Genetic Analysis of Root-to-Shoot Signaling and Rootstock-Mediated Tolerance to Water Deficit in Tomato

 ,
,  ,
,  ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material, Growth Conditions and Trait Evaluation
2.2. Statistical Analysis
2.3. Molecular Markers and QTL Analysis
3. Results
4. Discussion
4.1. S. pimpinellifolium Provides Water Deficit Tolerance Genes for Tomato Rootstocks
4.2. Root Acquisition and Long-Distance Transport of Nutrients.
4.3. Root-to-Shoot Hormone and Peptide Signalling
4.4. Transcription Factors as Candidate Genes at QTLs for Water-Shortage Tolerance
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gilliham, M.; Able, J.A.; Roy, S.J. Translating knowledge about abiotic stress tolerance to breeding programmes. Plant J. 2017, 90, 898–917. [Google Scholar] [CrossRef]
- FAO. Drought and Agriculture. 2017. Available online: http://www.fao.org/3/a-i7378e.pdf (accessed on 22 December 2020).
- Kim, Y.; Chung, Y.S.; Lee, E.; Tripathi, P.; Heo, S.; Kim, K.-H. Root Response to Drought Stress in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2020, 21, 1513. [Google Scholar] [CrossRef]
- Wood, A.J. Eco-Physiological Adaptations to Limited Water Environments. In Plant Abiotic Stress; Blackwell Publishing Ltd.: Noida, India, 2005; pp. 1–13. [Google Scholar]
- Bretó, M.P.; Asins, M.J.; Carbonell, E.A. Distribution of genetic variability in Lycopersicon species and their genetic relationships. Theor. Appl. Genet. 1993, 86, 113–120. [Google Scholar] [CrossRef]
- Peralta, I.E.; Spooner, D.M.; Knapp, S. Taxonomy of wild tomato and their relatives (Solanum Sect. Lycopersicoides, Sect. Jungladifolia, Sect. Lycopersicum; Solanaceae). Syst. Bot. Monogr. 2008, 84, 183. [Google Scholar]
- Lin, Y.-P.; Lu, C.-Y.; Lee, C.-R. The climatic association of population divergence and future extinction risk of Solanum pimpinellifolium. AoB Plants 2020, 12, plaa012. [Google Scholar] [CrossRef]
- Semel, Y.; Schauer, N.; Roessner, U.; Zamir, D.; Fernie, A.R. Metabolite analysis for the comparison of irrigated and non-irrigated field grown tomato of varying genotype. Metabolomics 2007, 3, 289–295. [Google Scholar] [CrossRef]
- Tardieu, F.; Tuberosa, R. Dissection and modelling of abiotic stress tolerance in plants. Curr. Opin. Plant Biol. 2010, 13, 206–212. [Google Scholar] [CrossRef]
- Wang, H.; Wang, H.; Shao, H.; Tang, X. Recent Advances in Utilizing Transcription Factors to Improve Plant Abiotic Stress Tolerance by Transgenic Technology. Front. Plant Sci. 2016, 7, 67. [Google Scholar] [CrossRef]
- Guo, Y.; Ping, W.; Chen, J.; Zhu, L.; Zhao, Y.; Guo, J.; Huang, Y. Meta-analysis of the effects of overexpression of WRKY transcription factors on plant responses to drought stress. BMC Genet. 2019, 20, 63. [Google Scholar] [CrossRef]
- Dong, C.; Ma, Y.; Wisniewski, M.; Cheng, Z.-M. Meta-analysis of the effect of overexpression of CBF/DREB family genes on drought stress response. Environ. Exp. Bot. 2017, 142, 1–14. [Google Scholar] [CrossRef]
- Zhao, P.-X.; Miao, Z.-Q.; Zhang, J.; Chen, S.-Y.; Liu, Q.-Q.; Xiang, C. Arabidopsis MADS-box factor AGL16 negatively regulates drought resistance via stomatal density and stomatal movement. J. Exp. Bot. 2020. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef]
- Argueso, C.T.; Ferreira, F.J.; Kieber, J.J. Environmental perception avenues: The interaction of cytokinin and environmental response pathways. Plant Cell Environ. 2009, 32, 1147–1160. [Google Scholar] [CrossRef]
- Thirumalaikumar, V.P.; Devkar, V.; Mehterov, N.; Ali, S.; Ozgur, R.; Turkan, I.; Mueller-Roeber, B.; Balazadeh, S. NAC transcription factor JUNGBRUNNEN1 enhances drought tolerance in tomato. Plant Biotechnol. J. 2017, 16, 354–366. [Google Scholar] [CrossRef]
- Tolosa, L.N.; Zhang, Z. The Role of Major Transcription Factors in Solanaceous Food Crops under Different Stress Conditions: Current and Future Perspectives. Plants 2020, 9, 56. [Google Scholar] [CrossRef]
- Ning, J.; Li, X.; Hicks, L.M.; Xiong, L. A Raf-Like MAPKKK Gene DSM1 Mediates Drought Resistance through Reactive Oxygen Species Scavenging in Rice. Plant Physiol. 2009, 152, 876–890. [Google Scholar] [CrossRef]
- He, X.; Wang, C.; Wang, H.; Li, L.; Wang, C. The Function of MAPK Cascades in Response to Various Stresses in Horticultural Plants. Front. Plant Sci. 2020, 11, 952. [Google Scholar] [CrossRef]
- Parmar, N.; Singh, K.H.; Sharma, D.; Singh, L.; Kumar, P.; Nanjundan, J.; Khan, Y.J.; Chauhan, D.K.; Thakur, A.K. Genetic engineering strategies for biotic and abiotic stress tolerance and quality enhancement in horticultural crops: A comprehensive review. 3 Biotech 2017, 7, 239. [Google Scholar] [CrossRef]
- Tardieu, F.; Simonneau, T.; Muller, B. The Physiological Basis of Drought Tolerance in Crop Plants: A Scenario-Dependent Probabilistic Approach. Annu. Rev. Plant Biol. 2018, 69, 733–759. [Google Scholar] [CrossRef]
- Cui, J.-T.; Shao, G.-C.; Lu, J.; Keabetswe, L.; Hoogenboom, G. Yield, quality and drought sensitivity of tomato to water deficit during different growth stages. Sci. Agric. 2020, 77, 20180390. [Google Scholar] [CrossRef]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Tanaka, M.; Seki, M.; Ham, L.H.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.-S.P. Differential Gene Expression in Soybean Leaf Tissues at Late Developmental Stages under Drought Stress Revealed by Genome-Wide Transcriptome Analysis. PLoS ONE 2012, 7, e49522. [Google Scholar] [CrossRef]
- Shanker, A.K.; Mandapaka, M.; Yadav, S.K.; Desai, S.; Bhanu, D.; Attal, N.B.; Bandi, V. Drought stress responses in crops. Funct. Integr. Genom. 2014, 14, 11–22. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, C.; Chen, H.Y.H.; Ruan, H. Response of Plants to Water Stress: A Meta-Analysis. Front. Plant Sci. 2020, 11, 978. [Google Scholar] [CrossRef]
- Davies, W.J.; Kudoyarova, G.R.; Hartung, W. Long-distance ABA Signaling and Its Relation to Other Signaling Pathways in the Detection of Soil Drying and the Mediation of the Plant’s Response to Drought. J. Plant Growth Regul. 2005, 24, 285–295. [Google Scholar] [CrossRef]
- Chan, K.X.; Wirtz, M.; Phua, S.Y.; Estavillo, G.M.; Pogson, B.J. Balancing metabolites in drought: The sulfur assimilation conundrum. Trends Plant Sci. 2013, 18, 18–29. [Google Scholar] [CrossRef]
- Reichardt, S.; Piepho, H.-P.; Stintzi, A.; Schaller, A. Peptide signaling for drought-induced tomato flower drop. Science 2020, 367, 1482–1485. [Google Scholar] [CrossRef]
- Rivas-Ubach, A.; Sardans, J.; Pérez-Trujillo, M.; Estiarte, M.; Peñuelas, J. Strong relationship between elemental stoichiometry and metabolome in plants. Proc. Natl. Acad. Sci. USA 2012, 109, 4181–4186. [Google Scholar] [CrossRef]
- Hughes, J.; Hepworth, C.; Dutton, C.; Dunn, J.; Hunt, L.; Stephens, J.; Waugh, R.; Cameron, D.D.; Gray, J.E. Reducing Stomatal Density in Barley Improves Drought Tolerance without Impacting on Yield. Plant Physiol. 2017, 174, 776–787. [Google Scholar] [CrossRef]
- Albert, E.; Gricourt, J.; Bertin, N.; Bonnefoi, J.; Pateyron, S.; Tamby, J.-P.; Bitton, F.; Causse, M. Genotype by watering regime interaction in cultivated tomato: Lessons from linkage mapping and gene expression. Theor. Appl. Genet. 2015, 129, 395–418. [Google Scholar] [CrossRef]
- Diouf, I.A.; Derivot, L.; Bitton, F.; Pascual, L.; Causse, M. Water Deficit and Salinity Stress Reveal Many Specific QTL for Plant Growth and Fruit Quality Traits in Tomato. Front. Plant Sci. 2018, 9, 279. [Google Scholar] [CrossRef]
- Gur, A.; Semel, Y.; Osorio, S.; Friedmann, M.; Seekh, S.; Ghareeb, B.; Mohammad, A.; Pleban, T.; Gera, G.; Fernie, A.R.; et al. Yield quantitative trait loci from wild tomato are predominately expressed by the shoot. Theor. Appl. Genet. 2010, 122, 405–420. [Google Scholar] [CrossRef] [PubMed]
- Easlon, H.M.; Clair, D.A.S.; Bloom, A.J. An Introgression from Wild Tomato (Solanum habrochaites) Affects Tomato Photosynthesis and Water Relations. Crop. Sci. 2014, 54, 779–784. [Google Scholar] [CrossRef]
- Arms, E.M.; Bloom, A.J.; Clair, D.A.S. High-resolution mapping of a major effect QTL from wild tomato Solanum habrochaites that influences water relations under root chilling. Theor. Appl. Genet. 2015, 128, 1713–1724. [Google Scholar] [CrossRef] [PubMed]
- Cantero-Navarro, E.; Romero-Aranda, R.; Fernández-Muñoz, R.; Martínez-Andújar, C.; Pérez-Alfocea, F.; Albacete, A. Improving agronomic water use efficiency in tomato by rootstock-mediated hormonal regulation of leaf biomass. Plant Sci. 2016, 251, 90–100. [Google Scholar] [CrossRef]
- Kumar, P.; Rouphael, Y.; Cardarelli, M.; Colla, G. Vegetable Grafting as a Tool to Improve Drought Resistance and Water Use Efficiency. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, Y.; Cao, B.; Chen, Z.; Xu, K. The effectiveness of grafting to improve drought tolerance in tomato. Plant Growth Regul. 2020, 91, 157–167. [Google Scholar] [CrossRef]
- Albacete, A.; Martínez-Andújar, C.; Ghanem, M.E.; Acosta, M.; Sánchez-Bravo, J.; Asins, M.J.; Cuartero, J.; Lutts, S.; Dodd, I.C.; Pérez-Alfocea, F. Rootstock-mediated changes in xylem ionic and hormonal status are correlated with delayed leaf senescence, and increased leaf area and crop productivity in salinized tomato. Plant Cell Environ. 2009, 32, 928–938. [Google Scholar] [CrossRef]
- Lee, J.-M.; Kubota, C.; Tsao, S.; Bie, Z.; Echevarria, P.H.; Morra, L.; Oda, M. Current status of vegetable grafting: Diffusion, grafting techniques, automation. Sci. Hortic. 2010, 127, 93–105. [Google Scholar] [CrossRef]
- Price, A.H. Believe it or not, QTLs are accurate! Trends Plant Sci. 2006, 11, 213–216. [Google Scholar] [CrossRef]
- The Tomato Genome Consortium. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [CrossRef]
- Monforte, A.; Asins, M.J.; Carbonell, E.A. Salt tolerance in Lycopersicon species. V. May genetic variability at quantitative trait loci affect their analysis? Theor. Appl. Genet. 1997, 95, 284–293. [Google Scholar] [CrossRef]
- Albacete, A.; Ghanem, M.E.; Martínez-Andújar, C.; Acosta, M.; Sánchez-Bravo, J.; Martínez, V.; Lutts, S.; Dodd, I.C.; Pérez-Alfocea, F. Hormonal changes in relation to biomass partitioning and shoot growth impairment in salinized tomato (Solanum lycopersicum L.) plants. J. Exp. Bot. 2008, 59, 4119–4131. [Google Scholar] [CrossRef] [PubMed]
- Villalta, I.; Bernet, G.P.; Carbonell, E.A.; Asins, M.J. Comparative QTL analysis of salinity tolerance in terms of fruit yield using two solanum populations of F7 lines. Theor. Appl. Genet. 2007, 114, 1001–1017. [Google Scholar] [CrossRef] [PubMed]
- Asins, M.J.; Raga, V.; Roca, D.; Belver, A.; Carbonell, E.A. Genetic dissection of tomato rootstock effects on scion traits under moderate salinity. Theor. Appl. Genet. 2015, 128, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Van Ooijen, J.W. MapQTL 6. Software for the Mapping of Quantitative Trait Loci in Experimental Populations of Diploid Species; Kyazma BV: Wageningen, The Netherlands, 2009. [Google Scholar]
- Kevei, Z.; King, R.C.; Mohareb, F.; Sergeant, M.J.; Awan, S.Z.; Thompson, A.J. Resequencing at ≥40-Fold Depth of the Parental Genomes of a Solanum lycopersicum × S. pimpinellifolium Recombinant Inbred Line Population and Characterization of Frame-Shift InDels That Are Highly Likely to Perturb Protein Function. G3 Genes Genomes Genet. 2015, 5, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. agriGO v2.0: A GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef]
- Asins, M.J.; Raga, M.V.; Torrent, D.; Roca, D.; Carbonell, E.A. QTL and candidate gene analyses of rootstock-mediated tomato fruit yield and quality traits under low iron stress. Euphytica 2020, 216, 1–19. [Google Scholar] [CrossRef]
- Jaime-Pérez, N.; Pineda, B.; García-Sogo, B.; Atares, A.; Athman, A.; Byrt, C.S.; Olias, R.; Asins, M.J.; Gilliham, M.; Moreno, V.; et al. The Na+ transporter encoded by the HKT1;2 gene modulates Na+/K+ homeostasis in tomato shoots under salinity. Plant Cell Environ. 2017, 40, 658–671. [Google Scholar] [CrossRef]
- Plouznikoff, K.; Asins, M.J.; De Boulois, H.D.; Carbonell, E.A.; Declerck, S. Genetic analysis of tomato root colonization by arbuscular mycorrhizal fungi. Ann. Bot. 2019, mcy240. [Google Scholar] [CrossRef]
- Rivero, J.; Álvarez, D.; Flors, V.; Azcón-Aguilar, C.; Pozo, M.J. Root metabolic plasticity underlies functional diversity in mycorrhiza-enhanced stress tolerance in tomato. New Phytol. 2018, 220, 1322–1336. [Google Scholar] [CrossRef]
- Ghanem, M.E.; Albacete, A.; Smigocki, A.C.; Frébort, I.; Pospíšilová, H.; Martínez-Andújar, C.; Acosta, M.; Sánchez-Bravo, J.; Lutts, S.; Dodd, I.C.; et al. Root-synthesized cytokinins improve shoot growth and fruit yield in salinized tomato (Solanum lycopersicum L.) plants. J. Exp. Bot. 2010, 62, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Hwang, J.-U.; Lee, M.; Kim, Y.-Y.; Assmann, S.M.; Martinoia, E.; Lee, Y. PDR-type ABC transporter mediates cellular uptake of the phytohormone abscisic acid. Proc. Natl. Acad. Sci. USA 2010, 107, 2355–2360. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fan, K.; Hung, S.; Chen, Y.-R. The role of peptides cleaved from protein precursors in eliciting plant stress reactions. New Phytol. 2020, 225, 2267–2282. [Google Scholar] [CrossRef] [PubMed]
- Fanourakis, D.; Tsaniklidis, G.; Pappi, P.; Markakis, E.A.; Doupis, G.; Charova, S.N.; Delis, C.; Tsaniklidis, G. The Role of Proteases in Determining Stomatal Development and Tuning Pore Aperture: A Review. Plants 2020, 9, 340. [Google Scholar] [CrossRef]
- Gonzalo, M.J.; Li, Y.-C.; Chen, K.-Y.; Gil, D.; Montoro, T.; Nájera, I.; Baixauli, C.; Granell, A.; Monforte, A.J. Genetic Control of Reproductive Traits in Tomatoes Under High Temperature. Front. Plant Sci. 2020, 11, 326. [Google Scholar] [CrossRef]



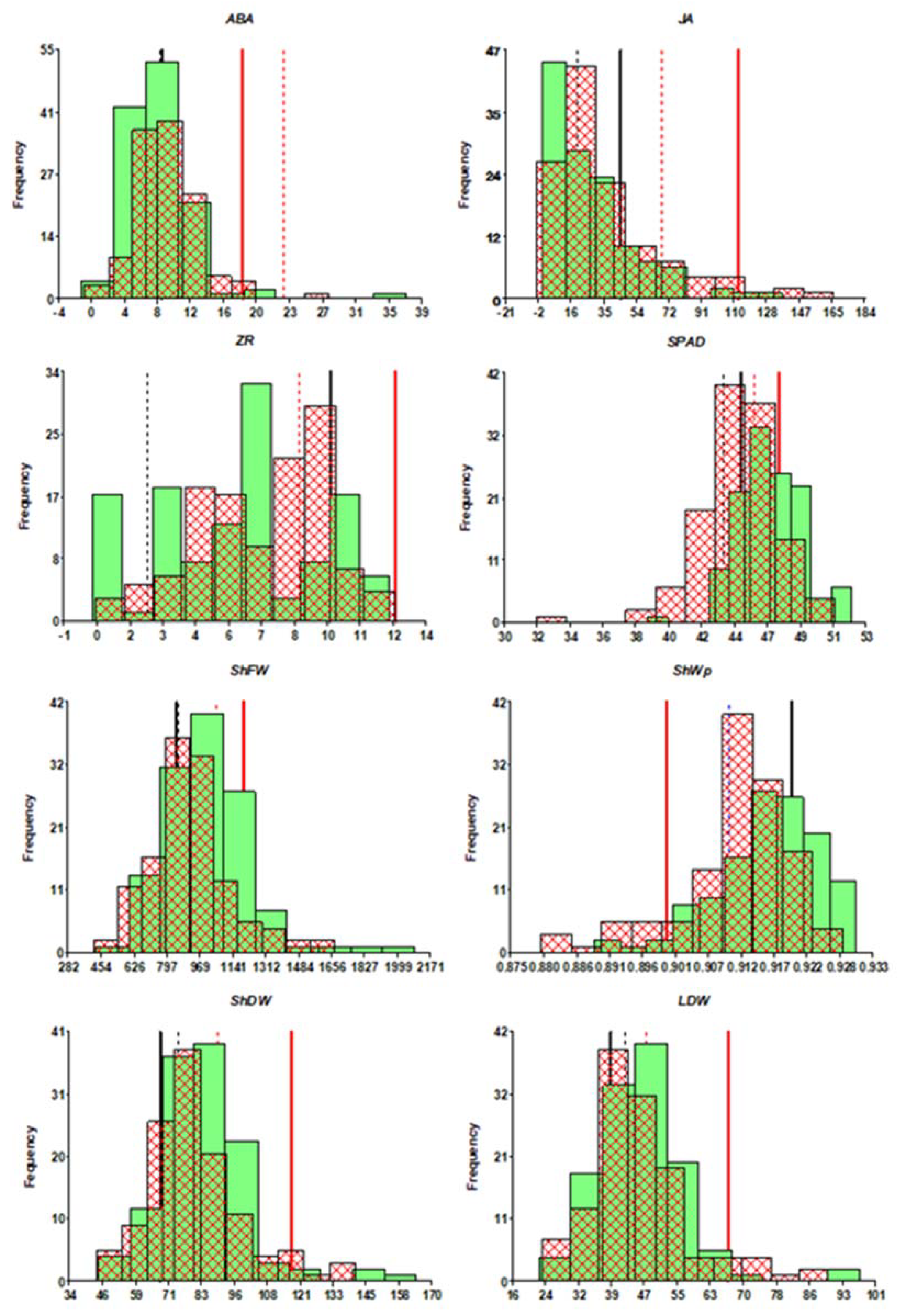{kind=link}
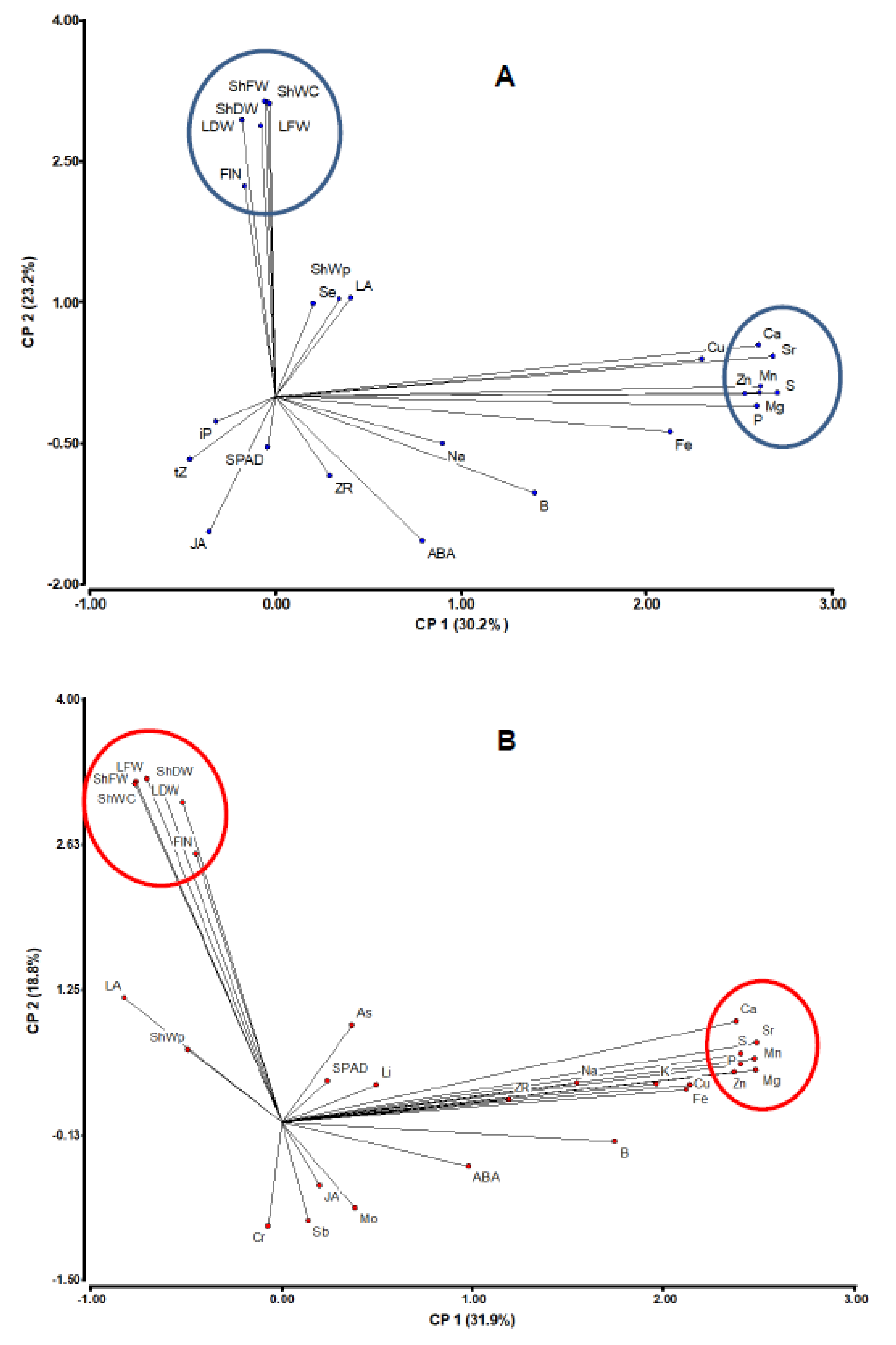{kind=link}
| Abbreviation | Trait | Bol/Bol_C | Bol_C | Bol/Bol_D | Bol_D | Mi_C | Ma_C | Mi_D | Ma_D | H2_C | H2_D | G | E | G × E |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ABA | Xylem sap [ABA] | 17.91 ± 6.2 | 8.48 ± 4.19 | 22.76 ± 8.94 | 8.3 ± 2.12 | –0.85 | 36.96 | –0.56 | 28.03 | 0.1505 | 0.1544 | <0.0001 | 0.1815 | 0.0932 |
| ACC | Xylem sap [ACC] | 9 ± 4.5 | 8.96 ± 4.48 | 4.66 ± 2.33 | 7.97 ± 3.99 | 0.00 | 23.32 | 0.00 | 26.93 | 0.0308 | 0.0724 | 0.0396 | 0.8182 | 0.3208 |
| As | Xylem sap [As] (mg/L) | 0.11 ± 0.03 | 0.12 ± 0.03 | 0.1 ± 0.01 | 0.15 ± 0.04 | 0.01 | 0.31 | 0.01 | 0.24 | 0.0000 | 0.1325 | 0.2856 | 0.7236 | 0.2377 |
| B | Xylem sap [B] (mg/L) | 0.32 ± 0.1 | 0.2 ± 0.02 | 0.25 ± 0.03 | 0.36 ± 0.09 | –0.06 | 0.83 | –0.02 | 0.81 | 0.3007 | 0.3249 | <0.0001 | 0.1699 | <0.0001 |
| Ca | Xylem sap [Ca] (mg/L) | 154.62 ± 23.41 | 85.25 ± 9.83 | 170.87 ± 44.94 | 269.57 ± 27.46 | 62.35 | 549.20 | 56.06 | 619.80 | 0.1539 | 0.2800 | 0.0021 | 0.3211 | <0.0001 |
| Cr | Xylem sap [Cr] (mg/L) | 0.03 ± 0.02 | 0.01 ± 0.01 | 0 ± 0 | 0 ± 0 | 0.00 | 0.14 | –0.01 | 0.09 | 0.0670 | 0.1894 | 0.0475 | 0.658 | 0.016 |
| Cu | Xylem sap [Cu] (mg/L) | 0.1 ± 0.01 | 0.08 ± 0.03 | 0.08 ± 0 | 0.12 ± 0.0033 | 0.04 | 0.33 | 0.04 | 0.43 | 0.1859 | 0.1218 | 0.0003 | 0.9083 | 0.1081 |
| Fe | Xylem sap [Fe] (mg/L) | 0.36 ± 0.2 | 0.19 ± 0.02 | 0.25 ± 0.08 | 0.37 ± 0.12 | 0.04 | 1.61 | 0.09 | 1.46 | 0.1711 | 0.2052 | <0.0001 | 0.997 | 0.2186 |
| FlN | Number of flowers | 5.67 ± 0.88 | 3.67 ± 0.33 | 4.33 ± 0.88 | 3.67 ± 0.88 | 2.69 | 9.40 | 1.54 | 7.04 | 0.0214 | 0.1638 | <0.0001 | 0.7691 | 0.5706 |
| iP | Xylem sap [isopentenyladenine] | 2.81 ± 0.33 | 2.43 ± 0.11 | 2.14 ± 0.16 | 2.01 ± 0.13 | –0.05 | 20.54 | 0.00 | 11.09 | 0.1316 | 0.0905 | <0.0001 | 0.2615 | 0.0237 |
| JA | Xylem sap [Jasmonic acid] | 111.67 ± 49.84 | 44.6 ± 42.45 | 67.79 ± 42.62 | 19.89 ± 11.72 | 0.00 | 137.08 | –3.03 | 166.23 | 0.1461 | 0.1970 | 0.0004 | 0.1956 | 0.0045 |
| K | Xylem sap [K] (mg/L) | 522.65 ± 104.91 | 398.33 ± 23.16 | 580.38 ± 140.63 | 622.03 ± 46.16 | 242.79 | 897.09 | 184.37 | 1053.70 | 0.0557 | 0.1324 | 0.0692 | 0.8391 | 0.0595 |
| LA | Area of fifth leaf (cm2) | 1052.67 ± 11.34 | 898.1 ± 73.44 | 594.13 ± 68.82 | 829.87 ± 79.35 | 563.44 | 1260.69 | 430.09 | 1194.51 | 0.1708 | 0.1189 | 0.0003 | 0.0685 | 0.013 |
| LDW | Leaf Dry Weight (g) | 66.79 ± 8.24 | 39.46 ± 0.82 | 47.69 ± 1.2 | 42.84 ± 1.27 | 22.64 | 97.41 | 23.37 | 89.95 | 0.0496 | 0.3147 | <0.0001 | 0.6744 | 0.1598 |
| LFW | Leaf Fresh Weight (g) | 666.67 ± 70.07 | 486.67 ± 38.75 | 561.67 ± 7.88 | 478 ± 28.38 | 235.44 | 1118.12 | 213.33 | 994.00 | 0.2912 | 0.4482 | <0.0001 | 0.1812 | 0.0004 |
| Li | Xylem sap [Li] (mg/L) | 0.09 ± 0.09 | 0.05 ± 0.05 | 0.18 ± 0.06 | 0.19 ± 0.05 | 0.00 | 0.18 | 0.00 | 0.32 | 0.0000 | 0.2978 | 0.0005 | 0.0093 | 0.0003 |
| Mg | Xylem sap [Mg] (mg/L) | 39.87 ± 4.24 | 20.44 ± 3.51 | 48.91 ± 13.91 | 62.56 ± 6.4 | 15.03 | 120.00 | 14.90 | 158.58 | 0.1763 | 0.2142 | 0.0001 | 0.5589 | 0.0015 |
| Mn | Xylem sap [Mn] (mg/L) | 0.68 ± 0.13 | 0.39 ± 0.04 | 0.73 ± 0.25 | 1.14 ± 0.12 | 0.26 | 3.94 | 0.25 | 3.26 | 0.1946 | 0.2906 | 0.0003 | 0.923 | <0.0001 |
| Mo | Xylem sap [Mo] (mg/L) | 0.01 ± 0.0033 | 0.02 ± 0.01 | 0.01 ± 0.01 | 0.02 ± 0.01 | 0.00 | 0.36 | 0.00 | 0.27 | 0.0773 | 0.1752 | 0.0294 | 0.5448 | 0.0466 |
| Na | Xylem sap [Na] (mg/L) | 4.04 ± 1.39 | 2.43 ± 0.39 | 4.51 ± 2.26 | 3.33 ± 2.73 | 1.52 | 34.14 | 0.71 | 29.21 | 0.3642 | 0.1961 | <0.0001 | 0.8547 | 0.1227 |
| P | Xylem sap [P] | 30.82 ± 9.56 | 23.76 ± 4.69 | 30.77 ± 9.13 | 42.27 ± 7.99 | 11.51 | 81.16 | 5.71 | 92.95 | 0.1535 | 0.2151 | 0.0001 | 0.314 | 0.0017 |
| S | Xylem sap [S] (mg/L) | 40.6 ± 8.49 | 24.4 ± 3.33 | 57.22 ± 19.81 | 75.52 ± 20.7 | 16.04 | 137.62 | 2.97 | 183.80 | 0.1971 | 0.2894 | <0.0001 | 0.5098 | <0.0001 |
| Sb | Xylem sap [Sb] (mg/L) | 0.01 ± 0.01 | 0 ± 0 | 0.0033 ± 0.0033 | 0 ± 0 | 0.00 | 0.15 | –0.01 | 0.11 | 0.0000 | 0.1722 | 0.0866 | 0.5837 | 0.4646 |
| Se | Xylem sap [Se] (mg/L) | 0.04 ± 0.04 | 0.04 ± 0.03 | 0.03 ± 0.02 | 0.01 ± 0.01 | 0.00 | 0.57 | 0.00 | 0.13 | 0.1869 | 0.0594 | <0.0001 | 0.0134 | 0.0011 |
| ShDW | Total Shoot Dry Weight (g) | 117.47 ± 13.9 | 68.13 ± 4.45 | 89.67 ± 2.69 | 74.83 ± 1.53 | 44.90 | 164.19 | 44.01 | 141.53 | 0.1299 | 0.3449 | <0.0001 | 0.4472 | 0.1096 |
| ShFW | Total Shoot Fresh Weight (g) | 1203 ± 125.05 | 849.67 ± 90.71 | 1056.67 ± 5.93 | 860 ± 46.36 | 430.58 | 2087.98 | 414.00 | 1669.82 | 0.2858 | 0.4469 | <0.0001 | 0.1178 | 0.0003 |
| ShWC | Shoot water content (g) | 1085.55 ± 111.74 | 781.54 ± 86.32 | 967.01 ± 5.73 | 785.17 ± 47.32 | 385.96 | 1923.97 | 369.91 | 1536.15 | 0.2975 | 0.4492 | <0.0001 | 0.1063 | 0.0002 |
| ShWp | Shoot water proportion | 0.9 ± 0.0033 | 0.92 ± 0.0033 | 0.91 ± 0.0033 | 0.91 ± 0.0033 | 0.89 | 0.93 | 0.88 | 0.93 | 0.2000 | 0.2408 | <0.0001 | 0.024 | 0.1937 |
| SPAD | SPAD readings | 47.23 ± 0.54 | 44.83 ± 0.88 | 45.67 ± 0.38 | 43.77 ± 0.87 | 39.00 | 51.80 | 31.97 | 50.73 | 0.6843 | 0.3698 | <0.0001 | 0.007 | <0.0001 |
| Sr | Xylem sap [Sr] (mg/L) | 0.33 ± 0.04 | 0.21 ± 0.04 | 0.43 ± 0.12 | 0.65 ± 0.05 | 0.14 | 1.31 | 0.13 | 1.53 | 0.1944 | 0.2579 | 0.0003 | 0.8104 | <0.0001 |
| tZ | Xylem sap [trans–Zeatin] | 17.02 ± 3.33 | 9.88 ± 2.35 | 13.97 ± 2.42 | 8.34 ± 2 | 5.17 | 29.39 | 5.07 | 32.72 | 0.1809 | 0.0565 | 0.0006 | 0.5577 | 0.0059 |
| Zn | Xylem sap [Zn] (mg/L) | 0.46 ± 0.06 | 0.28 ± 0.06 | 0.42 ± 0.09 | 0.57 ± 0.03 | 0.15 | 1.03 | 0.17 | 1.47 | 0.1510 | 0.1742 | 0.0004 | 0.3494 | 0.0172 |
| ZR | Xylem sap [trans–Zeatin Riboside] | 12.6 ± 0.82 | 9.88 ± 0.18 | 8.59 ± 1.13 | 2.28 ± 2.28 | 0.00 | 12.34 | 0.11 | 12.56 | 0.1039 | 0.1369 | 0.0108 | 0.0888 | 0.0162 |
| Cluster | QTL | Chr. | Position | SNP | LOD | PEV | a |
|---|---|---|---|---|---|---|---|
| ABA_WD_11 | 11 | 75.154 | 56,353–32,118 | 2.04 | 5.8 | 0.97 | |
| ABA_WD_2.1 | 2 | 8.856 | 36,400 | 3.64 | 10.7 | 1.39 | |
| ABA_WD_2.2 | 2 | 76.525 | 29,914 | 2.91 | 8.5 | −1.14 | |
| XII | ABA_WD_5 | 5 | 108.514 | 339–354 | 2.9 | 8.4 | −1.15 |
| B_C_3 | 3 | 64.335 | 35,397–35,459 | 2.48 | 6.1 | 0.04 | |
| B_C_5 | 5 | 62.107 | 51,061 | 2.12 | 5.2 | −0.04 | |
| B_C_6 | 6 | 10.733 | 65,686 | 2.3 | 5.7 | 0.04 | |
| VI | B_WD_6 | 6 | 96.06 | C2_At1g20050_230_b | 3.44 | 11.4 | 0.05 |
| B_WD_9 | 9 | 115.383 | 63,663 | 2.44 | 7.9 | 0.04 | |
| IV | Ca_WD_4 | 4 | 28.574 | 41,577 | 2.82 | 7.5 | −33.11 |
| VI | Ca_WD_6 | 6 | 96.06 | C2_At1g20050_230_b | 2.43 | 6.4 | 29.42 |
| VIII | Ca_WD_8 | 8 | 11.65 | 56,575 | 3.48 | 9.4 | 35.43 |
| Cu_WD_1 | 1 | 42.436 | 59,885 | 5.67 | 19.4 | 0.04 | |
| FlN_C_4 | 4 | 39.43 | 17,956 | 5.01 | 13.4 | 0.46 | |
| FlN_C_5 | 5 | 60.926 | CL017527-0194 | 3.04 | 7.8 | 0.34 | |
| FlN_C_6 | 6 | 0.182 | 26,804 | 2.63 | 6.7 | −0.31 | |
| FlN_C_8 | 8 | 102.434 | 58,992 | 2.1 | 5.3 | 0.27 | |
| I | FlN_WD_1 | 1 | 47.252 | 51,462 | 2.97 | 4.9 | 0.26 |
| FlN_WD_11 | 11 | 40.373 | 9508 | 2.36 | 3.8 | 0.23 | |
| III | FlN_WD_3 | 3 | 52.869 | 30,678 | 3.22 | 5.3 | 0.26 |
| FlN_WD_4.1 | 4 | 39.43 | 17,956 | 4.14 | 6.9 | 0.49 | |
| V | FlN_WD_4.2 | 4 | 49.35 | 47,259 | 2.42 | 3.9 | −0.35 |
| FlN_WD_5 | 5 | 32.533 | 23,804 | 4.82 | 8.2 | 0.33 | |
| FlN_WD_8 | 8 | 86.586 | 65,161–65,188 | 2.37 | 3.5 | −1.67 | |
| IX | FlN_WD_9.1 | 9 | 18.634 | 57,922–57,900 | 2.72 | 4.4 | 0.25 |
| X | FlN_WD_9.2 | 9 | 93.838 | 36,845–69,503 | 2.59 | 4.2 | 0.25 |
| I | JA_C_1 | 1 | 50.094 | 50,470 | 2.41 | 6.5 | 7.11 |
| JA_C_7 | 7 | 30.461 | 68,008 | 2.14 | 5.7 | −6.39 | |
| LA_C_11 | 11 | 24.632 | 66,141 | 2.25 | 6.4 | 32.18 | |
| LA_C_4 | 4 | 3.085 | 21,317 | 2.74 | 7.8 | −36.11 | |
| LA_C_6 | 6 | 101.994 | 54,183 | 3.24 | 9.3 | −39.51 | |
| V | LDW_C_4 | 4 | 59.661 | 47,590 | 2.51 | 8.5 | 3.14 |
| IX | LDW_WD_9 | 9 | 21.4 | 39,886 | 2.32 | 7.6 | 3.44 |
| V | LFW_C_4 | 4 | 56.517 | 47,487 | 2.79 | 9.3 | 39.27 |
| X | LFW_WD_9 | 9 | 94.89 | 69,503 | 2.45 | 8.7 | 37.52 |
| Mg_C_3 | 3 | 29.052 | 55,002 | 2.26 | 8.1 | 6.72 | |
| XI | Mg_WD_10 | 10 | 22.598 | 17,838 | 2.24 | 5.7 | 8.14 |
| II | Mg_WD_2 | 2 | 46.39 | T0562-49,497 | 2.5 | 6.4 | 8.41 |
| IV | Mg_WD_4 | 4 | 26.383 | 16,978 | 2.15 | 5.5 | −7.79 |
| VI | Mg_WD_6 | 6 | 96.06 | C2_At1g20050_230_b | 2.88 | 7.4 | 8.97 |
| VII | Mg_WD_7 | 7 | 68.714 | 53,534 | 2.52 | 6.5 | −8.48 |
| XI | Mn_WD_10 | 10 | 22.598 | 17,838 | 3.61 | 8.8 | 0.19 |
| II | Mn_WD_2 | 2 | 46.39 | T0562-49,497 | 3.13 | 7.5 | 0.17 |
| IV | Mn_WD_4 | 4 | 26.174 | 41,648–41,623 | 2.65 | 6.3 | −0.16 |
| VII | Mn_WD_7 | 7 | 68.714 | 53,534 | 2.56 | 6.1 | −0.16 |
| IX | Mn_WD_9 | 9 | 19.634 | 57,922–57,900 | 2.62 | 6.2 | −0.16 |
| Na_C_7 | 7 | 38.818 | 67,869 | 12.26 | 36.8 | −3.49 | |
| Na_WD_11 | 11 | 89.544 | 56,158 | 4.09 | 11.4 | 2.08 | |
| Na_WD_7 | 7 | 38.818 | 67,869 | 6.23 | 18.2 | −2.65 | |
| XI | P_WD_10 | 10 | 24.053 | 46,416 | 2.76 | 7.8 | 5.12 |
| VI | P_WD_6 | 6 | 95.563 | 2819_5_183_b | 4.06 | 11.8 | 6.43 |
| VI | S_WD_6 | 6 | 96.06 | C2_At1g20050_230_b | 3.86 | 10.2 | 13.00 |
| VIII | S_WD_8 | 8 | 11.65 | 56,575 | 2.2 | 5.6 | 9.46 |
| V | ShDW_C_4 | 4 | 56.517 | 47,487 | 2.5 | 8.6 | 5.19 |
| III | ShDW_DW_3 | 3 | 52.651 | 30,704 | 3.16 | 9 | 5.55 |
| V | ShFW_C_4 | 4 | 56.517 | 47,487 | 2.39 | 8.4 | 68.79 |
| X | ShFW_WD_9 | 9 | 83.507 | 58,234 | 2.41 | 8.6 | 61.68 |
| V | ShWC_C_4 | 4 | 56.517 | 47,487 | 3.05 | 9.8 | 68.25 |
| ShWC_WD_1 | 1 | 12.421 | 60,303 | 2.33 | 6 | −49.63 | |
| ShWC_WD_3 | 3 | 91.561 | 62,473 | 2.26 | 5.7 | −52.09 | |
| V | ShWC_WD_4 | 4 | 63.868 | 3872 | 2.81 | 7.3 | 53.67 |
| X | ShWC_WD_9 | 9 | 94.89 | 69,503 | 3.94 | 10.4 | 64.86 |
| ShWp_C_12 | 12 | 88.198 | 32,743 | 2.9 | 7.1 | 0.00 | |
| ShWp_C_3.1 | 3 | 4.081 | 63,211 | 4.17 | 10.4 | 0.00 | |
| ShWp_C_3.2 | 3 | 90.075 | 62,473 | 2.4 | 5.8 | 0.00 | |
| ShWp_C_9 | 9 | 103.792 | 69,686–63,663 | 3.22 | 7.9 | 0.00 | |
| ShWp_WD_1 | 1 | 78.318 | 27,588–27,600 | 2.76 | 6.8 | 0.00 | |
| ShWp_WD_6.1 | 6 | 51.444 | 27,197 | 2.16 | 5.2 | 0.00 | |
| ShWp_WD_6.2 | 6 | 60.181 | 42,119 | 6.5 | 17.2 | −0.01 | |
| ShWp_WD_6.3 | 6 | 74.588 | 57,435 | 2.91 | 7.2 | 0.00 | |
| X | ShWp_WD_9 | 9 | 84.784 | 58,253 | 2.97 | 7.3 | 0.00 |
| Sr_C_1 | 1 | 152.18 | SGN-U313729_snp305 | 3.58 | 10.8 | −0.25 | |
| Sr_C_9 | 9 | 32.383 | 39,735 | 2.43 | 7.2 | 0.08 | |
| IV | Sr_WD_4 | 4 | 26.383 | 16,978 | 2.94 | 7.7 | −0.08 |
| VI | Sr_WD_6 | 6 | 96.06 | C2_At1g20050_230_b | 3.11 | 8.1 | 0.08 |
| VIII | Sr_WD_8 | 8 | 11.65 | 56,575 | 2.31 | 5.9 | 0.07 |
| tZ_C_11 | 11 | 52.492 | 2966 | 2.3 | 8.3 | −1.45 | |
| II | Zn_WD_2 | 2 | 46.39 | T0562-49,497 | 2.69 | 6.8 | 0.07 |
| VI | Zn_WD_6 | 6 | 96.06 | C2_At1g20050_230_b | 2.94 | 7.4 | 0.16 |
| (XI) | ZR_WD_10 | 10 | 45.374 | 52,018 | 3.42 | 9.3 | 0.86 |
| V | ZR_WD_4 | 4 | 48.186 | 47,229 | 2.42 | 5.9 | 0.89 |
| XII | ZR_WD_5 | 5 | 110.72 | 37,819 | 2.31 | 6.2 | −0.72 |
| (IX) | ZR_WD_9 | 9 | 2.545 | 14,673–58,096 | 2.81 | 7.6 | −0.83 |
| Cluster/QTL | Mut | Exp | Start | Annotation | mRNA | Ord |
|---|---|---|---|---|---|---|
| Sr_C_1 | M | 97,531,670 | Magnesium transporter NIPA2 | Solyc01g111190.2.1 | 6 | |
| Sr_C_9 | N | 3,902,289 | Cation/H+ antiporter | Solyc09g010530.2.1 | 53 | |
| VIII | L | 876,173 | ChaC cation transport regulator-like 1 | Solyc08g006150.2.1 | 38 | |
| VIII | Max | 1,096,757 | Glutamate-gated kainate-type ion channel receptor | Solyc08g006500.2.1 | 4 | |
| VIII | L5 | Max | 2,169,906 | Subtilisin-like protease (4 copies in tandem) | Solyc08g007670.1.1 | 113 |
| VIII | Max | 2,541,278 | Aquaporin (aquaporin PIP-type pTOM75) | Solyc08g008050.2.1 | 149 | |
| VIII | L5 | VL | 2,654,348 | Cation/H(+) antiporter 18 | Solyc08g008190.2.1 | 162 |
| VIII | L5 | N | 2,658,313 | Cation/H(+) antiporter 18 | Solyc08g008200.1.1 | 163 |
| VI | H | 47,496,433 | Zinc-transporter-like protein | Solyc06g076440.1.1 | 151 | |
| VI | L5 | Max | 47,705,394 | Cytochrome P450 86A1 | Solyc06g076800.2.1 | 117 |
| VI | L5 | Max | 48,070,379 | SAGA-associated factor 11 homolog | Solyc06g082160.2.1 | 80 |
| VI | Med | 48,142,943 | Cation/H(+) antiporter 14 | Solyc06g082230.2.1 | 73 | |
| VI | L5 | Max | 48,687,538 | E3 ubiquitin-protein ligase CHIP | Solyc06g083150.2.1 | 17 |
| VI | L5 | Max | 49,364,970 | High affinity sulfate transporter 1 | Solyc06g084140.2.1 | 116 |
| IV | L5 | Max | 4,029,691 | ER glycerol-phosphate acyltransferase | Solyc04g011600.2.1 | 110 |
| IV | E9 | Max | 4,150,679 | BZIP transcription factor | Solyc04g011670.2.1 | 103 |
| IV | L5 | N | 4,226,837 | Glutaredoxin (2 copies) | Solyc04g011780.1.1 | 92 |
| IV | Max | 4,376,146 | Ethylene responsive transcription factor 2a (ABR1-like) | Solyc04g012050.2.1 | 66 | |
| IV | E9 | VL | 4,872,992 | Universal stress protein (PHOS32) | Solyc04g014600.2.1 | 9 |
| IV | M | 5,226,676 | Heavy metal transport/detoxification protein (2 copies) | Solyc04g015020.2.1 | 0 | |
| IV | L5 | N | 5,133,669 | Outward rectifying potassium channel | Solyc04g014880.2.1 | 0 |
| IV | L5 | Max | 5,458,419 | NRC1 | Solyc04g015250.1.1 | 6 |
| IV | H | 5,491,465 | MYB transcription factor (protein PHR1-LIKE 3) | Solyc04g015290.2.1 | 10 | |
| IV | VL | 6,590,404 | Cation/H(+) antiporter 28 | Solyc04g015990.1.1 | 78 | |
| XI | Max | 763,795 | Ca-activated outward-rectifying potassium channel 1 | Solyc10g006010.2.1 | 128 | |
| XI | Max | 950,380 | Purple acid phosphatase | Solyc10g006300.2.1 | 99 | |
| XI | L5 | H | 1,911,958 | Organic anion transporter | Solyc10g007610.2.1 | 2 |
| XI | L5 | Max | 1,919,334 | Glutathione S-transferase (2 copies in tandem) | Solyc10g007620.1.1 | 3 |
| XI | H | 1,981,130 | Mg2+ transporter protein CorA family protein | Solyc10g007740.2.1 | 15 | |
| VII | E9 | M | 60,468,833 | MAPKKK54 | Solyc07g051920.1.1 | 107 |
| VII | VL | 60,504,547 | C4-dicarboxylate transporter/malic acid transport family | Solyc07g051950.2.1 | 104 | |
| VII | H | 60,921,179 | Glutamate-gated kainate-type ion channel receptor (2 copies) | Solyc07g052400.2.1 | 62 | |
| VII | Max | 62,362,157 | Two-pore calcium channel 2 | Solyc07g053970.2.1 | 92 | |
| II | M | 44,832,287 | Copper transport protein 86 (ataxin-10) | Solyc02g080650.2.1 | 8 | |
| II | Max | 45,468,042 | Yellow stripe-like protein 2.1 (YSL7) | Solyc02g081570.2.1 | 100 | |
| II | M | 45,492,312 | Zinc transporter protein (zinc transporter 3) | Solyc02g081600.2.1 | 103 | |
| II | M | 45,804,741 | High affinity copper uptake protein (Cu transporter 5) | Solyc02g082080.1.1 | 151 | |
| IX | L5 | N | 1,176,696 | MYB transcription factor (SRM1-like) | Solyc09g007580.1.1 | 79 |
| IX | M | 1,401,117 | Mn transporter mntH (ethylene signaling protein) | Solyc09g007870.2.1 | 50 | |
| IX | H | 1,607,740 | Membrane magnesium transporter 1 | Solyc09g008140.2.1 | 23 | |
| IX | Max | 1,718,596 | MYB transcription factor 38 (protein blind-like1, bli1) | Solyc09g008250.2.1 | 12 | |
| V | L5 | Max | 63,103,059 | Cytochrome P450 CYP2 subfamily (3 copies in tandem) | Solyc04g078340.2.1 | 15 |
| V | Max | 63,259,221 | WRKY transcription factor 7 | Solyc04g078550.2.1 | 6 | |
| V | L5 | Max | 63,627,437 | Anthocyanidin 3-O-glucosyltransferase | Solyc04g079030.2.1 | 54 |
| V | Max | 63,744,550 | Gibberellin receptor GID1L2 | Solyc04g079190.2.1 | 70 | |
| V | L5 | M | 63,888,688 | MYB transcription factor 77 | Solyc04g079360.1.1 | 87 |
| V | L5 | L | 64,050,601 | Cytochrome P450 CYP2 subfamily | Solyc04g079680.2.1 | 115 |
| V | L5 | M | 64,193,977 | Seryl-tRNA synthetase | Solyc04g079870.1.1 | 134 |
| XII | L5 | M | 63,229,667 | Zeatin O-β-D-xylosyltransferase | Solyc05g053120.1.1 | 223 |
| XII | M | 63,498,355 | Zeatin O-β-D-xylosyltransferase | Solyc05g053400.1.1 | 195 | |
| XII | M | 63,610,661 | 9-cis-epoxycarotenoid dioxygenase 6 | Solyc05g053530.1.1 | 182 | |
| XII | M | 64,775,972 | Cysteine desulfurase, molybdenum cofactor biosynthesis | Solyc05g055000.2.1 | 36 | |
| XII | M | 64,859,523 | Ethylene receptor | Solyc05g055070.2.1 | 29 | |
| XII | L5 | Max | 64,872,981 | Ubiquitin carboxyl-terminal hydrolase family | Solyc05g055090.2.1 | 27 |
| XII | Max | 65,269,467 | Isopentenyl-diphosphate delta-isomerase | Solyc05g055760.2.1 | 34 | |
| XII | L5 | Max | 65,399,111 | Serine/threonine-protein phosphatase (MAIN-LIKE 2) | Solyc05g056010.2.1 | 59 |
| ZR_WD_9 | L5 | Max | 21,705 | MYB transcription factor-like | Solyc09g005030.1.1 | 109 |
| ZR_WD_9 | VL | 1,356,808 | Cytokinin riboside 5’-monophosphate phosphoribohydrolase LOG3 | Solyc09g007830.2.1 | 70 | |
| ZR_WD_9 | L5 | Max | 2,036,208 | NCS1 family transporter | Solyc09g008550.2.1 | 142 |
| ABA_WD_2.1 | N | 36,230,064 | Short-chain dehydrogenase/reductase family protein | Solyc02g065060.2.1 | 46 | |
| ABA_WD_2.1 | L5 | N | 36,835,375 | Glucosyltransferase-like protein | Solyc02g065670.1.1 | 13 |
| ABA_WD_2.1 | E9 | VL | 37,817,307 | Glucosyltransferase-like protein | Solyc02g067690.2.1 | 116 |
| ABA_WD_11 | N | 54,668,129 | Histidine phosphotransfer protein | Solyc11g070150.1.1 | 49 | |
| ABA_WD_11 | L5 | Max | 55,006,926 | Aldehyde oxidase (abscisic-aldehyde oxidase-like) | Solyc11g071580.1.1 | 93 |
| ABA_WD_11 | L5 | Max | 55,013,280 | Aldehyde oxidase (abscisic-aldehyde oxidase-like) | Solyc11g071590.1.1 | 94 |
| ABA_WD_11 | N | 55,049,892 | Sensor histidine kinase LuxQ | Solyc11g071630.1.1 | 98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asins, M.J.; Albacete, A.; Martínez-Andújar, C.; Celiktopuz, E.; Solmaz, İ.; Sarı, N.; Pérez-Alfocea, F.; Dodd, I.C.; Carbonell, E.A.; Topcu, S. Genetic Analysis of Root-to-Shoot Signaling and Rootstock-Mediated Tolerance to Water Deficit in Tomato. Genes 2021, 12, 10. https://doi.org/10.3390/genes12010010
Asins MJ, Albacete A, Martínez-Andújar C, Celiktopuz E, Solmaz İ, Sarı N, Pérez-Alfocea F, Dodd IC, Carbonell EA, Topcu S. Genetic Analysis of Root-to-Shoot Signaling and Rootstock-Mediated Tolerance to Water Deficit in Tomato. Genes. 2021; 12(1):10. https://doi.org/10.3390/genes12010010
Chicago/Turabian StyleAsins, Maria J., Alfonso Albacete, Cristina Martínez-Andújar, Eser Celiktopuz, İlknur Solmaz, Nebahat Sarı, Francisco Pérez-Alfocea, Ian C. Dodd, Emilio A. Carbonell, and Sevilay Topcu. 2021. "Genetic Analysis of Root-to-Shoot Signaling and Rootstock-Mediated Tolerance to Water Deficit in Tomato" Genes 12, no. 1: 10. https://doi.org/10.3390/genes12010010
APA StyleAsins, M. J., Albacete, A., Martínez-Andújar, C., Celiktopuz, E., Solmaz, İ., Sarı, N., Pérez-Alfocea, F., Dodd, I. C., Carbonell, E. A., & Topcu, S. (2021). Genetic Analysis of Root-to-Shoot Signaling and Rootstock-Mediated Tolerance to Water Deficit in Tomato. Genes, 12(1), 10. https://doi.org/10.3390/genes12010010