
Identification and Comprehensive Structural and Functional Analyses of the EXO70 Gene Family in Cotton

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of EXO70 Family Members in Cotton
2.2. Phylogeny, Gene Structure, Conserved Domains, and Motif Analysis
2.3. Expression Pattern Analysis
2.4. Cloning of GhEXO70A1-A Gene
2.5. Real-Time Quantitative PCR (RT-qPCR) Analysis
2.6. Subcellular Localization Detection of GhEXO70A1-A
2.7. Yeast Double-Hybrid Assay
2.8. Virus-Induced GhEXO70A1-A Gene Silencing
2.9. Transcriptome Analysis
3. Results
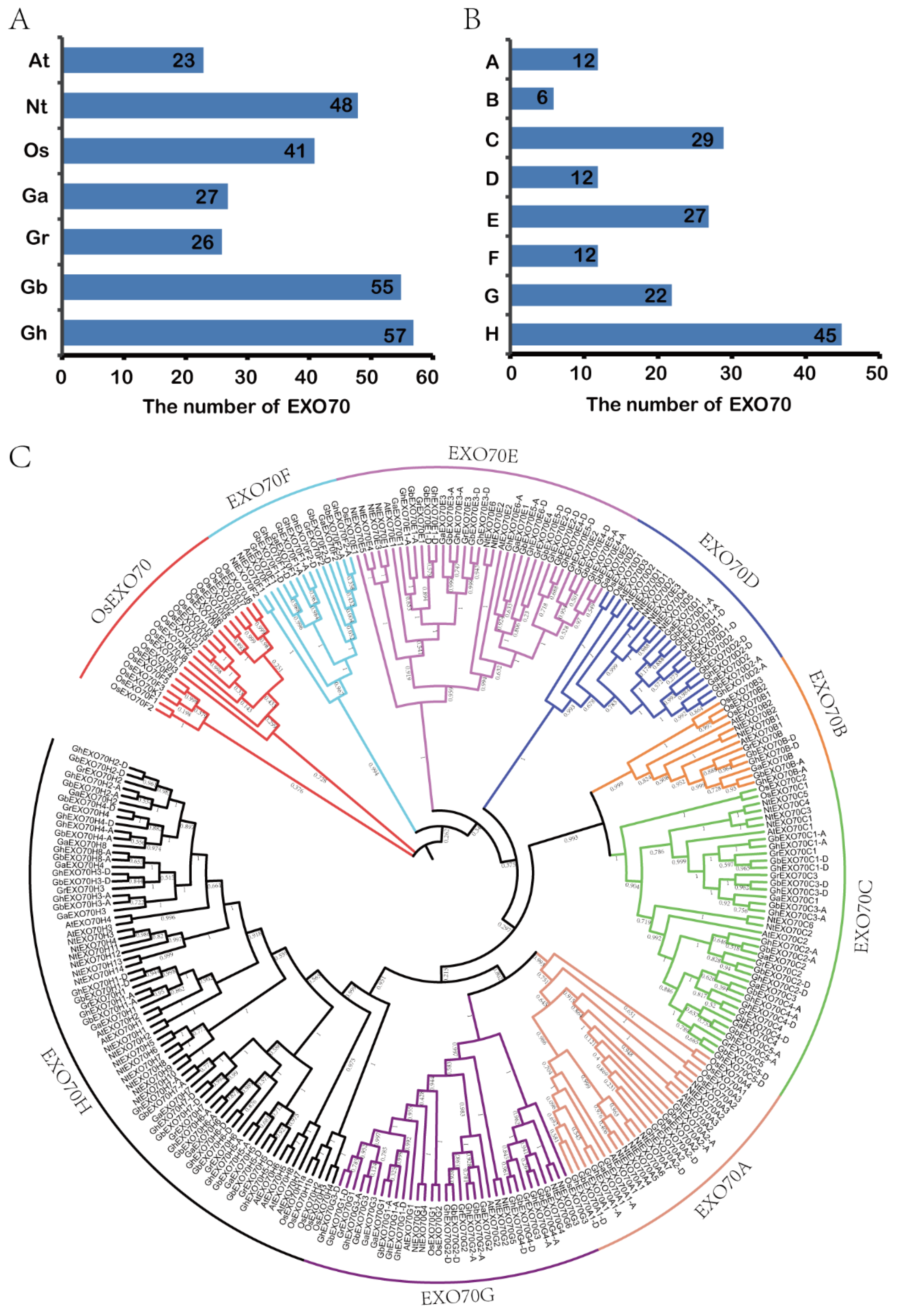
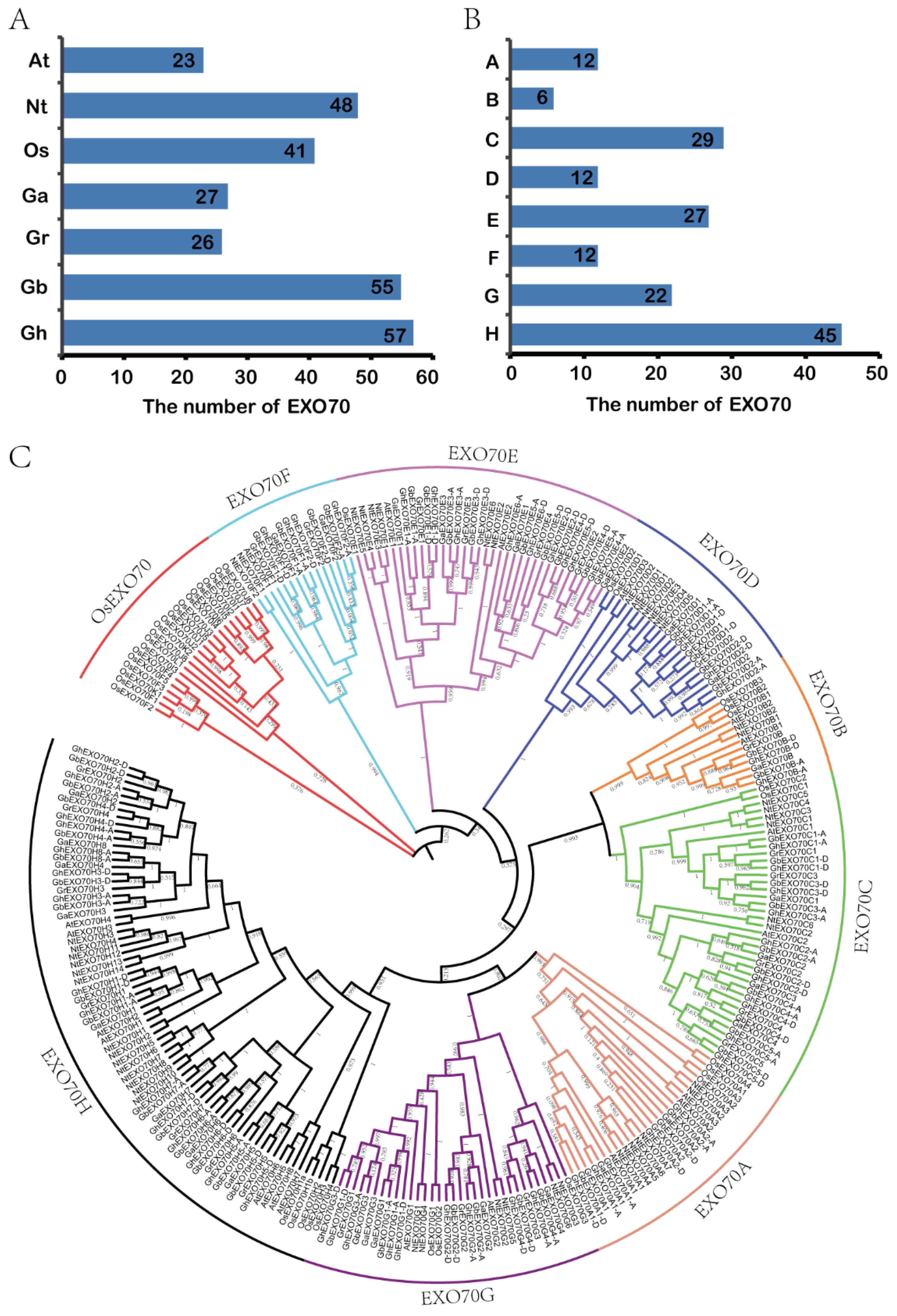
3.1. Identification and Analysis of the Phylogenetic Relationship of the EXO70 Gene Family in Cotton
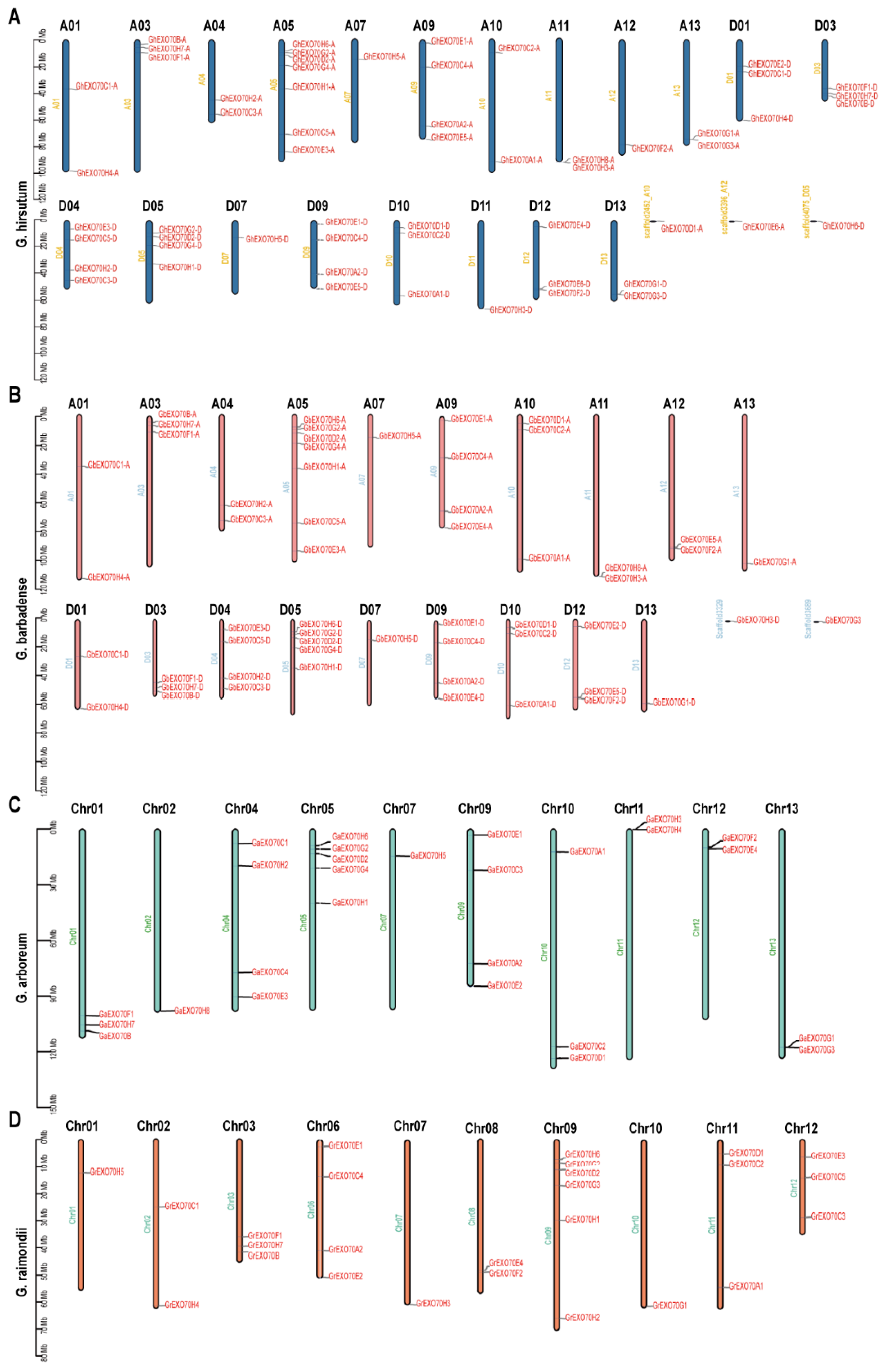
3.2. Chromosome Distribution Analysis of EXO70 in the Cotton Genome
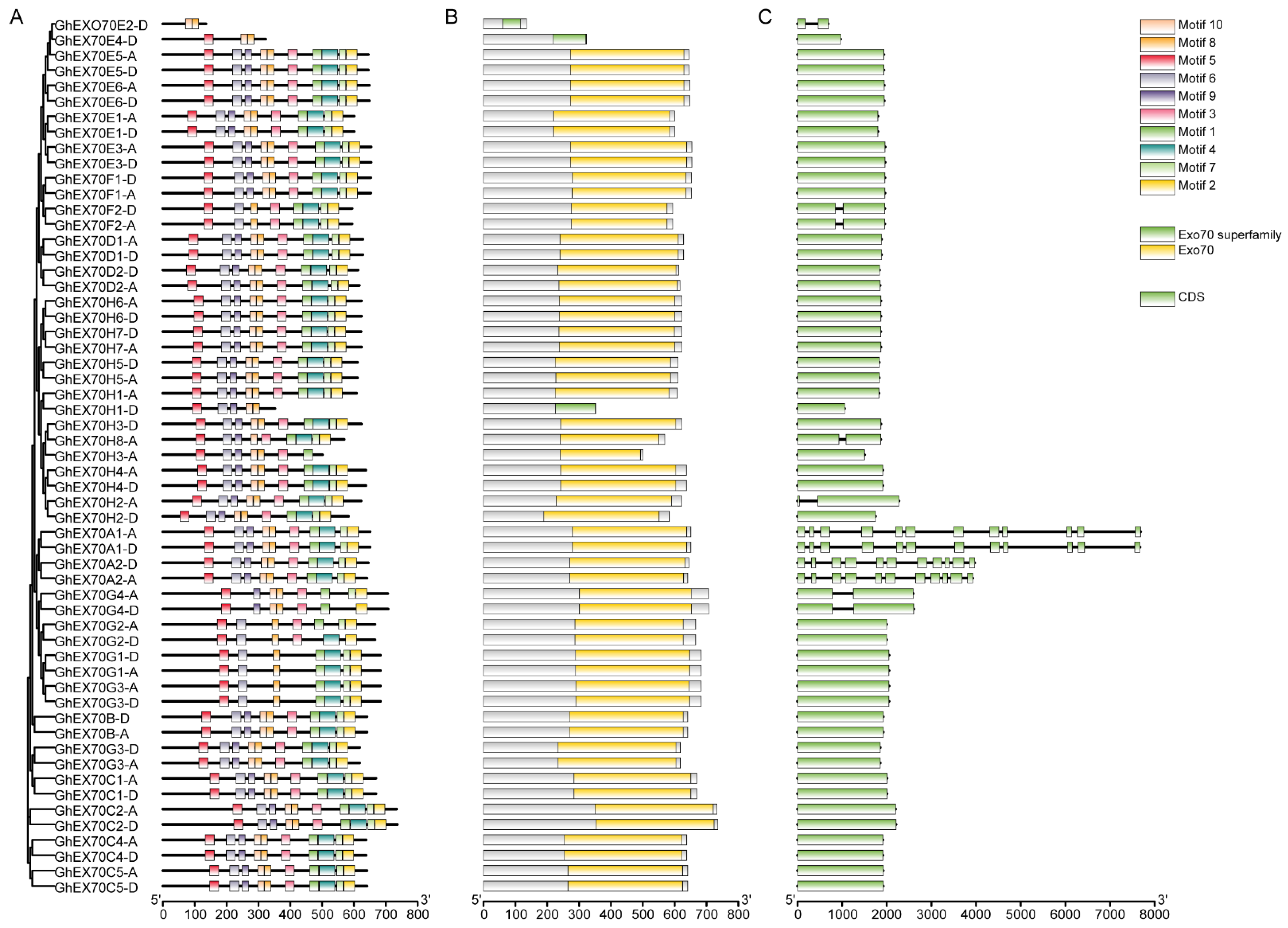
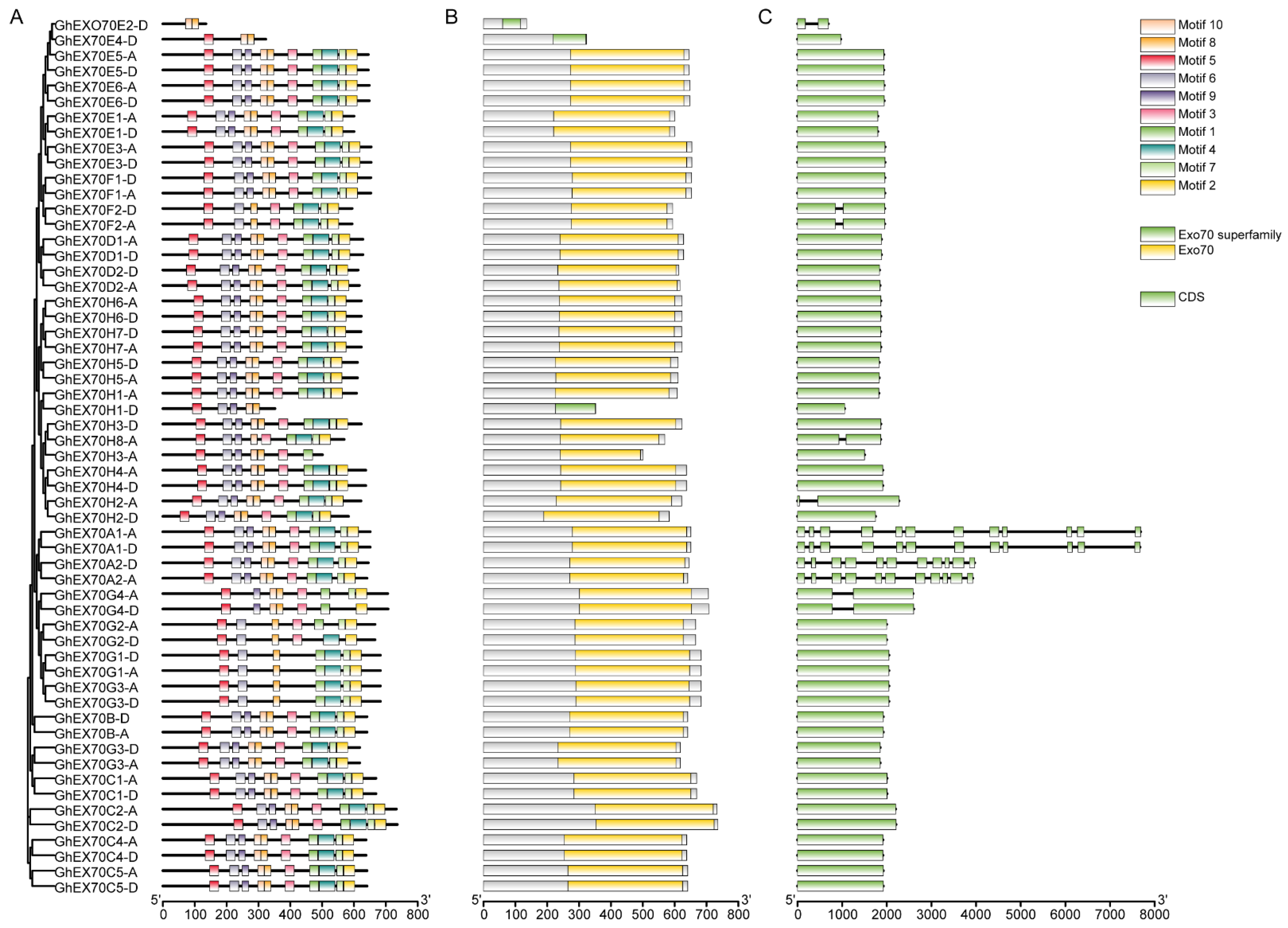
3.3. Analysis of EXO70 Gene Structure in G. hirsutum
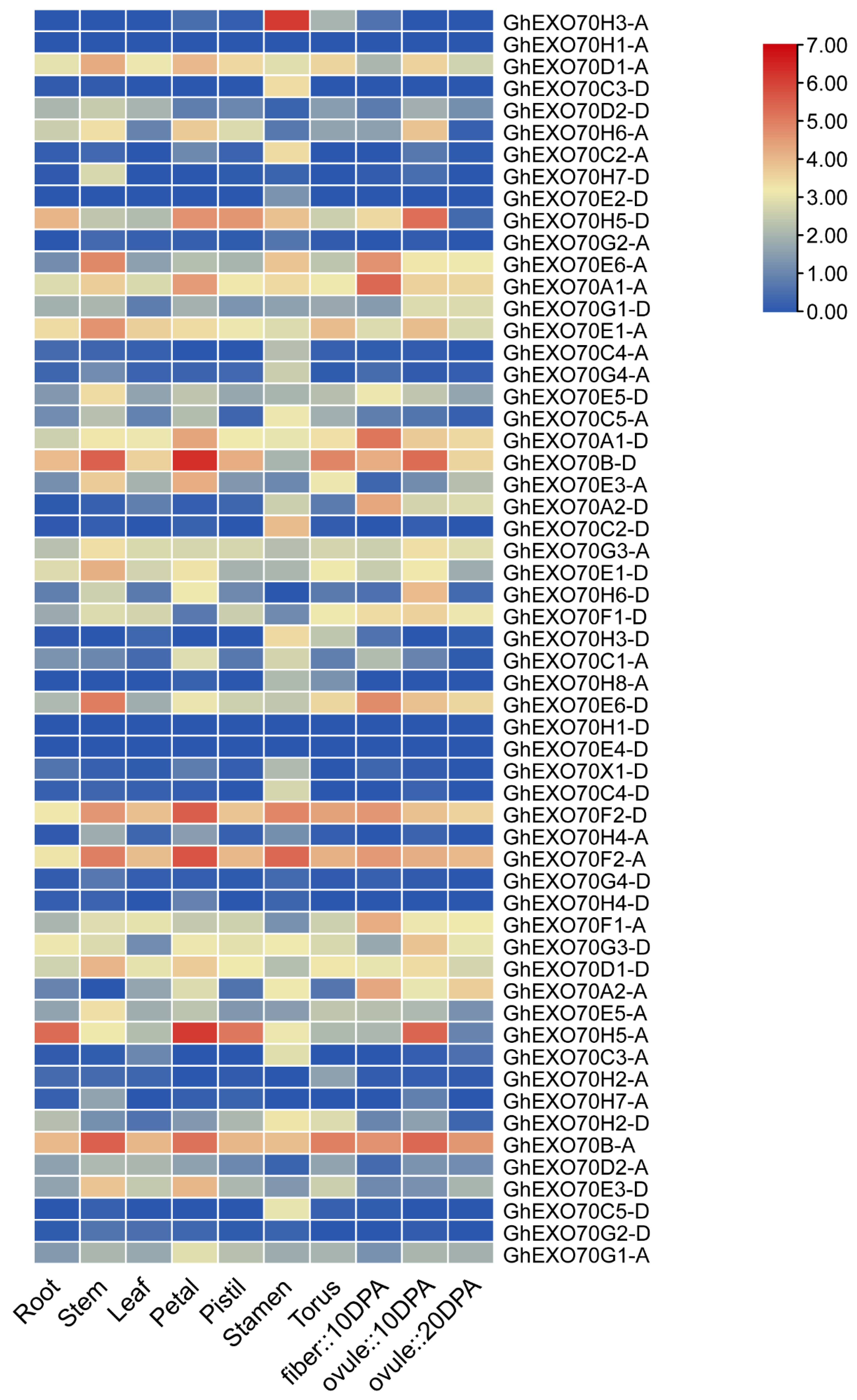
3.4. Analysis of EXO70 Gene Expression Patterns in G. arboretum and G. hirsutum
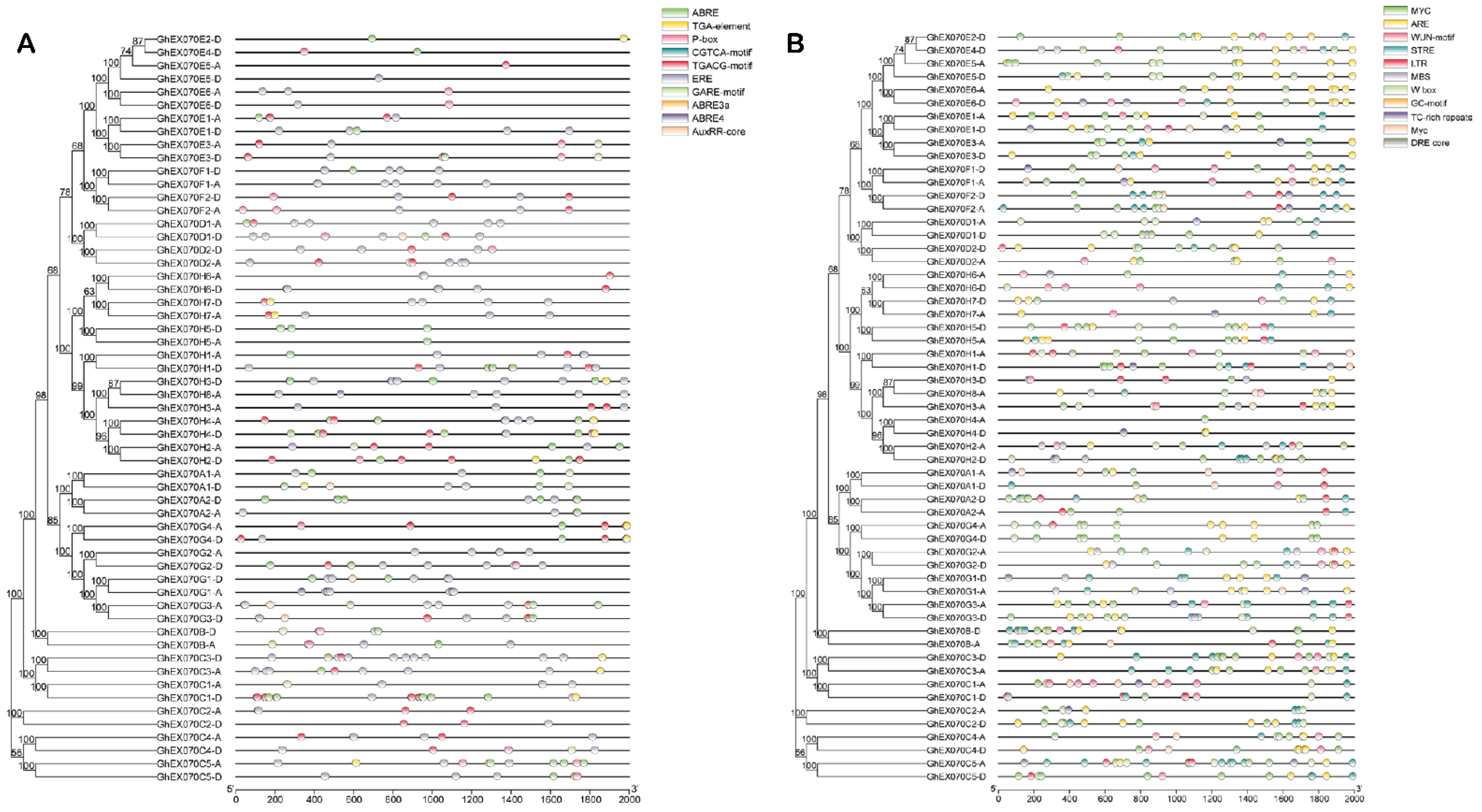
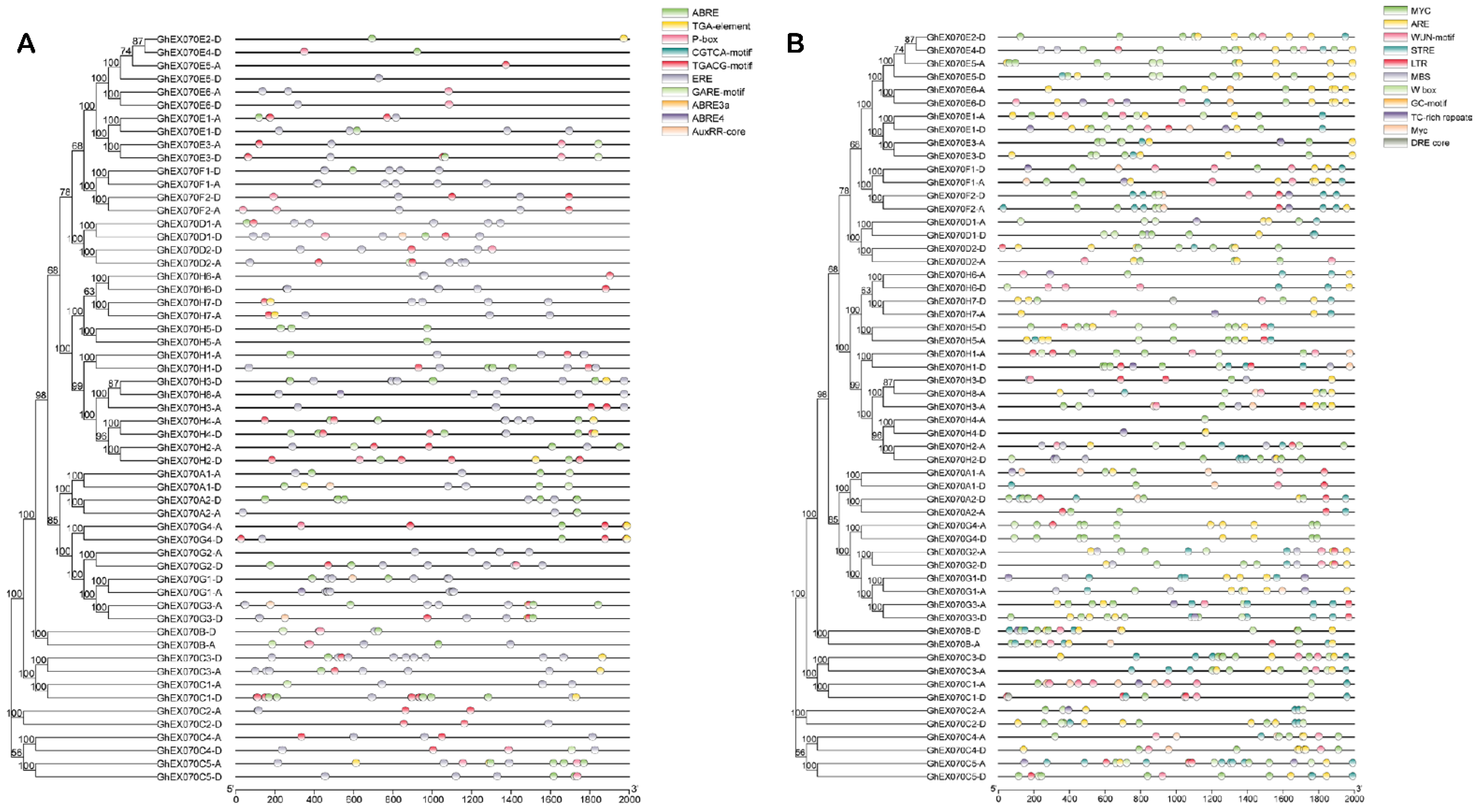
3.5. EXO70 Gene Transcription Regulation Analysis
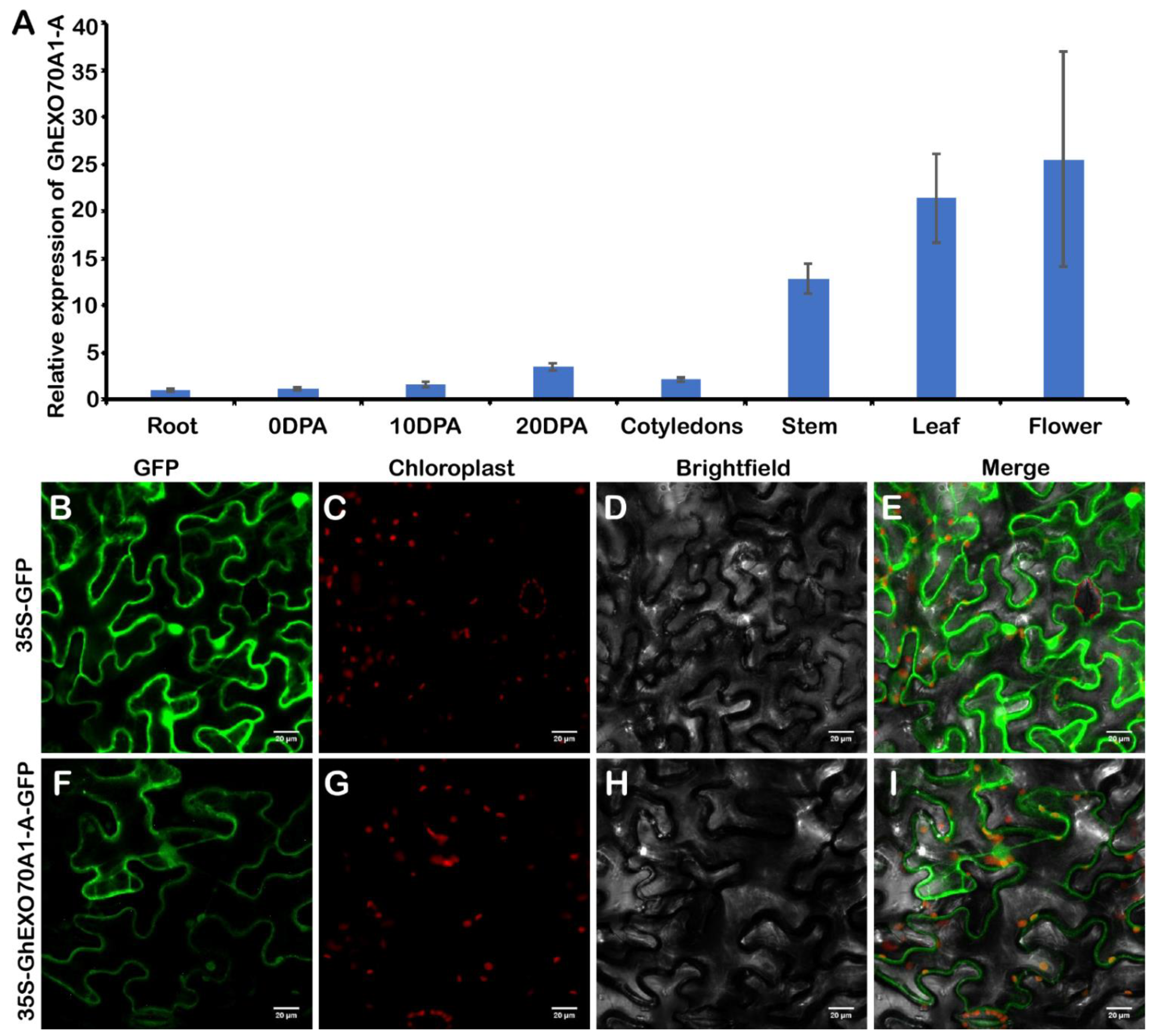
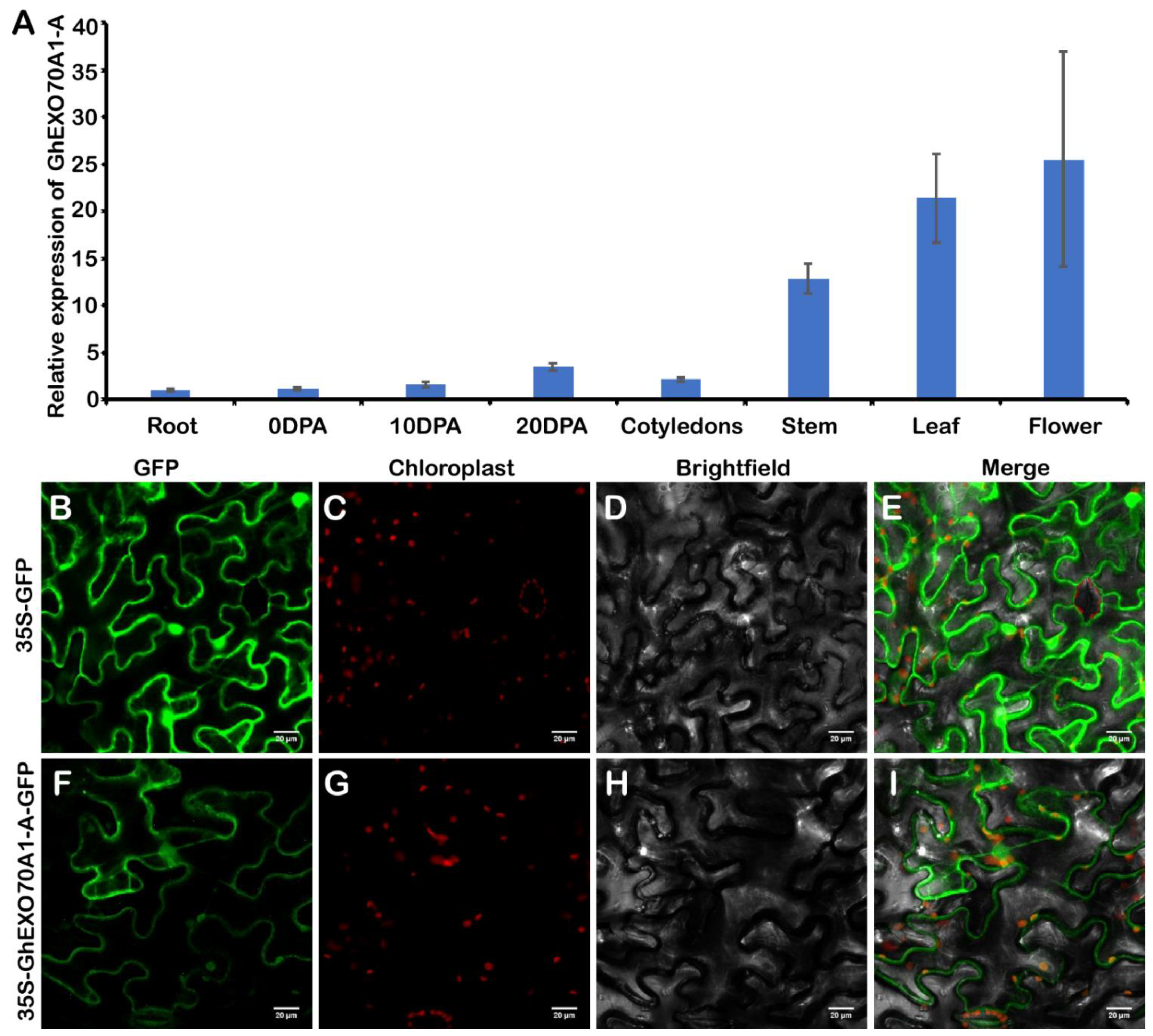
3.6. Expression Analysis and Subcellular Location of GhEXO70A1-A
3.7. GhEXO70A1-A Protein Interaction Analysis
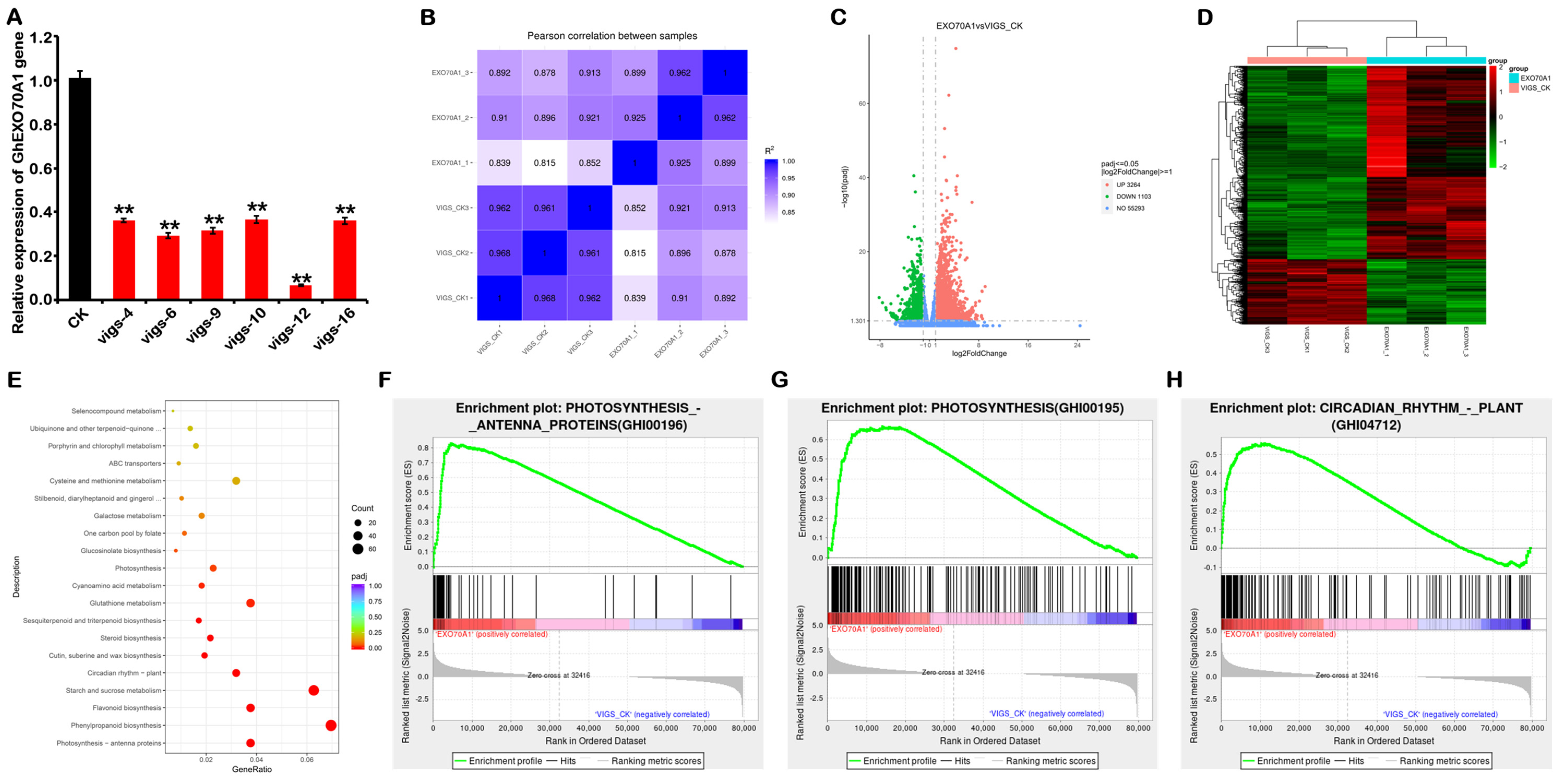
3.8. VIGS Silencing of GhEXO70A1-A Causes Changes in Signaling Pathways and Gene Expression
4. Discussion
4.1. Evolutionary Relationships of the Cotton EXO70 Gene Family
4.2. Biological Processes Implicating GhEXO70A1-A
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonifacino, J.S.; Glick, B.S. The mechanisms of vesicle budding and fusion. Cell 2004, 116, 153–166. [Google Scholar] [CrossRef] [Green Version]
- Whyte, J.R.; Munro, S. Vesicle tethering complexes in membrane traffic. J. Cell Sci. 2002, 115, 2627–2637. [Google Scholar] [CrossRef] [PubMed]
- Munson, M.; Novick, P. The exocyst defrocked, a framework of rods revealed. Nat. Struct. Mol. Biol. 2006, 13, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.C.; Ting, A.E.; Hazuka, C.D.; Davanger, S.; Kenny, J.W.; Kee, Y.; Scheller, R.H. The mammalian brain rsec6/8 complex. Neuron 1996, 17, 1209–1219. [Google Scholar] [CrossRef] [Green Version]
- TerBush, D.R.; Maurice, T.; Roth, D.; Novick, P. The Exocyst is a multiprotein complex required for exocytosis in Saccharomyces cerevisiae. EMBO J. 1996, 15, 6483–6494. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Guo, W. The exocyst complex in polarized exocytosis. Curr. Opin. Cell Biol. 2009, 21, 537–542. [Google Scholar] [CrossRef] [Green Version]
- Sivaram, M.V.; Saporita, J.A.; Furgason, M.L.; Boettcher, A.J.; Munson, M. Dimerization of the exocyst protein Sec6p and its interaction with the t-SNARE Sec9p. Biochemistry 2005, 44, 6302–6311. [Google Scholar] [CrossRef]
- Wu, H.; Turner, C.; Gardner, J.; Temple, B.; Brennwald, P. The Exo70 subunit of the exocyst is an effector for both Cdc42 and Rho3 function in polarized exocytosis. Mol. Biol. Cell 2010, 21, 430–442. [Google Scholar] [CrossRef] [Green Version]
- Elias, M.; Drdova, E.; Ziak, D.; Bavlnka, B.; Hala, M.; Cvrckova, F.; Soukupova, H.; Zarsky, V. The exocyst complex in plants. Cell. Biol. Int. 2003, 27, 199–201. [Google Scholar] [CrossRef]
- Synek, L.; Schlager, N.; Elias, M.; Quentin, M.; Hauser, M.T.; Zarsky, V. AtEXO70A1, a member of a family of putative exocyst subunits specifically expanded in land plants, is important for polar growth and plant development. Plant J. 2006, 48, 54–72. [Google Scholar] [CrossRef] [Green Version]
- Cvrckova, F.; Grunt, M.; Bezvoda, R.; Hala, M.; Kulich, I.; Rawat, A.; Zarsky, V. Evolution of the land plant exocyst complexes. Front. Plant Sci. 2012, 3, 159. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; van Os, G.M.; Ren, S.; Yu, D.; Ketelaar, T.; Emons, A.M.; Liu, C.M. Expression and functional analyses of EXO70 genes in Arabidopsis implicate their roles in regulating cell type-specific exocytosis. Plant Physiol. 2010, 154, 1819–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, X.; Zhang, J.; Zhang, Y.; Hsu, S.C.; Zhou, D.; Guo, W. Exo70 interacts with the Arp2/3 complex and regulates cell migration. Nat. Cell Biol. 2006, 8, 1383–1388. [Google Scholar] [CrossRef]
- Dupraz, S.; Grassi, D.; Bernis, M.E.; Sosa, L.; Bisbal, M.; Gastaldi, L.; Jausoro, I.; Caceres, A.; Pfenninger, K.H.; Quiroga, S. The TC10-Exo70 complex is essential for membrane expansion and axonal specification in developing neurons. J. Neurosci. 2009, 29, 13292–13301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, X.; Xu, Q.; Huang, Y.; Singh, R.D.; Anderson, R.; Leof, E.; Hu, J.; Ling, K. An association between type Igamma PI4P 5-kinase and Exo70 directs E-cadherin clustering and epithelial polarization. Mol. Biol. Cell 2012, 23, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, J.; Yang, C.; Capraro, B.R.; Baumgart, T.; Bradley, R.P.; Ramakrishnan, N.; Xu, X.; Radhakrishnan, R.; Svitkina, T.; et al. Exo70 generates membrane curvature for morphogenesis and cell migration. Dev. Cell 2013, 26, 266–278. [Google Scholar] [CrossRef] [Green Version]
- Fujita, A.; Koinuma, S.; Yasuda, S.; Nagai, H.; Kamiguchi, H.; Wada, N.; Nakamura, T. GTP hydrolysis of TC10 promotes neurite outgrowth through exocytic fusion of Rab11- and L1-containing vesicles by releasing exocyst component Exo70. PLoS ONE 2013, 8, e79689. [Google Scholar] [CrossRef]
- Fendrych, M.; Synek, L.; Pecenkova, T.; Toupalova, H.; Cole, R.; Drdova, E.; Nebesarova, J.; Sedinova, M.; Hala, M.; Fowler, J.E.; et al. The Arabidopsis exocyst complex is involved in cytokinesis and cell plate maturation. Plant Cell 2010, 22, 3053–3065. [Google Scholar] [CrossRef] [Green Version]
- Pecenkova, T.; Hala, M.; Kulich, I.; Kocourkova, D.; Drdova, E.; Fendrych, M.; Toupalova, H.; Zarsky, V. The role for the exocyst complex subunits Exo70B2 and Exo70H1 in the plant-pathogen interaction. J. Exp. Bot. 2011, 62, 2107–2116. [Google Scholar] [CrossRef] [Green Version]
- Kulich, I.; Pecenkova, T.; Sekeres, J.; Smetana, O.; Fendrych, M.; Foissner, I.; Hoftberger, M.; Zarsky, V. Arabidopsis exocyst subcomplex containing subunit EXO70B1 is involved in autophagy-related transport to the vacuole. Traffic 2013, 14, 1155–1165. [Google Scholar] [CrossRef] [Green Version]
- Zarsky, V.; Kulich, I.; Fendrych, M.; Pecenkova, T. Exocyst complexes multiple functions in plant cells secretory pathways. Curr. Opin. Plant Biol. 2013, 16, 726–733. [Google Scholar] [CrossRef]
- Synek, L.; Vukasinovic, N.; Kulich, I.; Hala, M.; Aldorfova, K.; Fendrych, M.; Zarsky, V. EXO70C2 Is a Key Regulatory Factor for Optimal Tip Growth of Pollen. Plant Physiol. 2017, 174, 223–240. [Google Scholar] [CrossRef] [Green Version]
- Kulich, I.; Vojtikova, Z.; Sabol, P.; Ortmannova, J.; Nedela, V.; Tihlarikova, E.; Zarsky, V. Exocyst Subunit EXO70H4 Has a Specific Role in Callose Synthase Secretion and Silica Accumulation. Plant Physiol. 2018, 176, 2040–2051. [Google Scholar] [CrossRef] [Green Version]
- Hong, D.; Jeon, B.W.; Kim, S.Y.; Hwang, J.U.; Lee, Y. The ROP2-RIC7 pathway negatively regulates light-induced stomatal opening by inhibiting exocyst subunit Exo70B1 in Arabidopsis. New Phytol. 2016, 209, 624–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Liu, N.; Gao, C.; Cai, H.; Romeis, T.; Tang, D. The Arabidopsis exocyst subunits EXO70B1 and EXO70B2 regulate FLS2 homeostasis at the plasma membrane. New Phytol. 2020, 227, 529–544. [Google Scholar] [CrossRef] [PubMed]
- Acheampong, A.K.; Shanks, C.; Cheng, C.; Schaller, G.E.; Dagdas, Y.; Kieber, J.J. EXO70D isoforms mediate selective autophagic degradation of type-A ARR proteins to regulate cytokinin sensitivity. Proc. Natl. Acad. Sci. USA 2020, 117, 27034–27043. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, M.; Yu, D.; Ren, S.; Sun, S.; Liu, L.; Ketelaar, T.; Emons, A.M.; Liu, C.M. EXO70A1-mediated vesicle trafficking is critical for tracheary element development in Arabidopsis. Plant Cell 2013, 25, 1774–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, B.; Hu, L.; Chen, W.; Li, T.; Hu, B.; Zheng, L.; Lv, Z.; You, S.; Wang, Y.; Ma, B.; et al. Disruption of OsEXO70A1 Causes Irregular Vascular Bundles and Perturbs Mineral Nutrient Assimilation in Rice. Sci. Rep. 2015, 5, 18609. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Wang, N.; Zhang, T.; Zhang, Q.; Du, D.; Chen, X.; Lu, X.; Zhang, Y.; Zhu, M.; Liu, M.; et al. SHORT-ROOT 1 is critical to cell division and tracheary element development in rice roots. Plant J. 2021, 105, 1179–1191. [Google Scholar] [CrossRef]
- Dowd, K.M.; Pelitire, S.M.; Delhom, C.D. Molecular biology and physiology: Seed-fiber ratio, seed index, and seed tissue and compositional properties of current cotton cultivars. J. Cotton Sci. 2018, 22, 60–74. [Google Scholar]
- Yu, Q.; Hu, Z.; Shen, Y.; Jiang, Y.; Pan, P.; Hou, T.; Pan, Z.Q.; Huang, J.; Sun, Y. Gossypol inhibits cullin neddylation by targeting SAG-CUL5 and RBX1-CUL1 complexes. Neoplasia 2020, 22, 179–191. [Google Scholar] [CrossRef]
- Ding, Y.P.; Ge, X.Y.; Wang, P.; Wu, J.; Wang, S.F.; Li, F.G. Functional analysis of cotton salt stress response gene GhEXO70B1. J. Cotton Sci. 2018, 30, 4–15. [Google Scholar]
- Zhao, Y.; Guo, A.; Wang, Y.; Hua, J. Evolution of PEPC gene family in Gossypium reveals functional diversification and GhPEPC genes responding to abiotic stresses. Gene 2019, 698, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [PubMed]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 22DDCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chong, Y.T.; Gidda, S.K.; Sanford, C.; Parkinson, J.; Mullen, R.T.; Goring, D.R. Characterization of the Arabidopsis thaliana exocyst complex gene families by phylogenetic, expression profiling, and subcellular localization studies. New Phytol. 2010, 185, 401–419. [Google Scholar] [CrossRef]
- Yang, K.; Zhang, Y.; Lv, J.; Zhao, Y.B.; Zhang, H.C.; Hang, X.; He, G. EXO70 doubling in Arabidopsis and rice genomes. Chin. Sci. Bull. 2015, 1, 38–51. [Google Scholar]
- Zhao, J.; Zhang, X.; Wan, W.; Zhang, H.; Liu, J.; Li, M.; Wang, H.; Xiao, J.; Wang, X. Identification and Characterization of the EXO70 Gene Family in Polyploid Wheat and Related Species. Int. J. Mol. Sci. 2019, 20, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaharwar, A.K.; Singh, I.; Khademhosseini, A. Engineered biomaterials for in situ tissue regeneration. Nat. Rev. Mater. 2020, 5, 686–705. [Google Scholar] [CrossRef]
- Kulich, I.; Cole, R.; Drdová, E.; Cvrcková, F.; Soukup, A.; Fowler, J.; Zárský, V. Arabidopsis exocyst subunits SEC8 and EXO70A1 and exocyst interactor ROH1 are involved in the localized deposition of seed coat pectin. New Phytol. 2010, 188, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Kalmbach, L.; Hématy, K.; De Bellis, D.; Barberon, M.; Fujita, S.; Ursache, R.; Daraspe, J.; Geldner, N. Transient cell-specific EXO70A1 activity in the CASP domain and Casparian strip localization. Nat. Plants 2017, 3, 17058. [Google Scholar] [CrossRef]
- Vukašinović, N.; Oda, Y.; Pejchar, P.; Synek, L.; Pečenková, T.; Rawat, A.; Sekereš, J.; Potocký, M.; Žárský, V. Microtubule-dependent targeting of the exocyst complex is necessary for xylem development in Arabidopsis. New Phytol. 2017, 213, 1052–1067. [Google Scholar] [CrossRef]
- Chen, Z.J.; Sreedasyam, A.; Ando, A.; Song, Q.; Santiago, L.M.D.; Hulse-Kemp, A.M.; Ding, M.; Ye, W.; Kirkbride, R.C.; Jenkins, J.; et al. Genomic diversifications of five Gossypium allopolyploid species and their impact on cotton improvement. Nat. Genet. 2020, 52, 525–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc, G.; Wolfe, K.H. Functional divergence of duplicated genes formed by polyploidy during Arabidopsis evolution. Plant Cell 2004, 16, 1679–1691. [Google Scholar] [CrossRef] [Green Version]
- Synek, L.; Pleskot, R.; Sekereš, J.; Serrano, N.; Vukašinović, N.; Ortmannová, J.; Klejchová, M.; Pejchar, P.; Batystová, K.; Gutkowska, M.; et al. Plasma membrane phospholipid signature recruits the plant exocyst complex via the EXO70A1 subunit. Proc. Natl. Acad. Sci. USA 2021, 118, e2105287118. [Google Scholar] [CrossRef]
- Paponov, I.A.; Papono, M.; Teale, W.; Menges, M.; Chakrabortee, S.; Murray, J.A.H.; Palme, K. Comprehensive transcriptome analysis of auxin responses in Arabidopsis. Mol. Plant 2008, 1, 321–337. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.Q.; Chen, D.K.; Li, P.P.; Bao, H.N.; Liu, H.Z.; Yin, J.; Zeng, H.Y.; Yang, Y.B.; Li, Y.K.; Xiao, S. Jasmonates modulate sphingolipid metabolism and accelerate cell death in the ceramide kinase mutant acd5. Plant Physiol. 2021, kiab362. [Google Scholar] [CrossRef] [PubMed]
- Drdová, E.J.; Synek, L.; Pečenková, T.; Hála, M.; Kulich, I.; Fowler, J.E.; Murphy, A.S.; Žárský, V. The exocyst complex contributes to PIN auxin efflux carrier recycling and polar auxin transport in Arabidopsis. Plant J. 2013, 73, 709–719. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Name | Chromosome | Start | End | Exon Number | Protein Length (aa) | Molecular Weight (kDa) | Isoelectric Point | Subcellular Location |
|---|---|---|---|---|---|---|---|---|---|
| Gh_A10G1765 | GhEXO70A1-A | A10 | 92,079,304 | 92,086,988 | 12 | 650 | 73.432 | 8.308 | Cell membrane |
| Gh_D10G2039 | GhEXO70A1-D | D10 | 56,152,841 | 56,160,513 | 12 | 650 | 73.429 | 8.131 | Cell membrane |
| Gh_A09G1270 | GhEXO70A2-A | A09 | 64,875,878 | 64,879,803 | 11 | 640 | 72.863 | 8.901 | Cell membrane |
| Gh_D09G1272 | GhEXO70A2-D | D09 | 39,824,276 | 39,828,245 | 11 | 644 | 73.467 | 9.329 | Cell membrane, cytoplasm |
| Gh_A03G0212 | GhEXO70B-A | A03 | 3,226,810 | 3,228,732 | 1 | 640 | 72.955 | 4.951 | Cell membrane, cytoplasm |
| Gh_D03G1369 | GhEXO70B-D | D03 | 42,280,554 | 42,282,476 | 1 | 640 | 72.962 | 4.914 | Cell membrane, cytoplasm |
| Gh_A01G1064 | GhEXO70C1-A | A01 | 36,760,999 | 36,763,005 | 1 | 668 | 77.345 | 8.781 | Cell membrane, cytoplasm |
| Gh_D01G1124 | GhEXO70C1-D | D01 | 23,935,803 | 23,937,809 | 1 | 668 | 76.941 | 8.482 | Cell membrane, cytoplasm |
| Gh_A10G0625 | GhEXO70C2-A | A10 | 9,989,979 | 9,992,177 | 1 | 732 | 84.71 | 4.555 | Nucleus |
| Gh_D10G0774 | GhEXO70C2-D | D10 | 9,213,316 | 9,215,523 | 1 | 735 | 85.065 | 4.537 | Nucleus |
| Gh_A04G0860 | GhEXO70C3-A | A04 | 55,909,662 | 55,911,515 | 1 | 617 | 70.763 | 4.975 | Cell membrane, cytoplasm |
| Gh_D04G1359 | GhEXO70C3-D | D04 | 44,235,768 | 44,237,621 | 1 | 617 | 70.799 | 5.024 | Cell membrane, cytoplasm |
| Gh_A09G0369 | GhEXO70C4-A | A09 | 20,318,531 | 20,320,444 | 1 | 637 | 73.612 | 5.53 | Cell membrane, cytoplasm |
| Gh_D09G0388 | GhEXO70C4-D | D09 | 14,114,761 | 14,116,674 | 1 | 637 | 73.682 | 5.35 | Cell membrane, cytoplasm |
| Gh_A05G2929 | GhEXO70C5-A | A05 | 70,962,860 | 70,964,782 | 1 | 640 | 73.595 | 6.664 | Cell membrane, cytoplasm |
| Gh_D04G0713 | GhEXO70C5-D | D04 | 14,452,693 | 14,454,615 | 1 | 640 | 73.566 | 6.384 | Cell membrane, cytoplasm |
| Gh_A10G2233 | GhEXO70D1-A | scaffold2452_A10 | 2396 | 4279 | 1 | 627 | 71.136 | 5.36 | Cell membrane, cytoplasm |
| Gh_D10G0529 | GhEXO70D1-D | D10 | 5,107,684 | 5,109,567 | 1 | 627 | 71.084 | 5.278 | Cell membrane, cytoplasm |
| Gh_A05G1157 | GhEXO70D2-A | A05 | 11,706,409 | 11,708,259 | 1 | 616 | 69.599 | 5.35 | Cell membrane |
| Gh_D05G1334 | GhEXO70D2-D | D05 | 11,742,012 | 11,743,850 | 1 | 612 | 69.264 | 4.972 | Cell membrane, cytoplasm |
| Gh_A09G0090 | GhEXO70E1-A | A09 | 2,303,006 | 2,304,805 | 1 | 599 | 69.086 | 4.988 | Cell membrane, cytoplasm |
| Gh_D09G0087 | GhEXO70E1-D | D09 | 2,312,361 | 2,314,160 | 1 | 599 | 69.308 | 5.089 | Cell membrane, cytoplasm |
| Gh_D01G1051 | GhEXO70E2-D | D01 | 19,584,687 | 19,585,379 | 2 | 134 | 15.208 | 6.674 | Cell membrane |
| Gh_A05G3215 | GhEXO70E3-A | A05 | 84,043,718 | 84,045,679 | 1 | 653 | 74.613 | 4.761 | Cell membrane |
| Gh_D04G0392 | GhEXO70E3-D | D04 | 6,206,299 | 6,208,260 | 1 | 653 | 74.511 | 4.731 | Cell membrane |
| Gh_D12G0327 | GhEXO70E4-D | D12 | 4,666,459 | 4,667,427 | 1 | 322 | 36.173 | 4.875 | Cell membrane |
| Gh_A09G2154 | GhEXO70E5-A | A09 | 74,578,905 | 74,580,839 | 1 | 644 | 73.351 | 5.784 | Cell membrane, cytoplasm |
| Gh_D09G2359 | GhEXO70E5-D | D09 | 50,553,799 | 50,555,733 | 1 | 644 | 73.402 | 6.24 | Cell membrane, cytoplasm |
| Gh_A12G2651 | GhEXO70E6-A | scaffold3396_A12 | 4667 | 6610 | 1 | 647 | 73.345 | 5.788 | Cell membrane |
| Gh_D12G1810 | GhEXO70E6-D | D12 | 50,610,382 | 50,612,325 | 1 | 647 | 73.122 | 5.417 | Cell membrane |
| Gh_A03G0449 | GhEXO70F1-A | A03 | 9,703,926 | 9,705,884 | 1 | 652 | 73.754 | 4.614 | Cell membrane, cytoplasm |
| Gh_D03G1089 | GhEXO70F1-D | D03 | 36,373,153 | 36,375,111 | 1 | 652 | 73.79 | 4.587 | Cell membrane |
| Gh_A12G1712 | GhEXO70F2-A | A12 | 78,884,906 | 78,886,861 | 2 | 593 | 67.361 | 4.566 | Cell membrane |
| Gh_D12G1873 | GhEXO70F2-D | D12 | 51,339,965 | 51,341,920 | 2 | 593 | 67.36 | 4.589 | Cell membrane, cytoplasm |
| Gh_A13G1576 | GhEXO70G1-A | A13 | 74,625,245 | 74,627,293 | 1 | 682 | 77.054 | 8.387 | Cell membrane, cytoplasm |
| Gh_D13G1935 | GhEXO70G1-D | D13 | 54,742,538 | 54,744,586 | 1 | 682 | 76.932 | 8.152 | Cell membrane, cytoplasm |
| Gh_A05G0971 | GhEXO70G2-A | A05 | 9,685,126 | 9,687,123 | 1 | 665 | 74.839 | 6.629 | Cell membrane, nucleus |
| Gh_D05G1080 | GhEXO70G2-D | D05 | 9,208,639 | 9,210,636 | 1 | 665 | 74.79 | 6.457 | Cell membrane, nucleus |
| Gh_A13G1577 | GhEXO70G3-A | A13 | 74,629,925 | 74,631,973 | 1 | 682 | 77.006 | 8.013 | Cell membrane, cytoplasm |
| Gh_D13G1936 | GhEXO70G3-D | D13 | 54,747,144 | 54,749,192 | 1 | 682 | 77.129 | 8.008 | Cell membrane, cytoplasm |
| Gh_A05G1829 | GhEXO70G4-A | A05 | 19,148,304 | 19,150,892 | 2 | 705 | 80.991 | 6.269 | Cell membrane, nucleus |
| Gh_D05G2026 | GhEXO70G4-D | D05 | 18,583,197 | 18,585,799 | 2 | 706 | 81.133 | 6.088 | Cell membrane |
| Gh_A05G2577 | GhEXO70H1-A | A05 | 36,616,771 | 36,618,594 | 1 | 607 | 68.055 | 7.626 | Cell membrane |
| Gh_D05G2864 | GhEXO70H1-D | D05 | 32,263,469 | 32,264,524 | 1 | 351 | 39.036 | 8.216 | Cell membrane, nucleus |
| Gh_A04G0671 | GhEXO70H2-A | A04 | 45,359,837 | 45,362,111 | 2 | 621 | 70.248 | 5.721 | Cell membrane, cytoplasm |
| Gh_D04G1136 | GhEXO70H2-D | D04 | 37,216,953 | 37,218,701 | 1 | 582 | 65.703 | 5.697 | Cell membrane, cytoplasm |
| Gh_A11G2905 | GhEXO70H3-A | A11 | 92,971,783 | 92,973,285 | 1 | 500 | 56.067 | 7.521 | Cell membrane, nucleus |
| Gh_D11G3290 | GhEXO70H3-D | D11 | 65,820,199 | 65,822,067 | 1 | 622 | 69.832 | 7.178 | Cell membrane, cytoplasm |
| Gh_A01G1870 | GhEXO70H4-A | A01 | 98,692,379 | 98,694,289 | 1 | 636 | 71.827 | 7.493 | Cell membrane, cytoplasm |
| Gh_D01G2127 | GhEXO70H4-D | D01 | 60,327,492 | 60,329,402 | 1 | 636 | 72.148 | 7.783 | Cell membrane, cytoplasm |
| Gh_A07G0865 | GhEXO70H5-A | A07 | 15,194,291 | 15,196,123 | 1 | 610 | 69.021 | 6.068 | Cell membrane, cytoplasm |
| Gh_D07G0937 | GhEXO70H5-D | D07 | 12,444,178 | 12,446,010 | 1 | 610 | 68.925 | 6.316 | Cell membrane, cytoplasm |
| Gh_A05G0839 | GhEXO70H6-A | A05 | 8,379,960 | 8,381,828 | 1 | 622 | 70.171 | 5.757 | Cell membrane |
| Gh_D05G3898 | GhEXO70H6-D | scaffold4075_D05 | 141,419 | 143,287 | 1 | 622 | 70.049 | 5.361 | Cell membrane, cytoplasm |
| Gh_A03G0316 | GhEXO70H7-A | A03 | 5,680,420 | 5,682,288 | 1 | 622 | 70.772 | 5.209 | Cell membrane, cytoplasm |
| Gh_D03G1262 | GhEXO70H7-D | D03 | 40,093,224 | 40,095,089 | 1 | 621 | 70.603 | 5.889 | Cell membrane, cytoplasm |
| Gh_A11G2904 | GhEXO70H8-A | A11 | 92,966,319 | 92,968,183 | 2 | 568 | 63.859 | 8.556 | Cell membrane, cytoplasm |
| Chromosome | Ga (27) | Gr (26) | Gb (55) | Gh (57) | Total | ||
|---|---|---|---|---|---|---|---|
| A | D | A | D | A | D | ||
| Chr.1 | 3 | 1 | 2 | 2 | 2 | 3 | 13 |
| Chr.2 | 1 | 2 | 0 | 0 | 0 | 0 | 3 |
| Chr.3 | 0 | 3 | 3 | 3 | 3 | 3 | 15 |
| Chr.4 | 4 | 0 | 2 | 4 | 2 | 4 | 16 |
| Chr.5 | 5 | 0 | 7 | 5 | 7 | 4 | 28 |
| Chr.6 | 0 | 4 | 0 | 0 | 0 | 0 | 4 |
| Chr.7 | 1 | 1 | 1 | 1 | 1 | 1 | 6 |
| Chr.8 | 0 | 2 | 0 | 0 | 0 | 0 | 2 |
| Chr.9 | 4 | 6 | 4 | 4 | 4 | 4 | 26 |
| Chr.10 | 3 | 1 | 3 | 3 | 2 | 3 | 15 |
| Chr.11 | 2 | 3 | 2 | 0 | 2 | 1 | 10 |
| Chr.12 | 2 | 3 | 2 | 3 | 1 | 3 | 14 |
| Chr.13 | 2 | 0 | 1 | 1 | 2 | 2 | 8 |
| total | 27 | 26 | 27 | 26 | 26 | 28 | 160 |
| unknown | 0 | 0 | 1 | 1 | 2 | 1 | 5 |
| Subgroup | G. arboretum | G. raimondii | G. barbadense | G. hirsutum |
|---|---|---|---|---|
| A | 2 | 2 | 4 | 4 |
| B | 1 | 1 | 2 | 2 |
| C | 4 | 5 | 10 | 10 |
| D | 2 | 2 | 4 | 4 |
| E | 4 | 4 | 9 | 10 |
| F | 2 | 2 | 4 | 4 |
| G | 4 | 3 | 7 | 8 |
| H | 8 | 7 | 15 | 15 |
| KEGG ID | Description | Gene Ratio | Bg Ratio | p Value | Up | Down |
|---|---|---|---|---|---|---|
| ghi00196 | Photosynthesis antenna proteins | 33/877 | 53/11,853 | 1.32 × 10−24 | 33 | 0 |
| ghi00940 | Phenylpropanoid biosynthesis | 61/877 | 285/11,853 | 1.91 × 10−14 | 50 | 11 |
| ghi00941 | Flavonoid biosynthesis | 33/877 | 101/11,853 | 9.05 × 10−14 | 32 | 1 |
| ghi00500 | Starch and sucrose metabolism | 55/877 | 324/11,853 | 4.27 × 10−9 | 45 | 10 |
| ghi04712 | Circadian rhythm—plant | 28/877 | 127/11,853 | 1.26 × 10−7 | 25 | 3 |
| ghi00073 | Cutin, suberine, and wax biosynthesis | 17/877 | 64/11,853 | 2.56 × 10−6 | 15 | 2 |
| ghi00100 | Steroid biosynthesis | 19/877 | 84/11,853 | 8.89 × 10−6 | 18 | 1 |
| ghi00909 | Sesquiterpenoid and triterpenoid biosynthesis | 15/877 | 58/11,853 | 1.41 × 10−5 | 11 | 4 |
| ghi00480 | Glutathione metabolism | 33/877 | 249/11,853 | 0.00076 | 20 | 13 |
| ghi00460 | Cyanoamino acid metabolism | 16/877 | 91/11,853 | 0.00095 | 16 | 0 |
| ghi00195 | Photosynthesis | 20/877 | 138/11,853 | 0.002806 | 20 | 0 |
| ghi00966 | Glucosinolate biosynthesis | 7/877 | 29/11,853 | 0.004374 | 1 | 6 |
| ghi00670 | One carbon pool by folate | 10/877 | 56/11,853 | 0.007353 | 10 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.-Q.; Qiu, L.; Liu, L.-L.; Luo, L.; Han, X.-P.; Zhai, Y.-H.; Wang, W.-J.; Ren, M.-Z.; Xing, Y.-D. Identification and Comprehensive Structural and Functional Analyses of the EXO70 Gene Family in Cotton. Genes 2021, 12, 1594. https://doi.org/10.3390/genes12101594
Zhu Y-Q, Qiu L, Liu L-L, Luo L, Han X-P, Zhai Y-H, Wang W-J, Ren M-Z, Xing Y-D. Identification and Comprehensive Structural and Functional Analyses of the EXO70 Gene Family in Cotton. Genes. 2021; 12(10):1594. https://doi.org/10.3390/genes12101594
Chicago/Turabian StyleZhu, Ya-Qian, Lu Qiu, Lu-Lu Liu, Lei Luo, Xin-Pei Han, Yao-Hua Zhai, Wen-Jing Wang, Mao-Zhi Ren, and Ya-Di Xing. 2021. "Identification and Comprehensive Structural and Functional Analyses of the EXO70 Gene Family in Cotton" Genes 12, no. 10: 1594. https://doi.org/10.3390/genes12101594
APA StyleZhu, Y.-Q., Qiu, L., Liu, L.-L., Luo, L., Han, X.-P., Zhai, Y.-H., Wang, W.-J., Ren, M.-Z., & Xing, Y.-D. (2021). Identification and Comprehensive Structural and Functional Analyses of the EXO70 Gene Family in Cotton. Genes, 12(10), 1594. https://doi.org/10.3390/genes12101594