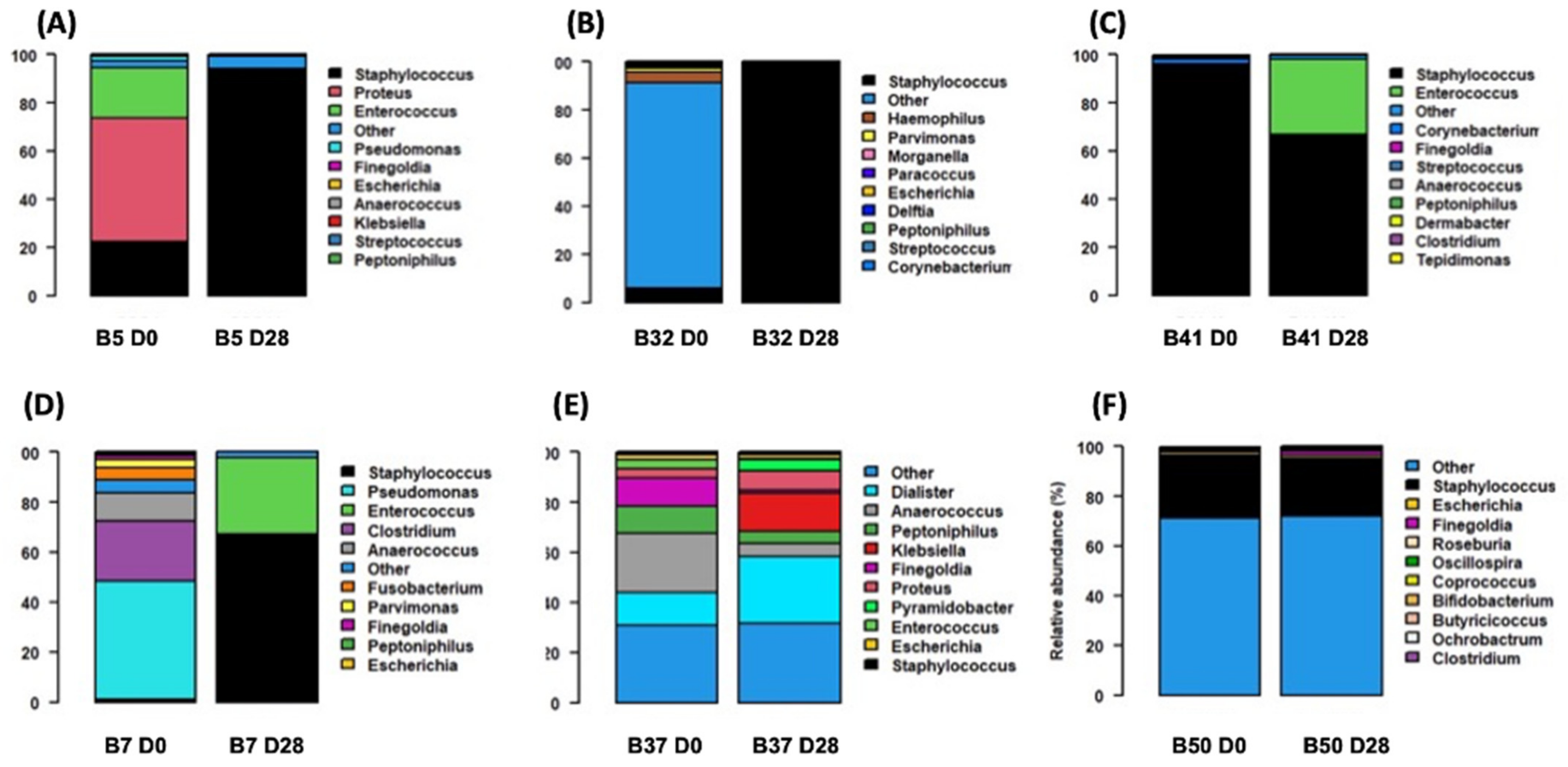
2.6. Description of Bacterial Communities Associated with S. aureus Using Metabarcoding Approach
2.7. Statistical Analysis
3. Results
3.1. Description of Patients
3.2. Genotypical Characteristics of S. aureus Strains
3.2.1. Genotyping of S. aureus Strains
3.2.2. Resistome and Virulome of S. aureus Strains
3.2.3. Evaluation of S. aureus Virulence Strains according to Clinical Evolution
3.3. Evolution of S. aureus Fitness and Ability to Form Biofilm in Accordance to Clinical Evolution
3.3.1. Fitness of S. aureus Strains
3.3.2. Ability to Form Biofilm in S. aureus Strains
3.4. S. aureus and Bacterial Interactions: Impact on Wound Evolution?
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whiteneck, G.G.; Charlifue, S.W.; Frankel, H.L.; Fraser, M.H.; Gardner, B.P.; Gerhart, K.A.; Krishnan, K.R.; Menter, R.R.; Nuseibeh, I.; Short, D.J. Mortality, Morbidity, and Psychosocial Outcomes of Persons Spinal Cord Injured More than 20 Years Ago. Paraplegia 1992, 30, 617–630. [Google Scholar] [CrossRef] [PubMed]
- National Pressure Ulcer Advisory Panel, European Pressure Ulcer Advisory Panel and Pan Pacific Pressure Injury Alliance. Prevention and Treatment of Pressure Ulcers: Quick Reference Guide; Haesler, E., Ed.; Cambridge Media: Osborne Park, Australia, 2014; pp. 1–75. [Google Scholar]
- Stroupe, K.; Manheim, L.; Evans, C.; Guihan, M.; Ho, C.; Li, K.; Cowper-Ripley, D.; Hogan, T.; St. Andre, J.; Huo, Z.; et al. Cost of Treating Pressure Ulcers for Veterans with Spinal Cord Injury. Top. Spinal Cord Inj. Rehabil. 2011, 16, 62–73. [Google Scholar] [CrossRef]
- Dowd, S.E.; Wolcott, R.D.; Sun, Y.; McKeehan, T.; Smith, E.; Rhoads, D. Polymicrobial Nature of Chronic Diabetic Foot Ulcer Biofilm Infections Determined Using Bacterial Tag Encoded FLX Amplicon Pyrosequencing (BTEFAP). PLoS ONE 2008, 3, e3326. [Google Scholar] [CrossRef] [PubMed]
- James, G.A.; Swogger, E.; Wolcott, R.; deLancey Pulcini, E.; Secor, P.; Sestrich, J.; Costerton, J.W.; Stewart, P.S. Biofilms in Chronic Wounds. Wound Repair Regen. 2008, 16, 37–44. [Google Scholar] [CrossRef]
- European Pressure Ulcer Advisory Panel; National Pressure Ulcer Advisory Panel and Pan Pacific Pressure Injury Alliance. Prevention and Treatment of Pressure Ulcers/Injuries: Clinical Practice Guideline: The International Guideline; Haesler, E., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 1–408. [Google Scholar]
- Thomson, P.D.; Smith, D.J. What Is Infection? Am. J. Surg. 1994, 167, S7–S11. [Google Scholar] [CrossRef]
- Dow, G.; Browne, A.; Sibbald, R.G. Infection in Chronic Wounds: Controversies in Diagnosis and Treatment. Ostomy Wound Manag. 1999, 45, 23–40. [Google Scholar]
- Sotto, A.; Lina, G.; Richard, J.-L.; Combescure, C.; Bourg, G.; Vidal, L.; Jourdan, N.; Etienne, J.; Lavigne, J.-P. Virulence Potential of Staphylococcus aureus Strains Isolated from Diabetic Foot Ulcers: A New Paradigm. Diabetes Care 2008, 31, 2318–2324. [Google Scholar] [CrossRef] [Green Version]
- Sotto, A.; Richard, J.-L.; Messad, N.; Molinari, N.; Jourdan, N.; Schuldiner, S.; Sultan, A.; Carrière, C.; Canivet, B.; Landraud, L.; et al. Distinguishing Colonization from Infection with Staphylococcus aureus in Diabetic Foot Ulcers with Miniaturized Oligonucleotide Arrays: A French Multicenter Study. Diabetes Care 2012, 35, 617–623. [Google Scholar] [CrossRef] [Green Version]
- Dana, A.N.; Bauman, W.A. Bacteriology of Pressure Ulcers in Individuals with Spinal Cord Injury: What We Know and What We Should Know. J. Spinal Cord Med. 2015, 38, 147–160. [Google Scholar] [CrossRef] [Green Version]
- Wolcott, R.D.; Hanson, J.D.; Rees, E.J.; Koenig, L.D.; Phillips, C.D.; Wolcott, R.A.; Cox, S.B.; White, J.S. Analysis of the Chronic Wound Microbiota of 2,963 Patients by 16S RDNA Pyrosequencing. Wound Repair Regen. 2016, 24, 163–174. [Google Scholar] [CrossRef]
- Dunyach-Remy, C.; Ngba Essebe, C.; Sotto, A.; Lavigne, J.-P. Staphylococcus aureus Toxins and Diabetic Foot Ulcers: Role in Pathogenesis and Interest in Diagnosis. Toxins 2016, 8, 209. [Google Scholar] [CrossRef] [Green Version]
- Messad, N.; Prajsnar, T.K.; Lina, G.; O’Callaghan, D.; Foster, S.J.; Renshaw, S.A.; Skaar, E.P.; Bes, M.; Dunyach-Remy, C.; Vandenesch, F.; et al. Existence of a Colonizing Staphylococcus aureus Strain Isolated in Diabetic Foot Ulcers. Diabetes 2015, 64, 2991–2995. [Google Scholar] [CrossRef] [Green Version]
- Josse, J.; Laurent, F.; Diot, A. Staphylococcal Adhesion and Host Cell Invasion: Fibronectin-Binding and Other Mechanisms. Front. Microbiol. 2017, 8, 2433. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Sun, Y.; Missiakas, D.; Schneewind, O. Staphylococcus aureus Decolonization of Mice with Monoclonal Antibody Neutralizing Protein A. J. Infect. Dis. 2019, 219, 884–888. [Google Scholar] [CrossRef]
- Missiakas, D.; Schneewind, O. Staphylococcus aureus Vaccines: Deviating from the Carol. J. Exp. Med. 2016, 213, 1645–1653. [Google Scholar] [CrossRef] [Green Version]
- Valeva, A.; Walev, I.; Pinkernell, M.; Walker, B.; Bayley, H.; Palmer, M.; Bhakdi, S. Transmembrane -Barrel of Staphylococcal -Toxin Forms in Sensitive but Not in Resistant Cells. Proc. Natl. Acad. Sci. USA 1997, 94, 11607–11611. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Braughton, K.R.; Kretschmer, D.; Bach, T.-H.L.; Queck, S.Y.; Li, M.; Kennedy, A.D.; Dorward, D.W.; Klebanoff, S.J.; Peschel, A.; et al. Identification of Novel Cytolytic Peptides as Key Virulence Determinants for Community-Associated MRSA. Nat. Med. 2007, 13, 1510–1514. [Google Scholar] [CrossRef]
- Lina, G.; Piemont, Y.; Godail-Gamot, F.; Bes, M.; Peter, M.-O.; Gauduchon, V.; Vandenesch, F.; Etienne, J. Involvement of Panton-Valentine Leukocidin--Producing Staphylococcus aureus in Primary Skin Infections and Pneumonia. Clin. Infect. Dis. 1999, 29, 1128–1132. [Google Scholar] [CrossRef]
- Dunyach-Remy, C.; Salipante, F.; Lavigne, J.-P.; Brunaud, M.; Demattei, C.; Yahiaoui-Martinez, A.; Bastide, S.; Palayer, C.; Sotto, A.; Gélis, A. Pressure Ulcers Microbiota Dynamics and Wound Evolution. Sci. Rep. 2021, 11, 18506. [Google Scholar] [CrossRef]
- EUCAST: Clinical Breakpoints and Dosing of Antibiotics. Available online: https://eucast.org/clinical_breakpoints/ (accessed on 26 September 2021).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Computat. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In Silico Detection and Typing of Plasmids Using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A Novel Web Tool for WGS-Based Detection of Antimicrobial Resistance Associated with Chromosomal Point Mutations in Bacterial Pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef] [Green Version]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Bartels, M.D.; Petersen, A.; Worning, P.; Nielsen, J.B.; Larner-Svensson, H.; Johansen, H.K.; Andersen, L.P.; Jarlov, J.O.; Boye, K.; Larsen, A.R.; et al. Comparing Whole-Genome Sequencing with Sanger Sequencing for Spa Typing of Methicillin-Resistant Staphylococcus aureus. J. Clin. Microbiol. 2014, 52, 4305–4308. [Google Scholar] [CrossRef] [Green Version]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid Large-Scale Prokaryote Pan Genome Analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Snippy: Fast Bacterial Variant Calling from NGS Reads. Available online: https://github.com/tseemann/snippy (accessed on 12 August 2021).
- Hadfield, J.; Croucher, N.J.; Goater, R.J.; Abudahab, K.; Aanensen, D.M.; Harris, S.R. Phandango: An Interactive Viewer for Bacterial Population Genomics. Bioinformatics 2018, 34, 292–293. [Google Scholar] [CrossRef]
- Ngba Essebe, C.; Visvikis, O.; Fines-Guyon, M.; Vergne, A.; Cattoir, V.; Lecoustumier, A.; Lemichez, E.; Sotto, A.; Lavigne, J.-P.; Dunyach-Remy, C. Decrease of Staphylococcus aureus Virulence by Helcococcus kunzii in a Caenorhabditis elegans Model. Front. Cell. Infect. Microbiol. 2017, 7, 77. [Google Scholar] [CrossRef] [Green Version]
- Lavigne, J.-P.; Nicolas-Chanoine, M.-H.; Bourg, G.; Moreau, J.; Sotto, A. Virulent Synergistic Effect between Enterococcus faecalis and Escherichia coli Assayed by Using the Caenorhabditis elegans Model. PLoS ONE 2008, 3, e3370. [Google Scholar] [CrossRef] [Green Version]
- Tjørve, K.M.C.; Tjørve, E. The Use of Gompertz Models in Growth Analyses, and New Gompertz-Model Approach: An Addition to the Unified-Richards Family. PLoS ONE 2017, 12, e0178691. [Google Scholar] [CrossRef] [PubMed]
- Peleg, M.; Corradini, M.G. Microbial Growth Curves: What the Models Tell Us and What They Cannot. Crit. Rev. Food Sci. Nutr. 2011, 51, 917–945. [Google Scholar] [CrossRef] [PubMed]
- Chavant, P.; Gaillard-Martinie, B.; Talon, R.; Hébraud, M.; Bernardi, T. A New Device for Rapid Evaluation of Biofilm Formation Potential by Bacteria. J. Microbiol. Methods 2007, 68, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Ranfaing, J.; Dunyach-Remy, C.; Lavigne, J.-P.; Sotto, A. Propolis Potentiates the Effect of Cranberry (Vaccinium macrocarpon) in Reducing the Motility and the Biofilm Formation of Uropathogenic Escherichia coli. PLoS ONE 2018, 13, e0202609. [Google Scholar] [CrossRef] [PubMed]
- Magoc, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Ankrum, A.; Hall, B.G. Population Dynamics of Staphylococcus aureus in Cystic Fibrosis Patients to Determine Transmission Events by Use of Whole-Genome Sequencing. J. Clin. Microbiol. 2017, 55, 2143–2152. [Google Scholar] [CrossRef] [Green Version]
- Lavigne, J.-P.; Hosny, M.; Dunyach-Remy, C.; Boutet-Dubois, A.; Schuldiner, S.; Cellier, N.; Yahiaoui-Martinez, A.; Molle, V.; La Scola, B.; Marchandin, H.; et al. Long-Term Intrahost Evolution of Staphylococcus aureus Among Diabetic Patients with Foot Infections. Front. Microbiol. 2021, 12, 741406. [Google Scholar] [CrossRef]
- Kahl, B.C.; Duebbers, A.; Lubritz, G.; Haeberle, J.; Koch, H.G.; Ritzerfeld, B.; Reilly, M.; Harms, E.; Proctor, R.A.; Herrmann, M.; et al. Population Dynamics of Persistent Staphylococcus aureus Isolated from the Airways of Cystic Fibrosis Patients during a 6-Year Prospective Study. J. Clin. Microbiol. 2003, 41, 4424–4427. [Google Scholar] [CrossRef] [Green Version]
- Lienard, A.; Hosny, M.; Jneid, J.; Schuldiner, S.; Cellier, N.; Sotto, A.; La Scola, B.; Lavigne, J.-P.; Pantel, A. Escherichia coli Isolated from Diabetic Foot Osteomyelitis: Clonal Diversity, Resistance Profile, Virulence Potential, and Genome Adaptation. Microorganisms 2021, 9, 380. [Google Scholar] [CrossRef]
- Uhlemann, A.-C.; Kennedy, A.D.; Martens, C.; Porcella, S.F.; Deleo, F.R.; Lowy, F.D. Toward an Understanding of the Evolution of Staphylococcus aureus Strain USA300 during Colonization in Community Households. Genome Biol. Evol. 2012, 4, 1275–1285. [Google Scholar] [CrossRef] [Green Version]
- Senneville, E.; Brière, M.; Neut, C.; Messad, N.; Lina, G.; Richard, J.-L.; Sotto, A.; Lavigne, J.-P. First Report of the Predominance of Clonal Complex 398 Staphylococcus aureus Strains in Osteomyelitis Complicating Diabetic Foot Ulcers: A National French Study. Clin. Microbiol. Infect. 2014, 20, O274–O277. [Google Scholar] [CrossRef] [Green Version]
- Bouiller, K.; Bertrand, X.; Hocquet, D.; Chirouze, C. Human Infection of Methicillin-Susceptible Staphylococcus aureus CC398: A Review. Microorganisms 2020, 8, 1737. [Google Scholar] [CrossRef]
- Peskine, A.; Laurent-Vannier, A.; Ackermann, C.N.; Anastassiades, C. Multiresistant bacteria colonisation in rehabilitation units. Ann. Readapt. Med. Phys. 2006, 49, 113–118. [Google Scholar] [CrossRef]
- Pouget, C.; Gustave, C.-A.; Ngba-Essebe, C.; Laurent, F.; Lemichez, E.; Tristan, A.; Sotto, A.; Dunyach-Rémy, C.; Lavigne, J.-P. Adaptation of Staphylococcus aureus in a Medium Mimicking a Diabetic Foot Environment. Toxins 2021, 13, 230. [Google Scholar] [CrossRef]
- Ramsey, M.M.; Freire, M.O.; Gabrilska, R.A.; Rumbaugh, K.P.; Lemon, K.P. Staphylococcus aureus shifts toward commensalism in response to Corynebacterium species. Front. Microbiol. 2016, 7, 1230. [Google Scholar] [CrossRef] [Green Version]
- Pouget, C.; Dunyach-Remy, C.; Pantel, A.; Schuldiner, S.; Sotto, A.; Lavigne, J.P. Biofilms in diabetic foot ulcers: Significance and clinical relevance. Microorganisms 2020, 8, 1580. [Google Scholar] [CrossRef]
- Iwase, T.; Uehara, Y.; Shinji, H.; Tajima, A.; Seo, H.; Takada, K.; Agata, T.; Mizunoe, Y. Staphylococcus epidermidis Esp inhibits Staphylococcus aureus biofilm formation and nasal colonization. Nature 2010, 465, 346–349. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Variables | Pressure Ulcers Evolution during 28 Days | ||||||
|---|---|---|---|---|---|---|---|
| Improvement (n = 3) | Worsening (n = 3) | ||||||
| Patient ID | P5 | P32 | P41 | P7 | P37 | P50 | |
| Sex | F | M | M | M | M | F | |
| Age | Years | 76 | 42 | 61 | 52 | 66 | 62 |
| Wound stage * (at D0) | III | III | III | III | III | III | |
| Wound stage (at D28) | III | III | III | III | III | III | |
| Wound localisation | Ischial/Sacral | Sacral | Ischial | Sacral | Sacral | Ischial | Ischial |
| CRP (D0) | mg/L | 26 | 22 | 11 | 9 | 154 | 5 |
| CRP (D28) | 23 | 34 | 10 | 22 | 69 | 9 | |
| Number of antibiotics used during wound management | 1 | 0 | 0 | 0 | 1 | 0 | |
| Wounds Evolution | Strains | Sequence Type Susceptibility to Methicillin 1 | SpA Type | Difference in SNP 2 Numbers (D0/D28) |
|---|---|---|---|---|
| Improvement | P5 D0 | 398—MSSA | t3625 | 6 |
| P5 D28 | ||||
| Improvement | P32 D0 | 398—MSSA | t571 | 43 |
| P32 D28 | ||||
| Improvement | P41 D0 | 8—MSSA | t008 | 12 |
| P41 D28 | ||||
| Worsening | P7 D0 | 8—MRSA | t008 | 32 |
| P7 D28 | ||||
| Worsening | P37 D0 | 5—MRSA | t777 | 8 |
| P37 D0 | ||||
| Worsening | P50 D0 | 5—MSSA | t002 | 82 |
| P50 D28 |
| Funtions | Genes | PU with Clinical Improvement 3 | PU with Worsening Evolution 3 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P5 D0 | P5 D28 | P32 D0 | P32 D28 | P41 D0 | P41 D28 | P7 D0 | P7 D28 | P37 D0 | P37 D28 | P50 D0 | P50 D28 | ||
| Adhesion/Colonisation | fnbpA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Biofilm Formation Regulatory System | icaABCDR 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| agrA, agrB sarA | 1 1 | 1 1 | 1 1 | 1 1 | 1 1 | 1 1 | 1 1 | 1 1 | 1 1 | 1 1 | 1 1 | 1 1 | |
| Pore Forming Toxins | lukDE | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| hlgA, hlgB, hlgC | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| hly | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| psm | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| lukF-PV, lukS-PV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Toxins that induce Lymphocyte T activation | sea and sel-X | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| sed, sej, ser | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | |
| seg, sel-i, sel-m, sel-n, sel-o, sel-u, sel-p | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | |
| tsst-1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Avoid Host Immune Response | clfB | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| coA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| clfA | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | |
| spA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| Protease activity | aur | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| etA, etB, etD | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| splA, splB, splE | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| Other | sak | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 |
| Resistance to β-lactams (MRSA 2) | mecA | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 |
| Resistance to quinolones | glrA and/or gyrA mutations | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 |
| Resistance to macrolides and related | ermA | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Resistance to fusidic acid | fusA (p.L461S) | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Strains | Clinical Evolution | LT50 (Days) | p (OP 50/Px) | p (NSA 739/Px) | p (D0/D28) |
|---|---|---|---|---|---|
| OP50 1 | Controls | 5.3 (±0.6) | NA | <0.001 | NA |
| NSA739 2 | 3.0 (±0.2) | <0.001 | NA | NA | |
| P5 D0 | Improvement | 4.0 (±0.1) | <0.001 | <0.01 | <0.001 |
| P5 D28 | 4.7 (±0.4) | <0.001 | <0.001 | ||
| P32 D0 | Improvement | 3.3 (±0.3) | <0.001 | 0.301 (NS) | 0.003 |
| P32 D28 | 3.7 (±0.6) | <0.001 | <0.001 | ||
| P41 D0 | Improvement | 4.0 (±0.3) | <0.001 | <0.001 | <0.001 |
| P41 D28 | 4.8 (±0.7) | <0.001 | <0.001 | ||
| P7 D0 | Worsening | 3.7 (±0.7) | <0.001 | <0.001 | <0.001 |
| P7 D28 | 4.3 (±0.5) | <0.001 | <0.001 | ||
| P37 D0 | Worsening | 4.0 (±0.1) | <0.001 | <0.001 | <0.001 |
| P37 D28 | 4.7 (±0.7) | 0.001 | <0.001 | ||
| P50 D0 | Worsening | 4.6 (±0.2) | <0.001 | <0.001 | 0.207 (NS) |
| P50 D28 | 5.0 (±0.1) | <0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fayolle, M.; Morsli, M.; Gelis, A.; Chateauraynaud, M.; Yahiaoui-Martinez, A.; Sotto, A.; Lavigne, J.-P.; Dunyach-Remy, C. The Persistence of Staphylococcus aureus in Pressure Ulcers: A Colonising Role. Genes 2021, 12, 1883. https://doi.org/10.3390/genes12121883
Fayolle M, Morsli M, Gelis A, Chateauraynaud M, Yahiaoui-Martinez A, Sotto A, Lavigne J-P, Dunyach-Remy C. The Persistence of Staphylococcus aureus in Pressure Ulcers: A Colonising Role. Genes. 2021; 12(12):1883. https://doi.org/10.3390/genes12121883
Chicago/Turabian StyleFayolle, Martin, Madjid Morsli, Anthony Gelis, Marion Chateauraynaud, Alex Yahiaoui-Martinez, Albert Sotto, Jean-Philippe Lavigne, and Catherine Dunyach-Remy. 2021. "The Persistence of Staphylococcus aureus in Pressure Ulcers: A Colonising Role" Genes 12, no. 12: 1883. https://doi.org/10.3390/genes12121883
APA StyleFayolle, M., Morsli, M., Gelis, A., Chateauraynaud, M., Yahiaoui-Martinez, A., Sotto, A., Lavigne, J.-P., & Dunyach-Remy, C. (2021). The Persistence of Staphylococcus aureus in Pressure Ulcers: A Colonising Role. Genes, 12(12), 1883. https://doi.org/10.3390/genes12121883