RECQ1 Helicase in Genomic Stability and Cancer



Abstract
1. Introduction
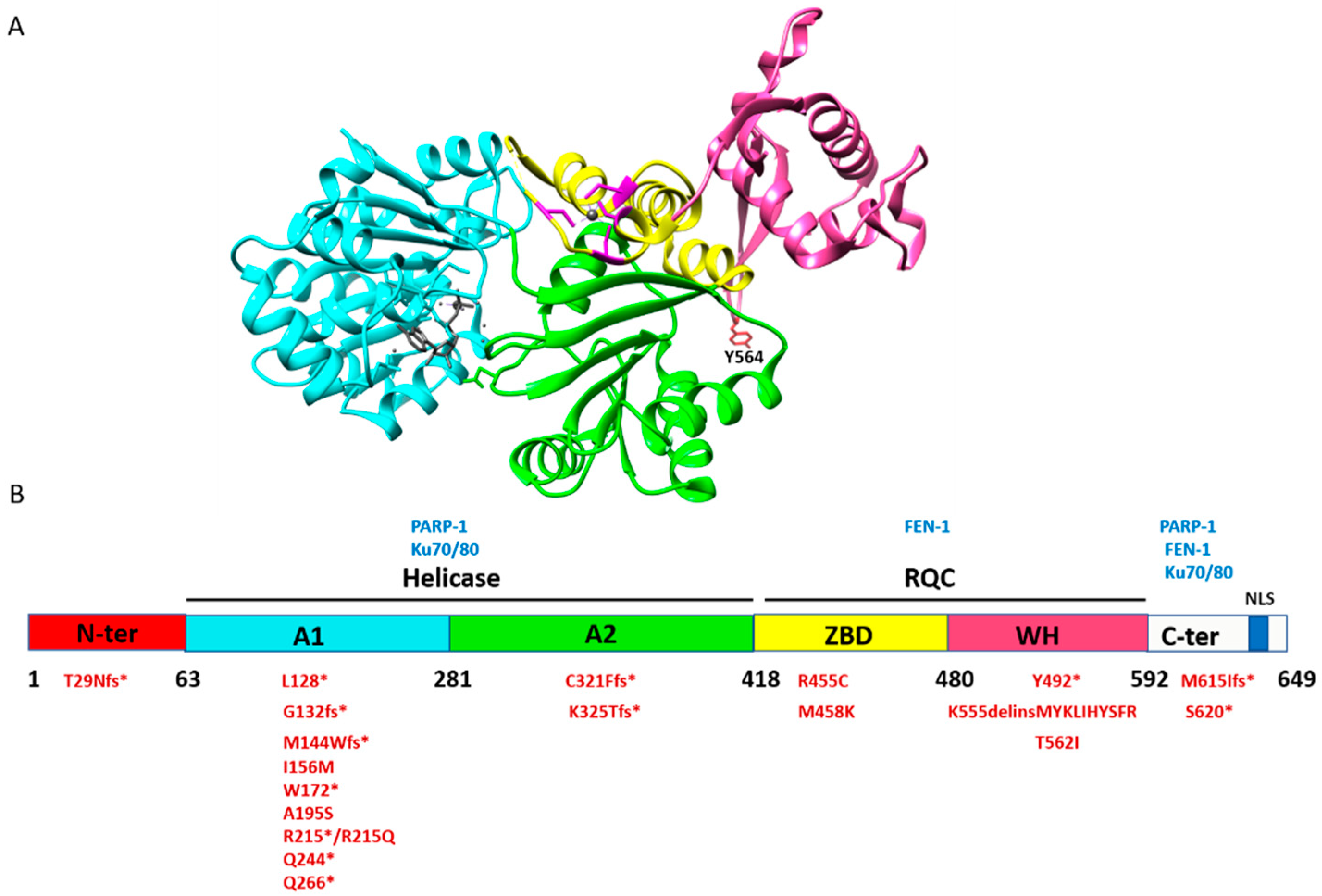
2. Structure and Biochemical Properties of RECQ1
3. Demonstrated Roles of RECQ1 in DNA Repair
3.1. RECQ1–RPA Interaction
3.2. RECQ1–PARP-1 Interaction
3.3. RECQ1–Ku70/80 Interaction
3.4. RECQ1–FEN-1 Interaction
4. Non-Canonical Roles of RECQ1
5. Association of RECQ1 Expression with Cancer
6. RECQ1 as a Candidate Breast Cancer Susceptibility Gene
7. Outlook for RECQ1 Relationship with Cancer
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tubbs, A.; Nussenzweig, A. Endogenous DNA Damage as a Source of Genomic Instability in Cancer. Cell 2017, 168, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Doherty, K.M.; Brosh, R.M. Mechanisms of RecQ helicases in pathways of DNA metabolism and maintenance of genomic stability. Biochem. J. 2006, 398, 319–337. [Google Scholar] [CrossRef] [PubMed]
- Croteau, D.L.; Popuri, V.; Opresko, P.L.; Bohr, V.A. Human RecQ helicases in DNA repair, recombination, and replication. Annu. Rev. Biochem. 2014, 83, 519–552. [Google Scholar] [CrossRef] [PubMed]
- Brosh, R.M. DNA helicases involved in DNA repair and their roles in cancer. Nat. Rev. Cancer 2013, 13, 542–558. [Google Scholar] [CrossRef]
- Cybulski, C.; Carrot-Zhang, J.; Kluzniak, W.; Rivera, B.; Kashyap, A.; Wokolorczyk, D.; Giroux, S.; Nadaf, J.; Hamel, N.; Zhang, S.; et al. Germline RECQL mutations are associated with breast cancer susceptibility. Nat. Genet. 2015, 47, 643–646. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, Y.; Xia, Y.; Xu, Y.; Ouyang, T.; Li, J.; Wang, T.; Fan, Z.; Fan, T.; Lin, B.; et al. Mutations in RECQL Gene Are Associated with Predisposition to Breast Cancer. PLoS Genet. 2015, 11, e1005228. [Google Scholar] [CrossRef]
- Seki, M.; Miyazawa, H.; Tada, S.; Yanagisawa, J.; Yamaoka, T.; Hoshino, S.; Ozawa, K.; Eki, T.; Nogami, M.; Okumura, K. Molecular cloning of cDNA encoding human DNA helicase Q1 which has homology to Escherichia coli Rec Q helicase and localization of the gene at chromosome 12p12. Nucleic Acids Res. 1994, 22, 4566–4573. [Google Scholar] [CrossRef]
- Puranam, K.L.; Blackshear, P.J. Cloning and characterization of RECQL, a potential human homologue of the Escherichia coli DNA helicase RecQ. J. Biol. Chem. 1994, 269, 29838–29845. [Google Scholar]
- Sami, F.; Sharma, S. Probing Genome Maintenance Functions of human RECQ1. Comput. Struct. Biotechnol. J. 2013, 6, e201303014. [Google Scholar] [CrossRef]
- Vindigni, A.; Marino, F.; Gileadi, O. Probing the structural basis of RecQ helicase function. Biophys. Chem. 2010, 149, 67–77. [Google Scholar] [CrossRef]
- Sharma, S.; Sommers, J.A.; Choudhary, S.; Faulkner, J.K.; Cui, S.; Andreoli, L.; Muzzolini, L.; Vindigni, A.; Brosh, R.M. Biochemical analysis of the DNA unwinding and strand annealing activities catalyzed by human RECQ1. J. Biol. Chem. 2005, 280, 28072–28084. [Google Scholar] [CrossRef]
- Cui, S.; Klima, R.; Ochem, A.; Arosio, D.; Falaschi, A.; Vindigni, A. Characterization of the DNA-unwinding activity of human RECQ1, a helicase specifically stimulated by human replication protein A. J. Biol. Chem. 2003, 278, 1424–1432. [Google Scholar] [CrossRef]
- Muzzolini, L.; Beuron, F.; Patwardhan, A.; Popuri, V.; Cui, S.; Niccolini, B.; Rappas, M.; Freemont, P.S.; Vindigni, A. Different quaternary structures of human RECQ1 are associated with its dual enzymatic activity. PLoS Biol. 2007, 5, e20. [Google Scholar] [CrossRef]
- Pike, A.C.; Shrestha, B.; Popuri, V.; Burgess-Brown, N.; Muzzolini, L.; Costantini, S.; Vindigni, A.; Gileadi, O. Structure of the human RECQ1 helicase reveals a putative strand-separation pin. Proc. Natl. Acad. Sci. USA 2009, 106, 1039–1044. [Google Scholar] [CrossRef]
- Pike, A.C.; Gomathinayagam, S.; Swuec, P.; Berti, M.; Zhang, Y.; Schnecke, C.; Marino, F.; von Delft, F.; Renault, L.; Costa, A.; et al. Human RECQ1 helicase-driven DNA unwinding, annealing, and branch migration: Insights from DNA complex structures. Proc. Natl. Acad. Sci. USA 2015, 112, 4286–4291. [Google Scholar] [CrossRef]
- Sami, F.; Gary, R.K.; Fang, Y.; Sharma, S. Site-directed mutants of human RECQ1 reveal functional importance of the zinc binding domain. Mutat. Res. 2016, 790, 8–18. [Google Scholar] [CrossRef]
- Sharma, S.; Stumpo, D.J.; Balajee, A.S.; Bock, C.B.; Lansdorp, P.M.; Brosh, R.M.; Blackshear, P.J. RECQL, a member of the RecQ family of DNA helicases, suppresses chromosomal instability. Mol. Cell. Biol. 2007, 27, 1784–1794. [Google Scholar] [CrossRef]
- Sharma, S.; Brosh, R.M. Human RECQ1 is a DNA damage responsive protein required for genotoxic stress resistance and suppression of sister chromatid exchanges. PLoS ONE 2007, 2, e1297. [Google Scholar] [CrossRef]
- Berti, M.; Ray Chaudhuri, A.; Thangavel, S.; Gomathinayagam, S.; Kenig, S.; Vujanovic, M.; Odreman, F.; Glatter, T.; Graziano, S.; Mendoza-Maldonado, R.; et al. Human RECQ1 promotes restart of replication forks reversed by DNA topoisomerase I inhibition. Nat. Struct. Mol. Biol. 2013, 20, 347–354. [Google Scholar] [CrossRef]
- Popuri, V.; Croteau, D.L.; Brosh, R.M.; Bohr, V.A. RECQ1 is required for cellular resistance to replication stress and catalyzes strand exchange on stalled replication fork structures. Cell Cycle 2012, 11, 4252–4265. [Google Scholar] [CrossRef]
- Lu, X.; Parvathaneni, S.; Hara, T.; Lal, A.; Sharma, S. Replication stress induces specific enrichment of RECQ1 at common fragile sites FRA3B and FRA16D. Mol. Cancer 2013, 12, 29. [Google Scholar] [CrossRef]
- Sharma, S.; Phatak, P.; Stortchevoi, A.; Jasin, M.; Larocque, J.R. RECQ1 plays a distinct role in cellular response to oxidative DNA damage. DNA Repair 2012, 11, 537–549. [Google Scholar] [CrossRef]
- Seki, T.; Tada, S.; Katada, T.; Enomoto, T. Cloning of a cDNA encoding a novel importin-α homologue, Qip1: Discrimination of Qip1 and Rch1 from hSrp1 by their ability to interact with DNA helicase Q1/RecQL. Biochem. Biophys. Res. Commun. 1997, 234, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Montes de Oca, R.; Shoemaker, C.J.; Gucek, M.; Cole, R.N.; Wilson, K.L. Barrier-to-autointegration factor proteome reveals chromatin-regulatory partners. PLoS ONE 2009, 4, e7050. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Levicoy, J.; Moreira-Ramos, S.; Rojas, D.A.; Urbina, F.; Maldonado, E. Transcription directed by human core promoters with a HomolD box sequence requires DDB1, RECQL and RNA polymerase II machinery. Gene 2012, 505, 318–323. [Google Scholar] [CrossRef]
- Doherty, K.M.; Sharma, S.; Uzdilla, L.A.; Wilson, T.M.; Cui, S.; Vindigni, A.; Brosh, R.M. RECQ1 helicase interacts with human mismatch repair factors that regulate genetic recombination. J. Biol. Chem. 2005, 280, 28085–28094. [Google Scholar] [CrossRef]
- Sami, F.; Lu, X.; Parvathaneni, S.; Roy, R.; Gary, R.K.; Sharma, S. RECQ1 interacts with FEN-1 and promotes binding of FEN-1 to telomeric chromatin. Biochem. J. 2015, 468, 227–244. [Google Scholar] [CrossRef]
- Parvathaneni, S.; Stortchevoi, A.; Sommers, J.A.; Brosh, R.M.; Sharma, S. Human RECQ1 interacts with Ku70/80 and modulates DNA end-joining of double-strand breaks. PLoS ONE 2013, 8, e62481. [Google Scholar] [CrossRef]
- Woodrick, J.; Gupta, S.; Camacho, S.; Parvathaneni, S.; Choudhury, S.; Cheema, A.; Bai, Y.; Khatkar, P.; Erkizan, H.V.; Sami, F.; et al. A new sub-pathway of long-patch base excision repair involving 5′ gap formation. EMBO J. 2017, 36, 1605–1622. [Google Scholar] [CrossRef]
- Cui, S.; Arosio, D.; Doherty, K.M.; Brosh, R.M.; Falaschi, A.; Vindigni, A. Analysis of the unwinding activity of the dimeric RECQ1 helicase in the presence of human replication protein A. Nucleic Acids Res. 2004, 32, 2158–2170. [Google Scholar] [CrossRef]
- Johnson, F.B.; Lombard, D.B.; Neff, N.F.; Mastrangelo, M.A.; Dewolf, W.; Ellis, N.A.; Marciniak, R.A.; Yin, Y.; Jaenisch, R.; Guarente, L. Association of the Bloom syndrome protein with topoisomerase IIIalpha in somatic and meiotic cells. Cancer Res. 2000, 60, 1162–1167. [Google Scholar]
- Chen, R.; Wold, M.S. Replication protein A: Single-stranded DNA’s first responder: Dynamic DNA-interactions allow replication protein A to direct single-strand DNA intermediates into different pathways for synthesis or repair. Bioessays 2014, 36, 1156–1161. [Google Scholar] [CrossRef]
- Banerjee, T.; Sommers, J.A.; Huang, J.; Seidman, M.M.; Brosh, R.M. Catalytic strand separation by RECQ1 is required for RPA-mediated response to replication stress. Curr. Biol. 2015, 25, 2830–2838. [Google Scholar] [CrossRef]
- Parvathaneni, S.; Sharma, S. The DNA repair helicase RECQ1 has a checkpoint-dependent role in mediating DNA damage responses induced by gemcitabine. J. Biol. Chem. 2019, 294, 15330–15345. [Google Scholar] [CrossRef]
- Liu, Y.; Kadyrov, F.A.; Modrich, P. PARP-1 enhances the mismatch-dependence of 5′-directed excision in human mismatch repair in vitro. DNA Repair 2011, 10, 1145–1153. [Google Scholar] [CrossRef]
- Lieber, M.R. The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway. Annu. Rev. Biochem. 2010, 79, 181–211. [Google Scholar] [CrossRef]
- Balakrishnan, L.; Bambara, R.A. Flap endonuclease 1. Annu. Rev. Biochem. 2013, 82, 119–138. [Google Scholar] [CrossRef]
- Saharia, A.; Guittat, L.; Crocker, S.; Lim, A.; Steffen, M.; Kulkarni, S.; Stewart, S.A. Flap endonuclease 1 contributes to telomere stability. Curr. Biol. 2008, 18, 496–500. [Google Scholar] [CrossRef]
- Sharma, S.; Sommers, J.A.; Brosh, R.M. Processing of DNA replication and repair intermediates by the concerted action of RecQ helicases and Rad2 structure-specific nucleases. Protein Pept. Lett. 2008, 15, 89–102. [Google Scholar] [CrossRef]
- Popuri, V.; Hsu, J.; Khadka, P.; Horvath, K.; Liu, Y.; Croteau, D.L.; Bohr, V.A. Human RECQL1 participates in telomere maintenance. Nucleic Acids Res. 2014, 42, 5671–5688. [Google Scholar] [CrossRef]
- Li, X.L.; Lu, X.; Parvathaneni, S.; Bilke, S.; Zhang, H.; Thangavel, S.; Vindigni, A.; Hara, T.; Zhu, Y.; Meltzer, P.S.; et al. Identification of RECQ1-regulated transcriptome uncovers a role of RECQ1 in regulation of cancer cell migration and invasion. Cell Cycle 2014, 13, 2431–2445. [Google Scholar] [CrossRef]
- Lu, X.; Parvathaneni, S.; Li, X.L.; Lal, A.; Sharma, S. Transcriptome guided identification of novel functions of RECQ1 helicase. Methods 2016, 108, 111–117. [Google Scholar] [CrossRef]
- Johnson, J.E.; Cao, K.; Ryvkin, P.; Wang, L.S.; Johnson, F.B. Altered gene expression in the Werner and Bloom syndromes is associated with sequences having G-quadruplex forming potential. Nucleic Acids Res. 2010, 38, 1114–1122. [Google Scholar] [CrossRef]
- Turaga, R.V.; Paquet, E.R.; Sild, M.; Vignard, J.; Garand, C.; Johnson, F.B.; Masson, J.Y.; Lebel, M. The Werner syndrome protein affects the expression of genes involved in adipogenesis and inflammation in addition to cell cycle and DNA damage responses. Cell Cycle 2009, 8, 2080–2092. [Google Scholar] [CrossRef]
- Kyng, K.J.; Bohr, V.A. Gene expression and DNA repair in progeroid syndromes and human aging. Ageing Res. Rev. 2005, 4, 579–602. [Google Scholar] [CrossRef]
- Aygun, O.; Xu, X.; Liu, Y.; Takahashi, H.; Kong, S.E.; Conaway, R.C.; Conaway, J.W.; Svejstrup, J.Q. Direct inhibition of RNA polymerase II transcription by RECQL5. J. Biol. Chem. 2009, 284, 23197–23203. [Google Scholar] [CrossRef]
- Popuri, V.; Bachrati, C.Z.; Muzzolini, L.; Mosedale, G.; Costantini, S.; Giacomini, E.; Hickson, I.D.; Vindigni, A. The Human RecQ helicases, BLM and RECQ1, display distinct DNA substrate specificities. J. Biol. Chem. 2008, 283, 17766–17776. [Google Scholar] [CrossRef]
- Fleming, A.M.; Zhu, J.; Ding, Y.; Burrows, C.J. 8-Oxo-7,8-dihydroguanine in the Context of a Gene Promoter G-Quadruplex Is an On-Off Switch for Transcription. ACS Chem. Biol. 2017, 12, 2417–2426. [Google Scholar] [CrossRef]
- Fleming, A.M.; Ding, Y.; Burrows, C.J. Oxidative DNA damage is epigenetic by regulating gene transcription via base excision repair. Proc. Natl. Acad. Sci. USA 2017, 114, 2604–2609. [Google Scholar] [CrossRef]
- Bokhari, B.; Sharma, S. Stress Marks on the Genome: Use or Lose? Int. J. Mol. Sci. 2019, 20, 364. [Google Scholar] [CrossRef]
- Li, X.; Wang, W.; Wang, J.; Malovannaya, A.; Xi, Y.; Li, W.; Guerra, R.; Hawke, D.H.; Qin, J.; Chen, J. Proteomic analyses reveal distinct chromatin-associated and soluble transcription factor complexes. Mol. Syst. Biol. 2015, 11, 775. [Google Scholar] [CrossRef]
- Sharma, S. An appraisal of RECQ1 expression in cancer progression. Front. Genet. 2014, 5, 426. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Maldonado, R.; Faoro, V.; Bajpai, S.; Berti, M.; Odreman, F.; Vindigni, M.; Ius, T.; Ghasemian, A.; Bonin, S.; Skrap, M.; et al. The human RECQ1 helicase is highly expressed in glioblastoma and plays an important role in tumor cell proliferation. Mol. Cancer 2011, 10, 83. [Google Scholar] [CrossRef] [PubMed]
- Viziteu, E.; Klein, B.; Basbous, J.; Lin, Y.L.; Hirtz, C.; Gourzones, C.; Tiers, L.; Bruyer, A.; Vincent, L.; Grandmougin, C.; et al. RECQ1 helicase is involved in replication stress survival and drug resistance in multiple myeloma. Leukemia 2017, 31, 2104–2113. [Google Scholar] [CrossRef] [PubMed]
- Sanada, S.; Futami, K.; Terada, A.; Yonemoto, K.; Ogasawara, S.; Akiba, J.; Yasumoto, M.; Sumi, A.; Ushijima, K.; Kamura, T.; et al. RECQL1 DNA repair helicase: A potential therapeutic target and a proliferative marker against ovarian cancer. PLoS ONE 2013, 8, e72820. [Google Scholar] [CrossRef]
- Viziteu, E.; Kassambara, A.; Pasero, P.; Klein, B.; Moreaux, J. RECQ helicases are deregulated in hematological malignancies in association with a prognostic value. Biomark. Res. 2016, 4, 3–4. [Google Scholar] [CrossRef]
- Futami, K.; Ogasawara, S.; Goto, H.; Yano, H.; Furuichi, Y. RecQL1 DNA repair helicase: A potential tumor marker and therapeutic target against hepatocellular carcinoma. Int. J. Mol. Med. 2010, 25, 537–545. [Google Scholar] [CrossRef]
- Arai, A.; Chano, T.; Futami, K.; Furuichi, Y.; Ikebuchi, K.; Inui, T.; Tameno, H.; Ochi, Y.; Shimada, T.; Hisa, Y.; et al. RECQL1 and WRN proteins are potential therapeutic targets in head and neck squamous cell carcinoma. Cancer Res. 2011, 71, 4598–4607. [Google Scholar] [CrossRef]
- Tao, J.; Tao, S.; Han, J.; Zhou, Z.; Zhang, X.; Wang, H.; Chen, R.; Ji, F.; Zhu, Y. RECQL1 plays an important role in the development of tongue squamous cell carcinoma. Cell. Physiol. Biochem. 2014, 33, 1579–1590. [Google Scholar] [CrossRef]
- Futami, K.; Kumagai, E.; Makino, H.; Goto, H.; Takagi, M.; Shimamoto, A.; Furuichi, Y. Induction of mitotic cell death in cancer cells by small interference RNA suppressing the expression of RecQL1 helicase. Cancer Sci. 2008, 99, 71–80. [Google Scholar] [CrossRef]
- Futami, K.; Kumagai, E.; Makino, H.; Sato, A.; Takagi, M.; Shimamoto, A.; Furuichi, Y. Anticancer activity of RecQL1 helicase siRNA in mouse xenograft models. Cancer Sci. 2008, 99, 1227–1236. [Google Scholar] [CrossRef]
- Vittori, M.; Breznik, B.; Hrovat, K.; Kenig, S.; Lah, T.T. RECQ1 Helicase Silencing Decreases the Tumour Growth Rate of U87 Glioblastoma Cell Xenografts in Zebrafish Embryos. Genes 2017, 8, 222. [Google Scholar] [CrossRef]
- Arora, A.; Parvathaneni, S.; Aleskandarany, M.A.; Agarwal, D.; Ali, R.; Abdel-Fatah, T.; Green, A.R.; Ball, G.R.; Rakha, E.A.; Ellis, I.O.; et al. Clinicopathological and Functional Significance of RECQL1 Helicase in Sporadic Breast Cancers. Mol. Cancer Ther. 2017, 16, 239–250. [Google Scholar] [CrossRef]
- Parvathaneni, S.; Lu, X.; Chaudhary, R.; Lal, A.; Madhusudan, S.; Sharma, S. RECQ1 expression is upregulated in response to DNA damage and in a p53-dependent manner. Oncotarget 2017, 8, 75924–75942. [Google Scholar] [CrossRef]
- Kwong, A.; Shin, V.Y.; Cheuk, I.W.Y.; Chen, J.; Au, C.H.; Ho, D.N.; Chan, T.L.; Ma, E.S.K.; Akbari, M.R.; Narod, S.A. Germline RECQL mutations in high risk Chinese breast cancer patients. Breast Cancer Res. Treat. 2016, 157, 211–215. [Google Scholar] [CrossRef]
- Sun, J.; Meng, H.; Yao, L.; Lv, M.; Bai, J.; Zhang, J.; Wang, L.; Ouyang, T.; Li, J.; Wang, T.; et al. Germline Mutations in Cancer Susceptibility Genes in a Large Series of Unselected Breast Cancer Patients. Clin. Cancer Res. 2017, 23, 6113–6119. [Google Scholar] [CrossRef]
- Tervasmaki, A.; Mantere, T.; Hartikainen, J.M.; Kauppila, S.; Lee, H.M.; Koivuluoma, S.; Grip, M.; Karihtala, P.; Jukkola-Vuorinen, A.; Mannermaa, A.; et al. Rare missense mutations in RECQL and POLG associate with inherited predisposition to breast cancer. Int. J. Cancer 2018, 142, 2286–2292. [Google Scholar] [CrossRef]
- Bogdanova, N.; Pfeifer, K.; Schurmann, P.; Antonenkova, N.; Siggelkow, W.; Christiansen, H.; Hillemanns, P.; Park-Simon, T.W.; Dork, T. Analysis of a RECQL splicing mutation, c.1667_1667+3delAGTA, in breast cancer patients and controls from Central Europe. Fam. Cancer 2017, 16, 181–186. [Google Scholar] [CrossRef]
- Li, N.; Rowley, S.M.; Goode, D.L.; Amarasinghe, K.C.; McInerny, S.; Devereux, L.; LifePool Investigators; Wong-Brown, M.W.; Lupat, R.; Lee, J.E.A.; et al. Mutations in RECQL are not associated with breast cancer risk in an Australian population. Nat. Genet. 2018, 50, 1346–1348. [Google Scholar] [CrossRef]
- Lek, M.; Karczewski, K.J.; Minikel, E.V.; Samocha, K.E.; Banks, E.; Fennell, T.; O’Donnell-Luria, A.H.; Ware, J.S.; Hill, A.J.; Cummings, B.B.; et al. Exome Aggregation Consortium Analysis of protein-coding genetic variation in 60,706 humans. Nature 2016, 536, 285–291. [Google Scholar] [CrossRef]
- Nguyen-Dumont, T.; Myszka, A.; Karpinski, P.; Sasiadek, M.M.; Akopyan, H.; Hammet, F.; Tsimiklis, H.; Park, D.J.; Pope, B.J.; Slezak, R.; et al. FANCM and RECQL genetic variants and breast cancer susceptibility: Relevance to South Poland and West Ukraine. BMC Med. Genet. 2018, 19, 12. [Google Scholar] [CrossRef]
- Hilz, P.; Heinrihsone, R.; Patzold, L.A.; Qi, Q.; Trofimovics, G.; Gailite, L.; Irmejs, A.; Gardovskis, J.; Miklasevics, E.; Daneberga, Z. Allelic variants of breast cancer susceptibility genes PALB2 and RECQL in the Latvian population. Hered. Cancer Clin. Pract. 2019, 17, 17. [Google Scholar] [CrossRef]
- Wendt, C.; Margolin, S. Identifying breast cancer susceptibility genes—A review of the genetic background in familial breast cancer. Acta Oncol. 2019, 58, 135–146. [Google Scholar] [CrossRef]
- Zheng, G.; Yu, H.; Hemminki, A.; Forsti, A.; Sundquist, K.; Hemminki, K. Familial associations of female breast cancer with other cancers. Int. J. Cancer 2017, 141, 2253–2259. [Google Scholar] [CrossRef]
- Salk, J.J.; Fox, E.J.; Loeb, L.A. Mutational heterogeneity in human cancers: Origin and consequences. Annu. Rev. Pathol. 2010, 5, 51–75. [Google Scholar] [CrossRef]


{kind=link}
| Protein | Functional Interaction | Reference |
|---|---|---|
| BAF | Unknown | [24] |
| DDB1 | Unknown | [25] |
| EXO-1 | RECQ1 stimulates EXO-1 nuclease activity | [26] |
| FEN-1 | RECQ1 stimulates FEN-1 nuclease activity | [27] |
| Importin-α (Qip1 and Rch1) | Unknown | [23] |
| Ku70/80 | RECQ1 regulates DNA binding of Ku70/80 | [28] |
| MLH1-PMS2 | Unknown | [26] |
| MSH2/6 | MSH2/6 stimulates RECQ1 helicase activity | [26] |
| PARP-1 | PARP-1 regulates ATPase and branch migration activities of RECQ1 | [19] |
| RECQ1 regulates poly-ADP-ribosylation activity of PARP-1 | [29] | |
| RAD51 | Unknown | [18] |
| RPA | RPA stimulates RECQ1 helicase activity | [30] |
| Top3α | Unknown | [31] |
| Study | Mutation | Change in Protein | No. of Carriers/Total No. of Cases | No. of Carriers/Total No. of Controls |
|---|---|---|---|---|
| Cybulski et al. (2015) [5] | c.643C>T c.1667_1667+3delAGTA | p.R215 * p.K555delinsMYKLIHYSFR | 7/1013 30/13136 | 1/7136 2/4702 |
| Sun et al. (2015) [6] | c.383T>G | p.L128 * | 1/448 | 0/1588 |
| c.516G>A | p.W172 * | 1/448 | 0/1588 | |
| c.796C>T | p.Q266 * | 1/448 | 0/1588 | |
| c.395-2A>G | p.G132fs * | 1/448 | 0/1588 | |
| c.644G>A | p.R215Q | 1/448 | 0/1588 | |
| c.1363C>T | p.R455C | 1/448 | 1/1588 | |
| c.1373T>A | p.M458K | 1/448 | 0/1588 | |
| c.1685C>T | p.T562I | 1/448 | 0/1588 | |
| c.583G>T | p.A195S | 1/448 | 0/1588 | |
| Kong et al. (2016) [65] | c.974_977delAAGA | p.K325Tfs * | 1/1110 | 0/88 |
| c.394+1G>A | - | 1/1110 | 0/88 | |
| c.867+1G>T | - | 2/1110 | 0/88 | |
| c.796C>T | p.Q266 * | 2/1110 | 0/88 | |
| Sun et al. (2017) [66] | c.1856dupA | p.N619fs | 1/8085 | 0/0 |
| c.C1685T | p.T562I | 1/8085 | 0/0 | |
| c.G1398A | p.W466 * | 2/8085 | 0/0 | |
| c.T1373A | p.M458K | 1/8085 | 0/0 | |
| c.C1363T | p.R455C | 1/8085 | 0/0 | |
| c.C796T | p.Q266 * | 3/8085 | 0/0 | |
| c.G644A | p.R215Q | 10/8085 | 0/0 | |
| c.G583T | p.A195S | 1/8085 | 0/0 | |
| c.A577T | p.K193 * | 1/8085 | 0/0 | |
| c.G516A | p.W172 * | 2/8085 | 0/0 | |
| c.490_491insAAATGCTT | p.S164_S165delins * | 1/8085 | 0/0 | |
| c.395-2A>G | - | 2/8085 | 0/0 | |
| c.T383G | p.L128 * | 1/8085 | 0/0 | |
| c.189_190insATGATTCT | p.S64fs | 1/8085 | 0/0 | |
| c.120dupA | p.V41fs | 2/8085 | 0/0 | |
| Tervasmaki et al. (2018) [67] | c.468T>G | p.I156M | 6/1946 | 0/1408 |
| Bogdanova et al. (2017) [68] | c.1667_1667+3delAGTA | p.K555delinsMYKLIHYSFR | 9/2596 | 6/2132 |
| Li et al. (2018) [69] | c.85dupA | p.T29Nfs * | 0/4536 | 1/4576 |
| c.426delT | p.M144Wfs * | 0/4536 | 2/4576 | |
| c.501+1G>A | - | 1/4536 | 0/4576 | |
| c.701-2A>C | - | 1/4536 | 0/4576 | |
| c.730C>T | p.Q244 * | 0/4536 | 1/4576 | |
| c.796C>T | p.Q266 * | 0/4536 | 1/4576 | |
| c.962_965delGTTT | p.C321Ffs * | 1/4536 | 0/4576 | |
| c.1098+1G>A | - | 0/4536 | 1/4576 | |
| c.1355+1G>A | - | 1/4536 | 1/4576 | |
| c.1476C>G | pY492 * | 1/4536 | 0/4576 | |
| c.1667_1667+3delAGTA | p.K555delinsMYKLIHYSFR | 2/4536 | 1/4576 | |
| c.1841_1844dupAGAT | p.M615Ifs * | 0/4536 | 1/4576 | |
| c.1859C>G | p.S620 * | 6/4536 | 16/4576 | |
| Nguyen-Dumont et al. (2018) [71] | c.1667_1667+3delAGTA | p.K555delinsMYKLIHYSFR | 0/427 | 0/0 |
| Hilz et al. (2019) [72] | c.1667_1667+3delAGTA | p.K555delinsMYKLIHYSFR | 0/715 | 2/916 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Debnath, S.; Sharma, S. RECQ1 Helicase in Genomic Stability and Cancer. Genes 2020, 11, 622. https://doi.org/10.3390/genes11060622
Debnath S, Sharma S. RECQ1 Helicase in Genomic Stability and Cancer. Genes. 2020; 11(6):622. https://doi.org/10.3390/genes11060622
Chicago/Turabian StyleDebnath, Subrata, and Sudha Sharma. 2020. "RECQ1 Helicase in Genomic Stability and Cancer" Genes 11, no. 6: 622. https://doi.org/10.3390/genes11060622
APA StyleDebnath, S., & Sharma, S. (2020). RECQ1 Helicase in Genomic Stability and Cancer. Genes, 11(6), 622. https://doi.org/10.3390/genes11060622