
Impact of Bicarbonate-β-Lactam Exposures on Methicillin-Resistant Staphylococcus aureus (MRSA) Gene Expression in Bicarbonate-β-Lactam-Responsive vs. Non-Responsive Strains

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. RNA Isolation, Library Construction, Sequencing and Analysis
2.3. qRT-PCR Validation of Select Genes
3. Results and Discussion
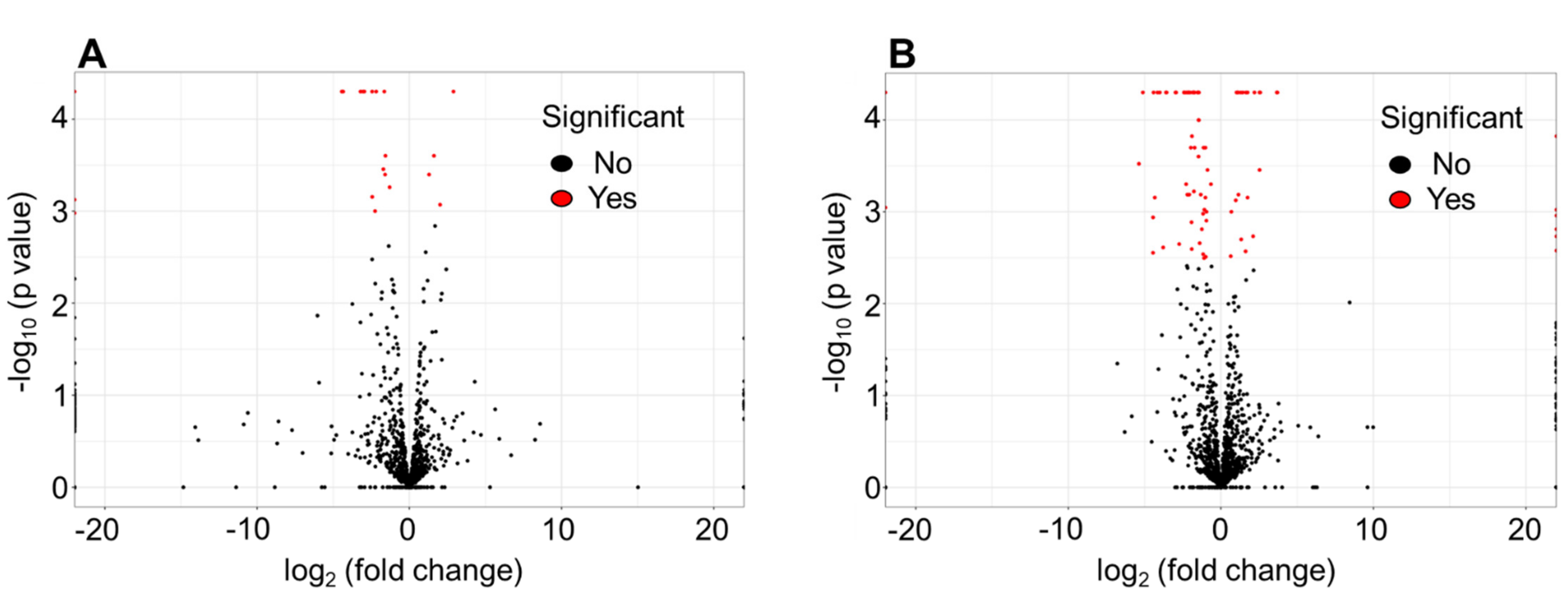
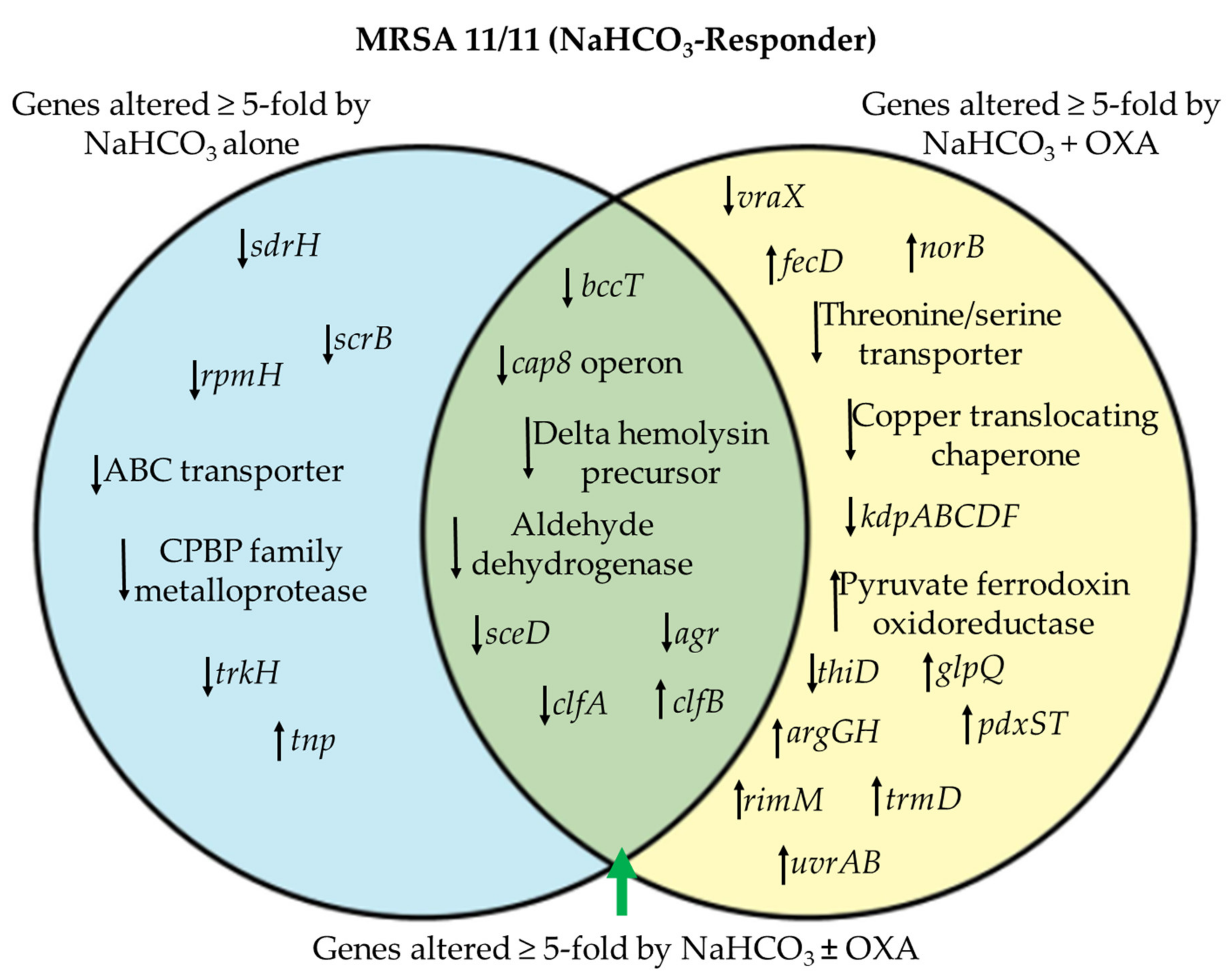
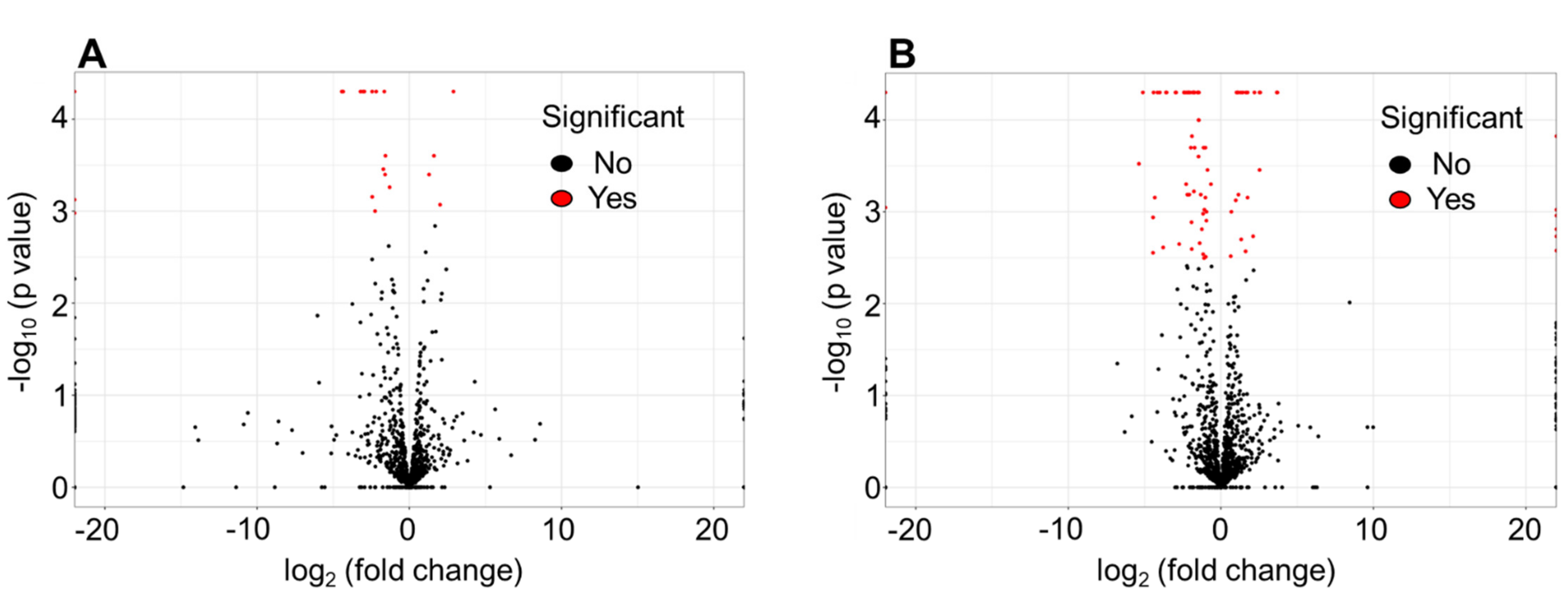
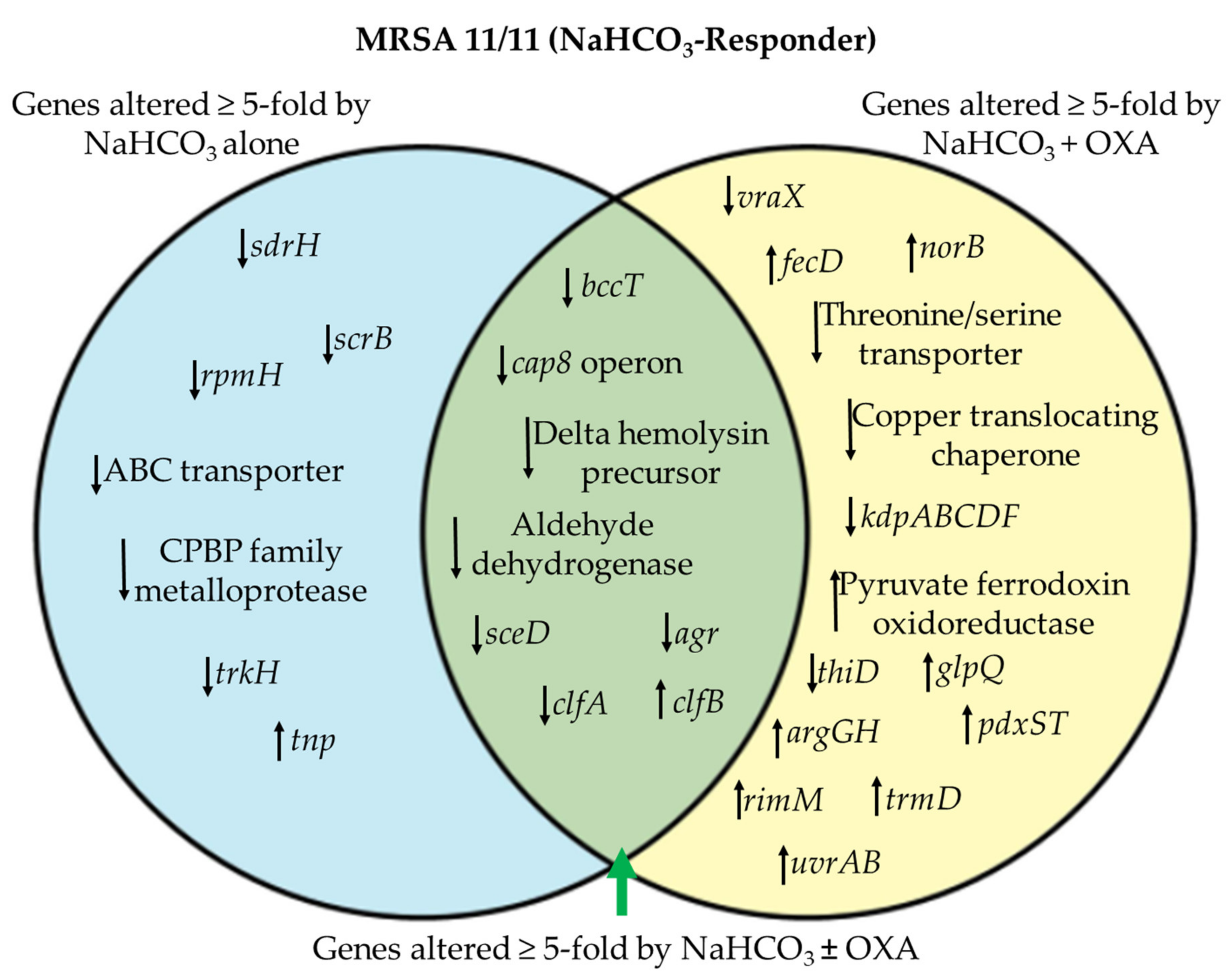
3.1. Differentially Expressed Genes by RNA-seq in NaHCO3-responsive vs. Non-responsive Strains
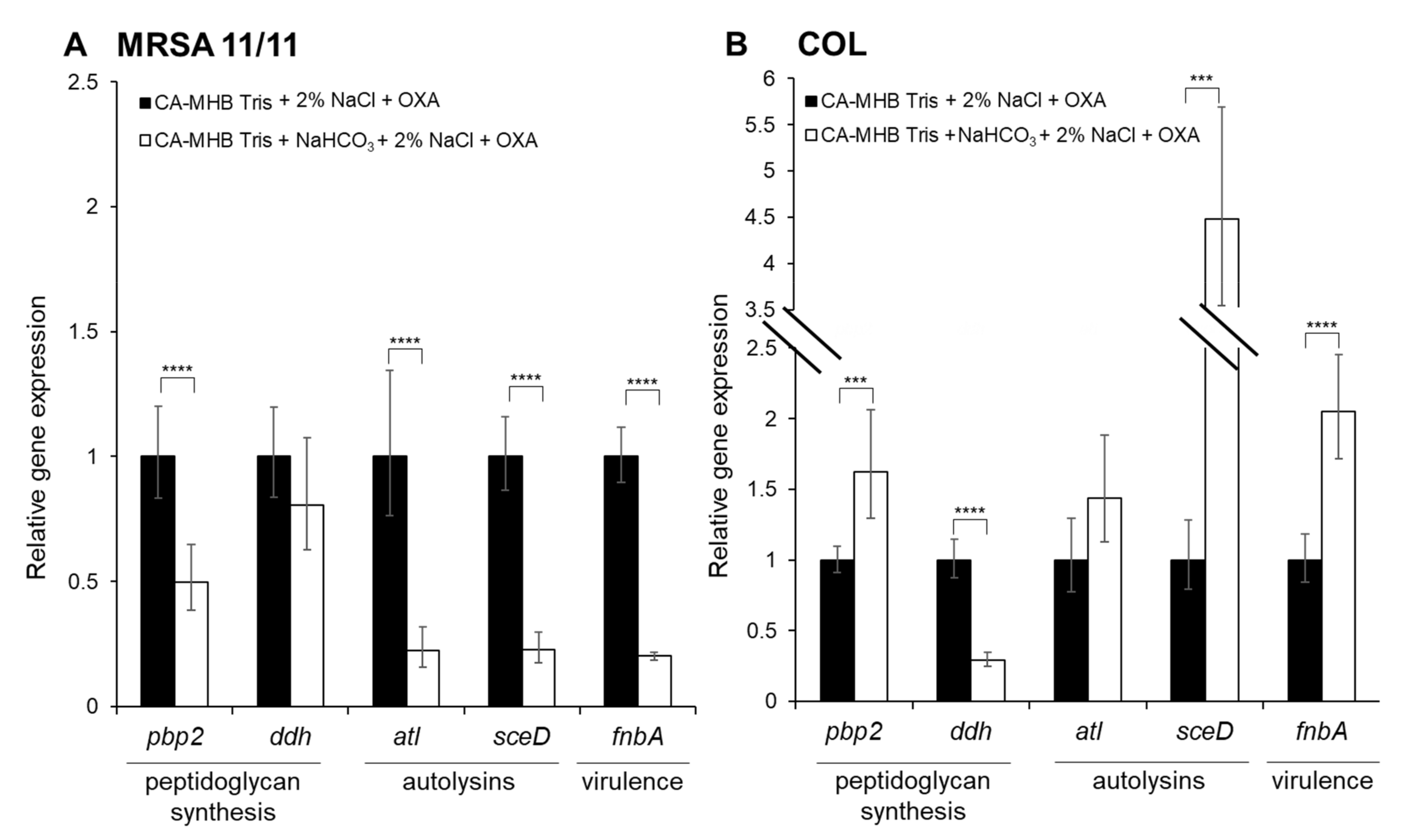
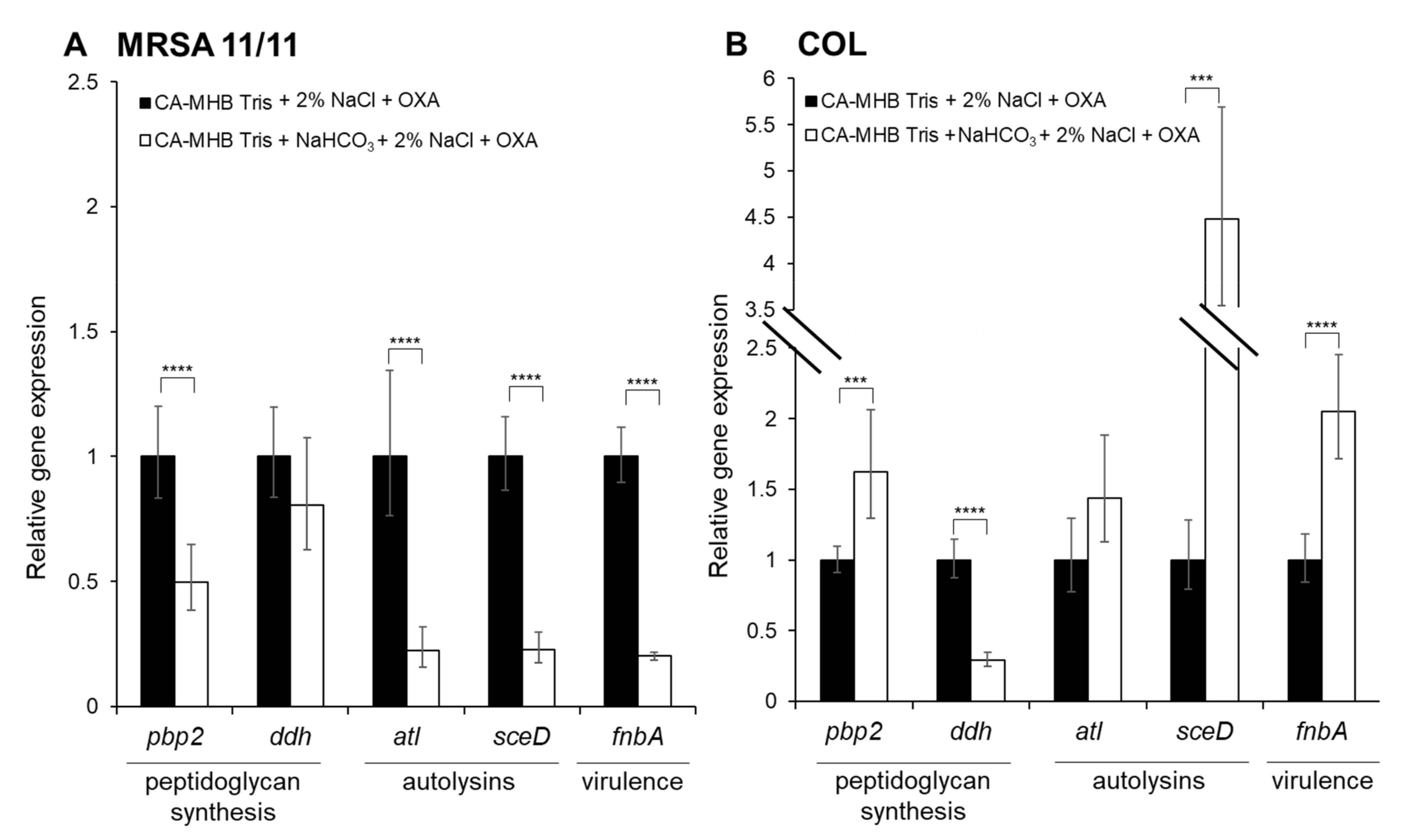
3.2. qRT-PCR Validation
3.3. Limitations of Study
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- CDC. Antibiotic Resistance Threats in the United States, 2019; U.S. Deppartment of Health and Human Services, CDC: Atlanta, GA, USA, 2019.
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277. [Google Scholar]
- Abadi, A.T.B.; Rizvanov, A.A.; Haertlé, T.; Blatt, N.L. World Health Organization report: Current crisis of antibiotic resistance. BioNanoScience 2019, 9, 778–788. [Google Scholar] [CrossRef]
- WHO. Antimicrobial resistance: Global report on surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Grundmann, H.; Aires-de-Sousa, M.; Boyce, J.; Tiemersma, E. Emergence and resurgence of meticillin-resistant Staphylococcus aureus as a public-health threat. Lancet 2006, 368, 874–885. [Google Scholar] [CrossRef] [Green Version]
- Goetghebeur, M.; Landry, P.-A.; Han, D.; Vicente, C. Methicillin-resistant Staphylococcus aureus: A public health issue with economic consequences. Can. J. Infect. Dis. Med. Microbiol. 2007, 18, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Hassoun, A.; Linden, P.K.; Friedman, B. Incidence, prevalence, and management of MRSA bacteremia across patient populations—a review of recent developments in MRSA management and treatment. Crit. Care 2017, 21, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, I.; Reilly, J.; Bunyan, D.; Walker, A. Costs of healthcare-associated methicillin-resistant Staphylococcus aureus and its control. Clin. Microbiol. Infect. 2010, 16, 1721–1728. [Google Scholar] [CrossRef] [Green Version]
- Purrello, S.; Garau, J.; Giamarellos, E.; Mazzei, T.; Pea, F.; Soriano, A.; Stefani, S. Methicillin-resistant Staphylococcus aureus infections: A review of the currently available treatment options. J. Glob. Antimicrob. Resist. 2016, 7, 178–186. [Google Scholar] [CrossRef] [Green Version]
- Edwards, B.; Andini, R.; Esposito, S.; Grossi, P.; Lew, D.; Mazzei, T.; Novelli, A.; Soriano, A.; Gould, I. Treatment options for methicillin-resistant Staphylococcus aureus (MRSA) infection: Where are we now? J. Glob. Antimicrob. Resist. 2014, 2, 133–140. [Google Scholar] [CrossRef]
- Bamgbola, O. Review of vancomycin-induced renal toxicity: An update. Ther. Adv. Endocrinol. Metab. 2016, 7, 136–147. [Google Scholar] [CrossRef]
- Abraham, G.; Finkelberg, D.; Spooner, L.M. Daptomycin-induced acute renal and hepatic toxicity without rhabdomyolysis. Ann. Pharmacother. 2008, 42, 719–721. [Google Scholar] [CrossRef]
- McConeghy, K.W.; Bleasdale, S.C.; Rodvold, K.A. The empirical combination of vancomycin and a β-lactam for staphylococcal bacteremia. Clin. Infect. Dis. 2013, 57, 1760–1765. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-H.; Kim, K.-H.; Kim, H.-B.; Kim, N.-J.; Kim, E.-C.; Oh, M.-D.; Choe, K.-W. Outcome of vancomycin treatment in patients with methicillin-susceptible Staphylococcus aureus bacteremia. Antimicrob. Agents Chemother. 2008, 52, 192–197. [Google Scholar] [CrossRef] [Green Version]
- Ersoy, S.C.; Abdelhady, W.; Li, L.; Chambers, H.F.; Xiong, Y.Q.; Bayer, A.S. Bicarbonate resensitization of methicillin-resistant Staphylococcus aureus to β-lactam antibiotics. Antimicrob. Agents Chemother. 2019, 63, e00496-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ersoy, S.C.; Otmishi, M.; Milan, V.T.; Li, L.; Pak, Y.; Mediavilla, J.; Chen, L.; Kreiswirth, B.; Chambers, H.F.; Proctor, R.A.; et al. Scope and predictive genetic/phenotypic signatures of ‘bicarbonate [NaHCO3]-responsiveness’ and β-Lactam sensitization among methicillin-resistant Staphylococcus aureus (MRSA). Antimicrob. Agents Chemother. 2020, 5, e02445-19. [Google Scholar] [CrossRef] [Green Version]
- Rose, W.E.; Bienvenida, A.M.; Xiong, Y.Q.; Chambers, H.F.; Bayer, A.S.; Ersoy, S.C. Ability of bicarbonate supplementation to sensitize selected methicillin-resistant Staphylococcus aureus (MRSA) strains to β-lactam antibiotics in an Ex Vivo simulated endocardial vegetation model. Antimicrob. Agents Chemother. 2019, 64, e02072-19. [Google Scholar]
- Ersoy, S.C.; Chambers, H.F.; Proctor, R.A.; Rosato, A.E.; Mishra, N.N.; Xiong, Y.Q.; Bayer, A.S. Impact of bicarbonate on PBP2a production, maturation, and functionality in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2021, 65, e02621-20. [Google Scholar] [CrossRef]
- Ersoy, S.C.; Heithoff, D.M.; Barnes, L.; Tripp, G.K.; House, J.K.; Marth, J.D.; Smith, J.W.; Mahan, M.J. Faculty opinions recommendation of correcting a fundamental flaw in the paradigm for antimicrobial susceptibility testing. EBioMedicine 2019, 20, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Fenn, W.O. The carbon dioxide dissociation curve of nerve and muscle. Am. J. Physiol. Leg. Content 1928, 85, 207–223. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef]
- Sahukhal, G.S.; Pandey, S.; Elasri, M.O. MsaABCR operon is involved in persister cell formation in Staphylococcus aureus. BMC Microbiol. 2017, 17, 218. [Google Scholar] [CrossRef] [Green Version]
- Grassi, L.; Di Luca, M.; Maisetta, G.; Rinaldi, A.C.; Esin, S.; Trampuz, A.; Batoni, G. Generation of persister cells of Pseudomonas aeruginosa and Staphylococcus aureus by chemical treatment and evaluation of their susceptibility to membrane-targeting agents. Front. Microbiol. 2017, 8, 1917. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Persister cells, dormancy and infectious disease. Nat. Rev. Genet. 2006, 5, 48–56. [Google Scholar] [CrossRef]
- Tuchscherr, L.; Löffler, B.; Proctor, R.A. Persistence of Staphylococcus aureus: Multiple metabolic pathways impact the expression of virulence factors in small-colony variants (SCVs). Front. Microbiol. 2020, 11, 1028. [Google Scholar] [CrossRef]
- Traber, K.E.; Lee, E.; Benson, S.; Corrigan, R.; Cantera, M.; Shopsin, B.; Novick, R.P. Agr function in clinical Staphylococcus aureus isolates. Microbiology 2008, 154, 2265. [Google Scholar] [CrossRef] [Green Version]
- Seidl, K.; Chen, L.; Bayer, A.S.; Hady, W.A.; Kreiswirth, B.N.; Xiong, Y.Q. Relationship of agr expression and function with virulence and vancomycin treatment outcomes in experimental endocarditis due to methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 5631–5639. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.K.; Luong, T.T.; Lee, C.Y. RNAIII of the Staphylococcus aureus agr system activates global regulator MgrA by stabilizing mRNA. Proc. Natl. Acad. Sci. USA 2015, 112, 14036–14041. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, C.P.; Boyle-Vavra, S.; Daum, R.S. Importance of the global regulators Agr and SaeRS in the pathogenesis of CA-MRSA USA300 infection. PLoS ONE 2010, 5, e15177. [Google Scholar] [CrossRef] [Green Version]
- Cheung, A.L.; Zhang, G. Global regulation of virulence determinants in Staphylococcus aureus by the SarA protein family. Front Biosci. 2002, 7, d1825–d1842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischoff, M.; Dunman, P.; Kormanec, J.; Macapagal, D.; Murphy, E.; Mounts, W.; Berger-Bächi, B.; Projan, S. Microarray-based analysis of the Staphylococcus aureus σB regulon. J. Bacteriol. 2004, 186, 4085–4099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Wamel, W.; Xiong, Y.-Q.; Bayer, A.; Yeaman, M.; Nast, C.; Cheung, A. Regulation of Staphylococcus aureus type 5 capsular polysaccharides by agr and sarA In Vitro and in an experimental endocarditis model. Microb. Pathog. 2002, 33, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.-Q.; Bayer, A.S.; Yeaman, M.R.; Van Wamel, W.; Manna, A.C.; Cheung, A.L. Impacts of sarA and agr in Staphylococcus aureus strain newman on fibronectin-binding protein A gene expression and fibronectin adherence capacity In Vitro and in experimental infective endocarditis. Infect. Immun. 2004, 72, 1832–1836. [Google Scholar] [CrossRef] [Green Version]
- Lacey, K.A.; Mulcahy, M.E.; Towell, A.M.; Geoghegan, J.A.; McLoughlin, R.M. Clumping factor B is an important virulence factor during Staphylococcus aureus skin infection and a promising vaccine target. PLoS Pathog. 2019, 15, e1007713. [Google Scholar] [CrossRef]
- McCrea, K.W.; Hartford, O.; Davis, S.; Eidhin, D.N.; Lina, G.; Speziale, P.; Foster, T.J.; Höök, M. The serine-aspartate repeat (Sdr) protein family in Staphylococcus epidermidis. Microbiology 2000, 146, 1535–1546. [Google Scholar] [CrossRef]
- Kong, C.; Chee, C.-F.; Richter, K.; Thomas, N.; Rahman, N.A.; Nathan, S. Suppression of Staphylococcus aureus biofilm formation and virulence by a benzimidazole derivative, UM-C162. Sci. Rep. 2018, 8, 1–16. [Google Scholar] [CrossRef]
- Ersoy, S.C.; Abdelhady, W.; Li, L.; Xiong, Y.Q.; Bayer, A.S. Bicarbonate represses the sarA/mecA regulatory axis, re-sensitizing selected methicillin-resistant Staphylococcus aureus (MRSA) to β-lactam antibiotics. Gordon Research Conference on Staphylococcal Diseases, poster 36. Castelldefels, Spain, 4–9 August 2019. [Google Scholar]
- Rahman, M.M.; Hunter, H.N.; Prova, S.; Verma, V.; Qamar, A.; Golemi-Kotra, D. Faculty opinions recommendation of the Staphylococcus aureus methicillin resistance factor FmtA is a d-amino esterase that acts on teichoic acids. mBio 2016, 7, 02070-15. [Google Scholar] [CrossRef] [Green Version]
- Straume, D.; Piechowiak, K.W.; Kjos, M.; Håvarstein, L.S. Class A PBPs: It is time to rethink traditional paradigms. Mol. Microbiol. 2021, 116, 41–52. [Google Scholar] [CrossRef]
- Łęski, T.A.; Tomasz, A. Role of penicillin-binding protein 2 (PBP2) in the antibiotic susceptibility and cell wall cross-linking of Staphylococcus aureus: Evidence for the cooperative functioning of PBP2, PBP4, and PBP2A. J. Bacteriol. 2005, 187, 1815–1824. [Google Scholar] [CrossRef] [Green Version]
- Boyle-Vavra, S.; De Jonge, B.; Ebert, C.C.; Daum, R.S. Cloning of the Staphylococcus aureus ddh gene encoding NAD+-dependent d-lactate dehydrogenase and insertional inactivation in a glycopeptide-resistant isolate. J. Bacteriol. 1997, 179, 6756–6763. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.-A.; Yoshii, Y.; Yamada, S.; Nagakura, M.; Kinjo, Y.; Mizunoe, Y.; Okuda, K.-i. Roles of lytic transglycosylases in biofilm formation and β-lactam resistance in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2019, 63, e01277-19. [Google Scholar] [CrossRef]
- Stapleton, M.R.; Horsburgh, M.J.; Hayhurst, E.J.; Wright, L.; Jonsson, I.-M.; Tarkowski, A.; Kokai-Kun, J.F.; Mond, J.J.; Foster, S.J. Characterization of IsaA and SceD, two putative lytic transglycosylases of Staphylococcus aureus. J. Bacteriol. 2007, 189, 7316–7325. [Google Scholar] [CrossRef] [Green Version]
- Biswas, R.; Martinez, R.E.; Göhring, N.; Schlag, M.; Josten, M.; Xia, G.; Hegler, F.; Gekeler, C.; Gleske, A.-K.; Götz, F. Proton-binding capacity of Staphylococcus aureus wall teichoic acid and its role in controlling autolysin activity. PLoS ONE 2012, 7, e41415. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Verma, V.; Belcheva, A.; Singh, A.; Fridman, M.; Golemi-Kotra, D. Staphylococcus aureus methicillin-resistance factor fmtA is regulated by the global regulator SarA. PLoS ONE 2012, 7, e43998. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Classifications of Gene Functions | Number of Genes/Transcriptional Units Per Category | Number Increased | Number Decreased |
|---|---|---|---|
| Virulence | 12 | 2 | 10 |
| Cell Wall Synthesis | 10 | 1 | 9 |
| Membrane | 19 | 4 | 15 |
| Metabolism | 22 | 12 | 10 |
| Transcription | 5 | 1 | 4 |
| Translation | 6 | 5 | 1 |
| Transposition | 2 | 2 | 0 |
| Stress Response | 2 | 1 | 1 |
| Totals: | 78 | 28 | 50 |
| Gene Name | Locus Tag | +NaHCO3 (no OXA) | +NaHCO3 with OXA B | Function | −2 | 0 | 2 |
|---|---|---|---|---|---|---|---|
| clfA | SAUSA300_0772 | −2.18 | −2.97 | virulence | |||
| clfB | SAUSA300_2565 | 1.29 | 1.11 | ||||
| agr | SAUSA300_1989 | −3.21 | −5.37 | ||||
| sdrH | SAUSA300_1985 | −2.44 | 0.00 | ||||
| fnbA | SAUSA300_2441 | 0.00 | −1.84 | ||||
| fnbB | SAUSA300_2440 | 0.00 | −1.47 | ||||
| atl | SAUSA300_0955 | −1.65 | −2.93 | autolysins | |||
| sceD | SAUSA300_2051 | −6.53 | −4.41 | ||||
| isaA | SAUSA300_2506 | 0.00 | −2.12 | ||||
| fmtA | SAUSA300_0959 | 0.00 | −1.32 | peptidoglycan/cell wall synthesis | |||
| ddh | SAUSA300_2463 | 0.00 | −4.00 | ||||
| pbp2 | SAUSA300_1341 | 0.00 | −1.81 | ||||
| bccT | SAUSA300_2549 | −2.45 | −2.42 | osmotic stress | |||
| usp | SAUSA300_0067 | −1.30 | −2.28 | stress response |
| Gene Name | Locus Tag | MRSA 11/11 +NaHCO3 (no OXA) | COL +NaHCO3 (no OXA) | MRSA +NaHCO3 with OXA B | COL +NaHCO3 with OXA B | Function |
|---|---|---|---|---|---|---|
| cap8 | SAUSA300_0152 | −3.05 | 2.36 | −4.45 | 0.00 | virulence |
| clpL | SAUSA300_2486 | 0.00 | 0.00 | −1.44 | −1.46 | |
| sasD | SAUSA300_0136 | 0.00 | 0.90 | 2.11 | −1.82 | |
| aaa | SAUSA300_0438 | 0.00 | −1.14 | −2.20 | 0.00 | |
| vraX | SAUSA300_RS03005 | 0.00 | 0.00 | −4.45 | 1.88 | |
| kdpABCDF | SAUSA300_2032 | 0.00 | 0.90 | −2.07 | 1.79 | osmotic stress |
| betAB | SAUSA300_2545 | −4.44 | −1.38 | −5.12 | −4.28 | |
| icaR | SAUSA300_2599 | 0.00 | 1.87 | 1.62 | 0.00 | transcriptional regulator |
| rsp | SAUSA300_2326 | −1.60 | −1.02 | 0.00 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ersoy, S.C.; Hanson, B.M.; Proctor, R.A.; Arias, C.A.; Tran, T.T.; Chambers, H.F.; Bayer, A.S. Impact of Bicarbonate-β-Lactam Exposures on Methicillin-Resistant Staphylococcus aureus (MRSA) Gene Expression in Bicarbonate-β-Lactam-Responsive vs. Non-Responsive Strains. Genes 2021, 12, 1650. https://doi.org/10.3390/genes12111650
Ersoy SC, Hanson BM, Proctor RA, Arias CA, Tran TT, Chambers HF, Bayer AS. Impact of Bicarbonate-β-Lactam Exposures on Methicillin-Resistant Staphylococcus aureus (MRSA) Gene Expression in Bicarbonate-β-Lactam-Responsive vs. Non-Responsive Strains. Genes. 2021; 12(11):1650. https://doi.org/10.3390/genes12111650
Chicago/Turabian StyleErsoy, Selvi C., Blake M. Hanson, Richard A. Proctor, Cesar A. Arias, Truc T. Tran, Henry F. Chambers, and Arnold S. Bayer. 2021. "Impact of Bicarbonate-β-Lactam Exposures on Methicillin-Resistant Staphylococcus aureus (MRSA) Gene Expression in Bicarbonate-β-Lactam-Responsive vs. Non-Responsive Strains" Genes 12, no. 11: 1650. https://doi.org/10.3390/genes12111650
APA StyleErsoy, S. C., Hanson, B. M., Proctor, R. A., Arias, C. A., Tran, T. T., Chambers, H. F., & Bayer, A. S. (2021). Impact of Bicarbonate-β-Lactam Exposures on Methicillin-Resistant Staphylococcus aureus (MRSA) Gene Expression in Bicarbonate-β-Lactam-Responsive vs. Non-Responsive Strains. Genes, 12(11), 1650. https://doi.org/10.3390/genes12111650