Comparative Genomics Provides Insights into the Taxonomy of Azoarcus and Reveals Separate Origins of Nif Genes in the Proposed Azoarcus and Aromatoleum Genera

 ,
,  , ,
, ,  , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Genomes
2.2. NCBI Sequence Datasets
2.3. Average Nucleotide Identity (ANI)
2.4. Protein Clusters
2.5. Core Genome Analysis Based on Core Protein Groups
2.6. Analysis of Nif and Other Functional Genes
2.7. SWeeP Phylogenies
2.8. Heat Maps
2.9. 16S rRNA Analysis
2.10. Nitrogenase Activity
3. Results and Discussion
3.1. Whole-Genome Sequences and Identity of Azoarcus sp. Strain TTM-91
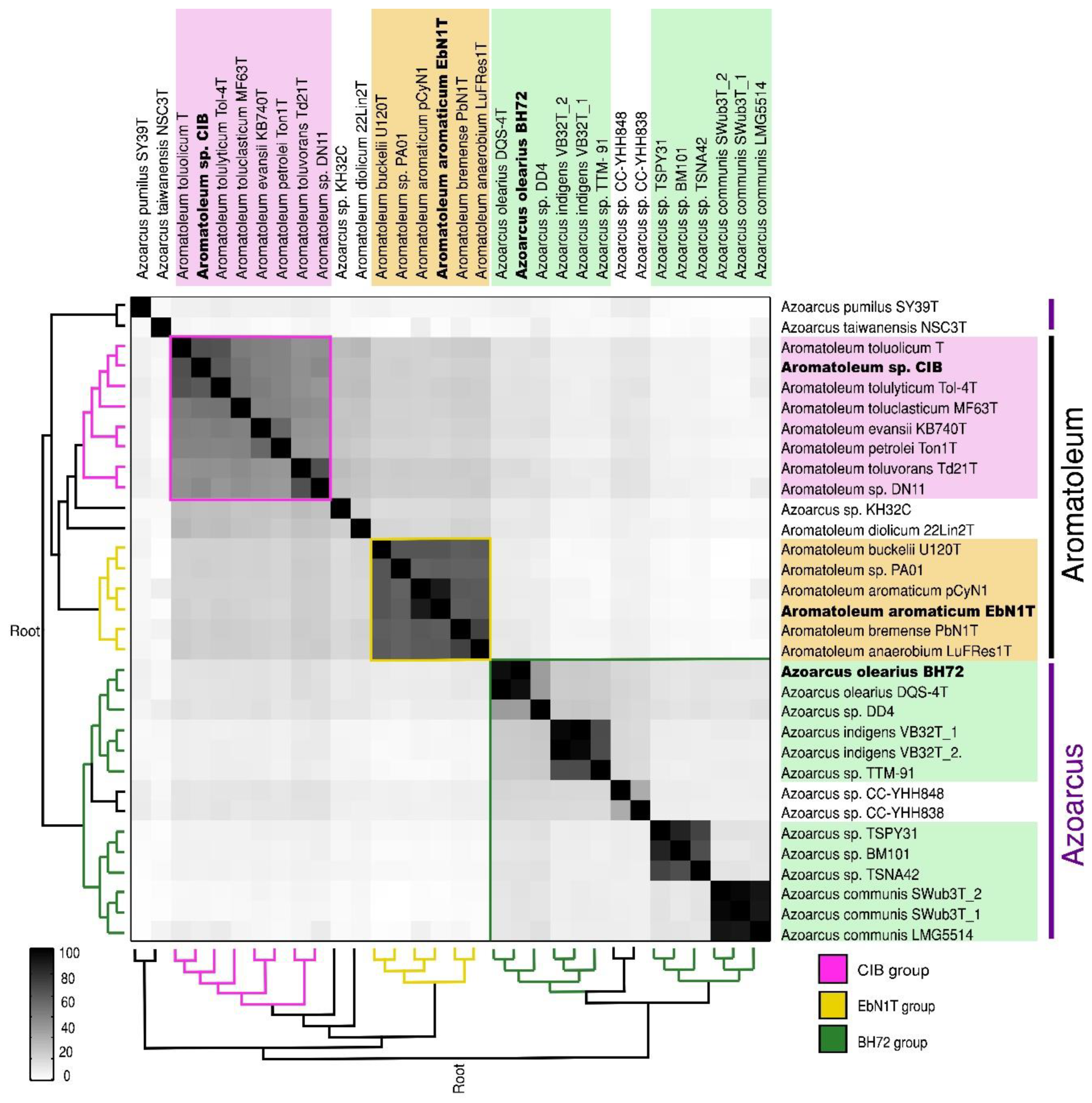
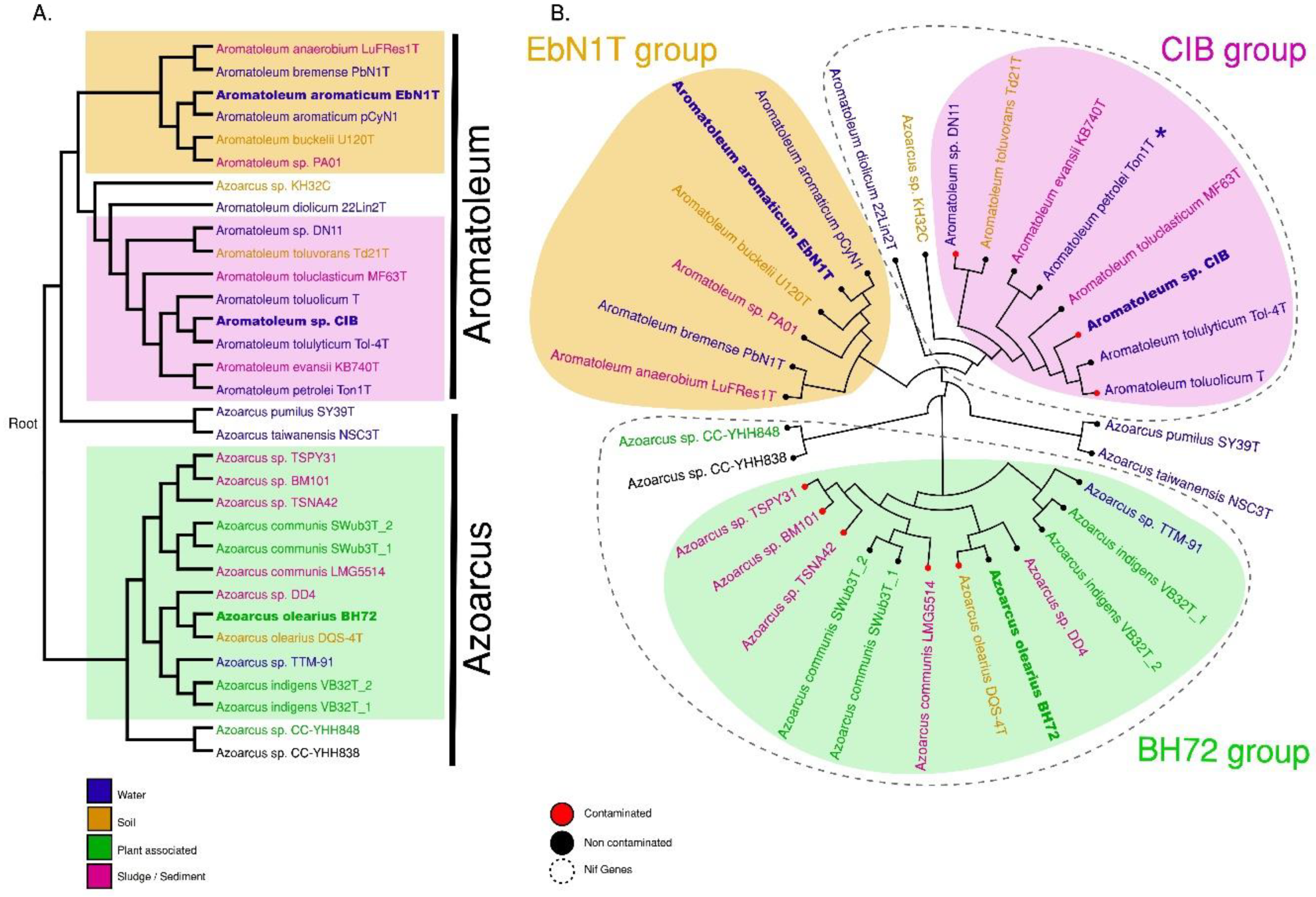
3.2. Average Nucleotide Identity (ANI) and Core Genome Analysis
3.3. Genes for Nitrate Reduction (Nar/Nap, Nir, Nor, Nos) and for the Anaerobic (Bzd) or Aerobic (Box) Degradation of Benzoate
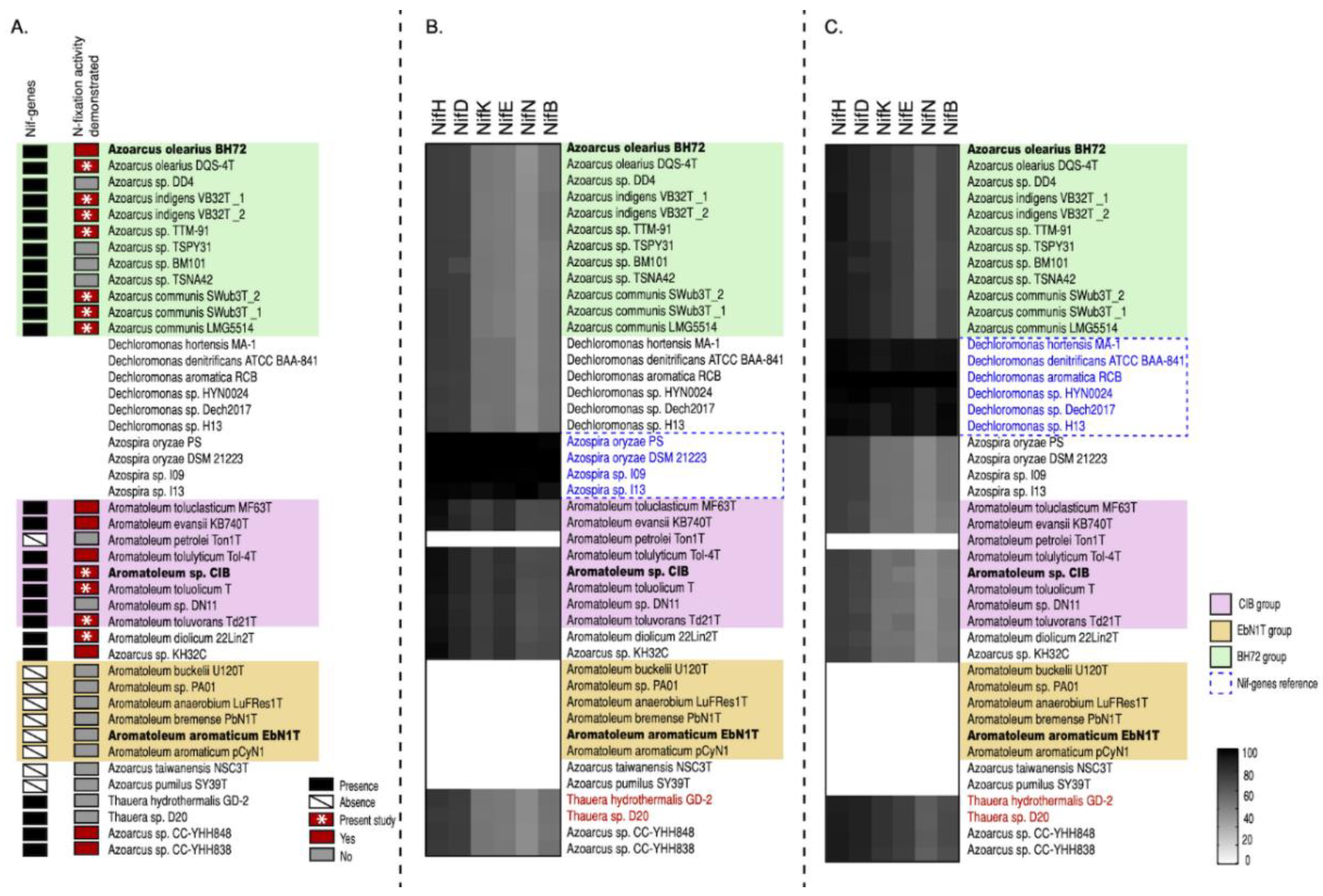
3.4. Nif Genes and Nitrogenase Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reinhold, B.; Hurek, T.; Niemann, E.-G.; Fendrik, I. Close association of Azospirillum and diazotrophic rods with different root zones of Kallar Grass. Appl. Environ. Microbiol. 1986, 52, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Reinhold, B.; Hurek, T.; Fendrik, I.; Pot, B.; Gillis, M.; Kersters, K.; Thielemans, S.; De Ley, J. Azospirillum halopraeferens sp. nov., a nitrogen-fixing organism associated with roots of Kallar Grass (Leptochloa fusca (L.) Kunth). Int. J. Syst. Evol. Microbiol. 1987, 37, 43–51. [Google Scholar] [CrossRef]
- Reinhold-Hurek, B.; Hurek, T.; Gillis, M.; Hoste, B.; Vancanneyt, M.; Kersters, K.; Ley, J. Azoarcus gen. nov., nitrogen-fixing proteobacteria associated with roots of Kallar Grass (Leptochloa fusca (L.) Kunth), and description of two species, Azoarcus indigens sp. nov. and Azoarcus communis sp. nov. Int. J. Syst. Evol. Microbiol. 1993, 43. [Google Scholar] [CrossRef]
- Chen, M.H.; Sheu, S.Y.; James, E.K.; Young, C.C.; Chen, W.M. Azoarcus olearius sp. nov., a nitrogen-fixing bacterium isolated from oil-contaminated soil. Int. J. Syst. Evol. Microbiol. 2013, 63, 3755–3761. [Google Scholar] [CrossRef] [PubMed]
- Faoro, H.; Rene Menegazzo, R.; Battistoni, F.; Gyaneshwar, P.; do Amaral, F.P.; Taule, C.; Rausch, S.; Goncalves Galvao, P.; de Los Santos, C.; Mitra, S.; et al. The oil-contaminated soil diazotroph Azoarcus olearius DQS-4(T) is genetically and phenotypically similar to the model grass endophyte Azoarcus sp. BH72. Environ. Microbiol. Rep. 2017, 9, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Anders, H.J.; Kaetzke, A.; Kampfer, P.; Ludwig, W.; Fuchs, G. Taxonomic position of aromatic-degrading denitrifying pseudomonad strains K 172 and KB 740 and their description as new members of the genera Thauera, as Thauera aromatica sp. nov., and Azoarcus, as Azoarcus evansii sp. nov., respectively, members of the beta subclass of the Proteobacteria. Int. J. Syst. Bacteriol. 1995, 45, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Fries, M.R.; Chee-Sanford, J.C.; Tiedje, J.M. Phylogenetic analyses of a new group of denitrifiers capable of anaerobic growth on toluene and description of Azoarcus tolulyticus sp. nov. Int. J. Syst. Bacteriol. 1995, 45, 500–506. [Google Scholar] [CrossRef]
- Springer, N.; Ludwig, W.; Philipp, B.; Schink, B. Azoarcus anaerobius sp. nov., a resorcinol-degrading, strictly anaerobic, denitrifying bacterium. Int. J. Syst. Bacteriol. 1998, 48, 953–956. [Google Scholar] [CrossRef]
- Song, B.; Haggblom, M.M.; Zhou, J.; Tiedje, J.M.; Palleroni, N.J. Taxonomic characterization of denitrifying bacteria that degrade aromatic compounds and description of Azoarcus toluvorans sp. nov. and Azoarcus toluclasticus sp. nov. Int. J. Syst. Bacteriol. 1999, 3, 1129–1140. [Google Scholar] [CrossRef]
- Mechichi, T.; Stackebrandt, E.; Gad’on, N.; Fuchs, G. Phylogenetic and metabolic diversity of bacteria degrading aromatic compounds under denitrifying conditions, and description of Thauera phenylacetica sp. nov., Thauera aminoaromatica sp. nov., and Azoarcus buckelii sp. nov. Arch. Microbiol. 2002, 178, 26–35. [Google Scholar] [CrossRef]
- Lee, D.-J.; Wong, B.-T.; Adav, S.S. Azoarcus taiwanensis sp. nov., a denitrifying species isolated from a hot spring. Appl. Microbiol. Biotechnol. 2014, 98, 1301–1307. [Google Scholar] [CrossRef] [PubMed]
- Rabus, R.; Kube, M.; Heider, J.; Beck, A.; Heitmann, K.; Widdel, F.; Reinhardt, R. The genome sequence of an anaerobic aromatic-degrading denitrifying bacterium, strain EbN1. Arch. Microbiol. 2005, 183, 27–36. [Google Scholar] [CrossRef]
- Nishizawa, T.; Tago, K.; Oshima, K.; Hattori, M.; Ishii, S.; Otsuka, S.; Senoo, K. Complete genome sequence of the denitrifying and N2O-reducing bacterium Azoarcus sp. strain KH32C. J. Bacteriol. 2012, 194, 06618-11. [Google Scholar] [CrossRef]
- Fernández-Llamosas, H.; Prandoni, N.; Fernández-Pascual, M.; Fajardo, S.; Morcillo, C.; Díaz, E.; Carmona, M. Azoarcus sp. CIB, an anaerobic biodegrader of aromatic compounds shows an endophytic lifestyle. PLoS ONE 2014, 9, e110771. [Google Scholar] [CrossRef]
- Junghare, M.; Patil, Y.; Schink, B. Draft genome sequence of a nitrate-reducing, o-phthalate degrading bacterium, Azoarcus sp. strain PA01(T). Stand Genom. Sci. 2015, 10, 90. [Google Scholar] [CrossRef] [PubMed]
- Deng, D.; Li, F.; Ye, L.; Li, M. Complete genome sequence of Azoarcus sp. strain DD4, a gram-negative propanotroph that degrades 1,4-dioxane and 1,1-dichloroethylene. Microbiol. Resour. Announc. 2019, 8, e00775-19. [Google Scholar] [CrossRef] [PubMed]
- Reinhold-Hurek, B.; Hurek, T. Reassessment of the taxonomic structure of the diazotrophic genus Azoarcus sensu lato and description of three new genera and new species, Azovibrio restrictus gen. nov., sp. nov., Azospira oryzae gen. nov., sp. nov. and Azonexus fungiphilus gen. nov., sp. nov. Int. J. Syst. Evol. Microbiol. 2000, 2, 649–659. [Google Scholar] [CrossRef]
- Loy, A.; Schulz, C.; Lücker, S.; Schöpfer-Wendels, A.; Stoecker, K.; Baranyi, C.; Lehner, A.; Wagner, M. 16S rRNA gene-based oligonucleotide microarray for environmental monitoring of the betaproteobacterial order “Rhodocyclales”. Appl. Environ. Microbiol. 2005, 71, 1373–1386. [Google Scholar] [CrossRef]
- Hurek, T.; Reinhold-Hurek, B. Azoarcus sp. strain BH72 as a model for nitrogen-fixing grass endophytes. J. Bacteriol. 2003, 106, 169–178. [Google Scholar] [CrossRef]
- Martin-Moldes, Z.; Zamarro, M.T.; Del Cerro, C.; Valencia, A.; Gomez, M.J.; Arcas, A.; Udaondo, Z.; Garcia, J.L.; Nogales, J.; Carmona, M.; et al. Whole-genome analysis of Azoarcus sp. strain CIB provides genetic insights to its different lifestyles and predicts novel metabolic features. Syst. Appl. Microbiol. 2015, 38, 462–471. [Google Scholar] [CrossRef]
- Rabus, R.; Wohlbrand, L.; Thies, D.; Meyer, M.; Reinhold-Hurek, B.; Kampfer, P. Aromatoleum gen. nov., a novel genus accommodating the phylogenetic lineage including Azoarcus evansii and related species, and proposal of Aromatoleum aromaticum sp. nov., Aromatoleum petrolei sp. nov., Aromatoleum bremense sp. nov., Aromatoleum toluolicum sp. nov. and Aromatoleum diolicum sp. nov. Int. J. Syst. Evol. Microbiol. 2019, 69, 982–997. [Google Scholar] [CrossRef] [PubMed]
- Hurek, T.; Egener, T.; Reinhold-Hurek, B. Divergence in nitrogenases of Azoarcus spp., Proteobacteria of the beta subclass. J. Bacteriol. 1997, 179, 4172–4178. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.-Y.; Yu, X.-Y.; Yu, X.-D.; Zhao, Z.; Chen, C.; Wang, R.-J.; Wu, M.; Zhang, X.-Q. Azoarcus pumilus sp. nov., isolated from seawater in Sanya, China. Int. J. Syst. Evol. Microbiol. 2019, 69, 1459–1464. [Google Scholar] [CrossRef] [PubMed]
- Beukes, C.W.; Palmer, M.; Manyaka, P.; Chan, W.Y.; Avontuur, J.R.; van Zyl, E.; Huntemann, M.; Clum, A.; Pillay, M.; Palaniappan, K.; et al. Genome data provides high support for generic boundaries in Burkholderia sensu lato. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Estrada-de Los Santos, P.; Palmer, M.; Chavez-Ramirez, B.; Beukes, C.; Steenkamp, E.T.; Briscoe, L.; Khan, N.; Maluk, M.; Lafos, M.; Humm, E.; et al. Whole genome analyses suggests that Burkholderia sensu lato contains two additional novel genera (Mycetohabitans gen. nov., and Trinickia gen. nov.): Implications for the evolution of diazotrophy and nodulation in the Burkholderiaceae. Genes 2018, 9, 389. [Google Scholar] [CrossRef]
- Avontuur, J.R.; Palmer, M.; Beukes, C.W.; Chan, W.Y.; Coetzee, M.P.A.; Blom, J.; Stepkowski, T.; Kyrpides, N.C.; Woyke, T.; Shapiro, N.; et al. Genome-informed Bradyrhizobium taxonomy: Where to from here? Syst. Appl. Microbiol. 2019, 42, 427–439. [Google Scholar] [CrossRef]
- Ormeño-Orrillo, E.; Martínez-Romero, E. A genomotaxonomy view of the Bradyrhizobium genus. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 5 January 2021).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bushnell, B. BBMap. Available online: https://sourceforge.net/p/bbmap/activity/?page=1&limit=100#5a6a7e4634309d1dd873dd30 (accessed on 20 April 2020).
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.; Korobeynikov, A.; Lapidus, A.; Prjibelsky, A.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling genomes and mini-metagenomes from highly chimeric reads. In Proceedings of the Annual International Conference on Research in Computational Molecular Biology, Beijing, China, 7–10 April 2013; Springer: Berlin/Heidelberg, Germany, 2013; pp. 158–170. [Google Scholar]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Rabus, R.; Kube, M.; Beck, A.; Widdel, F.; Reinhardt, R. Genes involved in the anaerobic degradation of ethylbenzene in a denitrifying bacterium, strain EbN1. Arch. Microbiol. 2002, 178, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Kube, M.; Heider, J.; Amann, J.; Hufnagel, P.; Kühner, S.; Beck, A.; Reinhardt, R.; Rabus, R. Genes involved in the anaerobic degradation of toluene in a denitrifying bacterium, strain EbN1. Arch. Microbiol. 2004, 181, 182–194. [Google Scholar] [CrossRef]
- Kühner, S.; Wöhlbrand, L.; Fritz, I.; Wruck, W.; Hultschig, C.; Hufnagel, P.; Kube, M.; Reinhardt, R.; Rabus, R. Substrate-dependent regulation of anaerobic degradation pathways for toluene and ethylbenzene in a denitrifying bacterium, strain EbN1. J. Bacteriol. 2005, 187, 1493–1503. [Google Scholar] [CrossRef] [PubMed]
- Devanadera, A.; Vejarano, F.; Zhai, Y.; Suzuki-Minakuchi, C.; Ohtsubo, Y.; Tsuda, M.; Kasai, Y.; Takahata, Y.; Okada, K.; Nojiri, H. Complete genome sequence of an anaerobic benzene-degrading bacterium, Azoarcus sp. Strain DN11. Microbiol. Resour. Announc. 2019, 8, e01699-18. [Google Scholar] [CrossRef] [PubMed]
- Zorraquino, V.; Toubiana, D.; Yan, D.; Blumwald, E. Draft genome sequence of the nitrogen-fixing endophyte Azoarcus communis SWub3. Microbiol. Resour. Announc. 2018, 7, e01080-18. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-S.; Cha, S.H.; Suk, H.Y.; Park, N.-H.; Woo, J.-H. Complete genome sequences of Azoarcus sp. TSPY31 and TSNA42 potentially having biosynthetic ability to produce indigo. Korean J. Microbiol. 2018, 54, 283–285. [Google Scholar] [CrossRef]
- Krause, A.; Ramakumar, A.; Bartels, D.; Battistoni, F.; Bekel, T.; Boch, J.; Bohm, M.; Friedrich, F.; Hurek, T.; Krause, L.; et al. Complete genome of the mutualistic, N2-fixing grass endophyte Azoarcus sp. strain BH72. Nat. Biotechnol. 2006, 24, 1385–1391. [Google Scholar] [CrossRef]
- Carlström, C.I.; Lucas, L.N.; Rohde, R.A.; Haratian, A.; Engelbrektson, A.L.; Coates, J.D. Characterization of an anaerobic marine microbial community exposed to combined fluxes of perchlorate and salinity. Appl. Microbiol. Biotechnol. 2016, 100, 9719–9732. [Google Scholar] [CrossRef]
- Lin, S.-Y.; Hameed, A.; Tsai, C.-F.; Huang, G.-H.; Hsu, Y.-H.; Young, C.-C. Description of Azoarcus nasutitermitis sp. nov. and Azoarcus rhizosphaerae sp. nov., two nitrogen-fixing species isolated from termite nest and rhizosphere of Ficus religiosa. Antonie Van Leeuwenhoek 2020, 113, 933–946. [Google Scholar] [CrossRef]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef]
- De Lima Nichio, B.T.; de Oliveira, A.M.R.; de Pierri, C.R.; Santos, L.G.C.; Lejambre, A.Q.; Vialle, R.A.; da Rocha Coimbra, N.A.; Guizelini, D.; Marchaukoski, J.N.; Pedrosa, F.O.; et al. RAFTS(3)G: An efficient and versatile clustering software to analyses in large protein datasets. BMC Bioinform. 2019, 20, 392. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, P.C.; Fang, Z.; Mason, S.W.; Setubal, J.C.; Dixon, R. Distribution of nitrogen fixation and nitrogenase-like sequences amongst microbial genomes. BMC Genom. 2012, 13, 1471–2164. [Google Scholar] [CrossRef] [PubMed]
- Vialle, R.A.; de Oliveira Pedrosa, F.; Weiss, V.A.; Guizelini, D.; Tibaes, J.H.; Marchaukoski, J.N.; de Souza, E.M.; Raittz, R.T. RAFTS3: Rapid Alignment-Free Tool for Sequence Similarity Search. bioRxiv 2016, 055269. [Google Scholar] [CrossRef]
- De Pierri, C.R.; Voyceik, R.; Santos de Mattos, L.G.C.; Kulik, M.G.; Camargo, J.O.; Repula de Oliveira, A.M.; de Lima Nichio, B.T.; Marchaukoski, J.N.; da Silva Filho, A.C.; Guizelini, D.; et al. SWeeP: Representing large biological sequences datasets in compact vectors. Sci. Rep. 2020, 10, 91. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Scornavacca, C. Dendroscope 3: An interactive tool for rooted phylogenetic trees and networks. Syst. Biol. 2012, 61, 1061–1067. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Dilworth, M.J. Acetylene reduction by nitrogen-fixing preparations from Clostridium pasteurianum. Biochim. Biophys. Acta 1966, 127, 285–294. [Google Scholar] [CrossRef]
- Schöllhorn, R.; Burris, R.H. Acetylene as a competitive inhibitor of N-2 fixation. Proc. Natl. Acad. Sci. USA 1967, 58, 213–216. [Google Scholar] [CrossRef]
- Zhang, W.; Wen, C.-K. Preparation of ethylene gas and comparison of ethylene responses induced by ethylene, ACC, and ethephon. Plant Physiol. Biochem. 2010, 48, 45–53. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Yang, L.; Muhadesi, J.B.; Wang, M.M.; Wang, B.J.; Liu, S.J.; Jiang, C.Y. Thauera hydrothermalis sp. nov., a thermophilic bacterium isolated from hot spring. Int. J. Syst. Evol. Microbiol. 2018, 68, 3163–3168. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.-S.; Liang, Q.-Y.; Du, Z.-J. Thauera lacus sp. nov., isolated from a saline lake in Inner Mongolia. Int. J. Syst. Evol. Microbiol. 2019, 69, 3786–3791. [Google Scholar] [CrossRef] [PubMed]
- Tindall, B.J.; Rosselló-Móra, R.; Busse, H.-J.; Ludwig, W.; Kämpfer, P. Notes on the characterization of prokaryote strains for taxonomic purposes. Int. J. Syst. Evol. Microbiol. 2010, 60, 249–266. [Google Scholar] [CrossRef]
- Qin, N.; Yang, F.; Li, A.; Prifti, E.; Chen, Y.; Shao, L.; Guo, J.; Le Chatelier, E.; Yao, J.; Wu, L.; et al. Alterations of the human gut microbiome in liver cirrhosis. Nature 2014, 513, 59–64. [Google Scholar] [CrossRef]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glockner, F.O.; Ludwig, W.; Schleifer, K.H.; Whitman, W.B.; Euzeby, J.; Amann, R.; Rossello-Mora, R. Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Barco, R.A.; Garrity, G.M.; Scott, J.J.; Amend, J.P.; Nealson, K.H.; Emerson, D. A genus definition for bacteria and archaea based on a standard genome relatedness index. mBio 2020, 11. [Google Scholar] [CrossRef]
- Qin, Q.-L.; Xie, B.-B.; Zhang, X.-Y.; Chen, X.-L.; Zhou, B.-C.; Zhou, J.; Oren, A.; Zhang, Y.-Z. A proposed genus boundary for the prokaryotes based on genomic insights. J. Bacteriol. 2014, 196, 2210–2215. [Google Scholar] [CrossRef]
- Kraft, B.; Strous, M.; Tegetmeyer, H.E. Microbial nitrate respiration—Genes, enzymes and environmental distribution. J. Biotechnol. 2011, 155, 104–117. [Google Scholar] [CrossRef]
- López Barragán, M.J.; Carmona, M.; Zamarro, M.T.; Thiele, B.; Boll, M.; Fuchs, G.; García, J.L.; Díaz, E. The bzd gene cluster, coding for anaerobic benzoate catabolism, in Azoarcus sp. strain CIB. J. Bacteriol. 2004, 186, 5762–5774. [Google Scholar] [CrossRef]
- Valderrama, J.A.; Durante-Rodríguez, G.; Blázquez, B.; García, J.L.; Carmona, M.; Díaz, E. Bacterial degradation of benzoate: Cross-regulation between aerobic and anaerobic pathways. J. Biol. Chem. 2012, 287, 10494–10508. [Google Scholar] [CrossRef] [PubMed]
- Zamarro, M.T.; Barragán, M.J.L.; Carmona, M.; García, J.L.; Díaz, E. Engineering a bzd cassette for the anaerobic bioconversion of aromatic compounds. Microb. Biotechnol. 2017, 10, 1418–1425. [Google Scholar] [CrossRef]
- Salinero, K.K.; Keller, K.; Feil, W.S.; Feil, H.; Trong, S.; Di Bartolo, G.; Lapidus, A. Metabolic analysis of the soil microbe Dechloromonas aromatica str. RCB: Indications of a surprisingly complex life-style and cryptic anaerobic pathways for aromatic degradation. BMC Genom. 2009, 10, 1471–2164. [Google Scholar] [CrossRef] [PubMed]
- Engelhard, M.; Hurek, T.; Reinhold-Hurek, B. Preferential occurrence of diazotrophic endophytes, Azoarcus spp., in wild rice species and land races of Oryza sativa in comparison with modern races. Environ. Microbiol. 2000, 2, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Morett, E.; Segovia, L. The sigma 54 bacterial enhancer-binding protein family: Mechanism of action and phylogenetic relationship of their functional domains. J. Bacteriol. 1993, 175, 6067–6074. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ledbetter, R.N.; Garcia Costas, A.M.; Lubner, C.E.; Mulder, D.W.; Tokmina-Lukaszewska, M.; Artz, J.H.; Patterson, A.; Magnuson, T.y.S.; Jay, Z.J.; Duan, H.; et al. The electron bifurcating FixABCX protein complex from Azotobacter vinelandii: Generation of low-potential reducing equivalents for nitrogenase catalysis. Biochemistry 2017, 56, 4177–4190. [Google Scholar] [CrossRef] [PubMed]
- Bueno Batista, M.; Dixon, R. Manipulating nitrogen regulation in diazotrophic bacteria for agronomic benefit. Biochem. Soc. Trans. 2019, 47, 603–614. [Google Scholar] [CrossRef]
- Fernández-Llamosas, H.; Ibero, J.; Thijs, S.; Imperato, V.; Vangronsveld, J.; Díaz, E.; Carmona, M. Enhancing the rice seedlings growth promotion abilities of Azoarcus sp. CIB by heterologous expression of ACC deaminase to improve performance of plants exposed to cadmium stress. Microorganisms 2020, 8, 1453. [Google Scholar] [CrossRef]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genome Size [bp] | Genome GC Content [%] | Nif Genes | Nitrogenase Activity Confirmed by Acetylene Reduction Assay (ARA) | Isolation Site and/or Type of Habitat | Country of Origin | Strain Origin and Description |
|---|---|---|---|---|---|---|---|
| Aromatoleum anaerobium LuFRes1T | 4,482,959 | 66.72 | No | No | Sewage sludge | Germany | [8] |
| Aromatoleum aromaticum EbN1T | 4,727,255 | 64.68 | No | No | River water | Germany | [12,21,34,35,36] |
| Aromatoleum aromaticum pCyN1 | 4,208,498 | 65.23 | No | No | River water | Germany | [21] |
| Aromatoleum bremense PbN1T | 4,319,217 | 65.57 | No | No | River water | Germany | [21] |
| Aromatoleum buckelii U120T | 4,227,546 | 65.33 | No | No | Oxic soil | Germany | [10] |
| Aromatoleum diolicum 22Lin2T | 5,138,050 | 64.22 | Yes | Yes | River water | Germany | [21] |
| Aromatoleum evansii KB740T | 5,883,543 | 65.98 | Yes | Yes | Creek sediment | USA | [6] |
| Aromatoleum petrolei Ton1T | 5,364,035 | 65.73 | No | No | River water | Germany | [21] |
| Aromatoleum sp. CIB | 5,257,030 | 65.84 | Yes | Yes | Diesel fuel-contaminated aquifer | Switzerland | [20] |
| Aromatoleum sp. DN11 | 4,956,835 | 66.30 | Yes | Yes | Gasoline-contaminated groundwater | Japan | [37] |
| Aromatoleum sp. PA01 | 3,908,240 | 66.10 | No | No | Sewage sludge, wastewater treatment plant | Germany | [15] |
| Aromatoleum toluclasticum MF63T | 5,925,983 | 66.00 | No | No | Shallow aquifer sediment | USA | [9] |
| Aromatoleum tolulyticum Tol-4T | 5,110,391 | 66.43 | Yes | Yes | Petroleum contaminated aquifer sediments | USA | [7] |
| Aromatoleum toluolicum TT | 6,025,652 | 65.93 | Yes | Yes | Petroleum contaminated aquifer | Switzerland | [21] |
| Aromatoleum toluvorans Td21T | 4,786,256 | 66.66 | Yes | Yes | Muck soil | USA | [9] |
| Azoarcus communis LMG5514 | 4,996,403 | 62.45 | Yes | Yes | Oily sludge, storage tank, petroleum refinery | France | [3] |
| Azoarcus communis SWub3T_1 | 4,976,326 | 62.36 | Yes | Yes | Kallar Grass roots | Pakistan | [3] |
| Azoarcus communis SWub3T_2 | 5,004,685 | 62.50 | Yes | Yes | Kallar Grass roots | Pakistan | [3,38] |
| Azoarcus sp. TSNA42 | 4,886,934 | 62.80 | Yes | nt * | Oil-contaminated sediment | South Korea | [39] |
| Azoarcus sp. TSPY31 | 4,572,081 | 63.20 | Yes | nt | Oil-contaminated sediment | South Korea | [39] |
| Azoarcus sp. TTM-91 | 5,393,782 | 67.70 | Yes | Yes | River water | Taiwan | This study |
| Azoarcus indigens VB32T | 5,562,509 | 67.31 | Yes | Yes | Kallar Grass roots | Pakistan | [3] |
| Azoarcus indigens VB32T | 5,464,470 | 67.60 | Yes | Yes | Kallar Grass roots | Pakistan | [3] |
| Azoarcus olearius BH72 | 4,376,040 | 67.90 | Yes | Yes | Kallar Grass roots | Pakistan | [3,40] |
| Azoarcus olearius DQS-4T | 4,451,751 | 67.83 | Yes | Yes | Oil-contaminated soil | Taiwan | [4,5] |
| Azoarcus pumilus SY39T | 3,225,512 | 66.53 | No | No | Sea water | China | [23] |
| Azoarcus sp. BM101 | 4,904,245 | 62.90 | Yes | nt | Estuary sediment | USA | [41] |
| Azoarcus sp. CC-YHH838 | 4,723,750 | 67.50 | Yes | Yes | Termite nest | Taiwan | [42] |
| Azoarcus sp. CC-YHH848 | 4,602,270 | 68.30 | Yes | Yes | Rhizosphere of Ficus religiosa | Taiwan | [42] |
| Azoarcus sp. DD4 | 5,400,077 | 66.70 | Yes | nt | Activated sludge, wastewater treatment plant | USA | [16] |
| Azoarcus sp. KH32C | 5,818,755 | 65.03 | Yes | Yes | Soil from field with paddy rice and soybean crops | Japan | [13] |
| Azoarcus taiwanensis NSC3T | 4,228,584 | 62.88 | No | No | Hot spring | Taiwan | [11] |
| Strain or Control | Activity ± SEM nmol C2H4 mg Protein−1 min−1 | Conditions 3 |
|---|---|---|
| Control 1 | 0.111 ± 0.088 | SM-semi-solid |
| Control 2 | not detected | SM-semi-solid |
| Az. olearius DQS-4T | 23.222 ± 0.826 | SM-semi-solid |
| Az. communis SWub3T | 10.611 ± 0.778 | SM-semi-solid |
| Az. communis LMG5514 | 26.025 ± 1.041 | SM-semi-solid |
| Az. indigens VB32T | 17.952 ± 0.653 | 1/2CVM-semi-solid |
| Azoarcus sp. TTM-91 | 3.960 ± 0.724 | 1/2CVM-semi-solid |
| Aromatoleum sp. CIB | 1.163 ± 0.041 | MNF + 3 mM Benzoate-liquid |
| Ar. toluvorans Td21T | 1.318 ± 0.354 | MNF + 3 mM Benzoate-liquid |
| Ar. diolicum 22Lin2T | 0.479 ± 0.022 | UMS + 30 mM Malate-liquid-1% O2 |
| Ar. toluolicum TT | 0.653 ± 0.049 | UMS + 30 mM Malate-liquid-1% O2 |
| Ar. evansii KB740T | not detected | All above conditions tested |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raittz, R.T.; Reginatto De Pierri, C.; Maluk, M.; Bueno Batista, M.; Carmona, M.; Junghare, M.; Faoro, H.; Cruz, L.M.; Battistoni, F.; Souza, E.d.; et al. Comparative Genomics Provides Insights into the Taxonomy of Azoarcus and Reveals Separate Origins of Nif Genes in the Proposed Azoarcus and Aromatoleum Genera. Genes 2021, 12, 71. https://doi.org/10.3390/genes12010071
Raittz RT, Reginatto De Pierri C, Maluk M, Bueno Batista M, Carmona M, Junghare M, Faoro H, Cruz LM, Battistoni F, Souza Ed, et al. Comparative Genomics Provides Insights into the Taxonomy of Azoarcus and Reveals Separate Origins of Nif Genes in the Proposed Azoarcus and Aromatoleum Genera. Genes. 2021; 12(1):71. https://doi.org/10.3390/genes12010071
Chicago/Turabian StyleRaittz, Roberto Tadeu, Camilla Reginatto De Pierri, Marta Maluk, Marcelo Bueno Batista, Manuel Carmona, Madan Junghare, Helisson Faoro, Leonardo M. Cruz, Federico Battistoni, Emanuel de Souza, and et al. 2021. "Comparative Genomics Provides Insights into the Taxonomy of Azoarcus and Reveals Separate Origins of Nif Genes in the Proposed Azoarcus and Aromatoleum Genera" Genes 12, no. 1: 71. https://doi.org/10.3390/genes12010071
APA StyleRaittz, R. T., Reginatto De Pierri, C., Maluk, M., Bueno Batista, M., Carmona, M., Junghare, M., Faoro, H., Cruz, L. M., Battistoni, F., Souza, E. d., Pedrosa, F. d. O., Chen, W.-M., Poole, P. S., Dixon, R. A., & James, E. K. (2021). Comparative Genomics Provides Insights into the Taxonomy of Azoarcus and Reveals Separate Origins of Nif Genes in the Proposed Azoarcus and Aromatoleum Genera. Genes, 12(1), 71. https://doi.org/10.3390/genes12010071