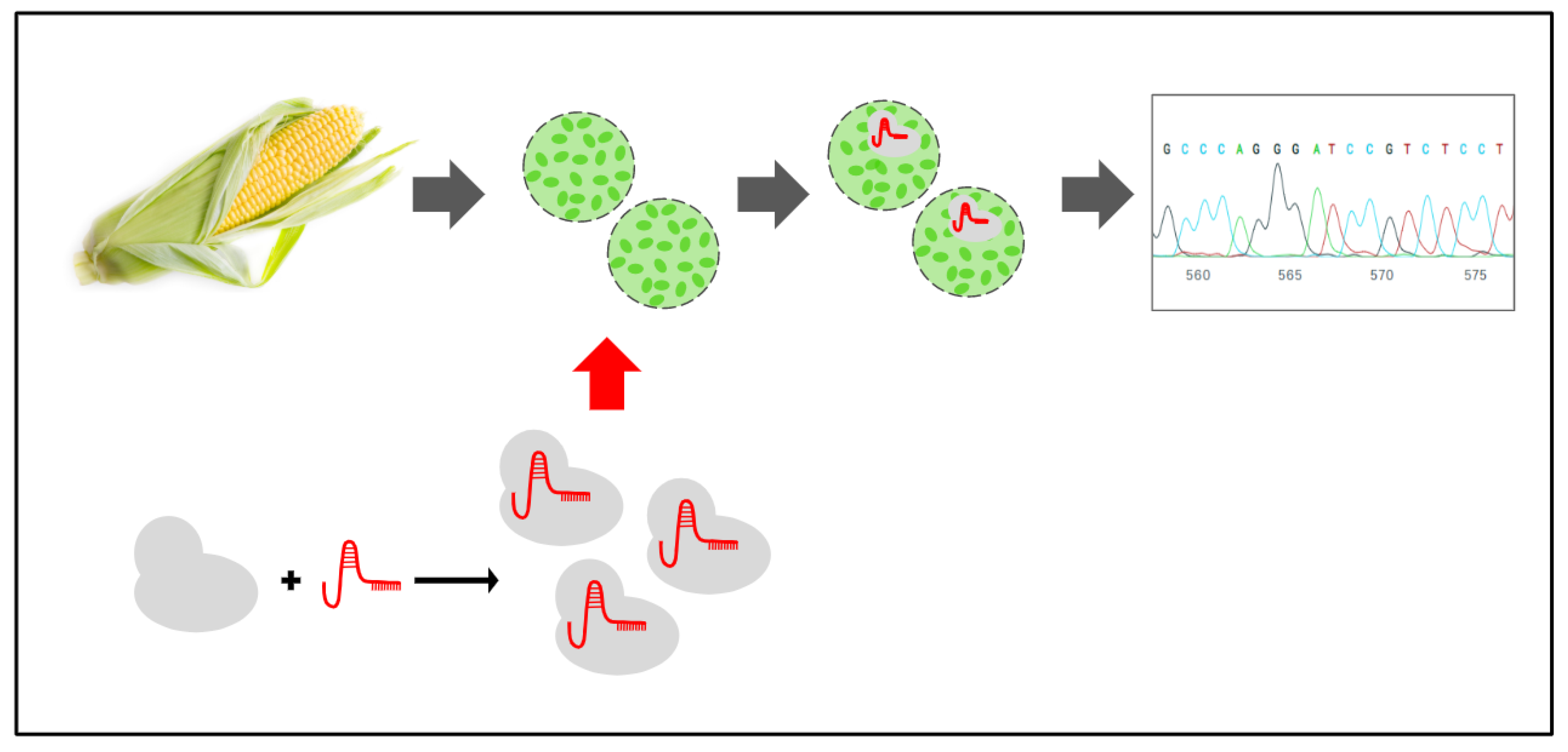
PEG-Delivered CRISPR-Cas9 Ribonucleoproteins System for Gene-Editing Screening of Maize Protoplasts

 ,
,  ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Target Site Selection and In Vitro Cleavage Assay
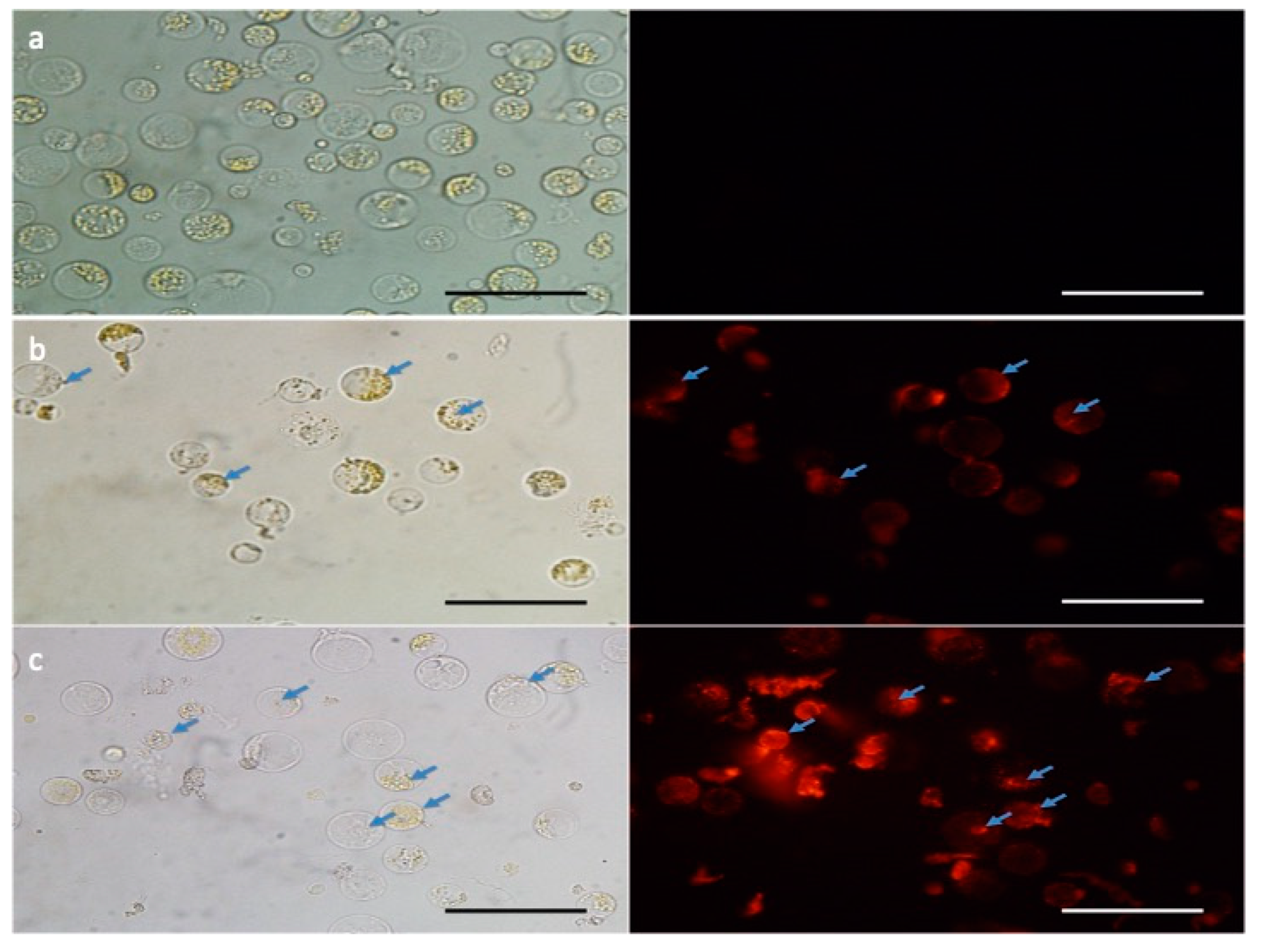
2.2. Maize Protoplast Isolation and Fluorescent Transfection Assay
2.3. Maize Protoplast Transformation
2.4. Gene-Editing Efficiency Analysis by Sanger Sequencing
3. Results
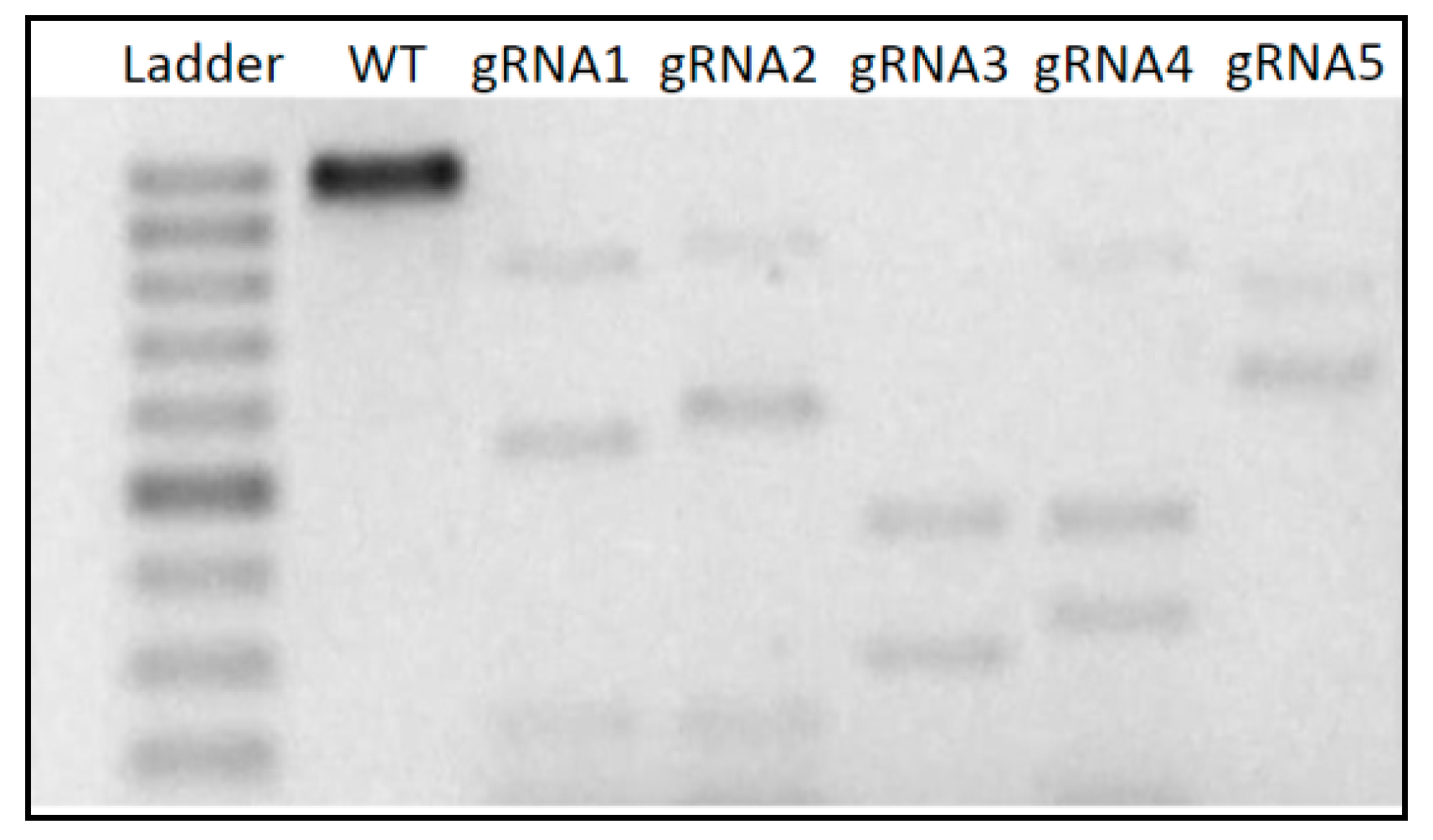
3.1. In Vitro Cleavage Assay
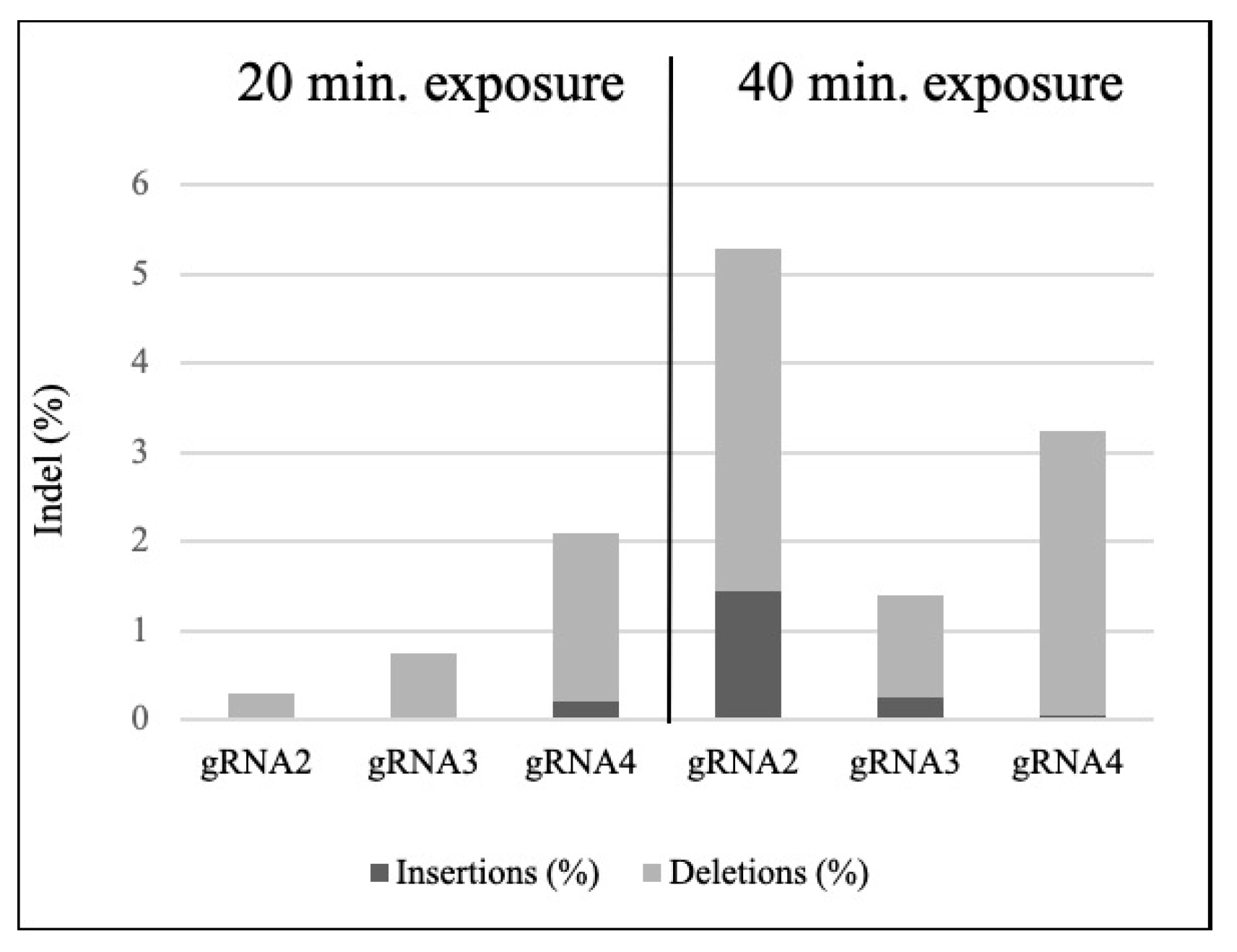
3.2. Targeted Mutagenesis in Maize Using CRISPR-Cas9 Ribonucleoproteins
4. Discussion
4.1. Ribonucleoprotein Delivery in Plants
4.2. CRISPR Delivery Methods in Maize
4.3. Analytical Platforms for Gene-Editing Detection
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.D.; Scott, D.; Weinstein, J.A.; Ran, F.A.; Konermann, S.; Agarwala, V.; Li, Y.; Fine, E.J.; Wu, X.; Shalem, O.; et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 2013, 31, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Bortesi, L.; Fischer, R. The CRISPR/Cas9 system for plant genome editing and beyond. Biotechnol. Adv. 2015, 33, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Grobman, A.; Bonavia, D. Origin, Domestication, and Evolution of Maize; Cambridge University Press (CUP): Cambridge, UK, 2013; pp. 329–486. [Google Scholar]
- Li, H.; Wang, M.; Li, W.; He, L.; Zhou, Y.; Zhu, J.; Che, R.; Warburton, M.L.; Yang, X.; Yan, J. Genetic variants and underlying mechanisms influencing variance heterogeneity in maize. Plant J. 2020, 103, 1089–1102. [Google Scholar] [CrossRef] [PubMed]
- Barrangou, R.; Doudna, J. Applications of CRISPR technologies in research and beyond. Nat. Biotechnol. 2016, 34, 933–941. [Google Scholar] [CrossRef]
- Xie, S.; Shen, B.; Zhang, C.; Huang, X.; Zhang, Y. sgRNAcas9: A software package for designing CRISPR sgRNA and evaluating potential off-target cleavage sites. PLoS ONE 2014, 9, e100448. [Google Scholar] [CrossRef]
- Metje-Sprink, J.; Menz, J.; Modrzejewski, D.; Sprink, T. DNA-free genome editing: Past, present and future. Front. Plant Sci. 2019, 9. [Google Scholar] [CrossRef]
- Liu, Q.; Yuan, Y.; Zhu, F.; Hong, Y.; Ge, R. Efficient genome editing using CRISPR/Cas9 ribonucleoprotein approach in cultured Medaka fish cells. Biol. Open 2018, 7, bio035170. [Google Scholar] [CrossRef]
- Kanchiswamy, C.N. DNA-free genome editing methods for targeted crop improvement. Plant Cell Rep. 2016, 35, 1469–1474. [Google Scholar] [CrossRef]
- Agarwal, A.; Yadava, P.; Kumar, K.; Singh, I.; Kaul, T.; Pattanayak, A.; Agrawal, P.K. Insights into maize genome editing via CRISPR/Cas9. Physiol. Mol. Biol. Plants 2018, 24, 175–183. [Google Scholar] [CrossRef]
- Woo, J.W.; Kim, J.; Kwon, S.I.; Corvalan, C.; Cho, S.W.; Kim, H.; Kim, S.-G.; Kim, S.T.; Choe, S.; Kim, J.-S. DNA-free genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins. Nat. Biotechnol. 2015, 33, 1162–1164. [Google Scholar] [CrossRef] [PubMed]
- Malnoy, M.; Viola, R.; Jung, M.-H.; Koo, O.-J.; Kim, S.; Kim, J.-S.; Velasco, R.; Kanchiswamy, C.N. DNA-free genetically edited grapevine and apple protoplast using CRISPR/Cas9 Ribonucleoproteins. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Subburaj, S.; Chung, S.J.; Lee, C.; Ryu, S.-M.; Kim, D.H.; Kim, J.-S.; Bae, S.; Lee, G. Site-directed mutagenesis in Petunia × hybrida protoplast system using direct delivery of purified recombinant Cas9 ribonucleoproteins. Plant Cell Rep. 2016, 35, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Turesson, H.; Olsson, N.; Fält, A.-S.; Ohlsson, P.; González, M.N.; Samuelsson, M.; Hofvander, P.; Olsson, P. Genome editing in potato via CRISPR-Cas9 ribonucleoprotein delivery. Physiol. Plant. 2018, 164, 378–384. [Google Scholar] [CrossRef]
- Svitashev, S.; Schwartz, C.; Lenderts, B.; Young, J.K.; Cigan, A.M. Genome editing in maize directed by CRISPR–Cas9 ribonucleoprotein complexes. Nat. Commun. 2016, 7, 13274. [Google Scholar] [CrossRef]
- Liang, Z.; Zong, Y.; Gao, C. An efficient targeted mutagenesis system using CRISPR/Cas in monocotyledons. Curr. Protoc. Plant Biol. 2016, 1, 329–344. [Google Scholar] [CrossRef]
- Char, S.N.; Neelakandan, A.; Nahampun, H.; Frame, B.; Main, M.; Spalding, M.H.; Becraft, P.W.; Meyers, B.C.; Walbot, V.; Wang, K.; et al. An agrobacterium-delivered CRISPR/Cas9 system for high-frequency targeted mutagenesis in maize. Plant Biotechnol. J. 2016, 15, 257–268. [Google Scholar] [CrossRef]
- Feng, C.; Su, H.; Bai, H.; Wang, R.; Liu, Y.; Guo, X.; Liu, C.; Zhang, J.; Yuan, J.; Birchler, J.A.; et al. High-efficiency genome editing using a dmc1 promoter-controlled CRISPR/Cas9 system in maize. Plant Biotechnol. J. 2018, 16, 1848–1857. [Google Scholar] [CrossRef]
- Dong, L.; Qi, X.; Zhu, J.; Liu, C.; Zhang, X.; Cheng, B.; Mao, L.; Xie, C. Supersweet and waxy: Meeting the diverse demands for specialty maize by genome editing. Plant Biotechnol. J. 2019, 17, 1853–1855. [Google Scholar] [CrossRef]
- Sheen, J. Molecular mechanisms underlying the differential expression of maize pyruvate, orthophosphate dikinase genes. Plant Cell 1991, 3, 225–245. [Google Scholar]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [PubMed]
- Hsiau, T.; Conant, D.; Rossi, N.; Maures, T.; Waite, K.; Yang, J.; Joshi, S.; Kelso, R.; Holden, K.; Enzmann, B.L.; et al. Inference of crispr edits from sanger trace data. bioRxiv 2019, 251082. [Google Scholar] [CrossRef]
- Svitashev, S.; Young, J.K.; Schwartz, C.; Gao, H.; Falco, S.C.; Cigan, A.M. Targeted mutagenesis, precise gene editing, and site-specific gene insertion in maize using Cas9 and guide RNA. Plant Physiol. 2015, 169, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-S.; Hsu, C.; Yang, L.; Lee, L.; Fu, J.; Cheng, Q.; Wu, F.; Hsiao, H.C.-W.; Zhang, Y.; Zhang, R.; et al. Application of protoplast technology to CRISPR/Cas9 mutagenesis: From single-cell mutation detection to mutant plant regeneration. Plant Biotechnol. J. 2018, 16, 1295–1310. [Google Scholar] [CrossRef] [PubMed]
- Bortesi, L.; Zhu, C.; Zischewski, J.; Perez, L.; Bassié, L.; Nadi, R.; Forni, G.; Lade, S.B.; Soto, E.; Jin, X.; et al. Patterns of CRISPR/Cas9 activity in plants, animals and microbes. Plant Biotechnol. J. 2016, 14, 2203–2216. [Google Scholar] [CrossRef]
- Kim, S.; Kim, D.; Cho, S.W.; Kim, J.; Kim, J.; Kim, J.-S. Highly efficient RNA-guided genome editing in human cells via delivery of purified Cas9 ribonucleoproteins. Genome Res. 2014, 24, 1012–1019. [Google Scholar] [CrossRef]
- Braatz, J.; Harloff, H.-J.; Mascher, M.; Stein, N.; Himmelbach, A.; Harloff, H.-J. CRISPR-Cas9 targeted mutagenesis leads to simultaneous modification of different homoeologous gene copies in polyploid oilseed rape (Brassica napus). Plant Physiol. 2017, 174, 935–942. [Google Scholar] [CrossRef]
- Sandhya, D.; Jogam, P.; Allini, V.R.; Abbagani, S.; Alok, A. The present and potential future methods for delivering CRISPR/Cas9 components in plants. J. Genet. Eng. Biotechnol. 2020, 18, 1–11. [Google Scholar] [CrossRef]
- Wada, N.; Ueta, R.; Osakabe, Y.; Osakabe, K. Precision genome editing in plants: State-of-the-art in CRISPR/Cas9-based genome engineering. BMC Plant Biol. 2020, 20, 1–12. [Google Scholar] [CrossRef]
- Liang, Z.; Chen, K.; Li, T.; Zhang, Y.; Wang, Y.; Zhao, Q.; Liu, J.; Zhang, H.; Liu, C.; Ran, Y.; et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat. Commun. 2017, 8, 14261. [Google Scholar] [CrossRef]
- Murovec, J.; Guček, K.; Bohanec, B.; Avbelj, M.; Jerala, R. DNA-free genome editing of brassica oleracea and B. rapa protoplasts using CRISPR-Cas9 Ribonucleoprotein complexes. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Toda, E.; Koiso, N.; Takebayashi, A.; Ichikawa, M.; Kiba, T.; Osakabe, K.; Osakabe, Y.; Sakakibara, H.; Kato, N.; Okamoto, T. An efficient DNA- and selectable-marker-free genome-editing system using zygotes in rice. Nat. Plants 2019, 5, 363. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Zhang, K.; Chen, K.; Gao, C. Targeted Mutagenesis in Zea mays Using TALENs and the CRISPR/Cas System. J. Genet. Genomics 2014, 41, 63–68. [Google Scholar] [CrossRef]
- Xing, H.; Dong, L.; Wang, Z.; Zhang, H.; Han, C.; Liu, B.; Wang, X.; Chen, Q. A CRISPR/Cas9 toolkit for multiplex genome editing in plants. BMC Plant Biol. 2014, 14, 327. [Google Scholar] [CrossRef]
- Qi, W.; Zhu, T.; Tian, Z.; Li, C.; Zhang, W.; Song, R. High-efficiency CRISPR/Cas9 multiplex gene editing using the glycine tRNA-processing system-based strategy in maize. BMC Biotechnol. 2016, 16, 58. [Google Scholar] [CrossRef]
- Zhu, J.; Song, N.; Sun, S.; Yang, W.; Zhao, H.; Song, W.; Lai, J. Efficiency and inheritance of targeted mutagenesis in maize using CRISPR-Cas9. J. Genet. Genomics 2016, 43, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Gao, H.; Wang, H.; Lafitte, H.R.; Archibald, R.L.; Yang, M.; Hakimi, S.M.; Mo, H.; Habben, J.E. ARGOS8 variants generated by CRISPR-Cas9 improve maize grain yield under field drought stress conditions. Plant Biotechnol. J. 2017, 15, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Xu, Q.; Liu, Y.; Zhang, J.; Ren, D.; Wang, G.; Liu, Y. Generation of transgene-free maize male sterile lines using the CRISPR/Cas9 system. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Zhang, Y.; Kleinstiver, B.P.; Guo, J.A.; Aryee, J.; Miller, J.; Malzahn, A.; Zarecor, S.; Lawrence-Dill, C.J.; Joung, J.L.; et al. Activities and specificities of CRISPR/Cas9 and Cas12a nucleases for targeted mutagenesis in maize. Plant Biotechnol. J. 2019, 17, 362–372. [Google Scholar] [CrossRef]
- Kelliher, T.; Starr, D.; Su, X.; Tang, G.; Chen, Z.; Carter, J.; Wittich, P.E.; Dong, S.; Green, J.; Burch, E. One-step genome editing of elite crop germplasm during haploid induction. Nat. Biotechnol. 2019, 37, 287–292. [Google Scholar] [CrossRef]
- Doll, N.M.; Gilles, L.M.; Gérentes, M.F.; Richard, C.; Just, J.; Fierlej, Y.; Borrelli, V.G.M.; Gendrot, G.; Ingram, C.G.; Rogowsky, P.M. Single and multiple gene knockouts by CRISPR–Cas9 in maize. Plant Cell Rep. 2019, 38, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Xu, F.; Liu, L.; Char, S.N.; Ding, Y.; Je, B.I.; Schmelz, E.; Yang, B.; Jackson, D. The maize heterotrimeric G protein β subunit controls shoot meristem development and immune responses. Proc. Natl. Acad. Sci. USA 2019, 117, 1799–1805. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Jian, L.; Xu, J.; Zhang, Q.; Zhang, M.; Jin, M.; Peng, Y.; Yan, J.; Han, B.; Liu, J.; et al. High-throughput CRISPR/Cas9 mutagenesis streamlines trait gene identification in maize. Plant Cell 2020, 32, 1397–1413. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Gadlage, M.J.; Lafitte, H.R.; Lenderts, B.; Yang, M.; Schroder, M.; Farrell, J.; Snopek, K.; Peterson, D.; Feigenbutz, L.; et al. Superior field performance of waxy corn engineered using CRISPR–Cas9. Nat. Biotechnol. 2020, 38, 579–581. [Google Scholar] [CrossRef]
- Gao, H.; Mutti, J.; Young, J.K.; Yang, M.; Schroder, M.; Lenderts, B.; Wang, L.; Peterson, D.; Clair, G.S.; Jones, S.; et al. Complex trait loci in maize enabled by CRISPR-Cas9 mediated gene insertion. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Barone, P.; Wu, E.; Lenderts, B.; Anand, A.; Gordon-Kamm, W.; Svitashev, S.; Kumar, S. Efficient gene targeting in maize using inducible CRISPR-Cas9 and marker-free donor template. Mol. Plant 2020, 13. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.; Chen, R.; Yang, L.; Fan, K.; Liu, Y.; Wang, G.; Ren, Z.; Liu, Y. Generation of transgene-free Semidwarf maize plants by gene editing of gibberellin-oxidase20-3 using CRISPR/Cas9. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Park, J.; Choe, S. DNA-free genome editing with preassembled CRISPR/Cas9 ribonucleoproteins in plants. Transgenic Res. 2019, 28, 61–64. [Google Scholar] [CrossRef]
- Vu, G.T.H.; Cao, X.H.; Fauser, F.; Reiss, B.; Puchta, H.; Schubert, I. Endogenous sequence patterns predispose the repair modes of CRISPR /Cas9-induced DNA double-stranded breaks in Arabidopsis thaliana. Plant J. 2017, 92, 57–67. [Google Scholar] [CrossRef]
- Agapito-Tenfen, S.Z.; Okoli, A.S.; Bernstein, M.J.; Wikmark, O.-G.; Myhr, A.I. Revisiting risk governance of GM plants: The need to consider new and emerging gene-editing techniques. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′–3′) | Amplicon Size (bp) |
| ZmIPK-F | GAAGAAGCAGCAGAGCTTCA | 876 |
| ZmIPK-R | CAGAAGAAATCCGTGAGGACAG | |
| crRNA | Sequence (5′–3′) | Cleavage Fragments (bp) |
| crRNA1 | AGCTCGACCACGCCGCCGAC | 279 | 597 |
| crRNA2 * | GGGATCCGTCTCCTTCTCCC | 617 | 259 |
| crRNA3 * | ATCTTCAAGGTCTACGTCGT | 525 | 351 |
| crRNA4 * | CAGGAGTTCGTCAACCATGG | 498 |378 |
| crRNA5 | ACAAGCTCTACGGAGACGAC | 141 | 735 |
| Sample | Incubation Time (min) | % of Indel | Model Fit (R2) | KO Score | Mutation Range (bp) | Greater Contribution (bp) |
|---|---|---|---|---|---|---|
| Cas9 only | 20 | 0 | 0 | 0 | 0 | |
| gRNA2 only | 20 | 0 | 0 | 0 | 0 | |
| gRNA3 only | 20 | 0 | 0 | 0 | 0 | |
| gRNA4 only | 20 | 0 | 0 | 0 | 0 | |
| Cas9 + gRNA2 rep1 | 20 | 0 | 0.99 | 0 | 0 | 0 |
| Cas9 + gRNA2 rep2 | 20 | 1 | 1 | 1 | −4 to −2 | −2 |
| Cas9 + gRNA3 rep1 | 20 | 0 | 1 | 0 | −7 to −1 | −2 |
| Cas9 + gRNA3 rep2 | 20 | 1 | 1 | 1 | −7 to−1 | −2 |
| Cas9 + gRNA4 rep1 | 20 | 3 | 0.99 | 2 | −7 to +12 | −2 |
| Cas9 + gRNA4 rep2 | 20 | 1 | 0.99 | 1 | −2 to +3 | −2 |
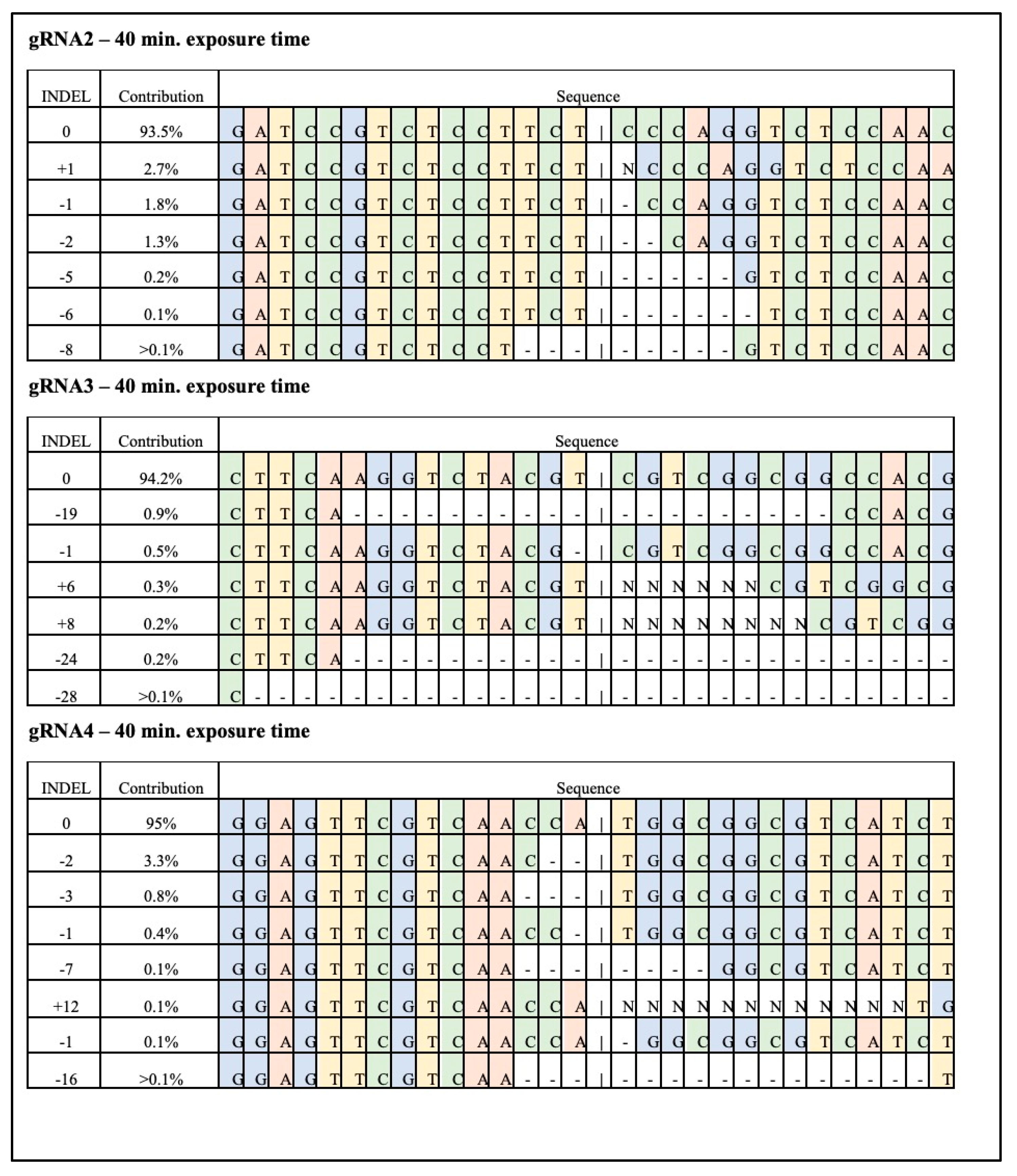
| Cas9 + gRNA2 rep1 | 40 | 4 | 0.99 | 4 | −7 to +1 | −1 |
| Cas9 + gRNA2 rep2 | 40 | 6 | 1 | 6 | -8 to +1 | +1 |
| Cas9 + gRNA3 rep1 | 40 | 1 | 1 | 1 | −19 to−1 | −1 |
| Cas9 + gRNA3 rep2 | 40 | 2 | 0.96 | 2 | −28 to +8 | −19 |
| Cas9 + gRNA4 rep1 | 40 | 2 | 0.99 | 2 | −3 to −2 | −2 |
| Cas9 + gRNA4 rep2 | 40 | 5 | 1 | 4 | −16 to +12 | −2 |
| Reference | Plant Species | Plant Material | Transfection Method | Gene-Editing Efficiency |
|---|---|---|---|---|
| RNP delivered in plants | ||||
| [12] | Arabidopsis thaliana, Lactuca sativa, Nicotiana attenuata, Oryza sativa | Protoplasts | PEG-mediated | 5.7–40.0% |
| [13] | Malus domestica, Vitis vinifera | Protoplasts | PEG-mediated | 0.1–6.9% |
| [14] | Petunia x hybrid | Protoplasts | PEG-mediated | 2.4–21.0% |
| [31] | Triticum aestivum | Protoplasts, immature embryos | PEG-mediated, Biolistics | 0.2–45.3% |
| [15] | Solanum tuberosum | Protoplasts | PEG-mediated | 1.0–25.0% |
| [32] | Brassica oleracea, Brassica rapa | Protoplasts | PEG-mediated | 0.1–24.5% |
| [33] | Oryza sativa | Zygotes | PEG-mediated | 14.0–64.0% |
| Other delivery methods in maize | ||||
| [34] | Zea mays | Protoplasts | Vector via PEG-mediated | 13.1% |
| Immature embryos | Agrobacterium-mediated | 16.4–19.1% | ||
| [35] | Zea mays | Protoplasts | Vector via PEG-mediated | N.A |
| [24] | Zea mays | Immature embryos | Vector via biolistics | 1.3–4.6% |
| [17] | Zea mays | Protoplasts | Vector via biolistics | 80.0–90.0% |
| [36] | Zea mays | Immature embryos | Agrobacterium-mediated | 57.1–71.4% |
| [19] | Zea mays | Protoplasts | Vector via PEG-mediated | 2.8–27.0% |
| Immature embryos | Agrobacterium-mediated | 19.0–31.0% | ||
| [37] | Zea mays | Protoplasts | Vector via PEG-mediated | 4.0–11.9% |
| Immature embryos | Agrobacterium-mediated | 65.8–86.9% | ||
| [16] | Zea mays | Immature embryos | RNP via Biolistics | 0.01–0.7% |
| [18] | Zea mays | Immature embryos | Agrobacterium-mediated | 12.0–74.0% |
| [38] | Zea mays | Immature embryos | Vector via Biolistics | 60.0–98.0% |
| [39] | Zea mays | Immature embryos | Agrobacterium-mediated | N.A |
| [19] | Zea mays | Immature embryos | Agrobacterium-mediated | 5.0–100% |
| [20] | Zea mays | Immature embryos | Agrobacterium-mediated | N.A |
| [40] | Zea mays | Immature embryos | Agrobacterium-mediated | 90.0–100% |
| [41] | Zea mays | Immature embryos | Agrobacterium-mediated | N.A |
| [42] | Zea mays | Immature embryos | Agrobacterium-mediated | N.A |
| [43] | Zea mays | Immature embryos | Agrobacterium-mediated | N.A |
| [44] | Zea mays | Immature embryos | Agrobacterium-mediated | N.A |
| [45] | Zea mays | Immature embryos | Agrobacterium-mediated | N.A |
| [46] | Zea mays | Immature embryos | Vector via biolistics | N.A |
| [47] | Zea mays | Immature embryos | Agrobacterium-mediated | 25.0–100% |
| [48] | Zea mays | Immature embryos | Agrobacterium-mediated | N.A |
| Present study | Zea mays | Protoplasts | RNA via PEG-mediated | 0.85–5.85% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sant’Ana, R.R.A.; Caprestano, C.A.; Nodari, R.O.; Agapito-Tenfen, S.Z. PEG-Delivered CRISPR-Cas9 Ribonucleoproteins System for Gene-Editing Screening of Maize Protoplasts. Genes 2020, 11, 1029. https://doi.org/10.3390/genes11091029
Sant’Ana RRA, Caprestano CA, Nodari RO, Agapito-Tenfen SZ. PEG-Delivered CRISPR-Cas9 Ribonucleoproteins System for Gene-Editing Screening of Maize Protoplasts. Genes. 2020; 11(9):1029. https://doi.org/10.3390/genes11091029
Chicago/Turabian StyleSant’Ana, Rodrigo Ribeiro Arnt, Clarissa Alves Caprestano, Rubens Onofre Nodari, and Sarah Zanon Agapito-Tenfen. 2020. "PEG-Delivered CRISPR-Cas9 Ribonucleoproteins System for Gene-Editing Screening of Maize Protoplasts" Genes 11, no. 9: 1029. https://doi.org/10.3390/genes11091029
APA StyleSant’Ana, R. R. A., Caprestano, C. A., Nodari, R. O., & Agapito-Tenfen, S. Z. (2020). PEG-Delivered CRISPR-Cas9 Ribonucleoproteins System for Gene-Editing Screening of Maize Protoplasts. Genes, 11(9), 1029. https://doi.org/10.3390/genes11091029