
Novel Candidate Genes Differentially Expressed in Glyphosate-Treated Horseweed (Conyza canadensis)

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Sample Preparation and RNA Extraction
2.2. cDNA Library Preparation and Illumina Sequencing
2.3. Read Quality Control and De novo Transcriptome Analysis
2.4. Differentially Expressed Gene (DEG) Analysis
2.5. Validation of Representative DEGs by Quantitative Reverse Transcription Polymerase Chain Reaction (RT-qPCR)
2.6. Transcript Alignments and Prediction of Gene Identities
2.7. Calculation Correlation Efficiency and Plot Generation
2.8. Gene Ontology (GO) Analysis
3. Results
3.1. Bioinformatic Information of Illumina Sequencing
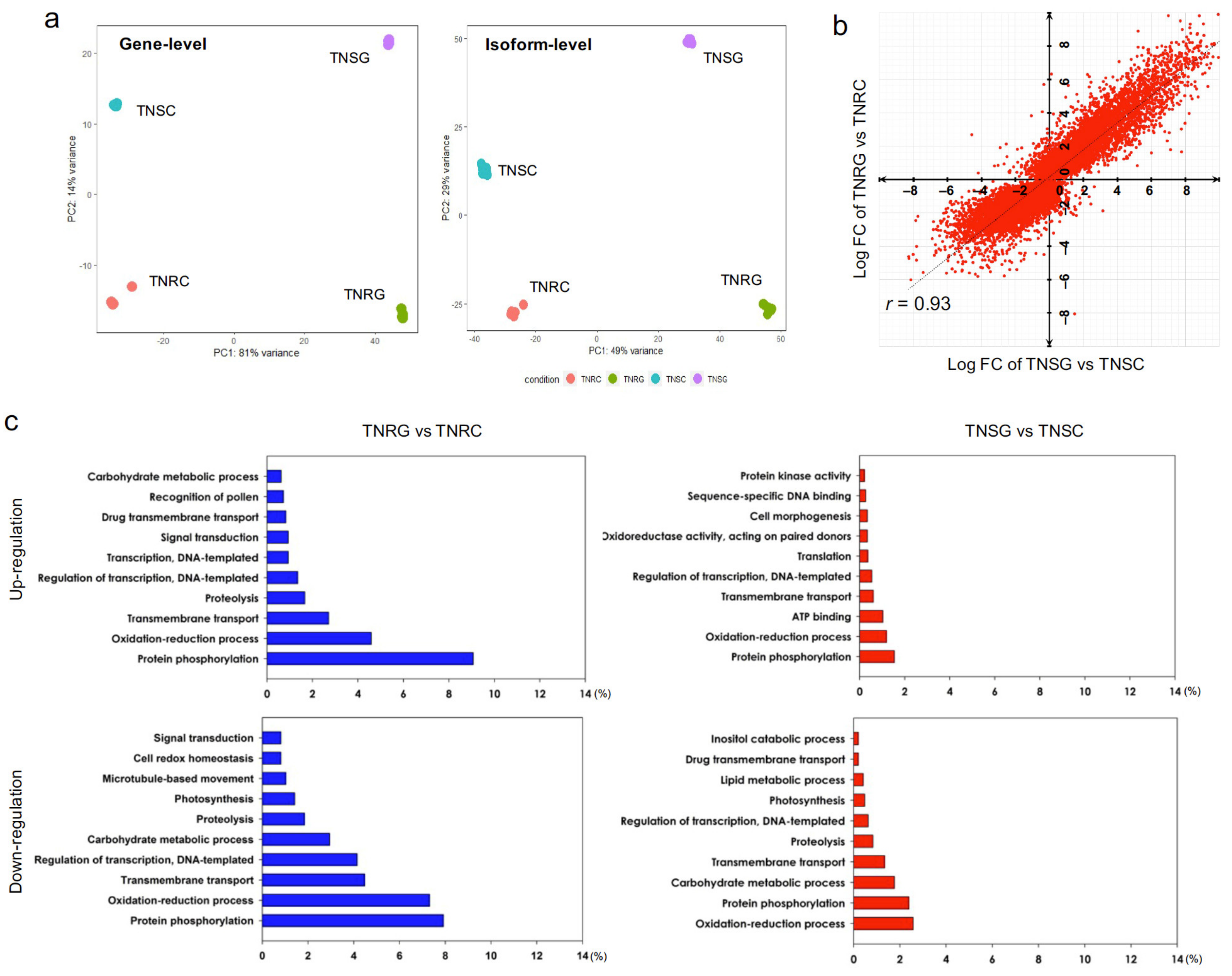
3.2. De novo Transcriptome Generation and DEG Analysis for TNRG versus (vs.) TNRC and TNSG vs. TNSC
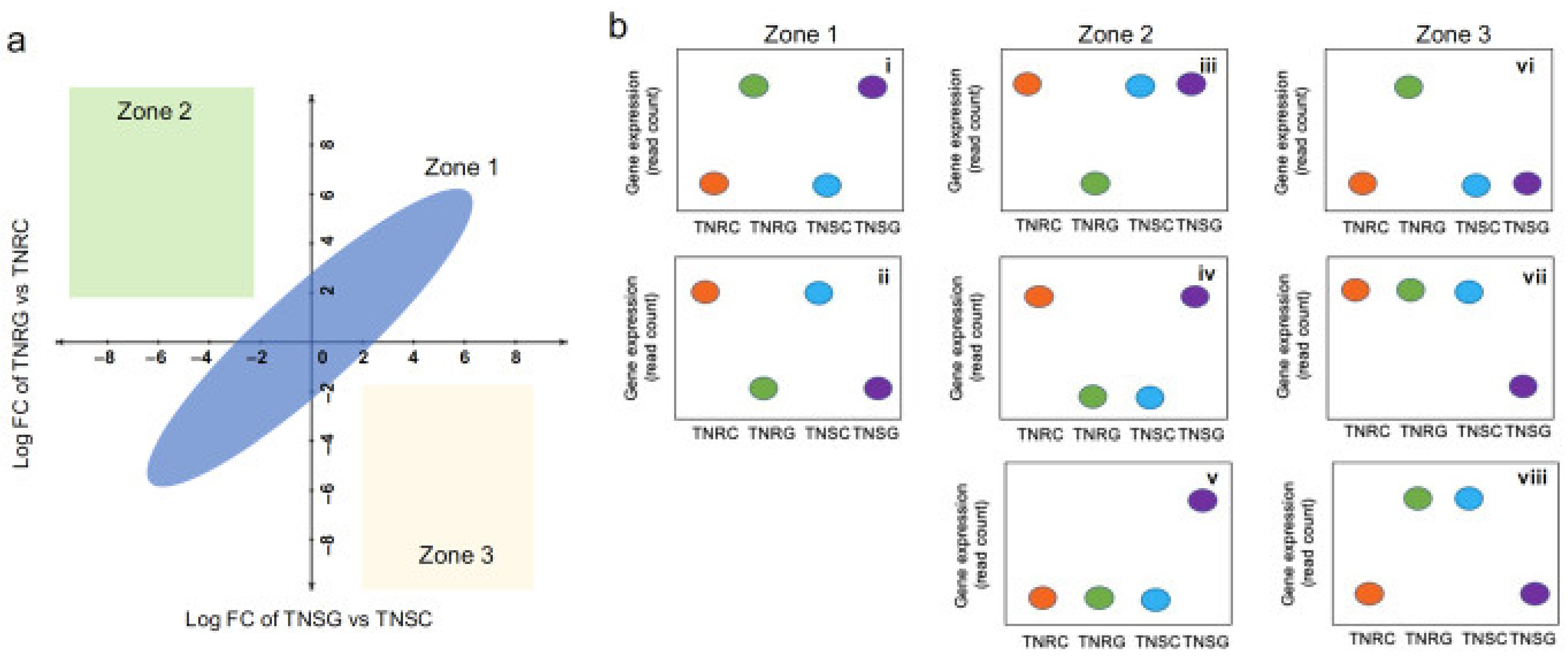
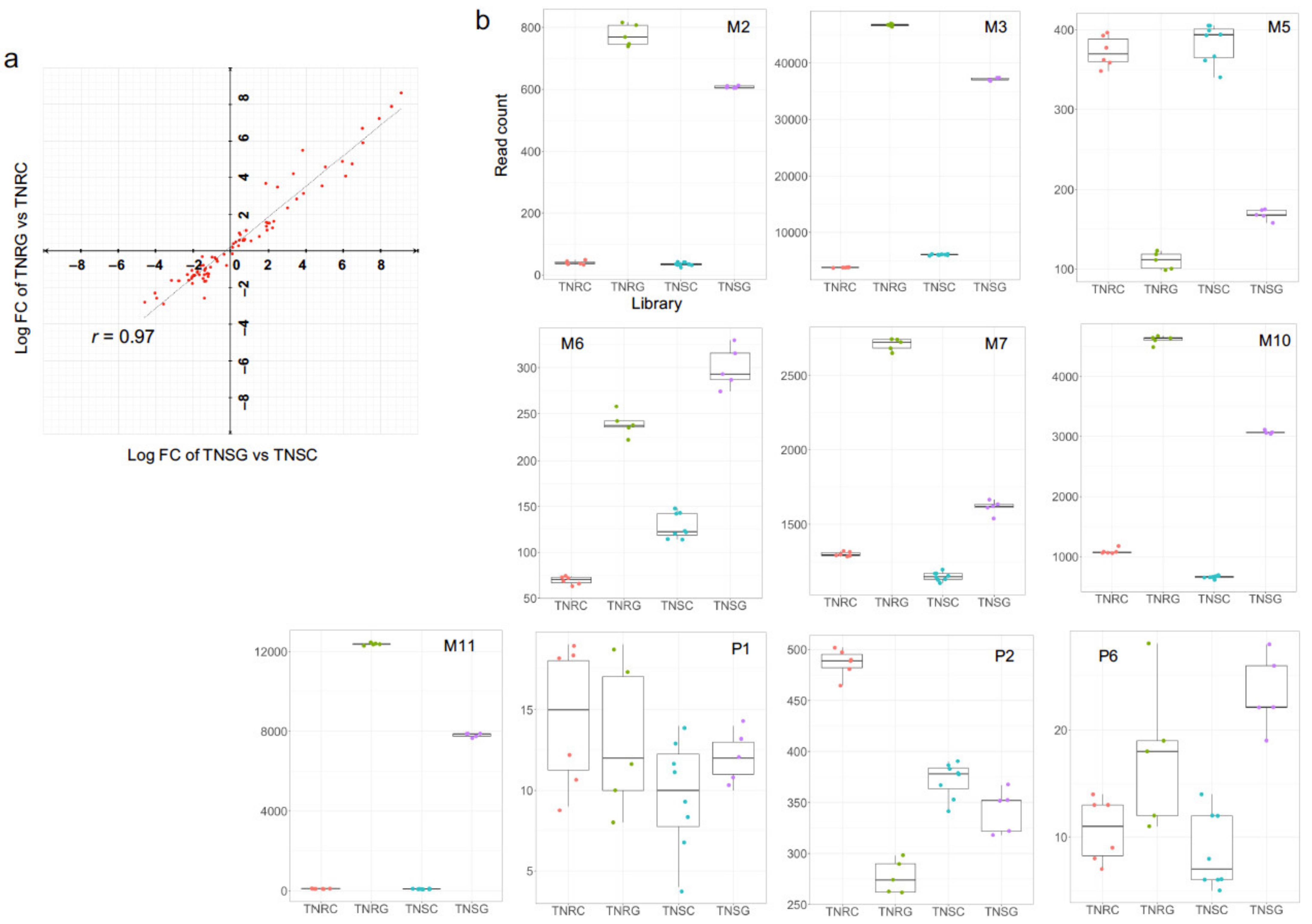
3.3. DEG Groups Dependent on Glyphosate Treatment
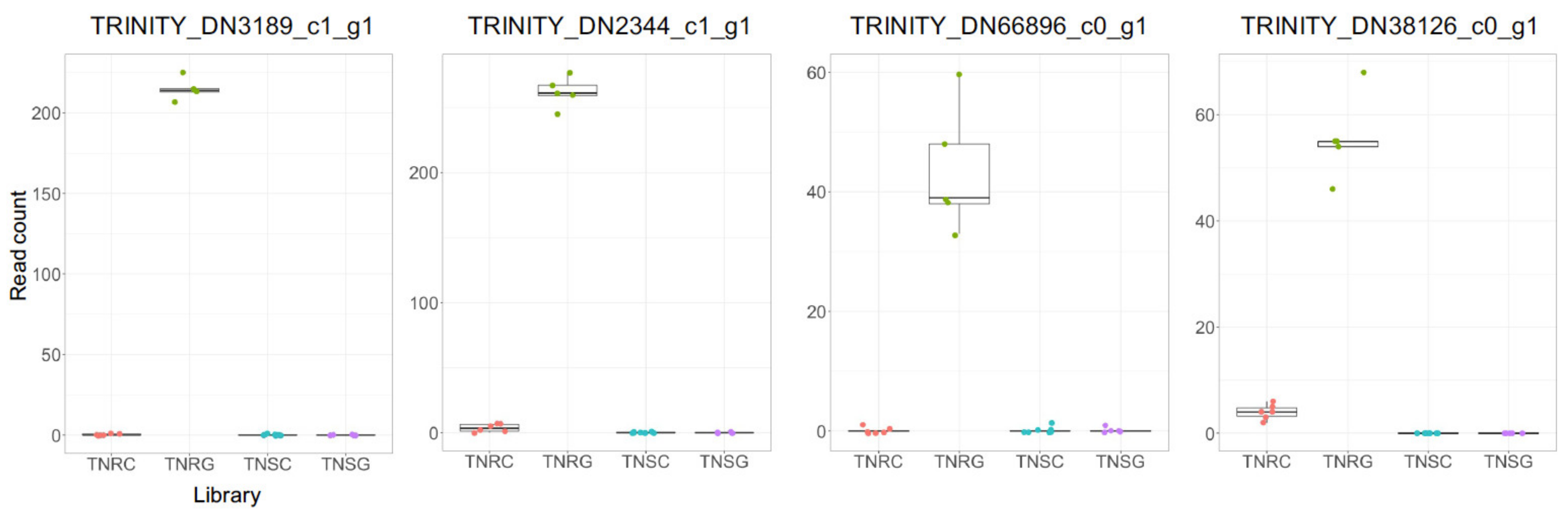
3.4. DEG Groups between TNR and TNS without Glyphosate Treatment
3.5. Differential Expression of Well-Known NTSR Genes Related to Metabolic Response to Glyphosate
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duke, S.O. The history and current status of glyphosate. Pest Manag. Sci. 2018, 74, 1027–1034. [Google Scholar] [CrossRef]
- Amrhein, N.; Deus, B.; Gehrke, P.; Steinrucken, H.C. The site of the inhibition of the shikimate pathway by glyphosate: II. Interference of glyphosate with chorismate formation in vivo and in vitro. Plant Physiol. 1980, 66, 830–834. [Google Scholar] [CrossRef] [Green Version]
- Steinrucken, H.C.; Amrhein, N. The herbicide glyphosate is a potent inhibitor of 5-enolpyruvyl-shikimic acid-3-phosphate synthase. Biochem. Biophys. Res. Commun. 1980, 94, 1207–1212. [Google Scholar] [CrossRef]
- Pratley, J.; Urwin, N.; Stanton, R.; Baines, P.; Broster, J.; Cullis, K.; Schafer, D.; Bohn, J.; Krueger, R. Resistance to glyphosate in Lolium rigidum. I. Bioevaluation. Weed Sci. 1999, 47, 405–411. [Google Scholar] [CrossRef]
- Heap, I.; Duke, S.O. Overview of glyphosate-resistant weeds worldwide. Pest Manag. Sci. 2018, 74, 1040–1049. [Google Scholar] [CrossRef]
- Wechsler, S.J.; McFadden, J.R.; Smith, D.J. What do farmers’ weed control decisions imply about glyphosate resistance? Evidence from surveys of US corn fields. Pest Manag. Sci. 2018, 74, 1143–1154. [Google Scholar] [CrossRef]
- Heap, I. Global perspective of herbicide-resistant weeds. Pest Manag. Sci. 2014, 70, 1306–1315. [Google Scholar] [CrossRef]
- Sammons, R.D.; Gaines, T.A. Glyphosate resistance: State of knowledge. Pest Manag. Sci. 2014, 70, 1367–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delye, C. Unravelling the genetic bases of non-target-site-based resistance (NTSR) to herbicides: A major challenge for weed science in the forthcoming decade. Pest Manag. Sci. 2013, 69, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Bell, M.S.; Hager, A.G.; Tranel, P.J. Multiple resistance to herbicides from four site-of-action groups in waterhemp (Amaranthus tuberculatus). Weed Sci. 2013, 61, 460–468. [Google Scholar] [CrossRef]
- Nandula, V.K.; Ray, J.D.; Ribeiro, D.N.; Pan, Z.; Reddy, K.N. Glyphosate resistance in tall waterhemp (Amaranthus tuberculatus) from Mississippi is due to both altered target-site and nontarget-site mechanisms. Weed Sci. 2013, 61, 374–383. [Google Scholar] [CrossRef]
- Moretti, M.L.; Alarcon-Reverte, R.; Pearce, S.; Morran, S.; Hanson, B.D. Transcription of putative tonoplast transporters in response to glyphosate and paraquat stress in Conyza bonariensis and Conyza canadensis and selection of reference genes for qRT-PCR. PLoS ONE 2017, 12, e0180794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hereward, J.P.; Werth, J.A.; Thornby, D.F.; Keenan, M.; Chauhan, B.S.; Walter, G.H. Gene expression in response to glyphosate treatment in fleabane (Conyza bonariensis)—glyphosate death response and candidate resistance genes. Pest Manag. Sci. 2018, 74, 2346–2355. [Google Scholar] [CrossRef]
- Piasecki, C.; Yang, Y.; Benemann, D.P.; Kremer, F.S.; Galli, V.; Millwood, R.J.; Cechin, J.; Agostinetto, D.; Maia, L.C.; Vargas, L.; et al. Transcriptomic analysis identifies new non-target site glyphosate-resistance genes in Conyza bonariensis. Plants 2019, 8, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcantara-de la Cruz, R.; Rojano-Delgado, A.M.; Gimenez, M.J.; Cruz-Hipolito, H.E.; Dominguez-Valenzuela, J.A.; Barro, F.; De Prado, R. First resistance mechanisms characterization in glyphosate-resistant Leptochloa virgata. Front. Plant Sci. 2016, 7, 1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunharo, C.A.; Patterson, E.L.; Carrijo, D.R.; de Melo, M.S.; Nicolai, M.; Gaines, T.A.; Nissen, S.J.; Christoffoleti, P.J. Confirmation and mechanism of glyphosate resistance in tall windmill grass (Chloris elata) from Brazil. Pest Manag. Sci. 2016, 72, 1758–1764. [Google Scholar] [CrossRef]
- Vila-Aiub, M.M.; Balbi, M.C.; Distefano, A.J.; Fernandez, L.; Hopp, E.; Yu, Q.; Powles, S.B. Glyphosate resistance in perennial Sorghum halepense (Johnsongrass), endowed by reduced glyphosate translocation and leaf uptake. Pest Manag. Sci. 2012, 68, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; d’Avignon, D.A.; Ackerman, J.J.; Collavo, A.; Sattin, M.; Ostrander, E.L.; Hall, E.L.; Sammons, R.D.; Preston, C. Vacuolar glyphosate-sequestration correlates with glyphosate resistance in ryegrass (Lolium spp.) from Australia, South America, and Europe: A 31P NMR investigation. J. Agric. Food Chem. 2012, 60, 1243–1250. [Google Scholar] [CrossRef]
- Ge, X.; d’Avignon, D.A.; Ackerman, J.J.; Sammons, R.D. Rapid vacuolar sequestration: The horseweed glyphosate resistance mechanism. Pest Manag. Sci. 2010, 66, 345–348. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.; d’Avignon, D.A.; Ackerman, J.J.; Sammons, R.D. In vivo 31P-nuclear magnetic resonance studies of glyphosate uptake, vacuolar sequestration, and tonoplast pump activity in glyphosate-resistant horseweed. Plant Physiol. 2014, 166, 1255–1268. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Abercrombie, L.L.; Yuan, J.S.; Riggins, C.W.; Sammons, R.D.; Tranel, P.J.; Stewart, C.N., Jr. Characterization of the horseweed (Conyza canadensis) transcriptome using GS-FLX 454 pyrosequencing and its application for expression analysis of candidate non-target herbicide resistance genes. Pest Manag. Sci. 2010, 66, 1053–1062. [Google Scholar] [CrossRef]
- Tani, E.; Chachalis, D.; Travlos, I.S.; Bilalis, D. Environmental conditions influence induction of key ABC-transporter genes affecting glyphosate resistance mechanism in Conyza canadensis. Int. J. Mol. Sci. 2016, 17, 342. [Google Scholar] [CrossRef] [Green Version]
- Laforest, M.; Martin, S.L.; Bisaillon, K.; Soufiane, B.; Meloche, S.; Page, E. A chromosome-scale draft sequence of the Canada fleabane genome. Pest Manag. Sci. 2020, 76, 2158–2169. [Google Scholar] [CrossRef]
- Peng, Y.; Lai, Z.; Lane, T.; Nageswara-Rao, M.; Okada, M.; Jasieniuk, M.; O’Geen, H.; Kim, R.W.; Sammons, R.D.; Rieseberg, L.H.; et al. De novo genome assembly of the economically important weed horseweed using integrated data from multiple sequencing platforms. Plant Physiol. 2014, 166, 1241–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewels, P.; Magnusson, M.; Lundin, S.; Kaller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NCBI Resource Coordinators. Database resources of the National Center for Biotechnology. Nucleic Acids Res. 2016, 44, D7–D19. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinformatics 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- RStudio Team. RStudio: Integrated Development for R; RStudio Inc.: Boston, MA, USA, 2020; Available online: http://www.rsutdio.com/ (accessed on 28 June 2021).
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Perotti, V.E.; Larran, A.S.; Palmieri, V.E.; Martinatto, A.K.; Permingeat, H.R. Herbicide resistant weeds: A call to integrate conventional agricultural practices, molecular biology knowledge and new technologies. Plant Sci. 2020, 290, 110255. [Google Scholar] [CrossRef] [PubMed]
- Duhoux, A.; Carrere, S.; Gouzy, J.; Bonin, L.; Delye, C. RNA-Seq analysis of rye-grass transcriptomic response to an herbicide inhibiting acetolactate-synthase identifies transcripts linked to non-target-site-based resistance. Plant Mol. Biol. 2015, 87, 473–487. [Google Scholar] [CrossRef]
- Gaines, T.A.; Lorentz, L.; Figge, A.; Herrmann, J.; Maiwald, F.; Ott, M.C.; Han, H.; Busi, R.; Yu, Q.; Powles, S.B.; et al. RNA-Seq transcriptome analysis to identify genes involved in metabolism-based diclofop resistance in Lolium rigidum. Plant J. 2014, 78, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Iwakami, S.; Uchino, A.; Kataoka, Y.; Shibaike, H.; Watanabe, H.; Inamura, T. Cytochrome P450 genes induced by bispyribac-sodium treatment in a multiple-herbicide-resistant biotype of Echinochloa phyllopogon. Pest Manag. Sci. 2014, 70, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Gao, H.; Xia, W.; Zhang, T.; Dong, L. Establishing a herbicide-metabolizing enzyme library in Beckmannia syzigachne to identify genes associated with metabolic resistance. J. Exp. Bot. 2016, 67, 1745–1757. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Yan, Y.; Ge, L.; Zhu, B.; Liu, W.; Wang, J. Target site mutations and cytochrome P450s confer resistance to fenoxaprop-P-ethyl and mesosulfuron-methyl in Alopecurus aequalis. Pest Manag. Sci. 2019, 75, 204–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leslie, T.; Baucom, R.S. De novo assembly and annotation of the transcriptome of the agricultural weed Ipomoea purpurea uncovers gene expression changes associated with herbicide resistance. G3 Genes Genomes Genet. 2014, 4, 2035–2047. [Google Scholar] [CrossRef] [Green Version]
- Conte, S.S.; Lloyd, A.M. Exploring multiple drug and herbicide resistance in plants-spotlight on transporter proteins. Plant Sci. 2011, 180, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.; Demosthenis, C.; Ilias, S.T. A glyphosate resistance mechanism in Conyza canadensis involves synchronization of EPSPS and ABC-transporter genes. Plant Mol. Biol. Rep. 2015, 33, 1721–1730. [Google Scholar] [CrossRef]
- Sun, Y.; Qiao, Z.; Muchero, W.; Chen, J.G. Lectin receptor-like kinases: The sensor and mediator at the plant cell surface. Front. Plant Sci. 2020, 11, 596301. [Google Scholar] [CrossRef]
- Teixeira, M.A.; Rajewski, A.; He, J.; Castaneda, O.G.; Litt, A.; Kaloshian, I. Classification and phylogenetic analyses of the Arabidopsis and tomato G-type lectin receptor kinases. BMC Genom. 2018, 19, 239. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Labbe, J.; Muchero, W.; Yang, X.; Jawdy, S.S.; Kennedy, M.; Johnson, J.; Sreedasyam, A.; Schmutz, J.; Tuskan, G.A.; et al. Genome-wide analysis of lectin receptor-like kinases in Populus. BMC Genom. 2016, 17, 699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, B.; Saikkonen, K.; Helander, M. Glyphosate-modulated biosynthesis driving plant defense and species interactions. Trends Plant Sci. 2021, 26, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Bonaventure, G. The Nicotiana attenuata Lectin receptor kinase 1 is involved in the perception of insect feeding. Plant Signal. Behav. 2011, 6, 2060–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilardoni, P.A.; Hettenhausen, C.; Baldwin, I.T.; Bonaventure, G. Nicotiana attenuata Lectin receptor kinase1 suppresses the insect-mediated inhibition of induced defense responses during Manduca sexta herbivory. Plant Cell 2011, 23, 3512–3532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, L.; Yu, Q.; Wang, J.; Han, H.; Mao, L.; Nyporko, A.; Maguza, A.; Fan, L.; Bai, L.; Powles, S. An ABCC-type transporter endowing glyphosate resistance in plants. Proc. Natl. Acad. Sci. USA 2021, 118, e2100136118. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | De novo Assembly Statistics |
|---|---|
| Total assembled reads | 100,204,924 |
| Total isoform level contigs | 105,354 |
| Total gene level contigs | 77,072 |
| Contig N50 (bp) | 1780 |
| Average contig length (bp) | 1039.7 |
| GC (%) | 38.21 |
| Contig ID | LogFC | FDR | Protein Accession ID | Description |
|---|---|---|---|---|
| TRINITY_DN905_c0_g1 | 11.8 | 0 | XP_021979261.1 | Uncharacterized protein LOC110875373 |
| TRINITY_DN13727_c0_g1 | 10.3 | 4.0 × 10−249 | OTG03028.1 | F-box domain-containing protein |
| TRINITY_DN27339_c0_g1 | 10.2 | 0 | XP_023758053.1 | Glutathione S-transferase parA |
| TRINITY_DN36883_c0_g1 | 10.3 | 0 | PWA57492.1 | Aminotransferase, class I/classII |
| TRINITY_DN1722_c0_g2 | 10.1 | 0 | PWA51732.1 | Hypothetical protein CTI12_AA461460 |
| TRINITY_DN66403_c0_g1 | 10 | 0 | XP_024958679.1 | Uncharacterized protein LOC112499602 |
| TRINITY_DN6438_c0_g1 | 9.9 | 0 | XP_024988661.1 | AAA-ATPase At3g50940-like |
| TRINITY_DN4388_c0_g1 | 9.9 | 0 | XP_022010209.1 | Uncharacterized protein LOC110909706 |
| TRINITY_DN11441_c0_g1 | 9.7 | 1.2 × 10−165 | XP_022008773.1 | Uncharacterized protein LOC110908182 |
| TRINITY_DN3079_c0_g1 | 9.7 | 0 | PWA86457.1 | NADH-ubiquinone reductase complex 1 MLRQ subunit |
| TRINITY_DN37237_c0_g1 | 9.6 | 0 | PLY74787.1 | Hypothetical protein LSAT_6X72041 |
| TRINITY_DN56890_c0_g1 | 9.4 | 1.3 × 10−136 | PWA59237.1 | Hypothetical protein CTI12_AA394960 |
| TRINITY_DN2953_c0_g1 | 9.4 | 1.5 × 10−131 | XP_022005344.1 | Inactive ATP-dependent zinc metalloprotease FTSHI 3 |
| TRINITY_DN11270_c0_g1 | 9.3 | 1.4 × 10−128 | XP_022000952.1 | Ultraviolet-B receptor UVR8-like |
| TRINITY_DN5461_c0_g1 | 9.3 | 2.3 × 10−294 | XP_021999019.1 | Phospholipase A1-Igamma2, chloroplastic-like |
| TRINITY_DN68599_c0_g1 | 9.3 | 1.1 × 10−122 | PLY95867.1 | Hypothetical protein LSAT_5X173860 |
| TRINITY_DN282_c0_g1 | 9.3 | 0 | AAT72931.1 | Cascarilladiene synthase |
| TRINITY_DN48830_c0_g1 | 9.2 | 2.9 × 10−112 | XP_023756578.1 | Flavin-containing monooxygenase 1 |
| TRINITY_DN669_c1_g1 | 9.2 | 0 | AAT72931.1 | Cascarilladiene synthase |
| TRINITY_DN310_c0_g3 | 9.1 | 1.0 × 10−248 | PWA98523.1 | ABC transporter Tap-like |
| Contig ID | LogFC | FDR | Protein Accession ID | Description |
|---|---|---|---|---|
| TRINITY_DN3992_c0_g1 | −8.3 | 0 | XP_021976551.1 | Chlorophyll a-b binding protein of LHCII type 1 |
| TRINITY_DN2551_c0_g1 | −8.3 | 0 | WP_124736833.1 | Hypothetical protein |
| TRINITY_DN9403_c0_g1 | −8.1 | 0 | PWA49348.1 | Chlorophyll A-B binding protein |
| TRINITY_DN11212_c0_g1 | −8.0 | 2.0 × 10−48 | XP_023760034.1 | Uncharacterized protein LOC111908441 |
| TRINITY_DN14399_c0_g1 | −7.9 | 3.9 × 10−45 | XP_023762447.1 | WD repeat and HMG-box DNA-binding protein 1 |
| TRINITY_DN42_c2_g1 | −7.8 | 0 | KVI00865.1 | Chlorophyll A-B binding protein |
| TRINITY_DN18783_c0_g1 | −7.8 | 7.2 × 10−43 | XP_019249685.1 | Uncharacterized protein LOC109228892 |
| TRINITY_DN38340_c0_g1 | −7.7 | 8.6 × 10−41 | XP_021973416.1 | Origin of replication complex subunit 1A-like |
| TRINITY_DN42_c4_g1 | −7.4 | 0 | XP_023739911.1 | Chlorophyll a-b binding protein of LHCII type 1 |
| TRINITY_DN14573_c0_g1 | −6.9 | 5.0 × 10−22 | XP_016071745.1 | Uncharacterized protein LOC107539687 |
| TRINITY_DN14043_c0_g1 | −6.8 | 5.3 × 10−53 | XP_023763882.1 | High mobility group B protein 7 |
| TRINITY_DN2800_c0_g1 | −6.8 | 2.1 × 10−114 | XP_023761206.1 | Palmitoyl-monogalactosyldiacylglycerol delta-7 desaturase |
| TRINITY_DN1183_c1_g1 | −6.8 | 2.1 × 10−51 | XP_024991237.1 | Zinc finger protein At1g68190 |
| TRINITY_DN200_c0_g2 | −6.7 | 0 | AEY78525.1 | Chlorophyll a/b-binding protein |
| TRINITY_DN1119_c0_g1 | −6.7 | 0 | KVI00865.1 | Chlorophyll A-B binding protein |
| TRINITY_DN70454_c0_g1 | −6.7 | 1.5 × 10−19 | XP_022016163.1 | Uncharacterized protein At1g04910 |
| TRINITY_DN63729_c0_g1 | −6.5 | 5.9 × 10−17 | PWA63735.1 | Phosphate-induced protein 1 |
| TRINITY_DN15048_c0_g1 | −6.4 | 3.8 × 10−16 | XP_022037435.1 | Pathogenesis-related protein PR-1-like |
| TRINITY_DN57166_c0_g1 | −6.4 | 3.6 × 10−39 | PWA93616.1 | Hypothetical protein CTI12_AA069020 |
| TRINITY_DN56621_c0_g1 | −6.4 | 2.1 × 10−15 | XP_022008540.1 | Uncharacterized protein LOC110907935 |
| Contig ID | LogFC | FDR | Protein Accession ID | Description |
|---|---|---|---|---|
| TRINITY_DN65437_c0_g1 | 10.4 | 0 | XP_021965748.1 | Protein mesh |
| TRINITY_DN1160_c1_g1 | 10.2 | 0 | XP_026462482.1 | Vitellogenin-A1-like |
| TRINITY_DN4388_c0_g1 | 9.9 | 0 | XP_022010209.1 | Uncharacterized protein LOC110909706 |
| TRINITY_DN1722_c0_g2 | 9.9 | 0 | XP_021975061.1 | Uncharacterized protein LOC110870166 |
| TRINITY_DN1160_c0_g1 | 9.8 | 0 | XP_026462481.1 | Vitellogenin-A1-like |
| TRINITY_DN56688_c0_g1 | 9.8 | 0 | PWA98881.1 | Alternative oxidase 1D |
| TRINITY_DN1160_c2_g1 | 9.7 | 2.1 × 10−201 | XP_017781017.1 | Vitellogenin-A1-like |
| TRINITY_DN2458_c0_g1 | 9.7 | 6.7 × 10−206 | AAH21837.1 | Ubc protein |
| TRINITY_DN905_c0_g1 | 9.7 | 0 | XP_021979261.1 | Uncharacterized protein LOC110875373 |
| TRINITY_DN14914_c0_g1 | 9.5 | 7.2 × 10−167 | XP_001654240.1 | Maternal protein exuperantia-2 |
| TRINITY_DN745_c0_g1 | 9.3 | 0 | OTG36297.1 | putative heat shock protein 70 family |
| TRINITY_DN6494_c0_g1 | 9.3 | 4.7 × 10−281 | XP_026272532.1 | Vitellogenin-1-like |
| TRINITY_DN16886_c0_g1 | 9.3 | 2.8 × 10−147 | ACT80192.1 | Cytochrome c oxidase subunit 1 (mitochondrion) |
| TRINITY_DN5166_c0_g1 | 9.2 | 0 | ABV60316.1 | Putative ADP/ATP translocase |
| TRINITY_DN10008_c0_g1 | 9.1 | 6.2 × 10−265 | PWA80350.1 | Hypothetical protein CTI12_AA197740 |
| TRINITY_DN28371_c0_g1 | 9.1 | 2.6 × 10−131 | XP_002066234.1 | Elongation factor 1-α 1 |
| TRINITY_DN1550_c0_g1 | 9.0 | 2.8 × 10−225 | AID52928.1 | 60S acidic ribosomal protein P0 |
| TRINITY_DN19097_c0_g1 | 9.0 | 1.5 × 10−117 | AXX71242.1 | Juvenile hormone |
| TRINITY_DN3463_c0_g1 | 8.8 | 0 | XP_024984476.1 | ADP, ATP carrier protein, mitochondrial |
| TRINITY_DN16233_c0_g1 | 8.7 | 4.0 × 10−105 | YP_009487737.1 | Cytochrome c oxidase subunit III (mitochondrion) |
| Contig ID | LogFC | FDR | Protein Accession ID | Description |
|---|---|---|---|---|
| TRINITY_DN28174_c0_g1 | −8.1 | 2.0 × 10−19 | XP_024970575.1 | Septum-site-determining protein minD homolog, chloroplastic |
| TRINITY_DN9403_c0_g1 | −6.0 | 0 | PWA49348.1 | Chlorophyll A-B binding protein |
| TRINITY_DN17513_c0_g1 | −5.7 | 5.4 × 10−10 | PWA54703.1 | Solute carrier family 25 member 44 |
| TRINITY_DN17284_c0_g1 | −5.6 | 9.4 × 10−10 | XP_023758730.1 | Uncharacterized protein LOC111907163 |
| TRINITY_DN20553_c0_g1 | −5.6 | 7.9 × 10−25 | PWA85753.1 | Protochlorophyllide oxidoreductase |
| TRINITY_DN42_c4_g1 | −5.6 | 0 | XP_023739911.1 | Chlorophyll a-b binding protein of LHCII type 1 |
| TRINITY_DN57253_c0_g1 | −5.5 | 2.4 × 10−9 | PWA80339.1 | Coiled-coil domain containing protein 109 |
| TRINITY_DN58008_c0_g1 | −5.5 | 2.1 × 10−9 | XP_022009704.1 | β-galactosidase 3 |
| TRINITY_DN37787_c0_g1 | −5.4 | 4.8 × 10−21 | XP_023765231.1 | Basic 7S globulin 2-like isoform X2 |
| TRINITY_DN67280_c0_g1 | −5.4 | 2.0 × 10−288 | PWA87944.1 | Hypothetical protein CTI12_AA124470 |
| TRINITY_DN51763_c0_g1 | −5.3 | 5.5 × 10−8 | PWA59983.1 | DNA-binding domain-containing protein |
| TRINITY_DN274_c0_g1 | −5.3 | 0 | PWA65034.1 | Peptidase T2, asparaginase 2, nucleophile aminohydrolase |
| TRINITY_DN57640_c0_g1 | −5.3 | 1.2 × 10−7 | PWA39470.1 | Protochlorophyllide oxidoreductase |
| TRINITY_DN58011_c0_g1 | −5.2 | 9.2 × 10−8 | XP_021990878.1 | Uncharacterized protein LOC110887609 |
| TRINITY_DN18290_c0_g1 | −5.1 | 3.1 × 10−7 | OMO80329.1 | Short-chain dehydrogenase/reductase SDR |
| TRINITY_DN8861_c0_g1 | −5.1 | 1.0 × 10−97 | XP_021986658.1 | Short-chain dehydrogenase TIC 32 |
| TRINITY_DN30954_c0_g1 | −5.1 | 1.5 × 10−6 | XP_023753002.1 | Peroxidase 19-like |
| TRINITY_DN24284_c0_g1 | −5.0 | 1.6 × 10−6 | OTG12714.1 | Putative serine/threonine/dual specificity protein kinase |
| TRINITY_DN43940_c0_g1 | −4.9 | 2.6 × 10−6 | PLY91593.1 | Hypothetical protein LSAT_7X10301 |
| TRINITY_DN48832_c0_g1 | −4.9 | 4.2 × 10−6 | PWA97992.1 | Cytochrome P450 |
| Contig ID | TNRG vs. TNRC | TNSG vs. TNSC | Protein Accession ID | Description | ||
|---|---|---|---|---|---|---|
| Log FC | FDR | LogFC | FDR | |||
| TRINITY_DN3189_c1_g1 | 8.9 | 0 | n.a. | n.a. | XP_023760056.1 | Uncharacterized protein LOC111908459 |
| TRINITY_DN66896_c0_g1 | 8.4 | 4.9 × 10−68 | n.a. | n.a. | XP_022012333.1 | Wall-associated receptor kinase-like 8 |
| TRINITY_DN51395_c0_g1 | 7.3 | 3.7 × 10−33 | n.a. | n.a. | PWA42696.1 | Homeodomain-like protein |
| TRINITY_DN17396_c0_g1 | 6.9 | 1.1 × 10−55 | n.a. | n.a. | XP_021968739.1 | Plasmamembrane calcium-transporting ATPase 12 |
| TRINITY_DN67025_c0_g1 | 6.6 | 1.3 × 10−46 | n.a. | n.a. | KVI07684.1 | Leucine rich repeat 4 |
| TRINITY_DN2344_c1_g1 | 6.1 | 0 | n.a. | n.a. | XP_023771967.1 | Kinesin light chain 3 isoform X2 |
| TRINITY_DN65261_c0_g1 | 5.7 | 3.9 × 10−30 | n.a. | n.a. | XP_023769670.1 | NADH dehydrogenase iron-sulfur protein 8 |
| TRINITY_DN17899_c0_g1 | 5.6 | 4.2 × 10−66 | n.a. | n.a. | PWA34905.1 | B-type lectin domain-containing protein |
| TRINITY_DN67848_c0_g1 | 4.7 | 2.3 × 10−51 | n.a. | n.a. | OTF95985.1 | Malectin-binding domain-containing protein |
| TRINITY_DN15755_c0_g1 | 3.9 | 5.6 × 10−27 | n.a. | n.a. | PWA66199.1 | Toll/interleukin-1 receptor (TIR) domain protein |
| TRINITY_DN7012_c0_g1 | 3.8 | 4.5 × 10−24 | n.a. | n.a. | PWA81859.1 | Toll/interleukin-1 receptor (TIR) domain protein |
| TRINITY_DN38126_c0_g1 | 3.7 | 1.6 × 10−60 | n.a. | n.a. | XP_022016073.1 | L-type lectin-domain containing receptor kinase |
| TRINITY_DN48964_c0_g1 | 3.5 | 7.7 × 10−53 | n.a. | n.a. | XP_022011897.1 | L-type lectin-domain containing receptor kinase |
| TRINITY_DN33216_c0_g1 | 3.2 | 7.7 × 10−41 | n.a. | n.a. | XP_022012440.1 | L-type lectin-domain containing receptor kinase |
| TRINITY_DN15265_c0_g2 | 2.2 | 3.3 × 10−16 | −0.2 | 0.8 | PWA39743.1 | Copper centre Cu(A) |
| Contig ID | TNRG vs. TNRC | TNSG vs. TNSC | Protein Accession ID | Description | ||
|---|---|---|---|---|---|---|
| Log FC | FDR | LogFC | FDR | |||
| TRINITY_DN46383_c0_g1 | −4.2 | 5.9 × 10−60 | 0.4 | 2.2 × 10−2 | XP_022011882.1 | Uncharacterized protein LOC110911561 |
| TRINITY_DN2887_c0_g1 | −3.3 | 0 | −0.1 | 1.2 × 10−3 | XP_022002942.1 | β-glucosidase 24-like |
| TRINITY_DN14379_c0_g1 | −2.7 | 2.8 × 10−52 | −0.5 | 2.4 × 10−5 | XP_023732528.1 | GRAVITROPIC IN THE LIGHT 1-like isoform X1 |
| TRINITY_DN22567_c0_g1 | −2.6 | 4.2 × 10−60 | −0.5 | 2.0 × 10−7 | PWA81344.1 | Hypothetical protein CTI12_AA064670 |
| TRINITY_DN14266_c0_g1 | −2.5 | 6.6 × 10−30 | −0.3 | 4.6 × 10−2 | XP_021982692.1 | Cysteine-rich repeat secretory protein 60-like |
| TRINITY_DN3134_c0_g1 | −2.5 | 1.2 × 10−86 | −0.1 | 4.7 × 10−1 | XP_021969889.1 | Isoflavone reductase homolog |
| TRINITY_DN21830_c0_g1 | −2.4 | 0 | −0.4 | 2.2 × 10−31 | XP_024983361.1 | Uncharacterized protein LOC112519462 |
| TRINITY_DN36753_c0_g1 | −2.4 | 6.7 × 10−70 | 0.5 | 5.3 × 10−10 | XP_010273654.1 | Mannose/glucose-specific lectin-like |
| TRINITY_DN14143_c0_g1 | −2.4 | 4.0 × 10−23 | −0.4 | 2.2 × 10−2 | PWA85337.1 | Hypothetical protein CTI12_AA151440 |
| TRINITY_DN1911_c0_g1 | −2.3 | 6.1 × 10−169 | −0.4 | 1.7 × 10−10 | XP_023773027.1 | Sulfate transporter 3.1-like |
| TRINITY_DN9813_c0_g1 | −2.3 | 1.9 × 10−32 | 0.4 | 7.9 × 10−3 | XP_024959450.1 | Protein LNK1-like isoform X1 |
| TRINITY_DN16564_c0_g1 | −2.3 | 1.4 × 10−31 | −0.2 | 1.2 × 10−1 | PWA68294.1 | WRKY domain-containing protein |
| TRINITY_DN65314_c0_g1 | −2.1 | 1.5 × 10−38 | −0.4 | 7.8 × 10−3 | XP_023763206.1 | Blue copper protein |
| Contig ID | TNRC vs. TNSC | TNRG vs. TNRC | TNSG vs. TNSC | Protein Accession ID | Description | ||||
|---|---|---|---|---|---|---|---|---|---|
| LogFC | FDR | LogFC | FDR | LogFC | FDR | ||||
| Up-regulated in TNR | TRINITY_DN4887_c0_g1 | 7.6 | 0 | 0.7 | 2.9 × 10−65 | 0.5 | 5.5 × 10−1 | XP_023740558.1 | Protein LURP-1-related 10-like |
| TRINITY_DN535_c0_g1 | 5.1 | 6.6 × 10−83 | 5.2 | 0 | 4.9 | 2.2 × 10−73 | PWA57088.1 | Cytochrome P450 | |
| TRINITY_DN5825_c3_g2 | 4.8 | 0 | 0.3 | 1.5 × 10−28 | 0.8 | 1.2 × 10−6 | PWA99700.1 | Armadillo-type fold | |
| TRINITY_DN5327_c0_g1 | 4.2 | 8.2 × 10−57 | 1.1 | 1.0 × 10−18 | 5.6 | 1.1 × 10−180 | KVH87673.1 | Ankyrin repeat-containing protein | |
| TRINITY_DN15014_c0_g1 | 4.2 | 2.8 × 10−89 | 0.4 | 0 | 0.2 | 8.5 × 10−1 | XP_023756443.1 | Uncharacterized protein LOC111904992 | |
| TRINITY_DN12399_c0_g1 | 3.7 | 3.0 × 10−39 | 0.7 | 1.6 × 10−5 | 1.2 | 1.6 × 10−2 | PWA74669.1 | Glutamyl-tRNA(Gln) amidotransferase | |
| TRINITY_DN3064_c0_g2 | 3.4 | 9.3 × 10−53 | 0.2 | 0.147264 | −0.2 | 6.6 × 10−1 | OWM86275.1 | Hypothetical protein CDL15_Pgr011099 | |
| TRINITY_DN4900_c1_g1 | 2.8 | 1.9 × 10−60 | 0.9 | 1.3 × 10−16 | 0.8 | 4.8 × 10−4 | PLY78555.1 | Hypothetical protein LSAT_1X84440 | |
| TRINITY_DN9714_c0_g1 | 2.6 | 4.5 × 10−26 | 2.1 | 3.0 × 10−66 | 3.4 | 1.6 × 10−54 | PWA81921.1 | O-methyltransferase, COMT-type | |
| TRINITY_DN830_c0_g2 | 2.3 | 0 | 1.5 | 0 | 0.3 | 4.0 × 10−160 | RCW19059.1 | Hypothetical protein GLYMA_13G018000 | |
| Up-regulated in TNS | TRINITY_DN3422_c0_g1 | −9.5 | 0 | n.a. | n.a. | −0.5 | 5.4 × 10−16 | PWA34949.1 | Ribonuclease H-like domain-containing protein |
| TRINITY_DN16145_c0_g1 | −9.5 | 5.3 × 10−151 | n.a. | n.a. | −0.1 | 5.9 × 10−1 | PWA56637.1 | RNA-directed DNA polymerase | |
| TRINITY_DN10659_c0_g1 | −9.5 | 2.2 × 10−159 | n.a. | n.a. | −0.2 | 1.2 × 10−2 | XP_018725631.1 | CHROMATIN REMODELING 24 | |
| TRINITY_DN10231_c0_g1 | −9.3 | 0 | n.a. | n.a. | −2.1 | 4.6 × 10−115 | XP_023748705.1 | Translocase of chloroplast 34, chloroplastic | |
| TRINITY_DN6562_c0_g1 | −9.1 | 1.0 × 10−276 | n.a. | n.a. | −0.6 | 9.2 × 10−15 | XP_021985853.1 | Uncharacterized protein LOC110882061 | |
| TRINITY_DN1776_c0_g1 | −8.8 | 0 | n.a. | n.a. | −0.1 | 8.1 × 10−2 | PLY61944.1 | Hypothetical protein LSAT_5X69860 | |
| TRINITY_DN11878_c0_g1 | −8.7 | 5.9 × 10−219 | n.a. | n.a. | 0.9 | 1.1 × 10−45 | XP_024984807.1 | FACT complex subunit SPT16 isoform X1 | |
| TRINITY_DN65613_c0_g1 | −8.7 | 2.2 × 10−211 | n.a. | n.a. | −0.7 | 4.6 × 10−14 | XP_021992293.1 | Uncharacterized protein LOC110889094 | |
| TRINITY_DN9729_c0_g1 | −8.6 | 4.1 × 10−217 | n.a. | n.a. | 0.2 | 2.0 × 10−2 | OTF97831.1 | Mitogen-activated protein (MAP) kinase kinase | |
| TRINITY_DN8641_c0_g1 | −8.6 | 1.4 × 10−87 | n.a. | n.a. | 0.6 | 2.3 × 10−9 | XP_022025290.1 | DNA-directed RNA polymerase | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Gardner, C.; Gupta, P.; Peng, Y.; Piasecki, C.; Millwood, R.J.; Ahn, T.-H.; Stewart, C.N., Jr. Novel Candidate Genes Differentially Expressed in Glyphosate-Treated Horseweed (Conyza canadensis). Genes 2021, 12, 1616. https://doi.org/10.3390/genes12101616
Yang Y, Gardner C, Gupta P, Peng Y, Piasecki C, Millwood RJ, Ahn T-H, Stewart CN Jr. Novel Candidate Genes Differentially Expressed in Glyphosate-Treated Horseweed (Conyza canadensis). Genes. 2021; 12(10):1616. https://doi.org/10.3390/genes12101616
Chicago/Turabian StyleYang, Yongil, Cory Gardner, Pallavi Gupta, Yanhui Peng, Cristiano Piasecki, Reginald J. Millwood, Tae-Hyuk Ahn, and C. Neal Stewart, Jr. 2021. "Novel Candidate Genes Differentially Expressed in Glyphosate-Treated Horseweed (Conyza canadensis)" Genes 12, no. 10: 1616. https://doi.org/10.3390/genes12101616
APA StyleYang, Y., Gardner, C., Gupta, P., Peng, Y., Piasecki, C., Millwood, R. J., Ahn, T.-H., & Stewart, C. N., Jr. (2021). Novel Candidate Genes Differentially Expressed in Glyphosate-Treated Horseweed (Conyza canadensis). Genes, 12(10), 1616. https://doi.org/10.3390/genes12101616