Genotyping by Sequencing Revealed QTL Hotspots for Trichome-Based Plant Defense in Gossypium hirsutum


 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Data Recording and Statistical Analysis
2.3. DNA Extraction and Genotyping by Sequencing
2.4. Linkage Map Construction
2.5. QTL Mapping and Gene Mining
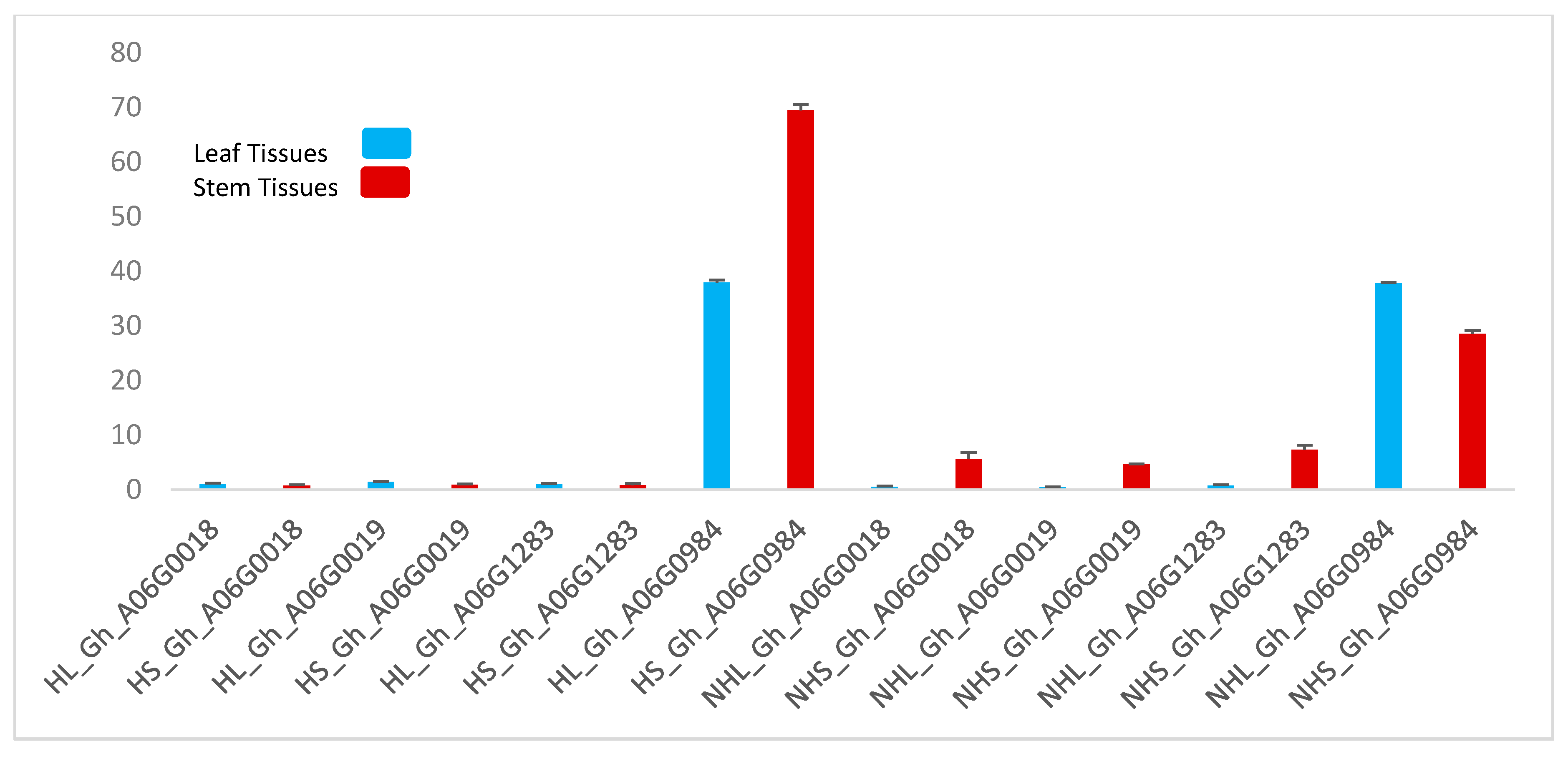
2.6. Gene Expression Level Analysis (qRT-PCR)
2.7. Synteny Analysis
3. Results
3.1. Correlation and Phenotypic Evaluation
3.2. Genetic Linkage Map for F2 Population
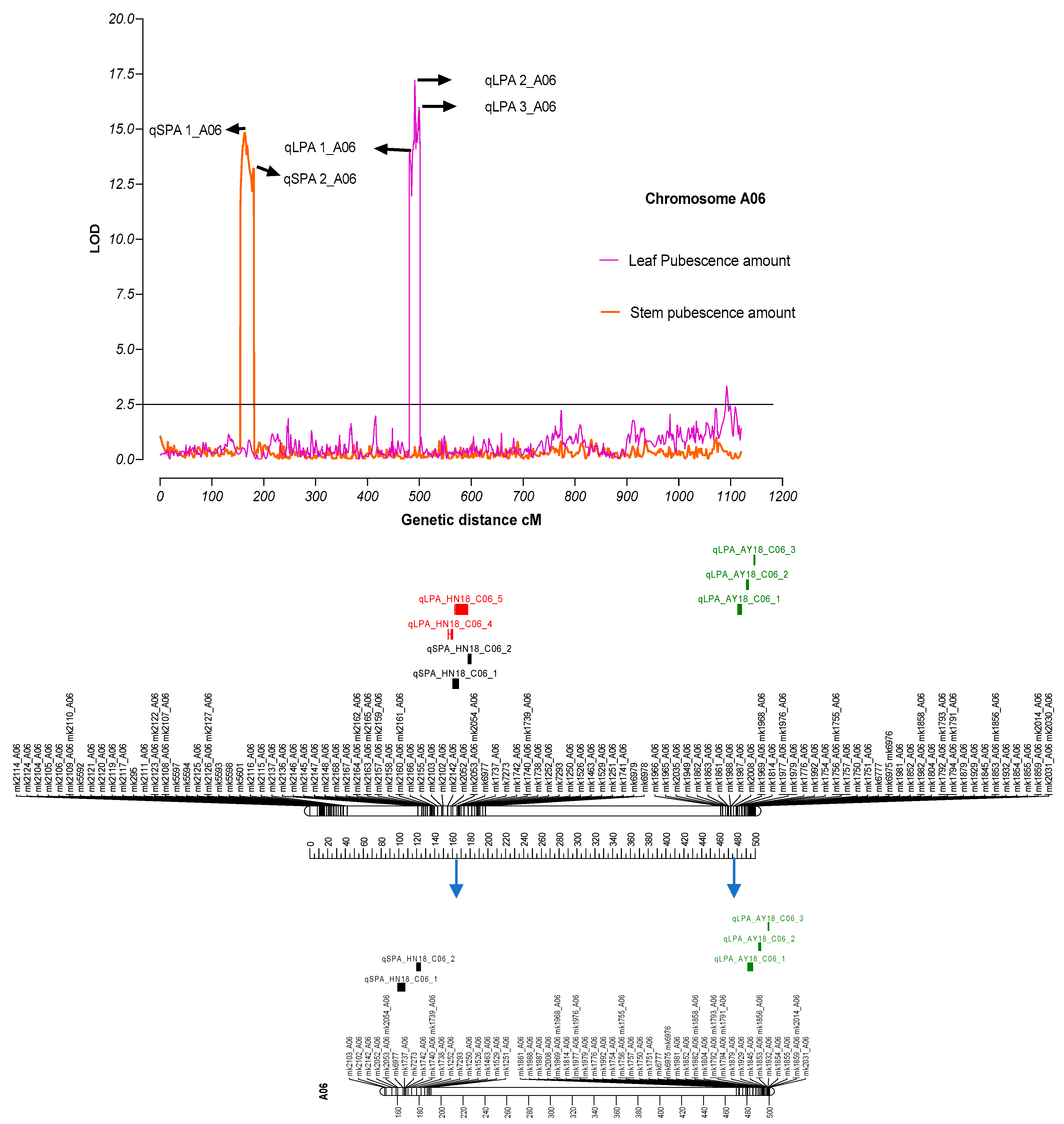
3.3. QTL Mapping
3.4. Candidate Gene Identification
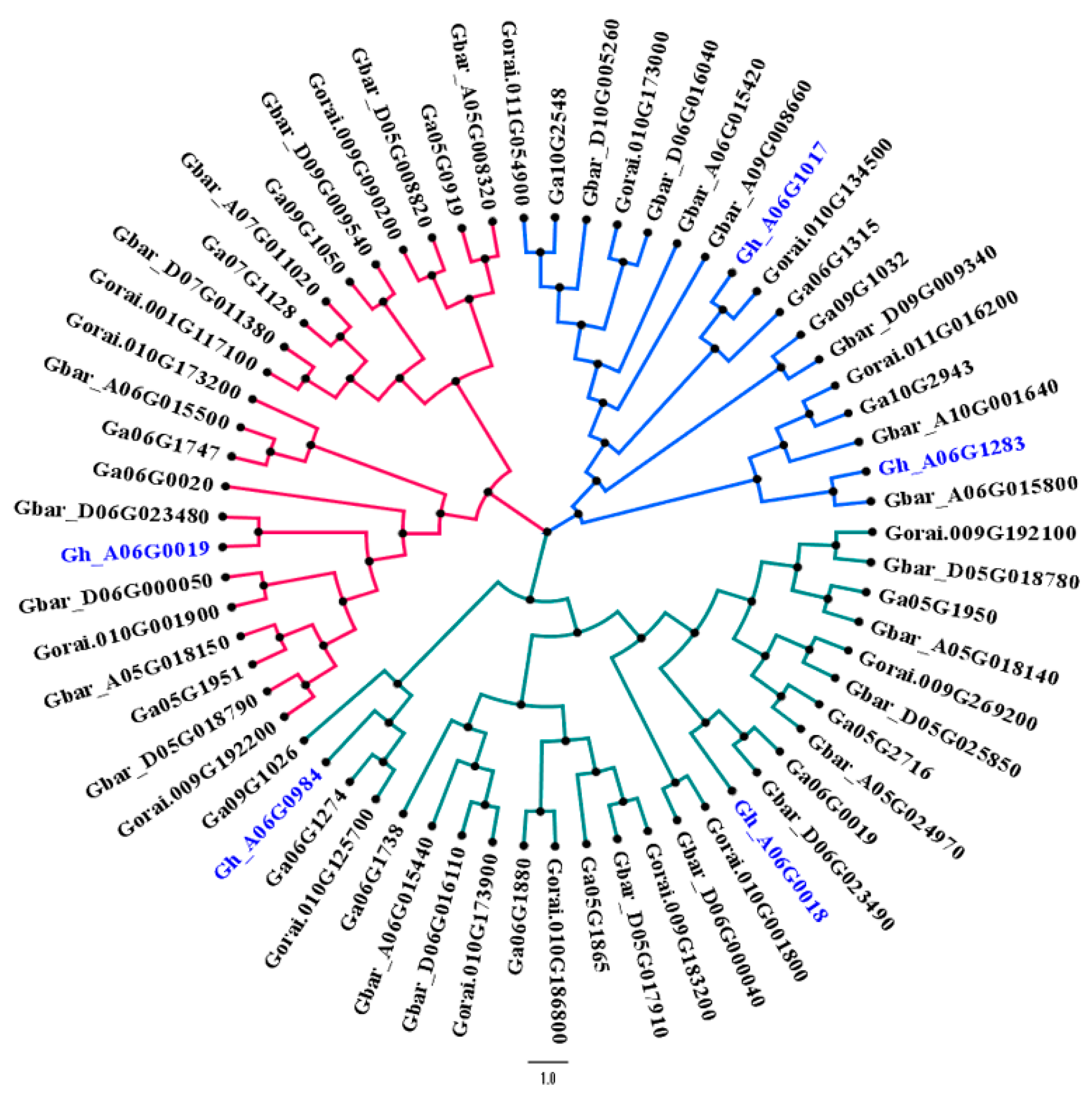
3.5. Phylogenetic Relationship of Hairy Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ding, M.; Ye, W.; Lin, L.; He, S.; Du, X.; Chen, A.; Cao, Y.; Qin, Y.; Yang, F.; Jiang, Y. The hairless stem phenotype of cotton (Gossypium barbadense) is linked to a copia-like retrotransposon insertion in a homeodomain-leucine zipper gene (HD1). Genetics 2015, 201, 143–154. [Google Scholar]
- Meagher, R.L., Jr.; Smith, W.C.; Smith, W.J. Preference of Gossypium genotypes to bemisia argentifolii (Homoptera: Aleyrodidae). J. Econ. Entomol. 1997, 90, 1046–1052. [Google Scholar] [CrossRef]
- Shukla, A.K.; Upadhyay, S.K.; Mishra, M.; Saurabh, S.; Singh, R.; Singh, H.; Thakur, N.; Rai, P.; Pandey, P.; Hans, A.L.; et al. Expression of an insecticidal fern protein in cotton protects against whitefly. Nat. Biotechnol. 2016, 34, 1046–1051. [Google Scholar] [CrossRef]
- Levin, D.A. The role of trichomes in plant defense. Q. Rev. Biol. 1973, 48, 3–15. [Google Scholar] [CrossRef]
- Hulskamp, M.; Schnittger, A.; Folkers, U. Pattern formation and cell differentiation: Trichomes in Arabidopsis as a genetic model system. In International Review of Cytology; Elsevier: Amsterdam, The Netherlands, 1998; Volume 186, pp. 147–178. [Google Scholar]
- Hülskamp, M. Plant trichomes: A model for cell differentiation. Nat. Rev. Mol. Cell Biol. 2004, 5, 471–480. [Google Scholar] [CrossRef]
- Schellmann, S.; Hulskamp, M. Epidermal differentiation: Trichomes in Arabidopsis as a model system. Int. J. Dev. Biol. 2004, 49, 579–584. [Google Scholar] [CrossRef]
- Mauricio, R.; Rausher, M.D. Experimental manipulation of putative selective agents provides evidence for the role of natural enemies in the evolution of plant defense. Evolution 1997, 51, 1435–1444. [Google Scholar] [CrossRef] [PubMed]
- Johnson, H.B. Plant pubescence: An ecological perspective. Bot. Rev. 1975, 41, 233–258. [Google Scholar] [CrossRef]
- Werker, E. Trichome diversity and development. Adv. Bot. Res. 2000, 31, 1–35. [Google Scholar]
- Nawab, N.; Khan, I.; Khan, A.; Amjad, M. Characterization and inheritance of cotton leaf pubescence. Pak. J. Bot. 2011, 43, 649–658. [Google Scholar]
- Telfer, A.; Bollman, K.M.; Poethig, R.S. Phase change and the regulation of trichome distribution in Arabidopsis thaliana. Development 1997, 124, 645–654. [Google Scholar] [PubMed]
- Pollard, A.; Briggs, D. Genecological studies of Urtica dioica L. III. Stinging hairs and plant-herbivore interactions. New Phytol. 1984, 97, 507–522. [Google Scholar] [CrossRef]
- Wan, Q.; Zhang, H.; Ye, W.; Wu, H.; Zhang, T. Genome-wide transcriptome profiling revealed cotton fuzz fiber development having a similar molecular model as Arabidopsis trichome. PLoS ONE 2014, 9, e97313. [Google Scholar] [CrossRef] [PubMed]
- Hülskamp, M.; Miséra, S.; Jürgens, G. Genetic dissection of trichome cell development in Arabidopsis. Cell 1994, 76, 555–566. [Google Scholar] [CrossRef]
- Kim, H.J.; Triplett, B.A. Cotton fiber growth in planta and in vitro. Models for plant cell elongation and cell wall biogenesis. Plant Physiol. 2001, 127, 1361–1366. [Google Scholar] [CrossRef]
- Bausher, M.G.; Singh, N.D.; Lee, S.-B.; Jansen, R.K.; Daniell, H. The complete chloroplast genome sequence of Citrus sinensis (L.) Osbeck var’Ridge Pineapple’: Organization and phylogenetic relationships to other angiosperms. BMC Plant Biol. 2006, 6, 21. [Google Scholar]
- Lee, S.-B.; Kaittanis, C.; Jansen, R.K.; Hostetler, J.B.; Tallon, L.J.; Town, C.D.; Daniell, H. The complete chloroplast genome sequence of Gossypium hirsutum: Organization and phylogenetic relationships to other angiosperms. BMC Genom. 2006, 7, 61. [Google Scholar]
- Soltis, P.S.; Soltis, D.E.; Chase, M.W. Angiosperm phylogeny inferred from multiple genes as a tool for comparative biology. Nature 1999, 402, 402–404. [Google Scholar] [CrossRef]
- Serna, L.; Martin, C. Trichomes: Different regulatory networks lead to convergent structures. Trends Plant Sci. 2006, 11, 274–280. [Google Scholar]
- Marks, M.D.; Wenger, J.P.; Gilding, E.; Jilk, R.; Dixon, R.A. Transcriptome analysis of Arabidopsis wild-type and gl3–sst sim trichomes identifies four additional genes required for trichome development. Mol. Plant 2009, 2, 803–822. [Google Scholar]
- Lacape, J.-M.; Nguyen, T.-B. Mapping quantitative trait loci associated with leaf and stem pubescence in cotton. J. Hered. 2005, 96, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.; Malik, T.; Malik, S. Tagging genes for velvet hairiness in upland cotton. J. Anim. Plant 2013, 23, 1666–1670. [Google Scholar]
- Tang, M.; Wu, X.; Cao, Y.; Qin, Y.; Ding, M.; Jiang, Y.; Sun, C.; Zhang, H.; Paterson, A.H.; Rong, J. Preferential insertion of a Ty1 LTR-retrotransposon into the A sub-genome’s HD1 gene significantly correlated with the reduction in stem trichomes of tetraploid cotton. Mol. Genet. Genom. 2020, 295, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Hegarty, M.; Yadav, R.; Lee, M.; Armstead, I.; Sanderson, R.; Scollan, N.; Powell, W.; Skøt, L. Genotyping by RAD sequencing enables mapping of fatty acid composition traits in perennial ryegrass (Lolium perenne (L.)). Plant Biotechnol. J. 2013, 11, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Davey, J.W.; Hohenlohe, P.A.; Etter, P.D.; Boone, J.Q.; Catchen, J.M.; Blaxter, M.L. Genome-wide genetic marker discovery and genotyping using next-generation sequencing. Nat. Rev. Genet. 2011, 12, 499–510. [Google Scholar] [PubMed]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar]
- Jaganathan, D.; Thudi, M.; Kale, S.; Azam, S.; Roorkiwal, M.; Gaur, P.M.; Kishor, P.K.; Nguyen, H.; Sutton, T.; Varshney, R.K. Genotyping-by-sequencing based intra-specific genetic map refines a “QTL-hotspot” region for drought tolerance in chickpea. Mol. Genet. Genom. 2015, 290, 559–571. [Google Scholar] [CrossRef]
- Lee, J.; Izzah, N.K.; Choi, B.-S.; Joh, H.J.; Lee, S.-C.; Perumal, S.; Seo, J.; Ahn, K.; Jo, E.J.; Choi, G.J. Genotyping-by-sequencing map permits identification of clubroot resistance QTLs and revision of the reference genome assembly in cabbage (Brassica oleracea L.). DNA Res. 2015, 23, 29–41. [Google Scholar] [CrossRef]
- Su, C.; Wang, W.; Gong, S.; Zuo, J.; Li, S.; Xu, S. High density linkage map construction and mapping of yield trait QTLs in maize (Zea mays) using the genotyping-by-sequencing (GBS) technology. Front. Plant Sci. 2017, 8, 706. [Google Scholar] [CrossRef]
- Gao, L.; Kielsmeier-Cook, J.; Bajgain, P.; Zhang, X.; Chao, S.; Rouse, M.N.; Anderson, J.A. Development of genotyping by sequencing (GBS)-and array-derived SNP markers for stem rust resistance gene Sr42. Mol. Breed. 2015, 35, 207. [Google Scholar] [CrossRef]
- Milczarski, P.; Hanek, M.; Tyrka, M.; Stojałowski, S. The application of GBS markers for extending the dense genetic map of rye (Secale cereale L.) and the localization of the Rfc1 gene restoring male fertility in plants with the C source of sterility-inducing cytoplasm. J. Appl. Genet. 2016, 57, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Hart, J.P.; Griffiths, P.D. Genotyping-by-sequencing enabled mapping and marker development for the by-2 potyvirus resistance allele in common bean. Plant Genome 2015, 8. [Google Scholar] [CrossRef]
- Wright, R.; Thaxton, P.; El-Zik, K.; Paterson, A. Molecular mapping of genes affecting pubescence of cotton. J. Hered. 1999, 90, 215–219. [Google Scholar] [CrossRef]
- Paterson, A.H.; Brubaker, C.L.; Wendel, J.F. A rapid method for extraction of cotton (Gossypium spp.) genomic DNA suitable for RFLP or PCR analysis. Plant Mol. Biol. Report. 1993, 11, 122–127. [Google Scholar]
- Paten, B.; Novak, A.; Haussler, D. Mapping to a reference genome structure. arXiv 2014, arXiv:1404.5010. [Google Scholar]
- Van Ooijen, J. MapQTL® 5, Software for the mapping of quantitative trait loci in experimental populations. Kyazma BV Wagening. 2004, 63. [Google Scholar]
- Wang, L.; Wang, A.; Huang, X.; Zhao, Q.; Dong, G.; Qian, Q.; Sang, T.; Han, B. Mapping 49 quantitative trait loci at high resolution through sequencing-based genotyping of rice recombinant inbred lines. Theor. Appl. Genet. 2011, 122, 327–340. [Google Scholar]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar]
- Saunders, J.H. Genetics of hairiness transferred from Gossypium raimondii to G. hirsutum. Euphytica 1965, 14, 276–282. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Qin, Y.; Sun, H.; Hao, P.; Wang, H.; Wang, C.; Ma, L.; Wei, H.; Yu, S. Transcriptome analysis reveals differences in the mechanisms of fiber initiation and elongation between long- and short-fiber cotton (Gossypium hirsutum L.) lines. BMC Genom. 2019, 20, 633. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, X.-L.; Huang, G.-Q.; Li, X.-B. Molecular characterization of cotton GhTUA9 gene specifically expressed in fibre and involved in cell elongation. J. Exp. Bot. 2007, 58, 3227–3238. [Google Scholar] [PubMed]
- Mellon, J.E.; Zelaya, C.A.; Dowd, M.K.; Beltz, S.B.; Klich, M.A. Inhibitory effects of gossypol, gossypolone, and apogossypolone on a collection of economically important filamentous fungi. J. Agric. Food Chem. 2012, 60, 2740–2745. [Google Scholar] [CrossRef] [PubMed]
- Dayan, F.E.; Duke, S.O. Trichomes and root hairs: Natural pesticide factories. Pestic. Outlook 2003, 14, 175–178. [Google Scholar]
- De Moraes, C.M.; Lewis, W.; Pare, P.; Alborn, H.; Tumlinson, J. Herbivore-infested plants selectively attract parasitoids. Nature 1998, 393, 570–573. [Google Scholar] [CrossRef]
- Zhu, Z.; Xu, F.; Zhang, Y.; Cheng, Y.T.; Wiermer, M.; Li, X.; Zhang, Y. Arabidopsis resistance protein SNC1 activates immune responses through association with a transcriptional corepressor. Proc. Natl. Acad. Sci. USA 2010, 107, 13960–13965. [Google Scholar]
- Brosseau, C.; Moffett, P. Functional and genetic analysis identify a role for Arabidopsis ARGONAUTE5 in antiviral RNA silencing. Plant Cell 2015, 27, 1742–1754. [Google Scholar] [CrossRef]
- Cabot, C.; Martos, S.; Llugany, M.; Gallego, B.; Tolrà, R.; Poschenrieder, C. A role for zinc in plant defense against pathogens and herbivores. Front. Plant Sci. 2019, 10, 1171. [Google Scholar]
- He, J.; Zhao, X.; Laroche, A.; Lu, Z.-X.; Liu, H.; Li, Z. Genotyping-by-sequencing (GBS), an ultimate marker-assisted selection (MAS) tool to accelerate plant breeding. Front. Plant Sci. 2014, 5, 484. [Google Scholar] [CrossRef]
- Poland, J.A.; Brown, P.J.; Sorrells, M.E.; Jannink, J.-L. Development of high-density genetic maps for barley and wheat using a novel two-enzyme genotyping-by-sequencing approach. PLoS ONE 2012, 7, e32253. [Google Scholar] [CrossRef]
- Miao, Y.; Zentgraf, U. A HECT E3 ubiquitin ligase negatively regulates Arabidopsis leaf senescence through degradation of the transcription factor WRKY53. Plant J. 2010, 63, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Traw, M.B.; Bergelson, J. Interactive effects of jasmonic acid, salicylic acid, and gibberellin on induction of trichomes in Arabidopsis. Plant Physiol. 2003, 133, 1367–1375. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Missihoun, T.D.; Bartels, D. The role of Arabidopsis aldehyde dehydrogenase genes in response to high temperature and stress combinations. J. Exp. Bot. 2017, 68, 4295–4308. [Google Scholar] [CrossRef]
- Desprez, T.; Vernhettes, S.; Fagard, M.; Refrégier, G.; Desnos, T.; Aletti, E.; Py, N.; Pelletier, S.; Höfte, H. Resistance against herbicide isoxaben and cellulose deficiency caused by distinct mutations in same cellulose synthase isoform CESA6. Plant Physiol. 2002, 128, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Shan, C.-M.; Shangguan, X.-X.; Zhao, B.; Zhang, X.-F.; Chao, L.-M.; Yang, C.-Q.; Wang, L.-J.; Zhu, H.-Y.; Zeng, Y.-D.; Guo, W.-Z. Control of cotton fibre elongation by a homeodomain transcription factor GhHOX3. Nat. Commun. 2014, 5, 5519. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.P.; Nelissen, I.; Eggermont, K.; Broekaert, W.F. Deficiency in phytoalexin production causes enhanced susceptibility of Arabidopsis thaliana to the fungus Alternaria brassicicola. Plant J. 1999, 19, 163–171. [Google Scholar] [CrossRef]
- Lin, F.; Fan, J.; Rhee, S.Y. QTG-Finder: A machine-learning based algorithm to prioritize causal genes of quantitative trait loci in Arabidopsis and rice. G3 Genes Genomes Genet. 2019, 9, 3129–3138. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Minimum | Maximum | Mean | Std. Deviation |
|---|---|---|---|---|
| LPA_HN | 0.330 | 40.670 | 13.230 | 10.040 |
| LPA_AY | 0.067 | 33.533 | 10.868 | 7.512 |
| AR_AY | 6.250 | 100.000 | 41.639 | 22.033 |
| SPA_HN | 1.000 | 3.000 | 2.099 | 0.634 |
| AG_AY | 1.000 | 5.000 | 2.746 | 1.577 |
| Variables | LPA_HN | SPA_HN | AR_AY | AG_AY | LPA_AY |
|---|---|---|---|---|---|
| LPA_HN | 1 | ||||
| SPA_HN | ** 0.452 | 1 | |||
| AR_AY | * 0.235 | ** 0.365 | 1 | ||
| AG_AY | ** 0.330 | ** 0.437 | ** 0.693 | 1 | |
| LPA_AY | ** 0.424 | ** 0.588 | ** 0.375 | ** 0.434 | 1 |
| QTLs | Chromosome | Position(cM) | Start bp | End bp | #Genes | LOD | R2 | Additive_effect | Dominant_effect | DPE |
|---|---|---|---|---|---|---|---|---|---|---|
| qSPA 1_A06 | A06 | 163.41 | 88454646 | 95426881 | 146 | 14.85 | 0.16 | 0.52 | 0.17 | LDM |
| qSPA 2_A06 | A06 | 180.71 | 45870860 | 63574389 | 85 | 13.22 | 0.19 | 0.51 | 0.08 | LDM |
| qLPA 1_A06 | A06 | 481.81 | 90237872 | 93497786 | 55 | 13.94 | 0.31 | 6.30 | −0.44 | LDM |
| qLPA 2_A06 | A06 | 491.21 | 46092 | 185343 | 12 | 17.22 | 0.39 | 6.8784 | −1.15 | LDM |
| qLPA 3_A06 | A06 | 499.41 | 91597800 | 91713664 | 2 | 16.00 | 0.33 | 6.7449 | −0.43 | LDM |
| qLPA 4_A06 | A06 | 158.81 | 95396635 | 95426819 | 1 | 6.45 | 0.05 | 5.19 | 2.95 | LDM |
| qLPA 5_A06 | A06 | 169.71 | 88651703 | 88804649 | 0 | 9.17 | 0.08 | 6.24 | 3.15 | LDM |
| qAR 1_D09 | D09 | 168.31 | 32855135 | 34029175 | 60 | 3.20 | 0.10 | −7.3756 | 6.20 | Zong 128 |
| qAG 1_A06 | A06 | 266.01 | 89207941 | 89941409 | 11 | 2.97 | 0.09 | −0.54 | 0.39 | Zong 128 |
| qAG 2_A06 | A06 | 274.01 | 88955098 | 89801937 | 12 | 3.21 | 0.10 | −0.50 | 0.56 | Zong 128 |
| Trait name | QTL | ID | Gene | Description | Start (bp) | End (bp) |
|---|---|---|---|---|---|---|
| Stem pub. | qSPA 1 | Gh_A06G1357 | UPL5 | E3 ubiquitin-protein ligase UPL5 | 95,039,259 | 95,042,363 |
| Stem pub. | qSPA 1 | Gh_A06G1244 | UBC34 | Ubiquitin-conjugating enzyme E2 34 | 89,677,173 | 89,679,975 |
| Stem and leaf pub. | qSPA 1 and qLPA 1 | Gh_A06G1257 | ALDH7B4 | Aldehyde dehydrogenase family 7 member B4 | 90,765,222 | 90,768,706 |
| Stem pub. | qSPA 1 | Gh_A06G1253 | MPE3 | Pectinesterase 3 | 90,103,159 | 90,105,870 |
| Stem pub. | qSPA 2 | Gh_A06G0984 | TUBA5 | Tubulin alpha-5 chain | 46,567,039 | 46,569,490 |
| Stem pub. | qSPA 2 | Gh_A06G1017 | CESA6 | Cellulose synthase A catalytic subunit 6 [UDP-forming] | 50,462,282 | 50,468,328 |
| Leaf pub. | qLPA 2 | Gh_A06G0019 | PER64 | Peroxidase 64 | 84,546 | 85,807 |
| Leaf pub. | qLPA 2 | Gh_A06G0018 | EXPA1 | Expansin-A1 | 82,222 | 83,190 |
| Aphid rate | qAR 1 | Gh_D09G0835 | TPR1 | Topless-related protein 1 | 32,915,885 | 32,923,301 |
| Aphid rate | qAR 1 | Gh_D09G0839 | AGO5 | Protein argonaute 5 | 33,188,202 | 33,202,318 |
| Aphid rate | qAR 1 | Gh_D09G0857 | ZAT5 | Zinc finger protein ZAT5 | 33,598,639 | 33,599,409 |
| Aphid grade and Stem pub. | qAG 1 and qSPA 1 | Gh_A06G1246 | GLO4 | Peroxisomal (S)-2-hydroxy-acid oxidase GLO4 | 89,813,435 | 89,822,109 |
| Leaf pub. | qLPA 1 | Gh_A06G1283 | PDF2 | Homeobox-leucine zipper protein PROTODERMAL FACTOR 2 | 92,578,328 | 92,581,427 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, H.; Nazir, M.F.; Pan, Z.; Gong, W.; Iqbal, M.S.; He, S.; Du, X. Genotyping by Sequencing Revealed QTL Hotspots for Trichome-Based Plant Defense in Gossypium hirsutum. Genes 2020, 11, 368. https://doi.org/10.3390/genes11040368
Ahmed H, Nazir MF, Pan Z, Gong W, Iqbal MS, He S, Du X. Genotyping by Sequencing Revealed QTL Hotspots for Trichome-Based Plant Defense in Gossypium hirsutum. Genes. 2020; 11(4):368. https://doi.org/10.3390/genes11040368
Chicago/Turabian StyleAhmed, Haris, Mian Faisal Nazir, Zhoe Pan, Wenfang Gong, Muhammad Shahid Iqbal, Shoupu He, and Xiongming Du. 2020. "Genotyping by Sequencing Revealed QTL Hotspots for Trichome-Based Plant Defense in Gossypium hirsutum" Genes 11, no. 4: 368. https://doi.org/10.3390/genes11040368
APA StyleAhmed, H., Nazir, M. F., Pan, Z., Gong, W., Iqbal, M. S., He, S., & Du, X. (2020). Genotyping by Sequencing Revealed QTL Hotspots for Trichome-Based Plant Defense in Gossypium hirsutum. Genes, 11(4), 368. https://doi.org/10.3390/genes11040368