
Two Rare Variants in PLAU and BACE1 Genes—Do They Contribute to Semantic Dementia Clinical Phenotype?

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Whole-Genome Sequencing
2.2. WGS Data Preprocessing
2.3. Quality Assessment
2.4. Rare Variant Analysis
2.5. Copy Number Variation Analysis
2.6. Mutation Screening of the PGRN, MAPT and C9orf72 Genes
2.7. Fibroblast Cultures and Inhibition of Nonsense-Mediated Decay (NMD)
2.8. mRNA Expression Analysis—Real Time PCR
2.9. Western Blot
2.10. Statistical Analysis
3. Results
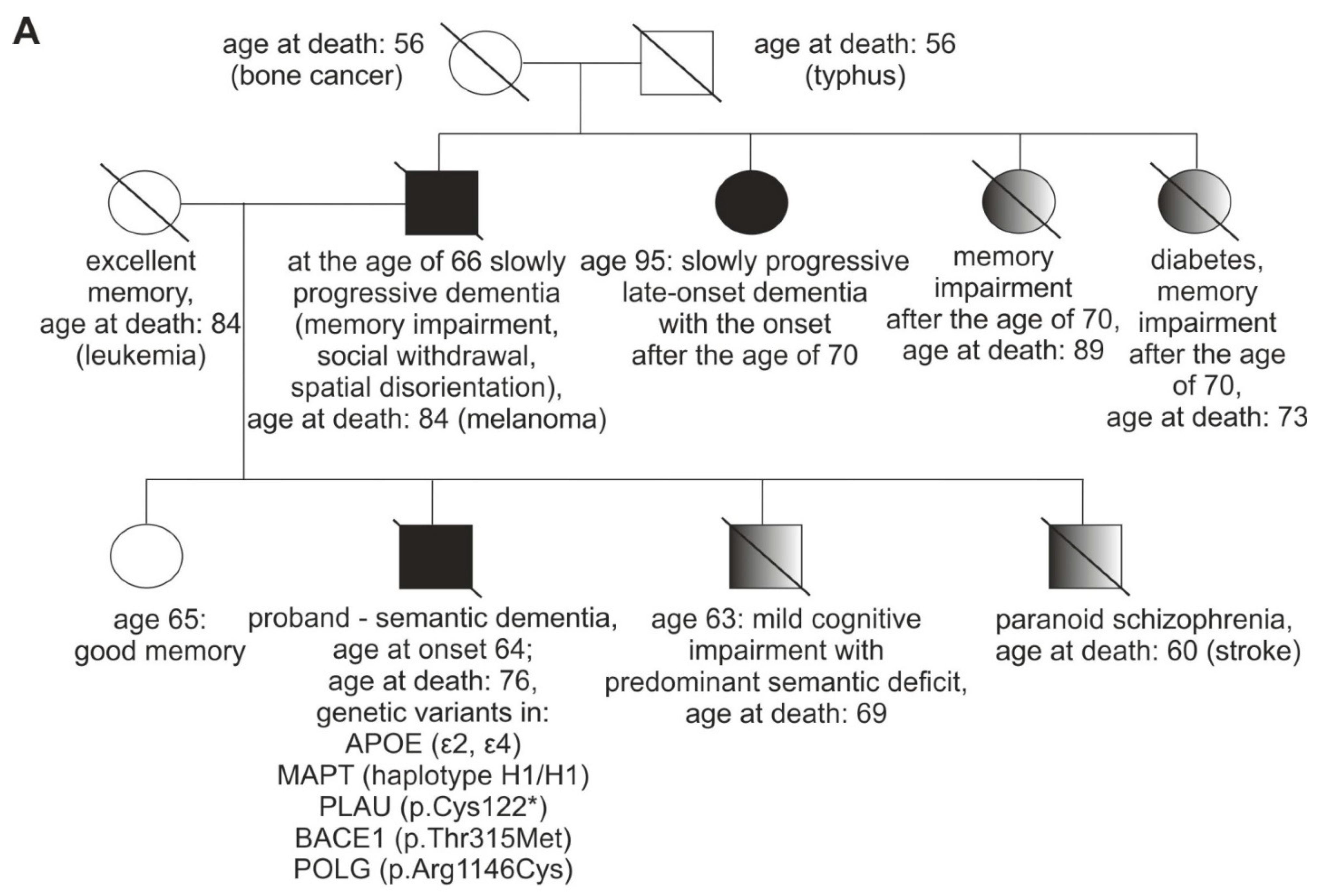
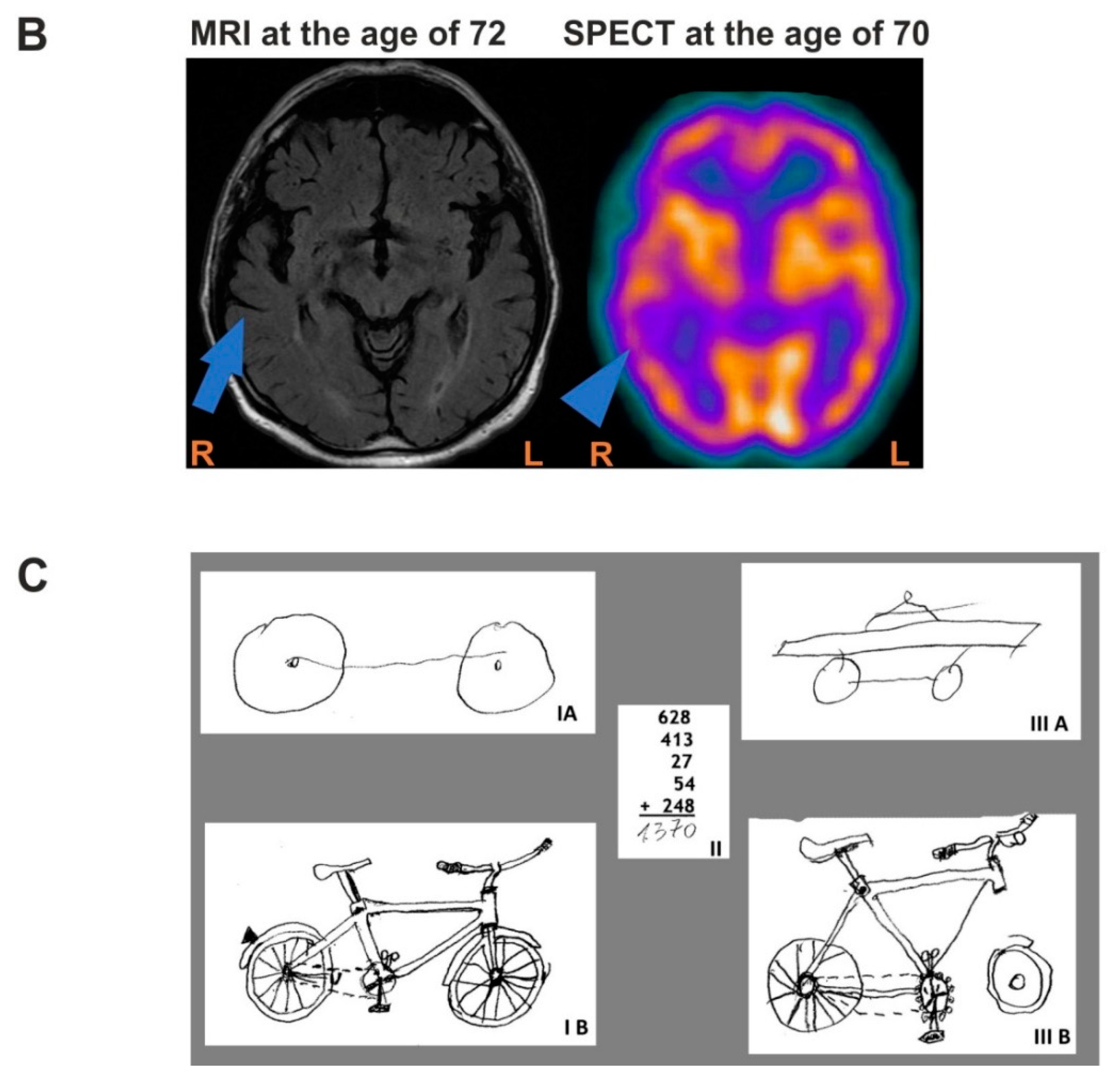
3.1. Case Description
3.2. Genetic Analyses
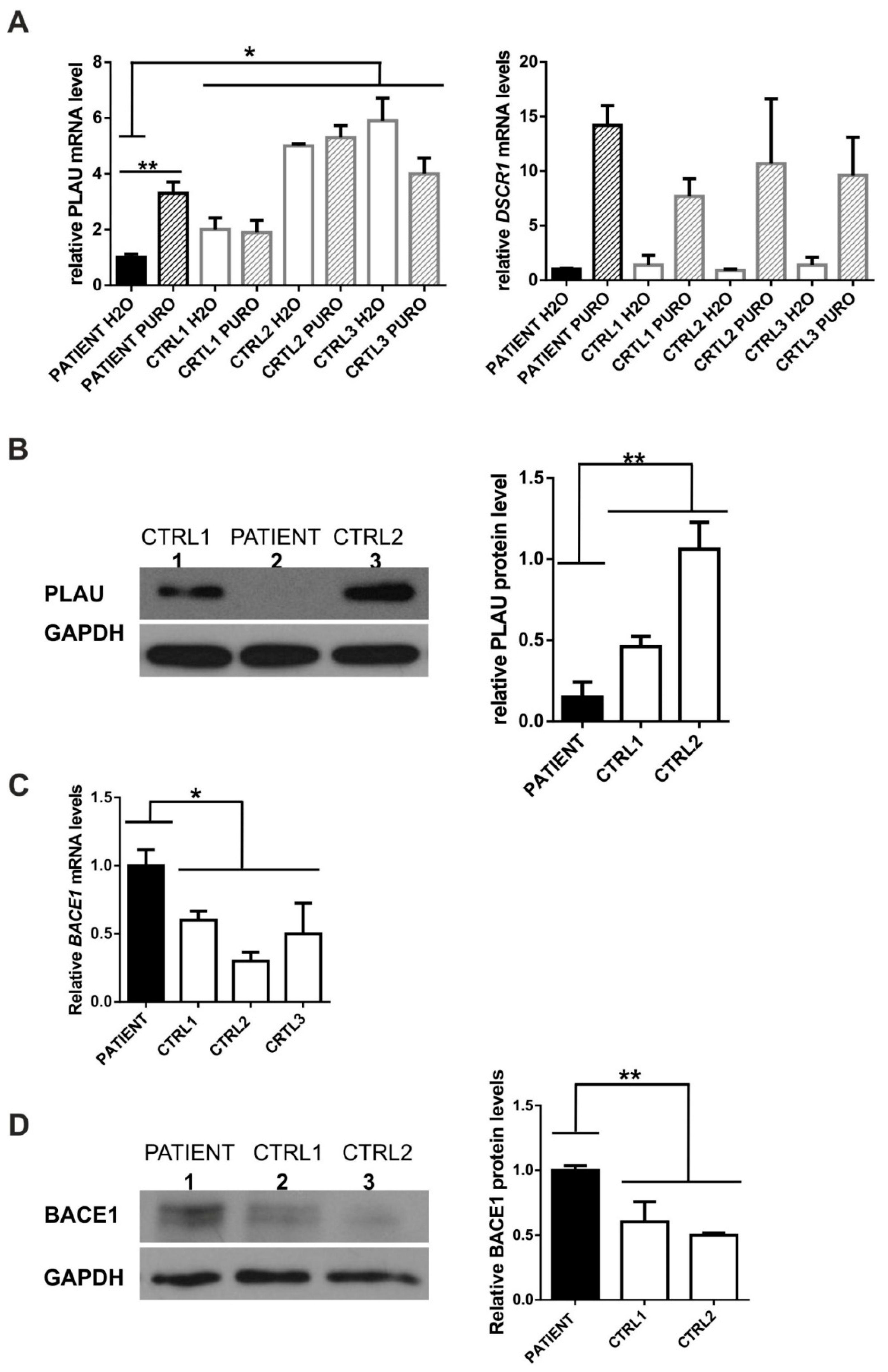
3.2.1. PLAU (Urokinase-Type Plasminogen Activator)
3.2.2. BACE1 (β-Site APP-Cleaving Enzyme 1, Beta-Secretase 1)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rabinovici, G.D.; Carrillo, M.C.; Forman, M.; DeSanti, S.; Miller, D.S.; Kozauer, N.; Petersen, R.C.; Randolph, C.; Knopman, D.S.; Smith, E.E.; et al. Multiple comorbid neuropathologies in the setting of Alzheimer’s disease neuropathology and implications for drug development. Alzheimer’s Dement 2017, 3, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Jellinger, K.A. Neuropathological aspects of Alzheimer disease, Parkinson disease and frontotemporal dementia. Neurodegener. Dis. 2008, 5, 118–121. [Google Scholar] [CrossRef]
- Cochran, J.N.; McKinley, E.C.; Cochran, M.; Amaral, M.D.; Moyers, B.A.; Lasseigne, B.N.; Gray, D.E.; Lawlor, J.M.J.; Prokop, J.W.; Geier, E.G.; et al. Genome sequencing for early-onset or atypical dementia: High diagnostic yield and frequent observation of multiple contributory alleles. Cold Spring Harb. Mol. Case Stud. 2019, 5, a003491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsanis, N. The continuum of causality in human genetic disorders. Genome Biol. 2016, 17, 233. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Layer, R.M.; Chiang, C.; Quinlan, A.R.; Hall, I.M. LUMPY: A probabilistic framework for structural variant discovery. Genome Biol. 2014, 15, R84. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, B.S.; Quinlan, A.R. Duphold: Scalable, depth-based annotation and curation of high-confidence structural variant calls. Gigascience 2019, 8, giz040. [Google Scholar] [CrossRef] [Green Version]
- DeJesus-Hernandez, M.; Mackenzie, I.R.; Boeve, B.F.; Boxer, A.L.; Baker, M.; Rutherford, N.J.; Nicholson, A.M.; Finch, N.A.; Flynn, H.; Adamson, J.; et al. Expanded GGGGCC hexanucleotide repeat in noncoding region of C9ORF72 causes chromosome 9p-linked FTD and ALS. Neuron 2011, 72, 245–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zekanowski, C.; Peplonska, B.; Styczynska, M.; Gustaw, K.; Kuznicki, J.; Barcikowska, M. Mutation screening of the MAPT and STH genes in Polish patients with clinically diagnosed frontotemporal dementia. Dement. Geriatr. Cogn. Disord. 2003, 16, 126–131. [Google Scholar] [CrossRef]
- Baker, M.; Mackenzie, I.R.; Pickering-Brown, S.M.; Gass, J.; Rademakers, R.; Lindholm, C.; Snowden, J.; Adamson, J.; Sadovnick, A.D.; Rollinson, S.; et al. Mutations in progranulin cause tau-negative frontotemporal dementia linked to chromosome 17. Nature 2006, 442, 916–919. [Google Scholar] [CrossRef]
- Gaweda-Walerych, K.; Walerych, D.; Berdynski, M.; Buratti, E.; Zekanowski, C. Parkin Levels Decrease in Fibroblasts With Progranulin (PGRN) Pathogenic Variants and in a Cellular Model of PGRN Deficiency. Front. Mol. Neurosci. 2021, 14, 676478. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Qu, Y.; Cao, Y.; Yang, L.; Ge, L.; Jin, Y.; Wang, H.; Song, F. The SMN1 common variant c.22 dupA in Chinese patients causes spinal muscular atrophy by nonsense-mediated mRNA decay in humans. Gene 2018, 644, 49–55. [Google Scholar] [CrossRef]
- Qu, Y.J.; Ge, L.; Bai, J.L.; Cao, Y.Y.; Jin, Y.W.; Wang, H.; Yang, L.; Song, F. p.Val19Glyfs*21 and p.Leu228* variants in the survival of motor neuron 1 trigger nonsense-mediated mRNA decay causing the SMN1 PTC+ transcripts degradation. Mutat. Res. 2017, 806, 31–38. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Sitek, N.; Brockhuis Sikorski, S. Otępienie semantyczne—Charakterystyka kliniczna ze szczególnym uwzględnieniem neuropsychologicznego profilu objawów [Semantic dementia–Clinical characteristics with special emphasis on neuropsychological pattern of symptoms]. Polski Przegląd Neurol. 2020, 16, 244–248. [Google Scholar]
- Harciarek, M.; Sitek, E.J.; Kertesz, A. The patterns of progression in primary progressive aphasia-implications for assessment and management. Aphasiology 2014, 28, 964–980. [Google Scholar] [CrossRef]
- Kumfor, F.; Landin-Romero, R.; Devenney, E.; Hutchings, R.; Grasso, R.; Hodges, J.R.; Piguet, O. On the right side? A longitudinal study of left- versus right-lateralized semantic dementia. Brain 2016, 139, 986–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greaves, C.V.; Rohrer, J.D. An update on genetic frontotemporal dementia. J. Neurol. 2019, 266, 2075–2086. [Google Scholar] [CrossRef] [Green Version]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Molinuevo, J.L.; Gramunt, N.; Gispert, J.D.; Fauria, K.; Esteller, M.; Minguillon, C.; Sanchez-Benavides, G.; Huesa, G.; Moran, S.; Dal-Re, R.; et al. The ALFA project: A research platform to identify early pathophysiological features of Alzheimer’s disease. Alzheimer’s Dement 2016, 2, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Hampel, H.; Vassar, R.; De Strooper, B.; Hardy, J.; Willem, M.; Singh, N.; Zhou, J.; Yan, R.; Vanmechelen, E.; De Vos, A.; et al. The beta-Secretase BACE1 in Alzheimer’s Disease. Biol. Psychiatry 2021, 89, 745–756. [Google Scholar] [CrossRef]
- Landin-Romero, R.; Tan, R.; Hodges, J.R.; Kumfor, F. An update on semantic dementia: Genetics, imaging, and pathology. Alzheimer’s Res. Ther. 2016, 8, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulugut Erkoyun, H.; Groot, C.; Heilbron, R.; Nelissen, A.; van Rossum, J.; Jutten, R.; Koene, T.; van der Flier, W.M.; Wattjes, M.P.; Scheltens, P.; et al. A clinical-radiological framework of the right temporal variant of frontotemporal dementia. Brain 2020, 143, 2831–2843. [Google Scholar] [CrossRef]
- Gorno-Tempini, M.L.; Hillis, A.E.; Weintraub, S.; Kertesz, A.; Mendez, M.; Cappa, S.F.; Ogar, J.M.; Rohrer, J.D.; Black, S.; Boeve, B.F.; et al. Classification of primary progressive aphasia and its variants. Neurology 2011, 76, 1006–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neary, D.; Snowden, J.S.; Gustafson, L.; Passant, U.; Stuss, D.; Black, S.; Freedman, M.; Kertesz, A.; Robert, P.H.; Albert, M.; et al. Frontotemporal lobar degeneration: A consensus on clinical diagnostic criteria. Neurology 1998, 51, 1546–1554. [Google Scholar] [CrossRef] [Green Version]
- Hodges, J.R.; Mitchell, J.; Dawson, K.; Spillantini, M.G.; Xuereb, J.H.; McMonagle, P.; Nestor, P.J.; Patterson, K. Semantic dementia: Demography, familial factors and survival in a consecutive series of 100 cases. Brain 2010, 133, 300–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulugut Erkoyun, H.; van der Lee, S.J.; Nijmeijer, B.; van Spaendonk, R.; Nelissen, A.; Scarioni, M.; Dijkstra, A.; Samanci, B.; Gurvit, H.; Yildirim, Z.; et al. The Right Temporal Variant of Frontotemporal Dementia Is Not Genetically Sporadic: A Case Series. J. Alzheimer’s Dis. 2021, 79, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Snowden, J.S.; Adams, J.; Harris, J.; Thompson, J.C.; Rollinson, S.; Richardson, A.; Jones, M.; Neary, D.; Mann, D.M.; Pickering-Brown, S. Distinct clinical and pathological phenotypes in frontotemporal dementia associated with MAPT, PGRN and C9orf72 mutations. Amyotroph. Lateral. Scler. Front. Degener. 2015, 16, 497–505. [Google Scholar] [CrossRef]
- Kyyriainen, J.; Bolkvadze, T.; Koivisto, H.; Lipponen, A.; Perez, L.O.; Ekolle Ndode-Ekane, X.; Tanila, H.; Pitkanen, A. Deficiency of urokinase-type plasminogen activator and its receptor affects social behavior and increases seizure susceptibility. Epilepsy Res. 2019, 151, 67–74. [Google Scholar] [CrossRef]
- Rantala, J.; Kemppainen, S.; Ndode-Ekane, X.E.; Lahtinen, L.; Bolkvadze, T.; Gurevicius, K.; Tanila, H.; Pitkanen, A. Urokinase-type plasminogen activator deficiency has little effect on seizure susceptibility and acquired epilepsy phenotype but reduces spontaneous exploration in mice. Epilepsy Behav. 2015, 42, 117–128. [Google Scholar] [CrossRef]
- Ertekin-Taner, N.; Ronald, J.; Feuk, L.; Prince, J.; Tucker, M.; Younkin, L.; Hella, M.; Jain, S.; Hackett, A.; Scanlin, L.; et al. Elevated amyloid beta protein (Abeta42) and late onset Alzheimer’s disease are associated with single nucleotide polymorphisms in the urokinase-type plasminogen activator gene. Hum. Mol. Genet. 2005, 14, 447–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juncker-Jensen, A.; Lund, L.R. Phenotypic overlap between MMP-13 and the plasminogen activation system during wound healing in mice. PLoS ONE 2011, 6, e16954. [Google Scholar]
- Bennett, R.E.; Robbins, A.B.; Hu, M.; Cao, X.; Betensky, R.A.; Clark, T.; Das, S.; Hyman, B.T. Tau induces blood vessel abnormalities and angiogenesis-related gene expression in P301L transgenic mice and human Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2018, 115, E1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riemenschneider, M.; Konta, L.; Friedrich, P.; Schwarz, S.; Taddei, K.; Neff, F.; Padovani, A.; Kolsch, H.; Laws, S.M.; Klopp, N.; et al. A functional polymorphism within plasminogen activator urokinase (PLAU) is associated with Alzheimer’s disease. Hum. Mol. Genet. 2006, 15, 2446–2456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papassotiropoulos, A.; Tsolaki, M.; Wollmer, M.A.; Molyva, D.; Thal, D.R.; Huynh, K.D.; Tracy, J.; Staehelin, H.B.; Monsch, A.U.; Nitsch, R.M.; et al. No association of a non-synonymous PLAU polymorphism with Alzheimer’s disease and disease-related traits. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2005, 132B, 21–23. [Google Scholar] [CrossRef] [PubMed]
- Bagnoli, S.; Tedde, A.; Cellini, E.; Rotondi, M.; Nacmias, B.; Sorbi, S. The urokinase-plasminogen activator (PLAU) gene is not associated with late onset Alzheimer’s disease. Neurogenetics 2005, 6, 53–54. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, M.; Ushiyama, Y.; Sakai, M.; Tamaki, S.; Hara, H.; Takahashi, K.; Sawasaki, Y.; Hanada, K. Characterization of single chain urokinase-type plasminogen activator with a novel amino-acid substitution in the kringle structure. Biochim. Biophys. Acta 1996, 1293, 83–89. [Google Scholar] [CrossRef]
- Vilarino-Guell, C.; Zimprich, A.; Martinelli-Boneschi, F.; Herculano, B.; Wang, Z.; Matesanz, F.; Urcelay, E.; Vandenbroeck, K.; Leyva, L.; Gris, D.; et al. Exome sequencing in multiple sclerosis families identifies 12 candidate genes and nominates biological pathways for the genesis of disease. PLoS Genet. 2019, 15, e1008180. [Google Scholar] [CrossRef]
- Cardoso, A.L.; Fernandes, A.; Aguilar-Pimentel, J.A.; de Angelis, M.H.; Guedes, J.R.; Brito, M.A.; Ortolano, S.; Pani, G.; Athanasopoulou, S.; Gonos, E.S.; et al. Towards frailty biomarkers: Candidates from genes and pathways regulated in aging and age-related diseases. Ageing Res. Rev. 2018, 47, 214–277. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.E.; Graff-Guerrero, A.; Houle, S.; Mizrahi, R.; Wilson, A.A.; Pollock, B.G.; Mulsant, B.H.; Felsky, D.; Voineskos, A.N.; Tang-Wai, D.F.; et al. Amyloid deposition in semantic dementia: A positron emission tomography study. Int. J. Geriatr. Psychiatry 2016, 31, 1064–1074. [Google Scholar] [CrossRef] [Green Version]
- Ulugut, H.; Dijkstra, A.A.; Scarioni, M.; Barkhof, F.; Scheltens, P.; Rozemuller, A.J.M.; Pijnenburg, Y.A.L. Right temporal variant frontotemporal dementia is pathologically heterogeneous: A case-series and a systematic review. Acta Neuropathol. Commun. 2021, 9, 131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Huang, J.; Shen, Y.; Li, R. BACE1-Dependent Neuregulin-1 Signaling: An Implication for Schizophrenia. Front. Mol. Neurosci. 2017, 10, 302. [Google Scholar] [CrossRef]
- Rossi, M.; Medina Escobar, A.; Radrizzani, M.; Tenembaum, S.; Perandones, C.; Merello, M. Dystonia in a Patient with Autosomal-Dominant Progressive External Ophthalmoplegia Type 1 Caused by Mutation in the POLG Gene. Mov. Disord. Clin. Pract. 2017, 4, 266–269. [Google Scholar] [CrossRef] [Green Version]
- Melberg, A.; Nennesmo, I.; Moslemi, A.R.; Kollberg, G.; Luoma, P.; Suomalainen, A.; Holme, E.; Oldfors, A. Alzheimer pathology associated with POLG1 mutation, multiple mtDNA deletions, and APOE4/4: Premature ageing or just coincidence? Acta Neuropathol. 2005, 110, 315–316. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.L.; Lee, E.B.; Xie, S.X.; Rennert, L.; Suh, E.; Bredenberg, C.; Caswell, C.; Van Deerlin, V.M.; Yan, N.; Yousef, A.; et al. Neurodegenerative disease concomitant proteinopathies are prevalent, age-related and APOE4-associated. Brain 2018, 141, 2181–2193. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Juan, P.; Moreno, S.; de Rojas, I.; Hernandez, I.; Valero, S.; Alegret, M.; Montrreal, L.; Garcia Gonzalez, P.; Lage, C.; Lopez-Garcia, S.; et al. The MAPT H1 Haplotype Is a Risk Factor for Alzheimer’s Disease in APOE epsilon4 Non-carriers. Front. Aging Neurosci. 2019, 11, 327. [Google Scholar] [CrossRef] [Green Version]
- Niemiec, E.; Vears, D.F.; Borry, P.; Howard, H.C. Readability of informed consent forms for whole-exome and whole-genome sequencing. J. Community Genet. 2018, 9, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Gene | HGVS 1 DNA/Protein | Predicted Effect | MAF gnomAD | CADD 2 | |
|---|---|---|---|---|---|
| Rare variants | PLAU | heterozygous c.366C>A/p.Cys122stop | mRNA nonsense mediated decay, haploinsuficiency | - | 38 |
| BACE1 | heterozygous c.944C>T/p.Thr315Met | missense (splicing variant) | 0.00002389 | 29.5 | |
| POLG | heterozygous c.3436C>T/p.Arg1146Cys | missense | 0.00018695 | 35 | |
| Common variants | APOE | ε2/ε4 | - | - | - |
| MAPT | H1/H1 | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaweda-Walerych, K.; Sitek, E.J.; Borczyk, M.; Berdyński, M.; Narożańska, E.; Brockhuis, B.; Korostyński, M.; Sławek, J.; Zekanowski, C. Two Rare Variants in PLAU and BACE1 Genes—Do They Contribute to Semantic Dementia Clinical Phenotype? Genes 2021, 12, 1806. https://doi.org/10.3390/genes12111806
Gaweda-Walerych K, Sitek EJ, Borczyk M, Berdyński M, Narożańska E, Brockhuis B, Korostyński M, Sławek J, Zekanowski C. Two Rare Variants in PLAU and BACE1 Genes—Do They Contribute to Semantic Dementia Clinical Phenotype? Genes. 2021; 12(11):1806. https://doi.org/10.3390/genes12111806
Chicago/Turabian StyleGaweda-Walerych, Katarzyna, Emilia J. Sitek, Małgorzata Borczyk, Mariusz Berdyński, Ewa Narożańska, Bogna Brockhuis, Michał Korostyński, Jarosław Sławek, and Cezary Zekanowski. 2021. "Two Rare Variants in PLAU and BACE1 Genes—Do They Contribute to Semantic Dementia Clinical Phenotype?" Genes 12, no. 11: 1806. https://doi.org/10.3390/genes12111806
APA StyleGaweda-Walerych, K., Sitek, E. J., Borczyk, M., Berdyński, M., Narożańska, E., Brockhuis, B., Korostyński, M., Sławek, J., & Zekanowski, C. (2021). Two Rare Variants in PLAU and BACE1 Genes—Do They Contribute to Semantic Dementia Clinical Phenotype? Genes, 12(11), 1806. https://doi.org/10.3390/genes12111806