Seascape Genetics and the Spatial Ecology of Juvenile Green Turtles

 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Genetic Data Collection and Analysis
2.1.1. Sample Collection
2.1.2. DNA Extraction, Sequencing and Regional Data Compilation
2.1.3. Data Analysis
2.2. Passive Drift Simulations
3. Results
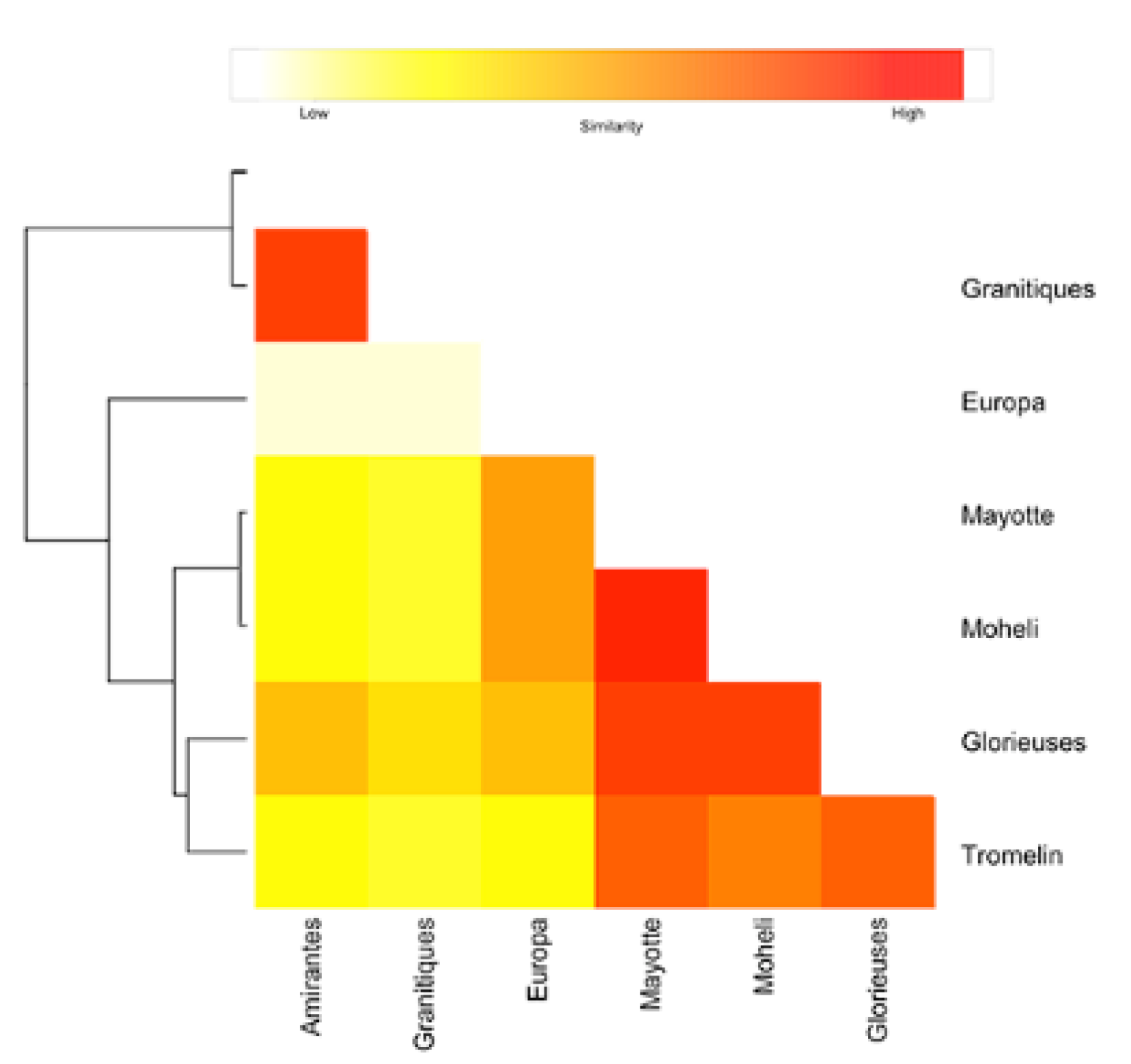
3.1. Genetic Diversity
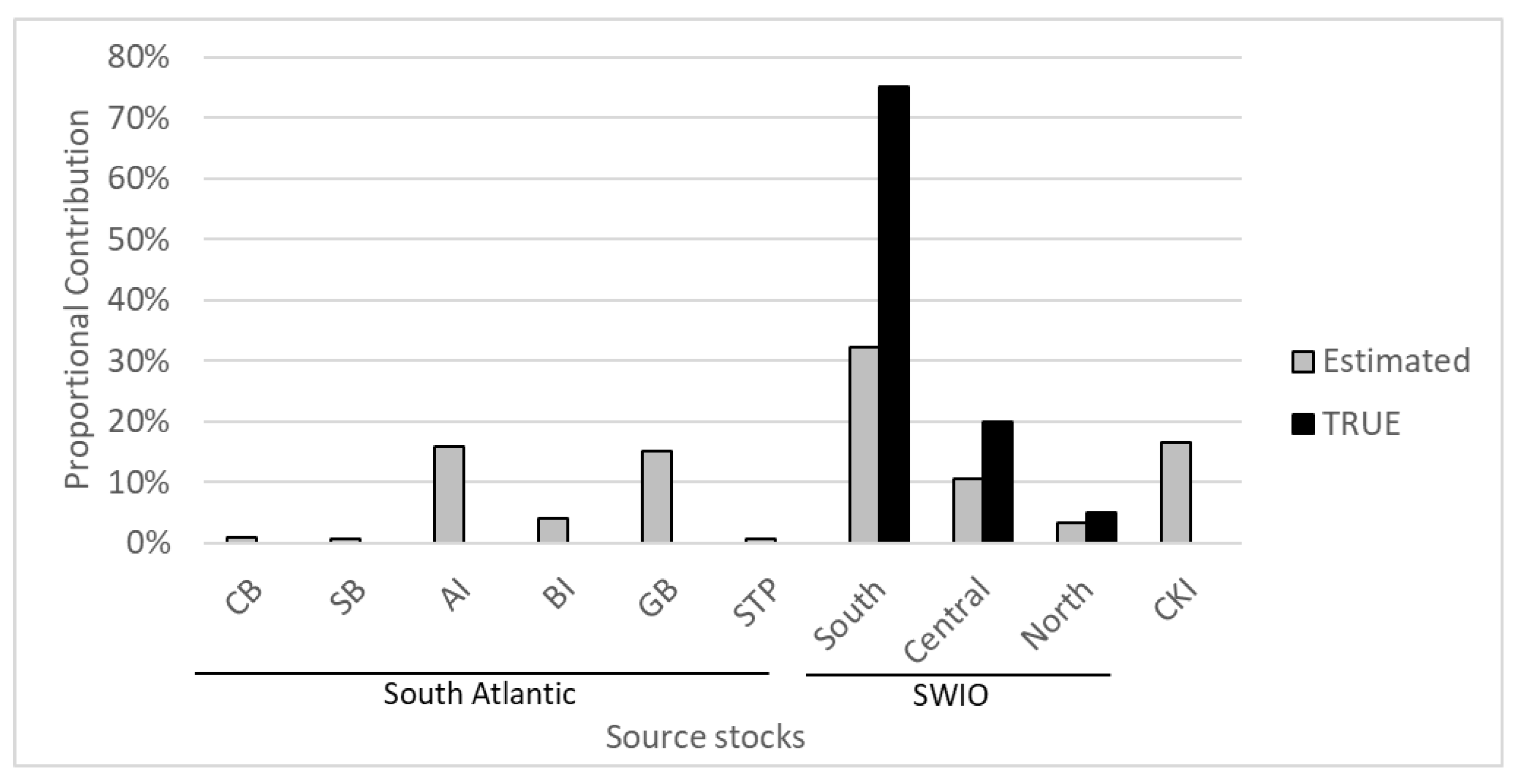
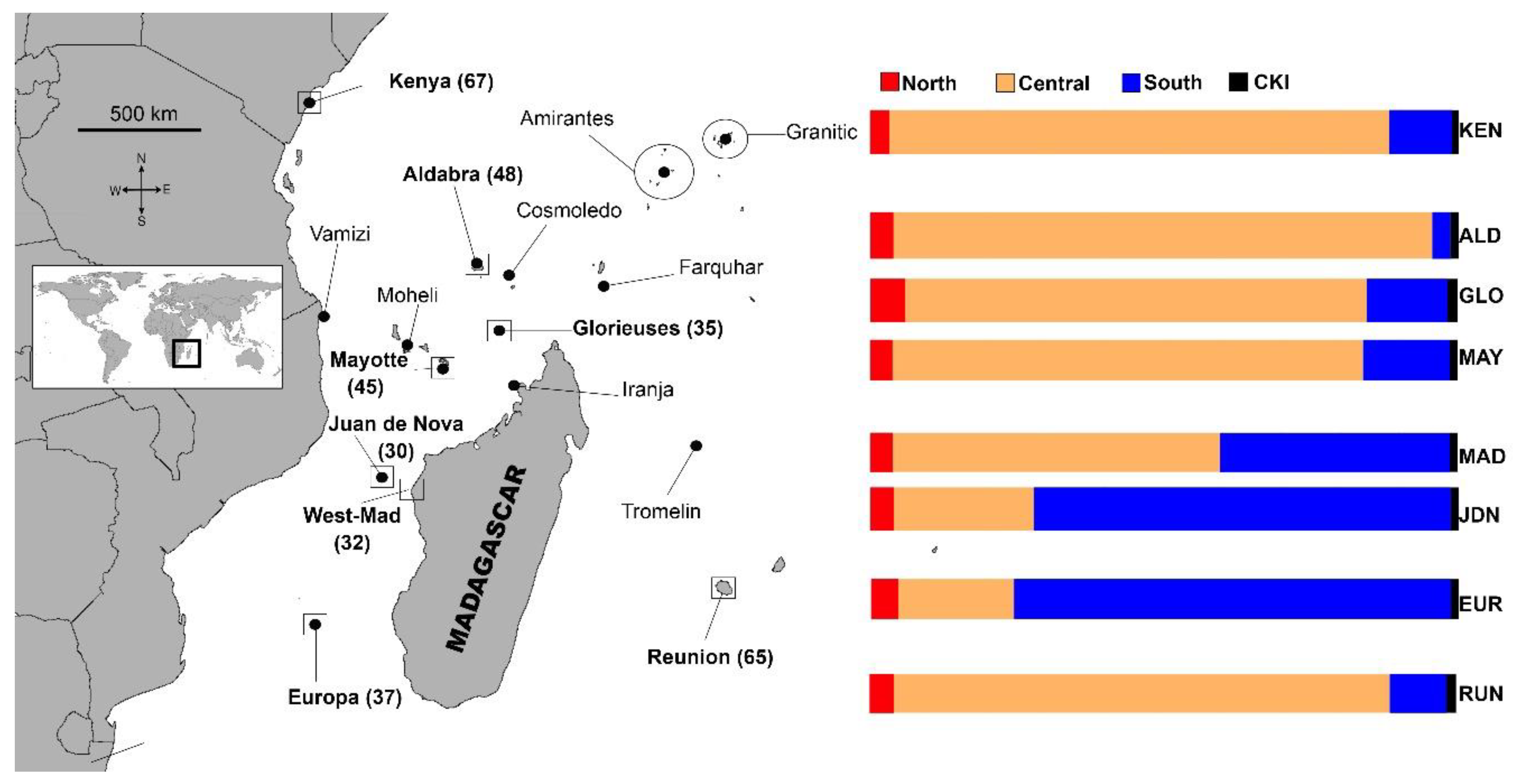
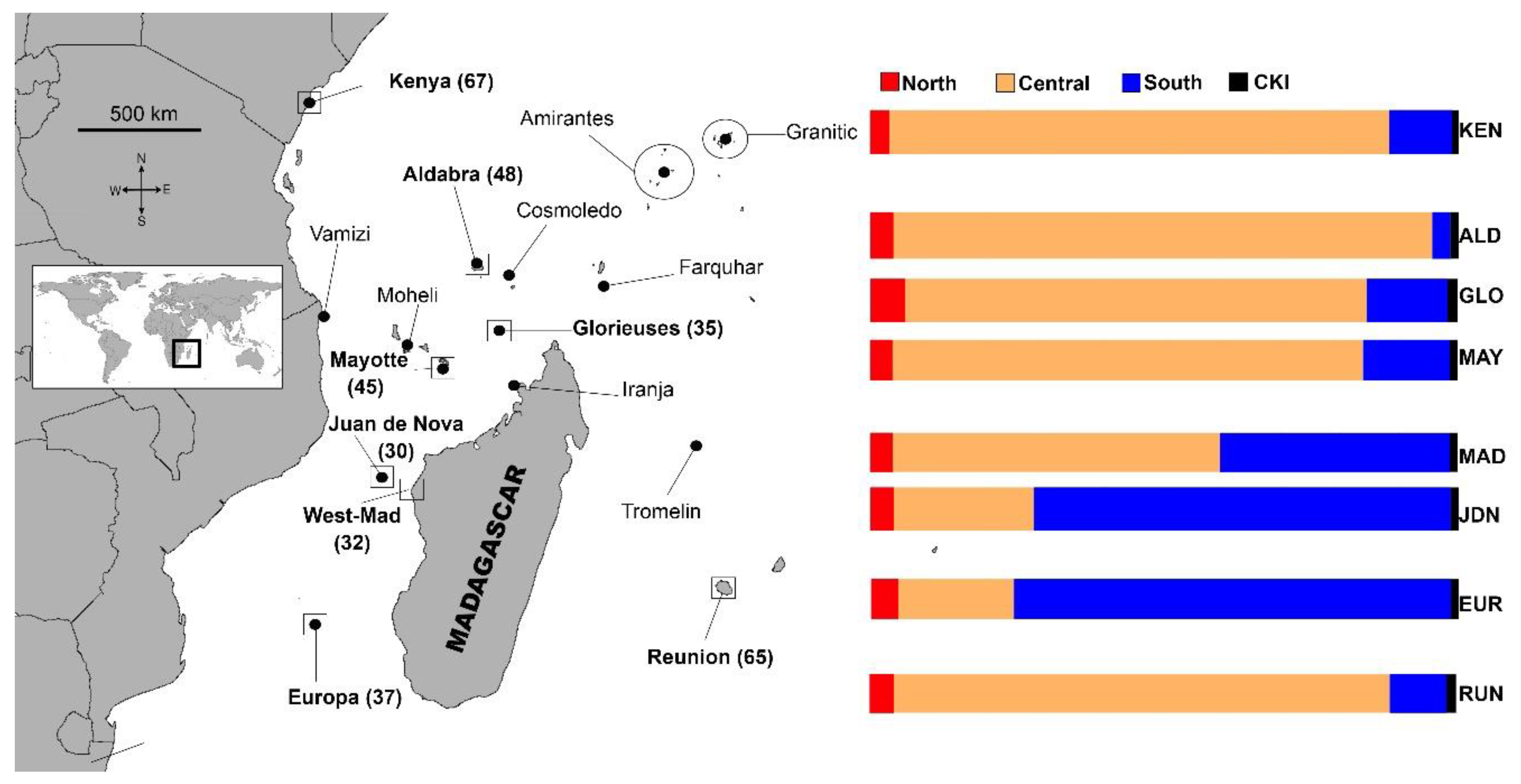
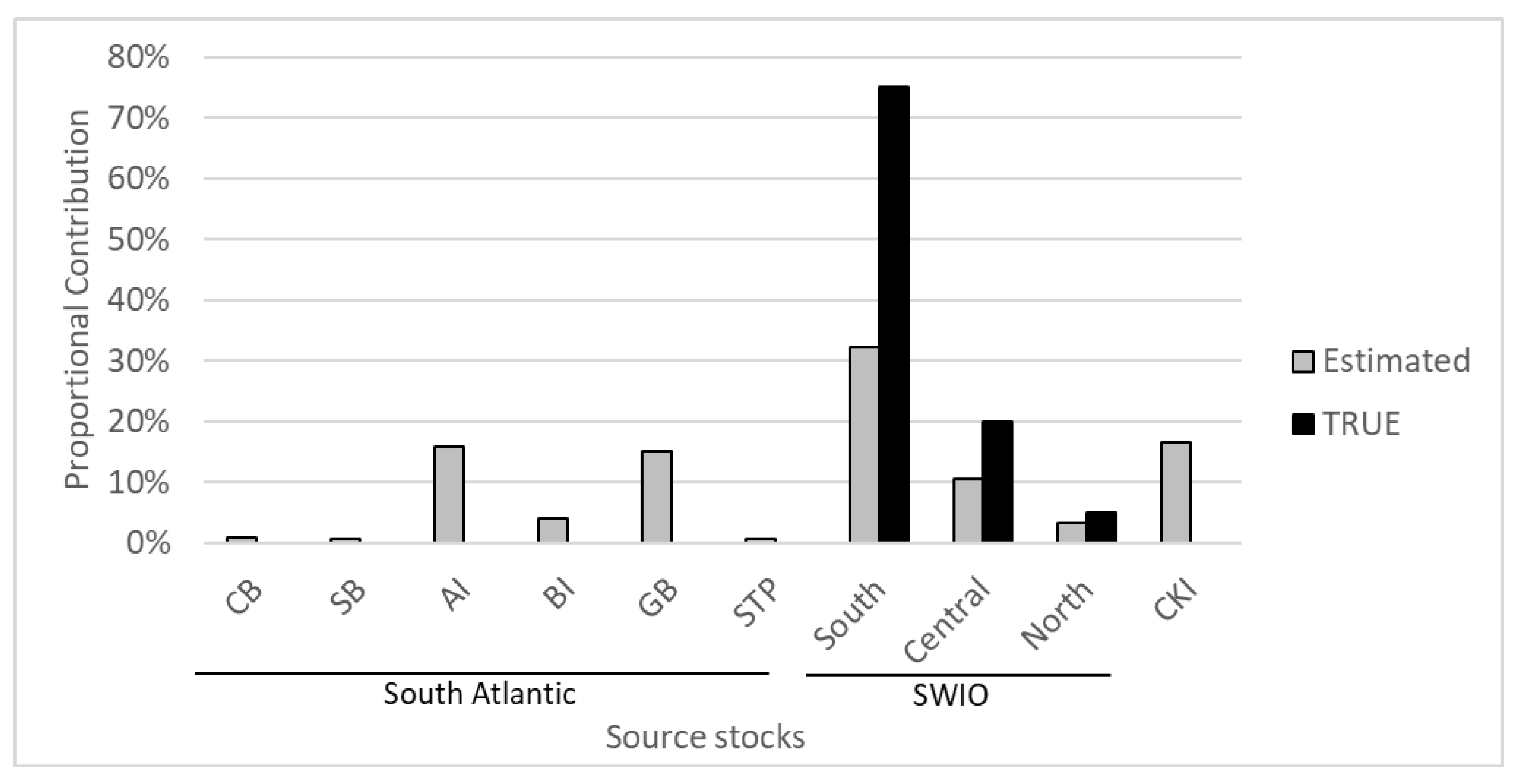
3.2. Mixed Stock Analysis
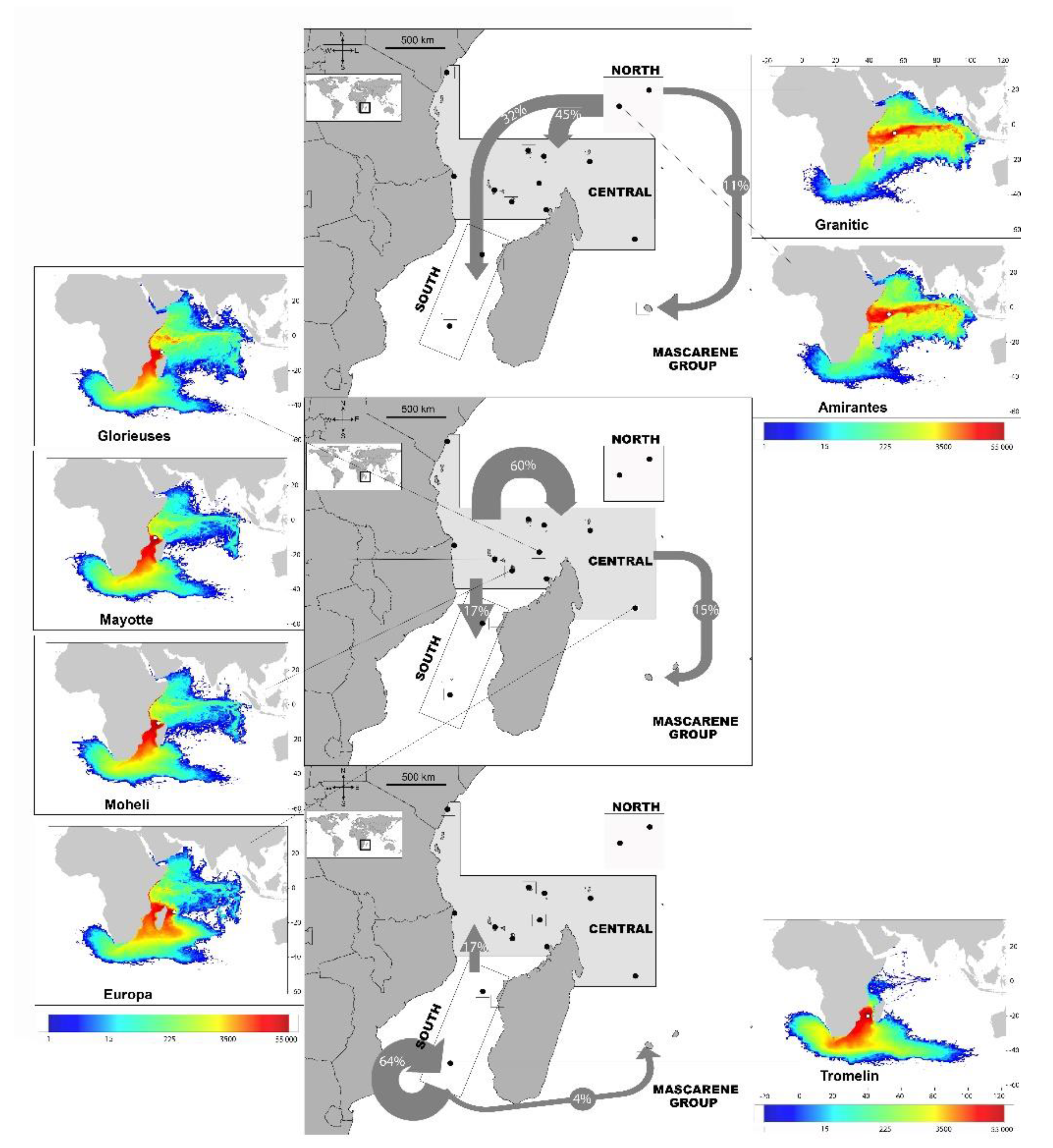
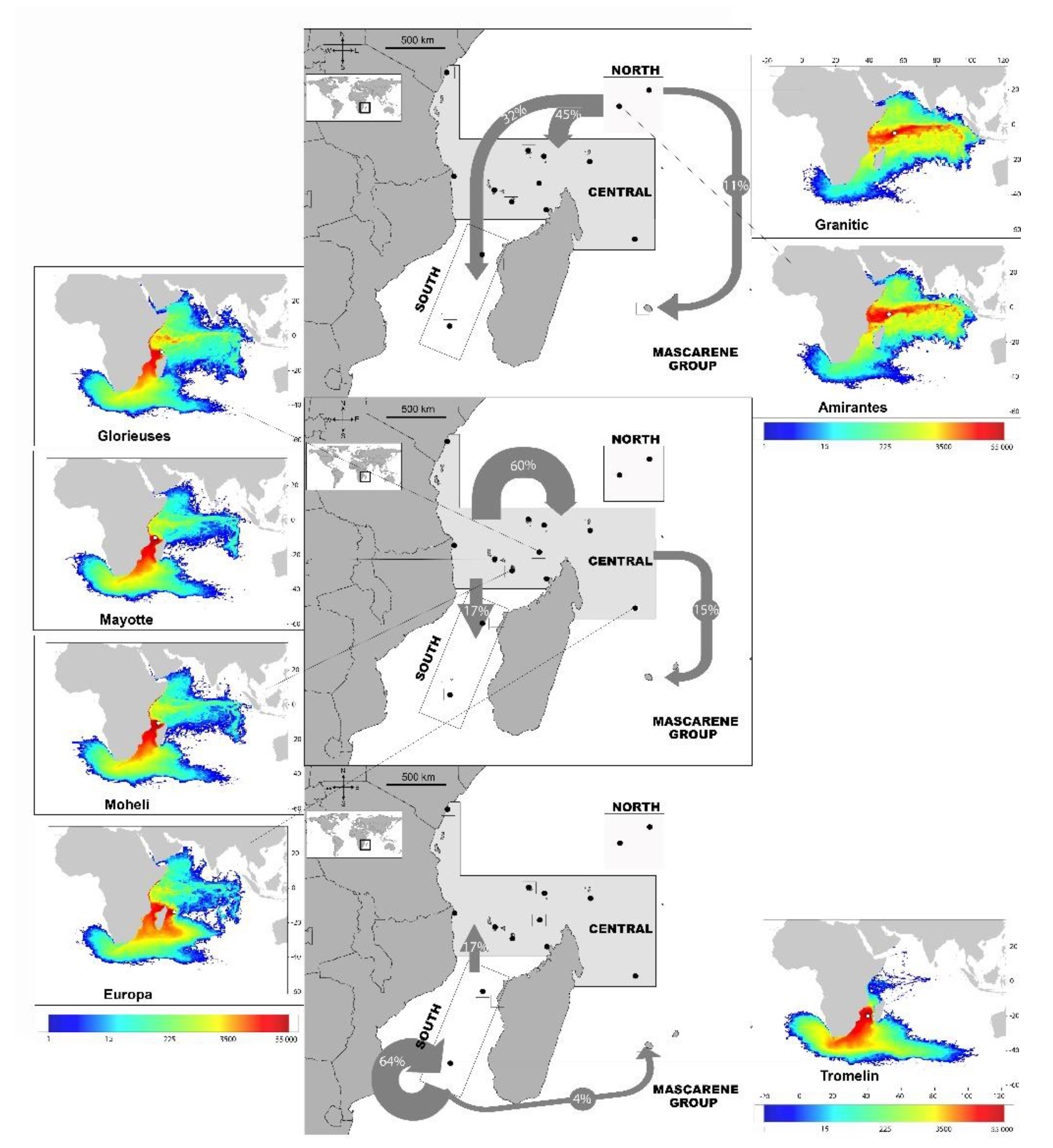
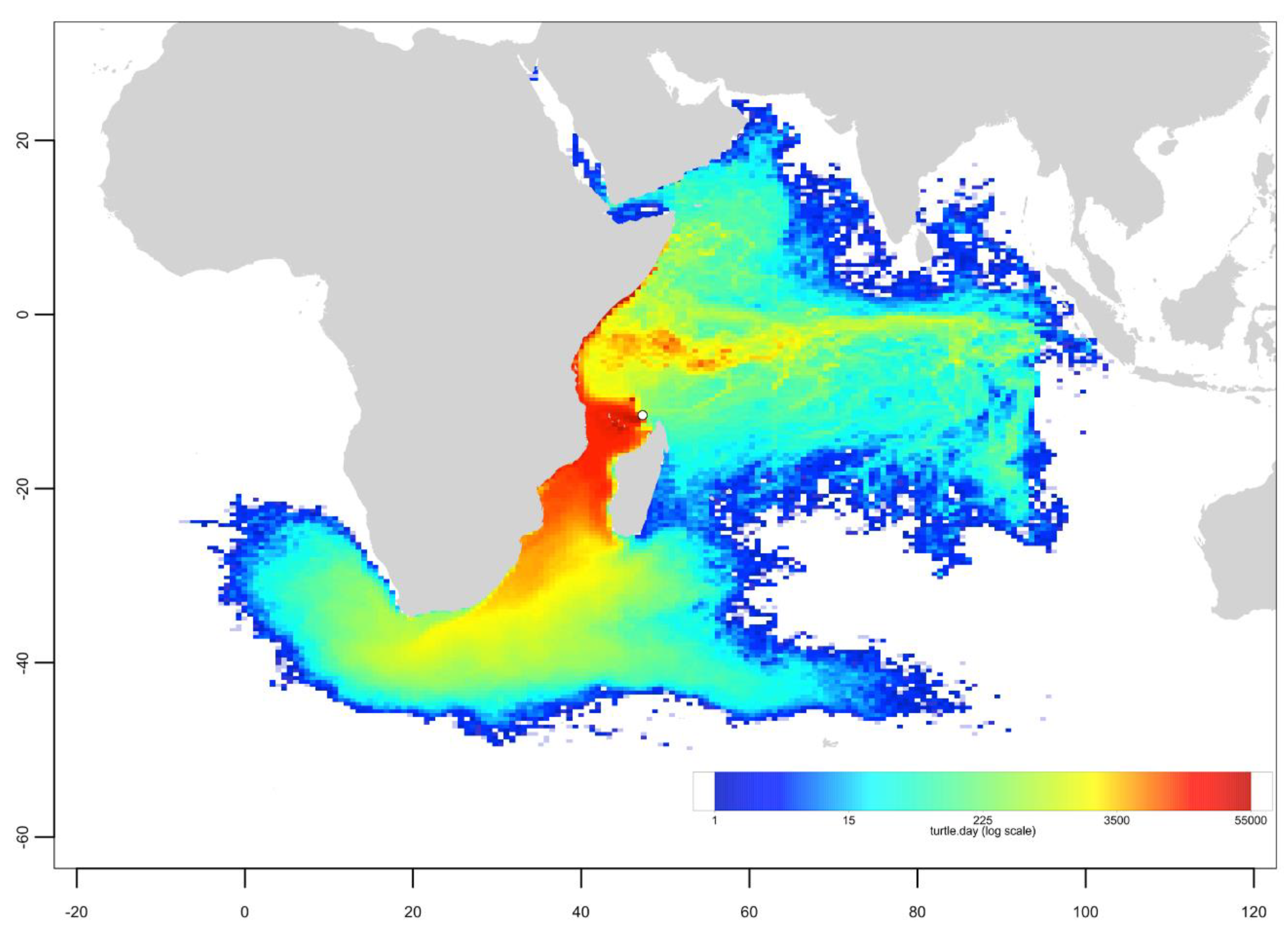
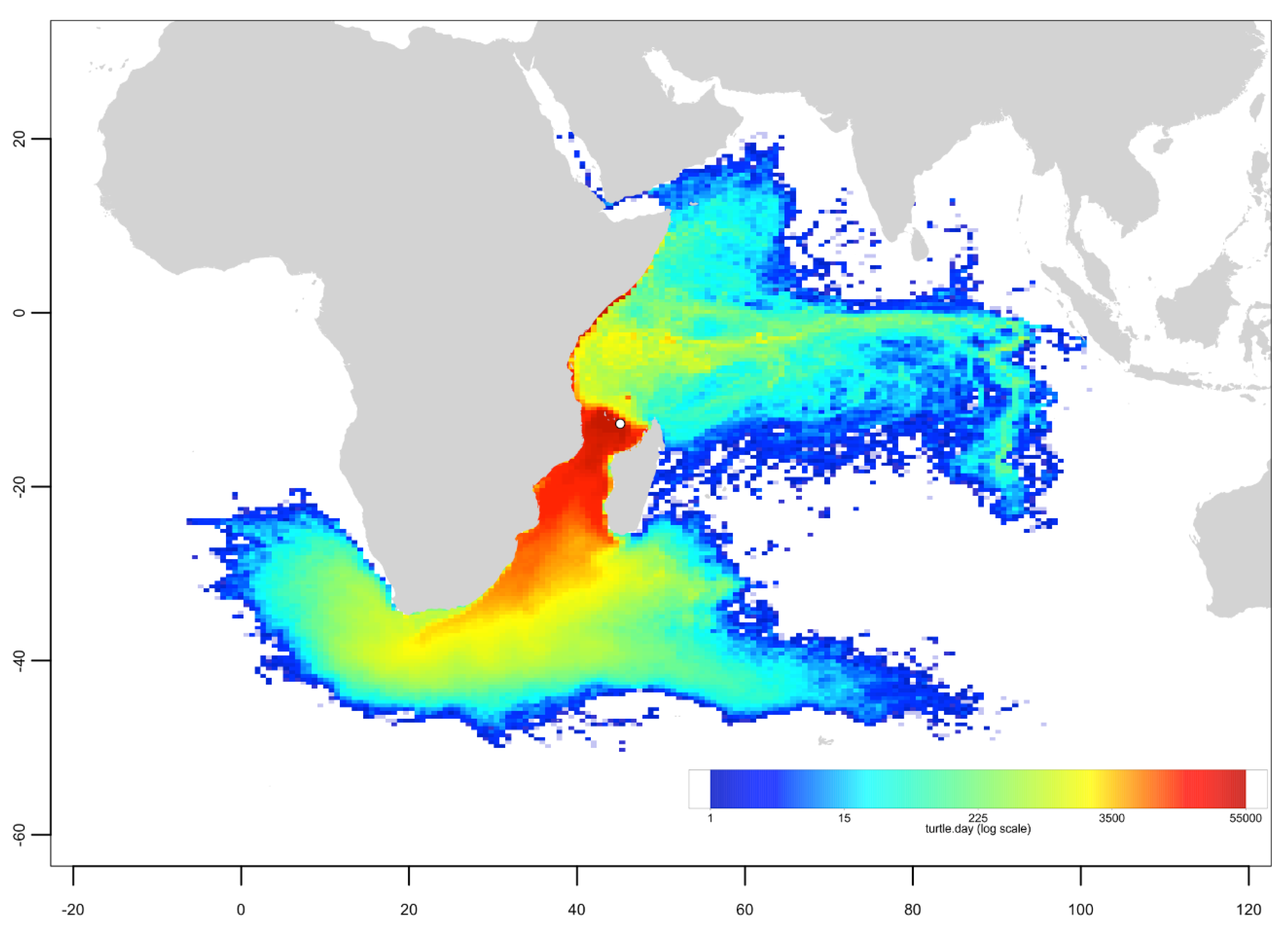
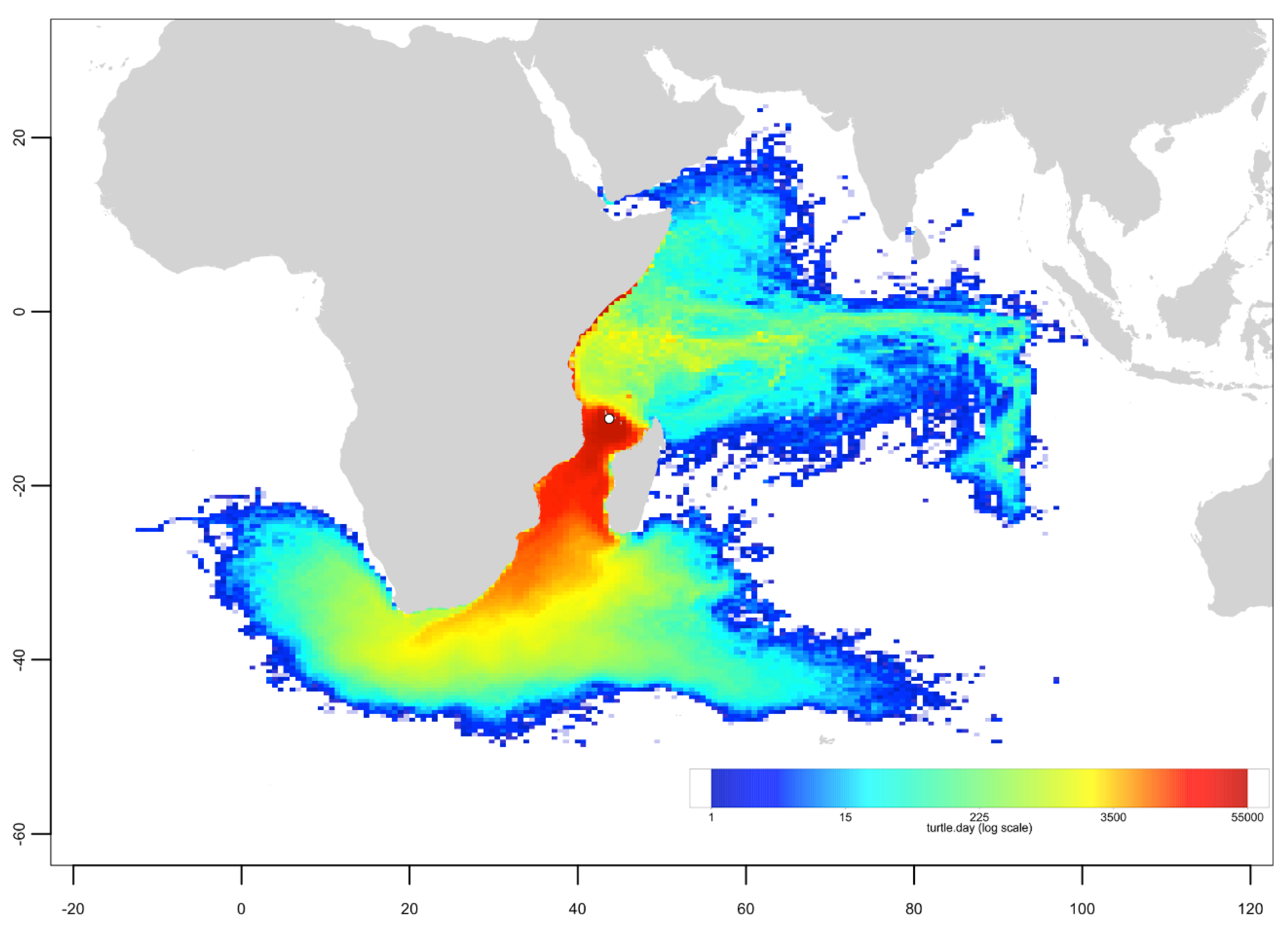
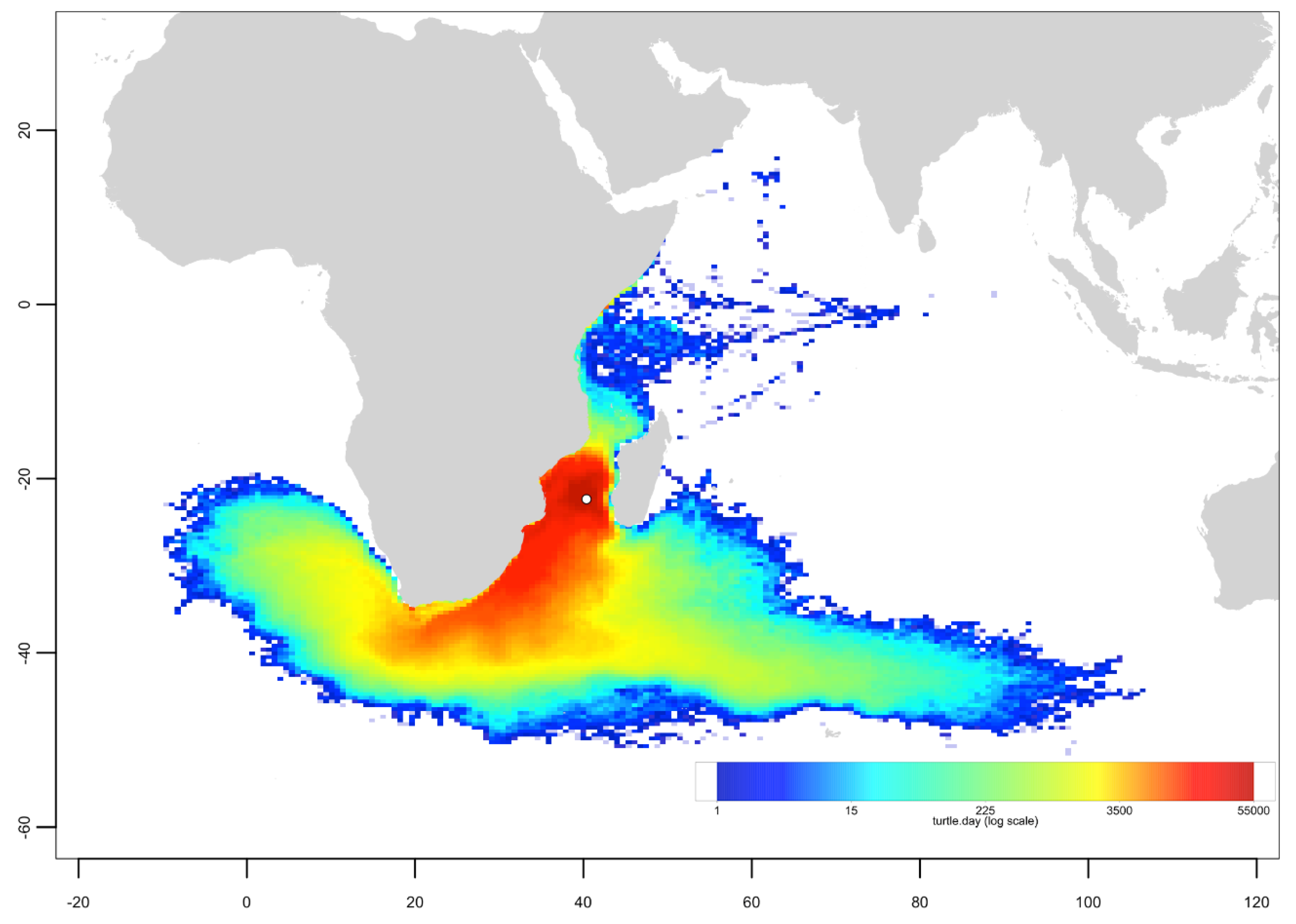
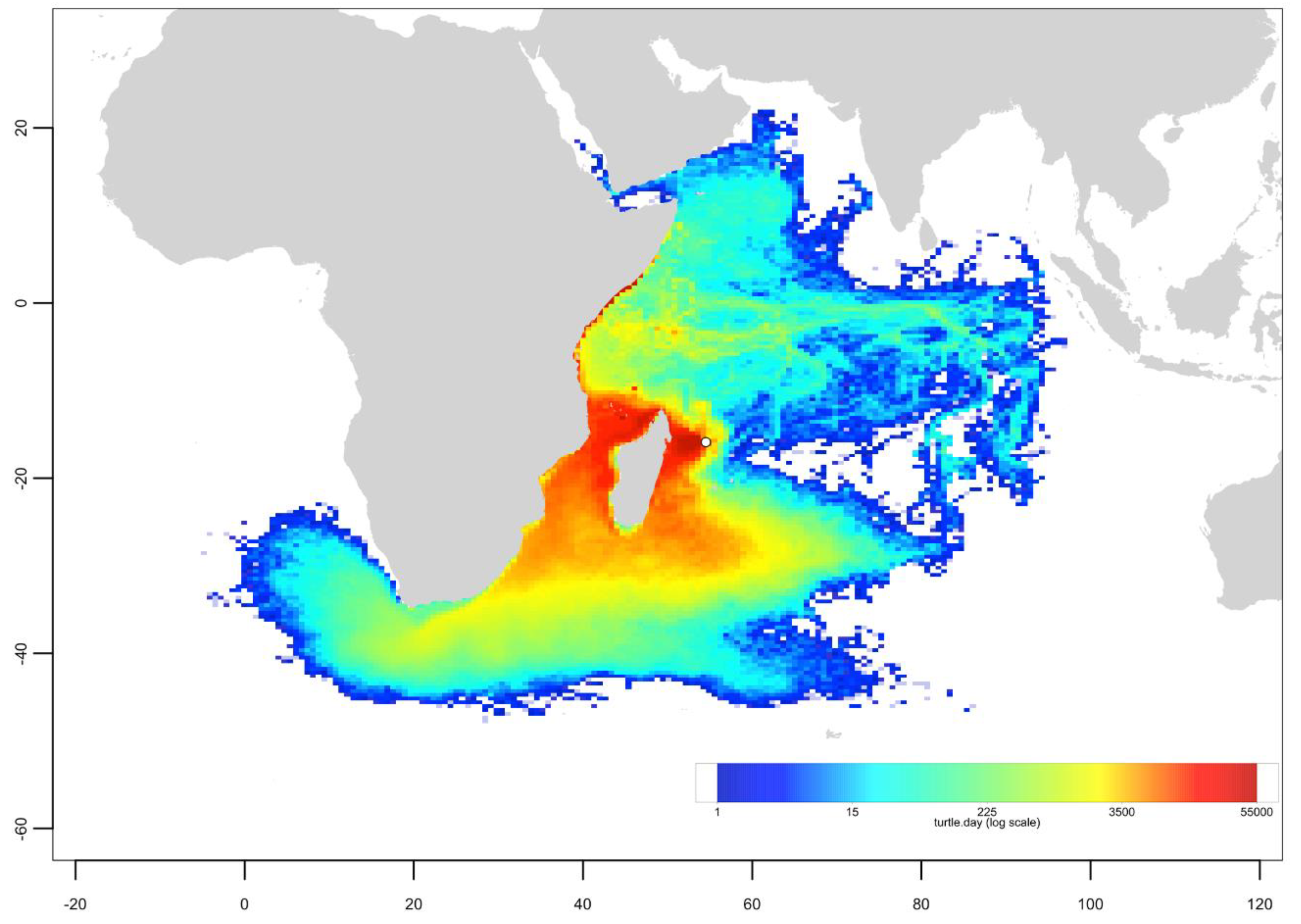
3.3. Passive Dispersal Simulations
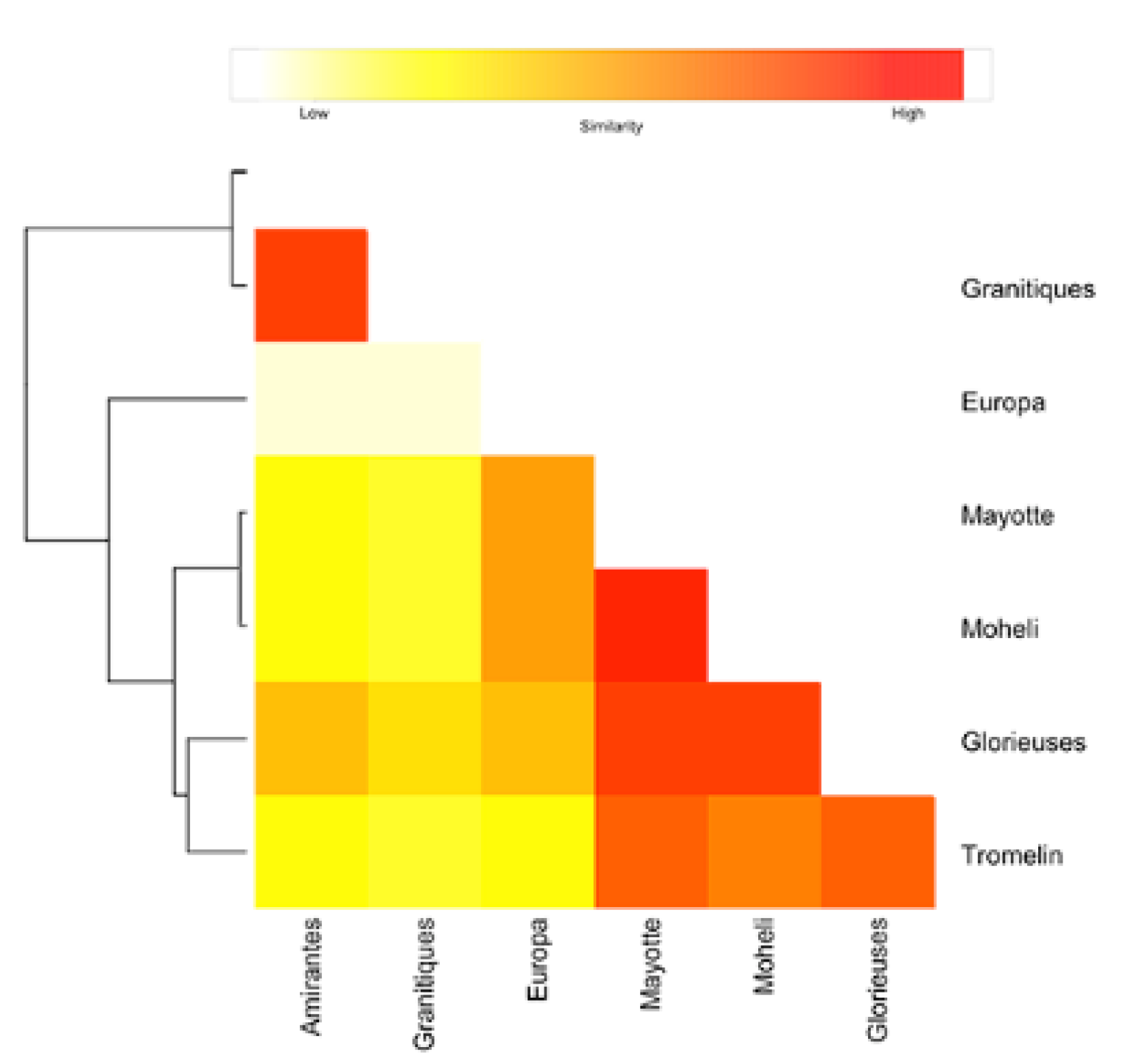
- The dispersal schemes (and corresponding density maps) associated with the nesting sites within the same Areas (North, Central and South) are similar.
- The largest differences (smallest connectivity) are observed between the juvenile density maps of the South (Europa) and North (Seychelles granitics and Amirantes) Areas.
- Nesting beaches of the Central Area display some connectivity with both the South and North Areas. As expected, Europa is more connected with Mayotte and Mohéli through the mesoscale eddy activity that mostly takes place on the western side of the Mozambique Channel. Connectivity with Glorieuses, lying further to the East, is more limited.
4. Discussion
4.1. Source Rookeries Included in the Analysis
4.2. The North Area
4.3. The Central Area
4.4. The South Area
4.5. The Case of La Réunion
4.6. Regional Connectivity Lessons from the SWIO
4.7. Limitations and Future Directions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
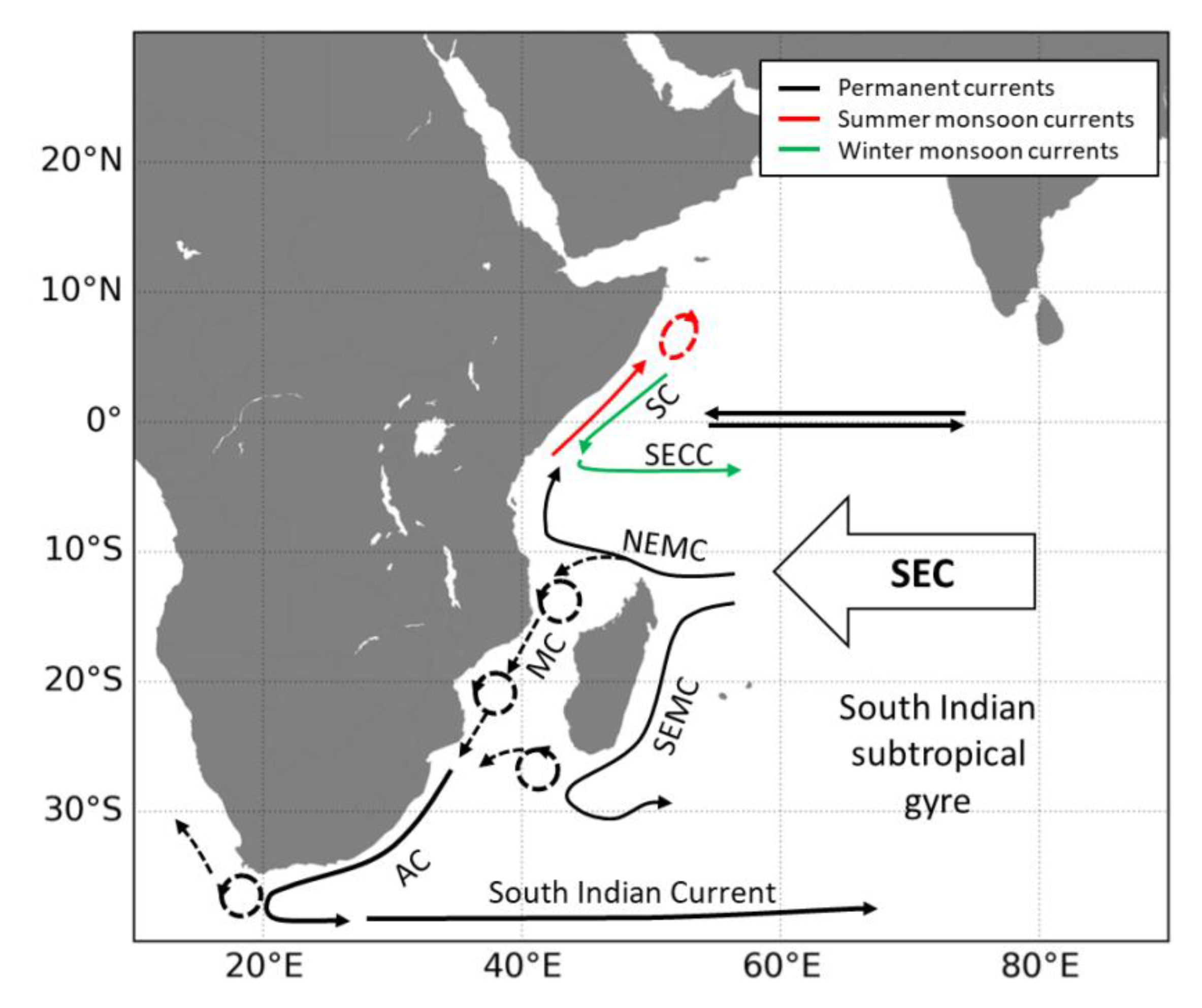
Appendix A. Main Ocean Circulation Patterns in the West Indian Ocean
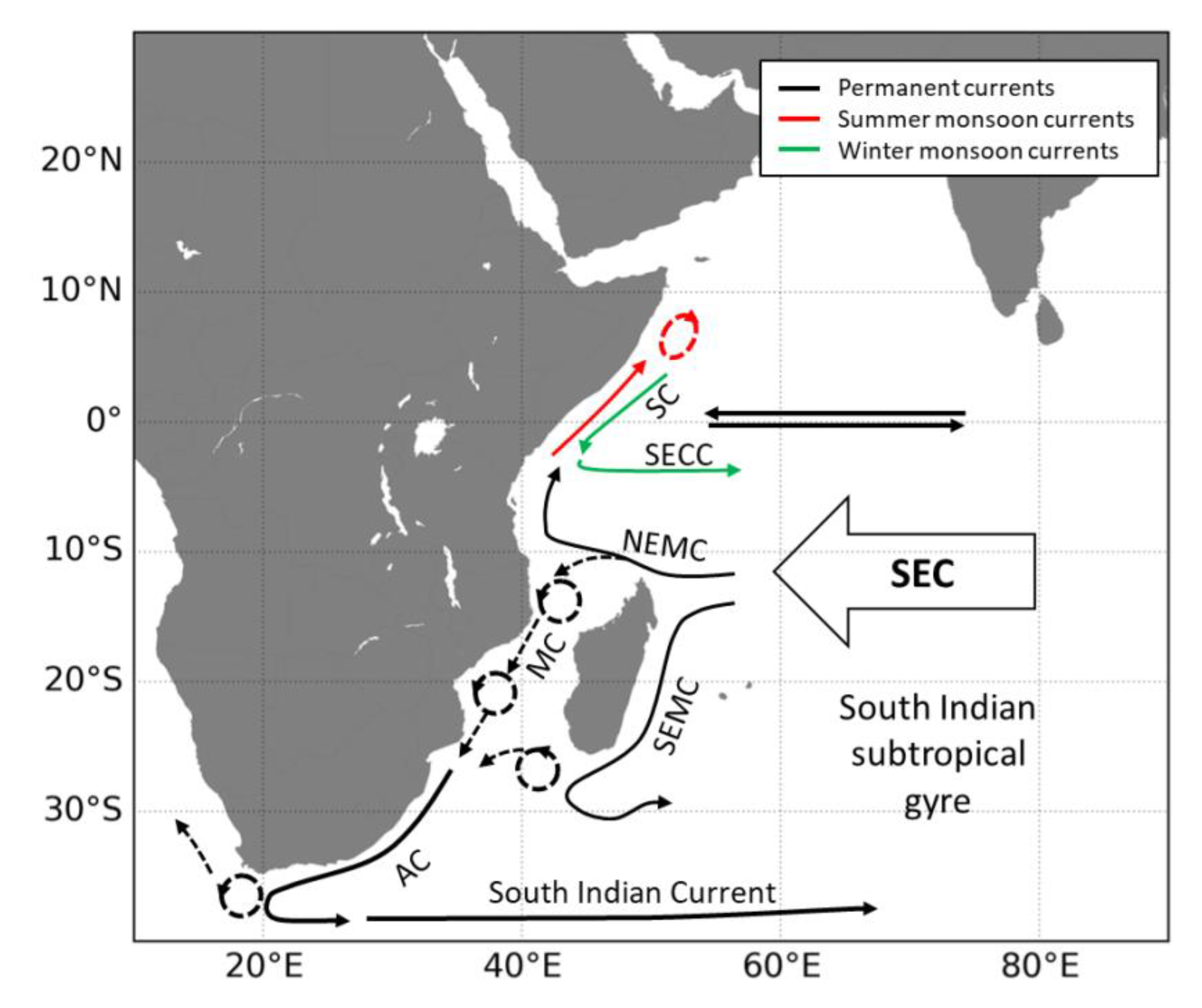
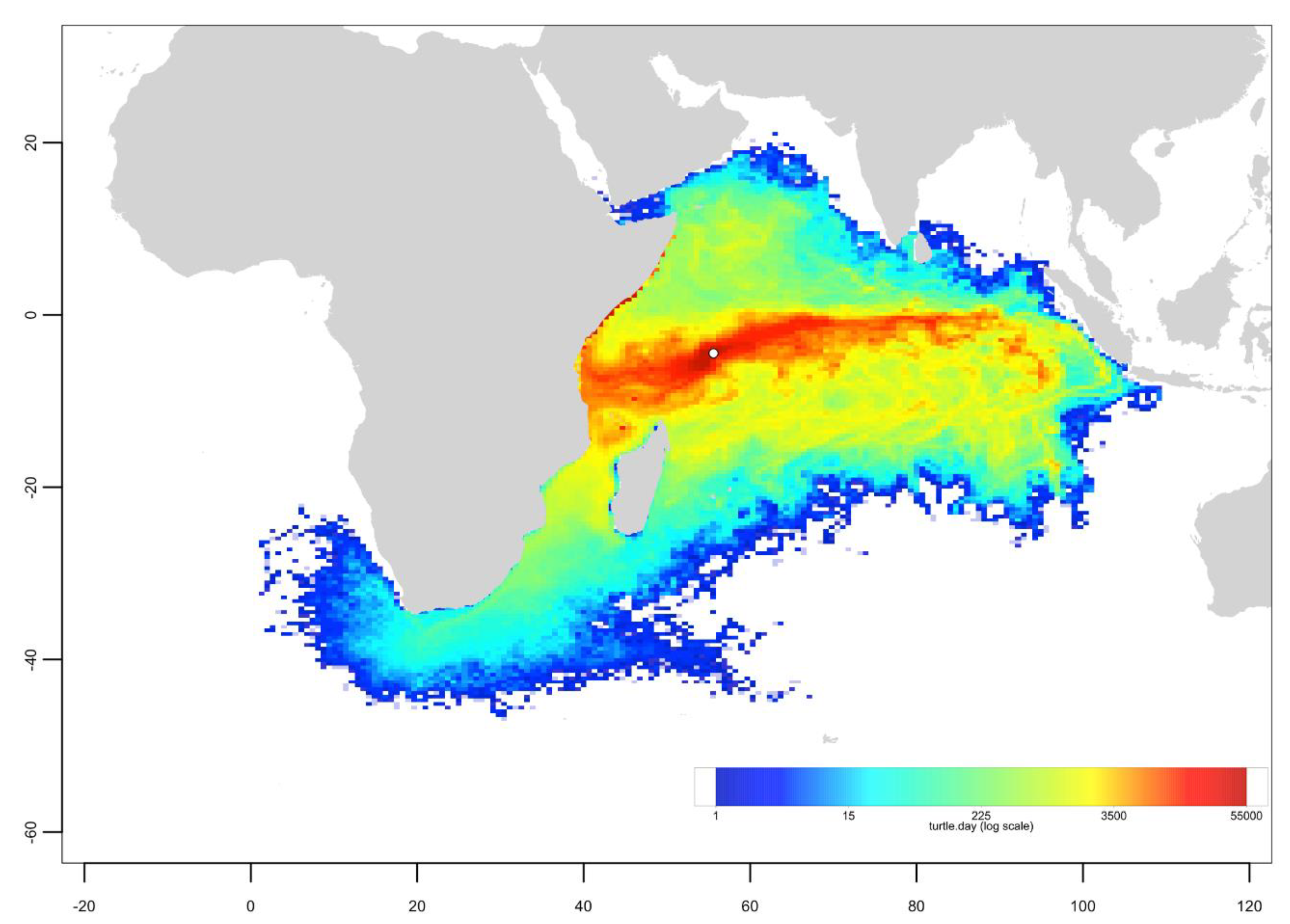
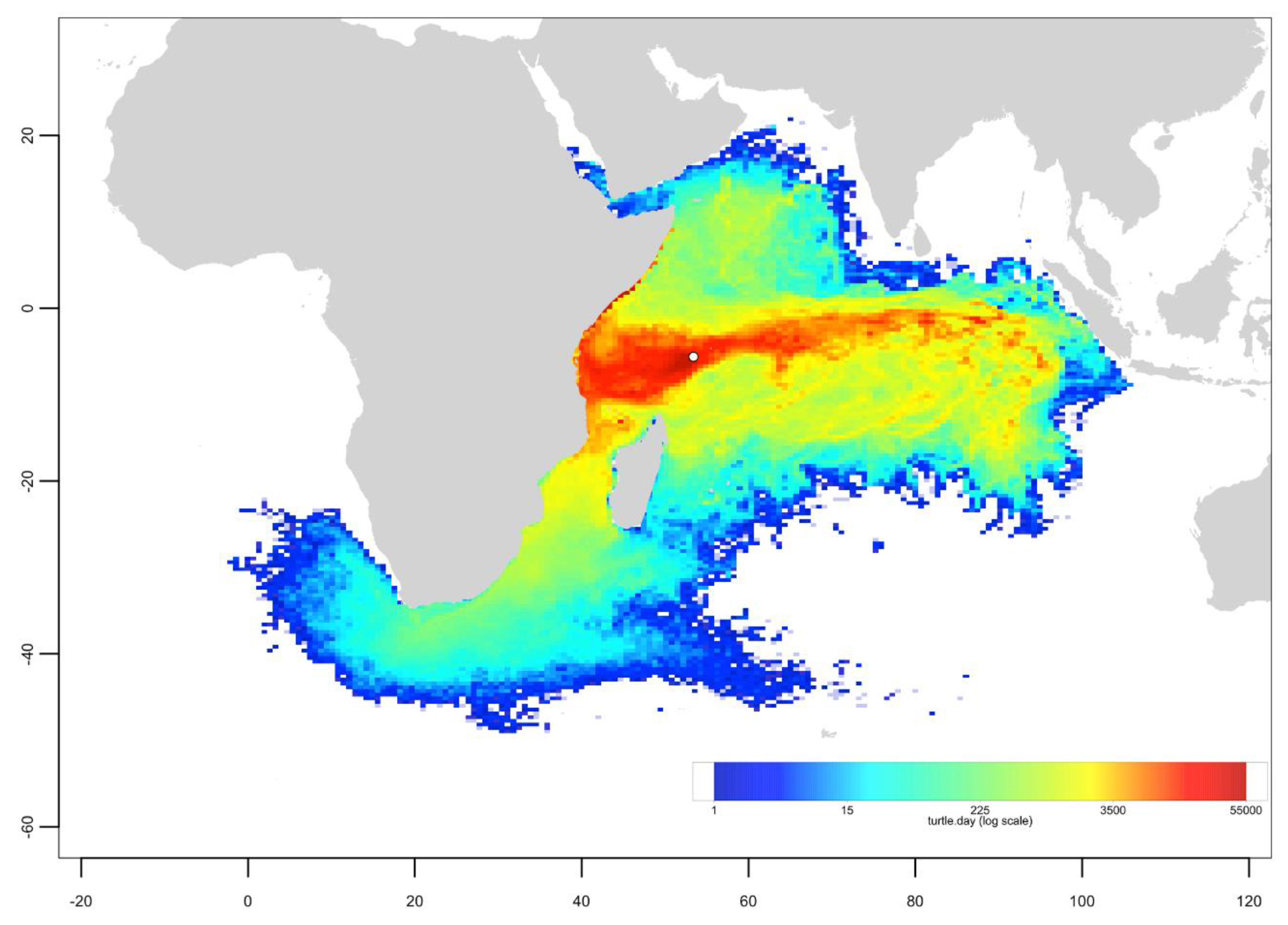
Appendix B. Particle Dispersal Maps
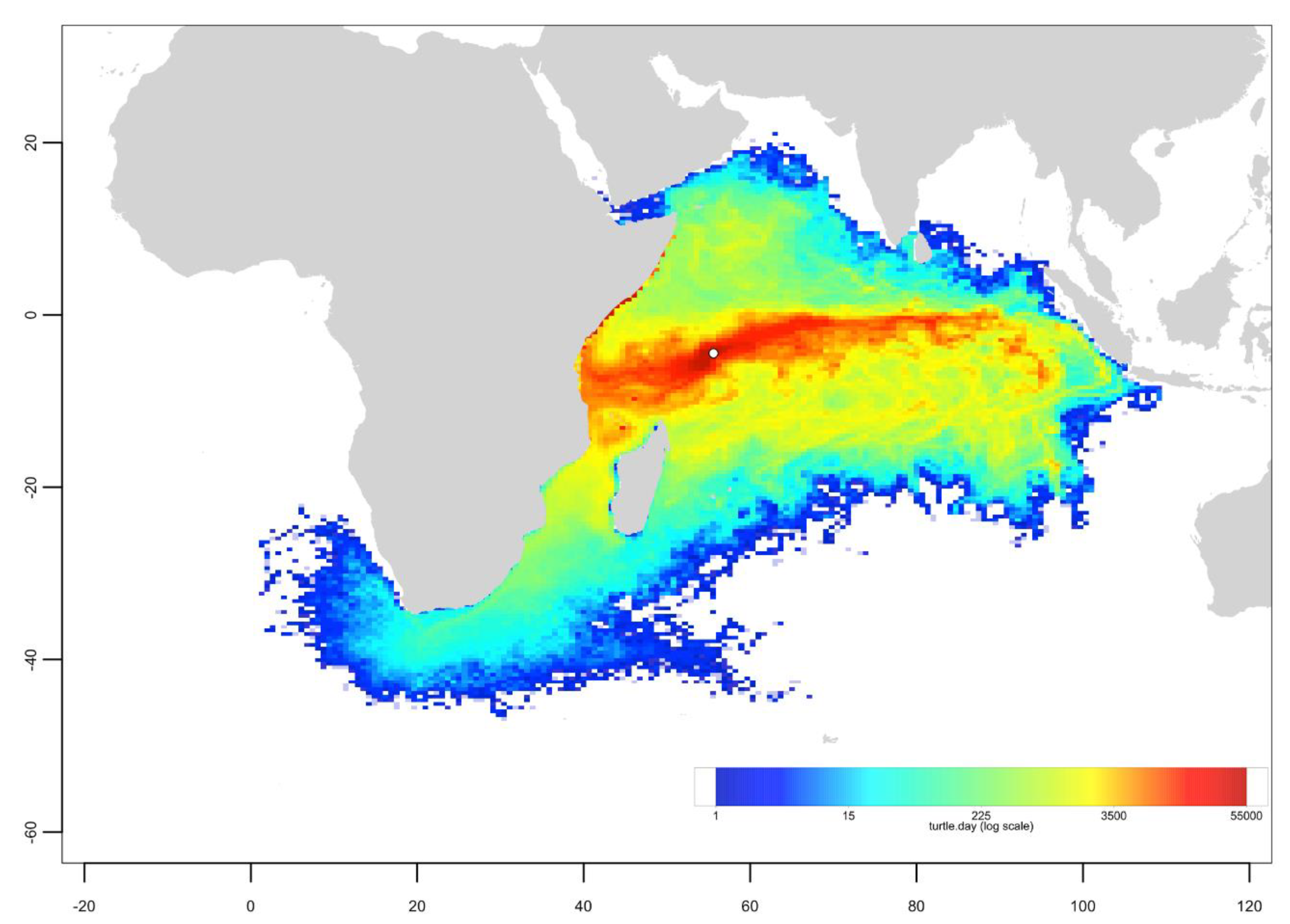
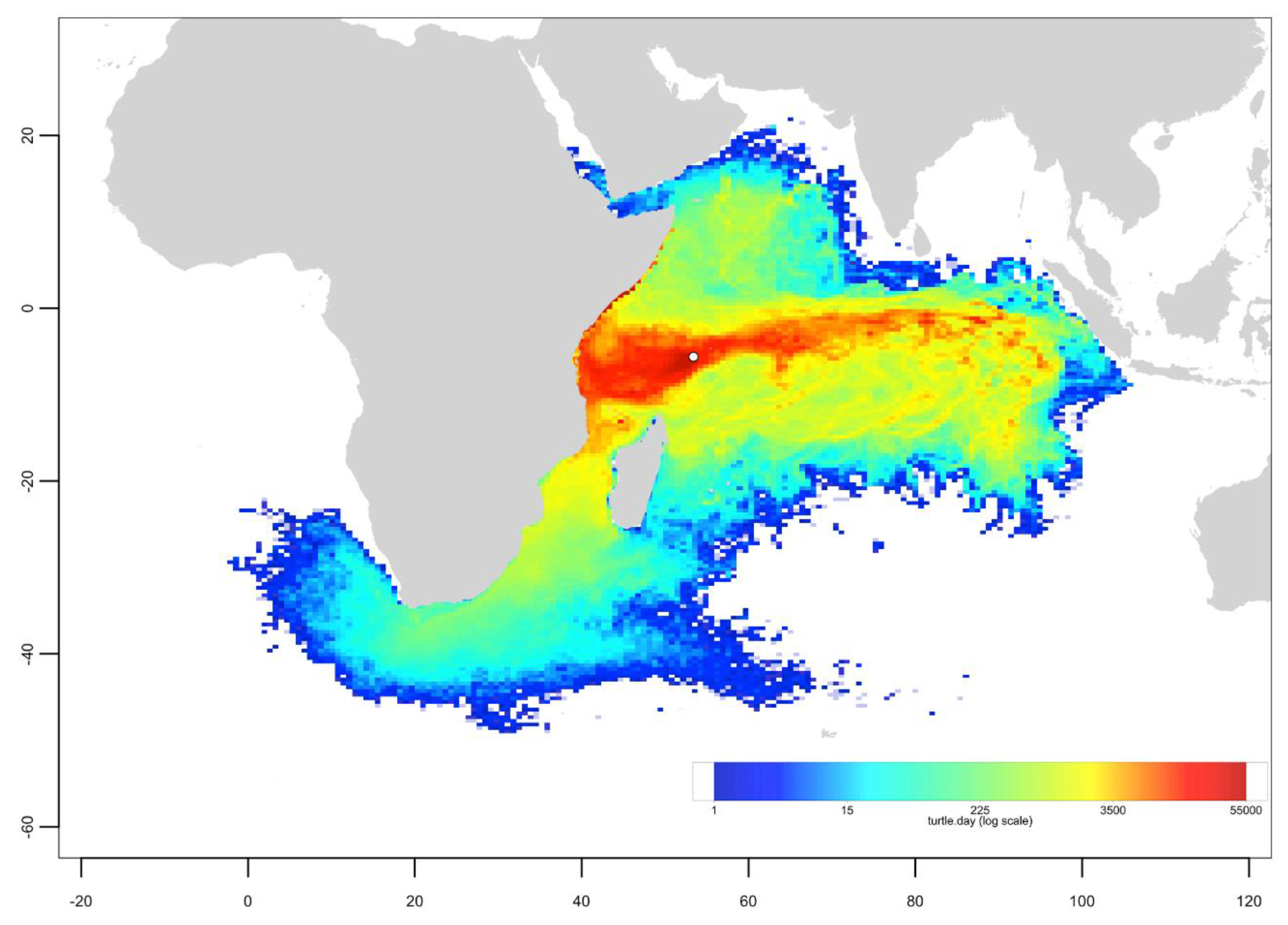
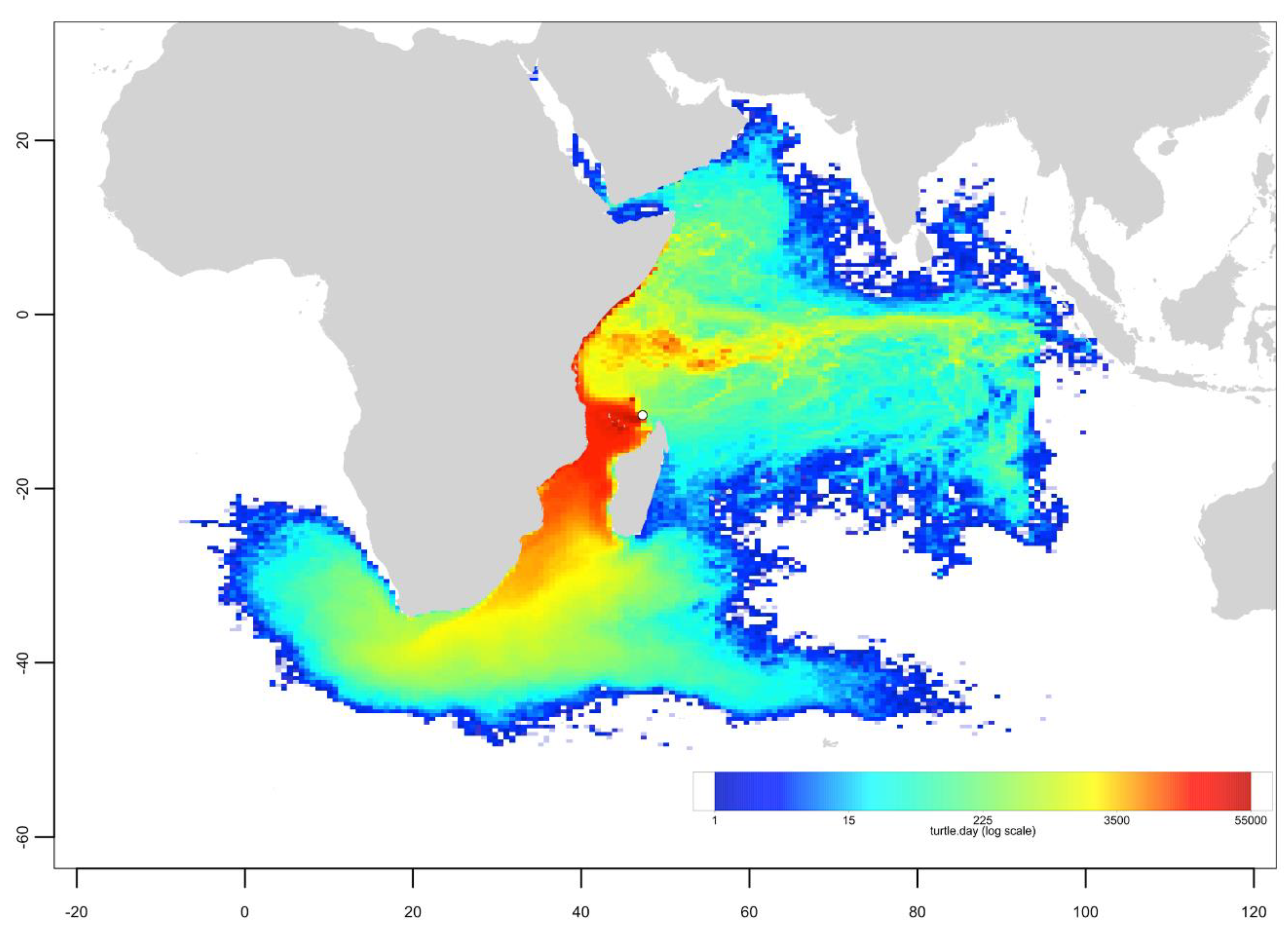
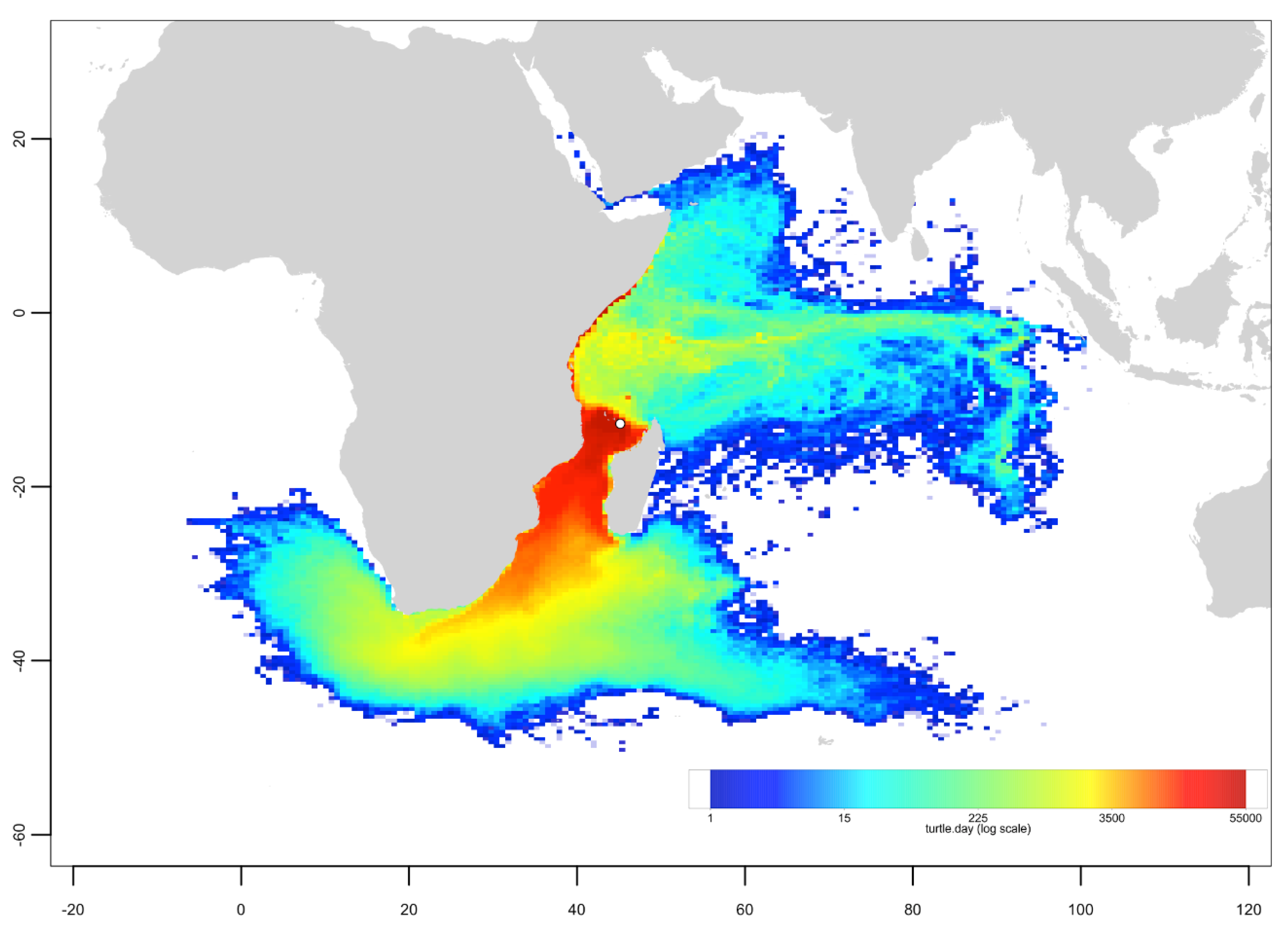


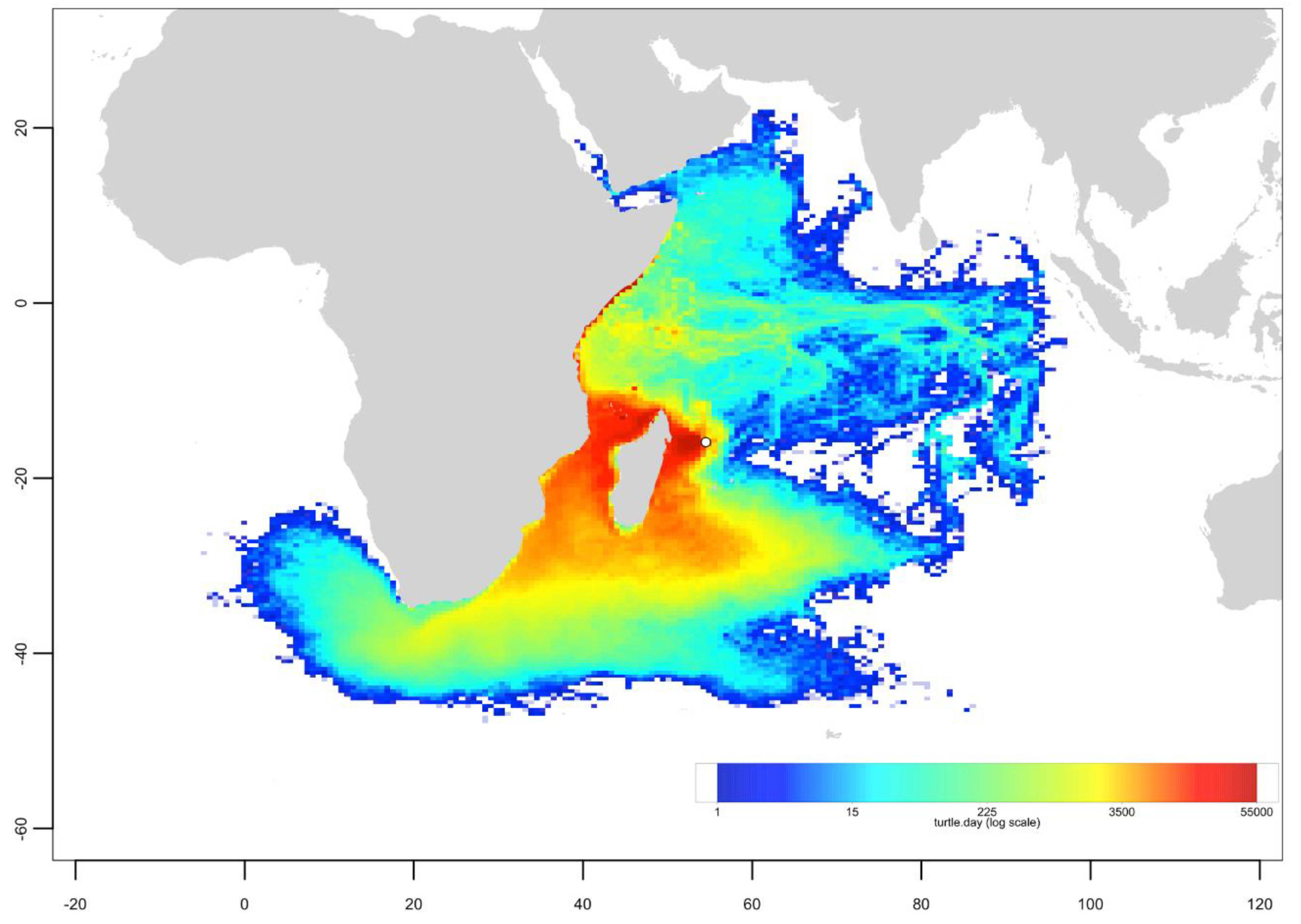
Appendix C. Mixed Stock Analysis Simulations
References
- Treml, E.A.; Halpin, P.N.; Urban, D.L.; Pratson, L.F. Modeling population connectivity by ocean currents, a graph-theoretic approach for marine conservation. Landsc. Ecol. 2007, 23, 19–36. [Google Scholar] [CrossRef]
- Hays, G.C. Ocean currents and marine life. Curr. Biol. 2017, 27, R470–R473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Block, B.A.; Jonsen, I.D.; Jorgensen, S.J.; Winship, A.J.; Shaffer, S.A.; Bograd, S.J.; Hazen, E.L.; Foley, D.G.; Breed, G.A.; Harrison, A.L.; et al. Tracking apex marine predator movements in a dynamic ocean. Nature 2011, 475, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Hart, K.M.; Hyrenbach, K.D. Satellite telemetry of marine megavertebrates: The coming of age of an experimental science. Endanger. Species Res. 2010, 10, 9–20. [Google Scholar] [CrossRef]
- Lalire, M.; Gaspar, P. Modeling the active dispersal of juvenile leatherback turtles in the North Atlantic Ocean. Mov. Ecol. 2019, 7, 1–17. [Google Scholar] [CrossRef]
- Mayorga-Adame, C.G.; Batchelder, H.P.; Spitz, Y.H. Modeling larval connectivity of coral reef organisms in the Kenya-Tanzania region. Front. Mar. Sci. 2017, 4, 92. [Google Scholar] [CrossRef] [Green Version]
- Caldwell, I.R.; Gergel, S.E. Thresholds in seascape connectivity: Influence of mobility, habitat distribution, and current strength on fish movement. Landsc. Ecol. 2013, 28, 1937–1948. [Google Scholar] [CrossRef]
- Hogan, J.D.; Thiessen, R.J.; Sale, P.F.; Heath, D.D. Local retention, dispersal and fluctuating connectivity among populations of a coral reef fish. Oecologia 2012, 168, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Vaudo, J.J.; Byrne, M.E.; Wetherbee, B.M.; Harvey, G.M.; Shivji, M.S. Long-term satellite tracking reveals region-specific movements of a large pelagic predator, the shortfin mako shark, in the western North Atlantic Ocean. J. Appl. Ecol. 2017, 54, 1765–1775. [Google Scholar] [CrossRef]
- Trudelle, L.; Cerchio, S.; Zerbini, A.N.; Geyer, Y.; Mayer, F.X.; Jung, J.L.; Hervé, M.R.; Pous, S.; Pous, S.; Rosenbaum, H.C.; et al. Influence of environmental parameters on movements and habitat utilization of humpback whales (Megaptera novaeangliae) in the Madagascar breeding ground. R. Soc. Open Sci. 2016, 3, 160616. [Google Scholar] [CrossRef] [Green Version]
- Shillinger, G.L.; Palacios, D.M.; Bailey, H.; Bograd, S.J.; Swithenbank, A.M.; Gaspar, P.; Wallace, B.P.; Spotila, J.R.; Paladino, F.V.; Piedra, R.; et al. Persistent leatherback turtle migrations present opportunities for conservation. PLoS Biol. 2008, 6, e171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musick, J.A.; Limpus, C.J. Habitat utilization and migration in juvenile sea turtles. In The Biology of Sea Turtles; Lutz, P.L., Musick, J.A., Eds.; CRC Press: Boca Raton, FL, USA, 1997; Volume 1, pp. 137–163. ISBN 0849384222. [Google Scholar]
- Limpus, C.J.; Couper, P.J.; Read, M.A. The green turtle, Chelonia mydas, in Queensland: Population structure in a warm temperate feeding area. Mem. Queensl. Mus. 1994, 35, 139–154. [Google Scholar]
- Jensen, M.P.; Bell, I.P.; Limpus, C.J.; Hamann, M.; Ambar, S.; Whap, T.; David, C.; FitzSimmons, N.N. Spatial and temporal genetic variation among size classes of green turtles (Chelonia mydas) provides information on oceanic dispersal and population dynamics. Mar. Ecol. Prog. Ser. 2016, 543, 241–256. [Google Scholar] [CrossRef] [Green Version]
- Jensen, M.P.; FitzSimmons, N.N.; Dutton, P.H. Molecular genetics of sea turtles. In The Biology of Sea Turtles; Wyneken, J., Lohmann, K.J., Musick, J.A., Eds.; CRC Press: Boca Raton, FL, USA, 2013; Volume 3, pp. 135–154. [Google Scholar]
- Hays, G.C.; Mortimer, J.A.; Ierodiaconou, D.; Esteban, N. Use of Long-Distance Migration Patterns of an Endangered Species to Inform Conservation Planning for the World’s Largest Marine Protected Area. Conserv. Biol. 2014, 28, 1636–1644. [Google Scholar] [CrossRef] [Green Version]
- Dethmers, K.E.M.E.M.; Jensen, M.P.; FitzSimmons, N.N.; Broderick, D.; Limpus, C.J.; Moritz, C. Migration of green turtles (Chelonia mydas) from Australasian feeding grounds inferred from genetic analyses. Mar. Freshw. Res. 2010, 61, 1376–1387. [Google Scholar] [CrossRef]
- Dalleau, M. Ecologie Spatiale des Tortues Marines Dans le Sud-Ouest de L’océan Indien: Apport de la Géomatique et de la Modélisation Pour la Conservation. Ph.D. Thesis, Université de la Réunion, La Réunion, France, 2013. (In French). [Google Scholar]
- Hays, G.C.; Hawkes, L.A. Satellite Tracking Sea Turtles: Opportunities and Challenges to Address Key Questions. Front. Mar. Sci. 2018, 5, 1–12. [Google Scholar] [CrossRef]
- Mansfield, K.L.; Wyneken, J.; Porter, W.P.; Luo, J. First satellite tracks of neonate sea turtles redefine the “lost years” oceanic niche. Proc. R. Soc. B 2014, 281, 20133039. [Google Scholar] [CrossRef] [Green Version]
- Bowen, B.W.; Karl, S.A. Population genetics and phylogeography of sea turtles. Mol. Ecol. 2007, 16, 4886–4907. [Google Scholar] [CrossRef]
- Bowen, B.W.; Bass, A.L.; Chow, S.-M.; Bostrom, M.; Bjorndal, K.A.; Bolten, A.B.; Okayama, T.; Bolker, B.; Epperly, S.P.; LaCasella, E.L.; et al. Natal homing in juvenile loggerhead turtles (Caretta caretta). Mol. Ecol. 2004, 13, 3797–3808. [Google Scholar] [CrossRef]
- Read, T.C.; FitzSimmons, N.N.; Wantiez, L.; Jensen, M.P.; Keller, F.; Chateau, O.; Farman, R.; Werry, J.; Mackay, K.T.; Petro, G.; et al. Mixed stock analysis of a resident green turtle, Chelonia mydas, population in New Caledonia links rookeries in the South Pacific. Wildl. Res. 2015, 42, 488–499. [Google Scholar] [CrossRef]
- Pella, J.; Masuda, M. Bayesian methods for analysis of stock mixtures from genetic characters. Fish. Bull. 2001, 99, 151–167. [Google Scholar]
- Bolker, B.; Okayama, T.; Bjorndal, K.A.; Bolten, A.B. Incorporating multiple mixed stocks in mixed stock analysis: “many-to-many” analyses. Mol. Ecol. 2007, 16, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Hays, G.C.; Marsh, R. Estimating the age of juvenile loggerhead sea turtles in the North Atlantic. Can. J. Zool. 1997, 75, 40–46. [Google Scholar] [CrossRef]
- Gaspar, P.; Benson, S.R.; Dutton, P.H.; Réveillère, A.; Jacob, G.; Meetoo, C.; Dehecq, A.; Fossette, S. Oceanic dispersal of juvenile leatherback turtles: Going beyond passive drift modeling. Mar. Ecol. Prog. Ser. 2012, 457, 265–284. [Google Scholar] [CrossRef]
- Scott, R.; Biastoch, A.; Agamboue, P.D.; Bayer, T.; Boussamba, F.L.; Formia, A.; Godley, B.J.; Mabert, B.D.K.; Manfoumbi, J.C.; Schwarzkopf, F.U.; et al. Spatio-temporal variation in ocean current-driven hatchling dispersion: Implications for the world’s largest leatherback sea turtle nesting region. Divers. Distrib. 2017, 23, 604–614. [Google Scholar] [CrossRef]
- Putman, N.F.; Seney, E.E.; Verley, P.; Shaver, D.J.; López-Castro, M.C.; Cook, M.; Guzmán, V.; Brost, B.; Ceriani, S.A.; de Mirón, R.J.G.D.; et al. Predicted distributions and abundances of the sea turtle ‘lost years’ in the western North Atlantic Ocean. Ecography 2019, 04929. [Google Scholar] [CrossRef] [Green Version]
- Abecassis, M.; Senina, I.; Lehodey, P.; Gaspar, P.; Parker, D.; Balazs, G.; Polovina, J. A Model of Loggerhead Sea Turtle (Caretta caretta) Habitat and Movement in the Oceanic North Pacific. PLoS ONE 2013, 8, e73274. [Google Scholar] [CrossRef]
- Gaspar, P.; Lalire, M. A model for simulating the active dispersal of juvenile sea turtles with a case study on western Pacific leatherback turtles. PLoS ONE 2017, 12, e0181595. [Google Scholar] [CrossRef] [Green Version]
- Naro-maciel, E.; Gaughran, S.J.; Putman, N.F.; Amato, G.; Arengo, F.; Dutton, P.H.; Mcfadden, K.W.; Vintinner, E.C.; Sterling, E.J. Predicting connectivity of green turtles at Palmyra Atoll, central Pacific: A focus on mtDNA and dispersal modelling. J. R. Soc. Interface 2014, 11, 20130888. [Google Scholar] [CrossRef] [Green Version]
- Putman, N.F.; Naro-maciel, E. Finding the “lost years” in green turtles: Insights from ocean circulation models and genetic analysis. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131468. [Google Scholar] [CrossRef]
- Blumenthal, J.M.; Abreu-Grobois, F.A.; Austin, T.J.; Broderick, A.C.; Bruford, M.W.; Coyne, M.S.; Ebanks-Petrie, G.; Formia, A.; Meylan, P.A.; Meylan, A.B.; et al. Turtle groups or turtle soup: Dispersal patterns of hawksbill turtles in the Caribbean. Mol. Ecol. 2009, 18, 4841–4853. [Google Scholar] [CrossRef] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Bourjea, J. Sea turtles; a review of status, distribution and interaction with fisheries in the Southwest Indian Ocean. In Offshore Fisheries of the Southwest Indian Ocean: Their Status and the Impact on Vulnerable Species; Van der Elst, R., Everett, B., Eds.; Oceanographic Research Institute: Durban, South Africa, 2015; pp. 325–349. [Google Scholar]
- Mortimer, J.A.; Esteban, N.; Guzman, A.N.; Hays, G.C. Estimates of marine turtle nesting populations in the south-west Indian Ocean indicate the importance of the Chagos Archipelago. Oryx 2020, in press. [Google Scholar] [CrossRef] [Green Version]
- Dalleau, M.; Ciccione, S.; Mortimer, J.A.; Garnier, J.; Benhamou, S.; Bourjea, J. Nesting Phenology of Marine Turtles: Insights from a Regional Comparative Analysis on Green Turtle (Chelonia mydas). PLoS ONE 2012, 7, e46920. [Google Scholar] [CrossRef] [Green Version]
- Bourjea, J.; Mortimer, J.A.; Garnier, J.; Okemwa, G.; Godley, B.; Hughes, G.; Dalleau, M.; Jean, C.; Ciccione, S.; Muths, D. Population structure enhances perspectives on regional management of the western Indian Ocean green turtle. Conserv. Genet. 2015, 16, 1069–1083. [Google Scholar] [CrossRef]
- Reyne, M.; Webster, I.; Huggins, A. A Preliminary Study on the Sea Turtle Density in Mauritius. Mar. Turt. Newsl. 2017, 152, 5–8. [Google Scholar]
- Jean, C.; Ciccione, S.; Ballorain, K.; Georges, J.Y.; Bourjea, J. Ultralight aircraft surveys reveal marine turtle population increases along the west coast of Reunion Island. Oryx 2010, 44, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Mortimer, J.A. Turtle & Tortoise Conservation Project J1 Environmental Management Plan of the Seychelles; Final report to the Ministry of Environment Republic of Seychelles and the Global Environment Facility, Seychelles; Ministry of Environment Republic of Seychelles and the Global Environment Facility: Victoria, Seychelles, 1998; Volume 1. [Google Scholar]
- Limpus, C.J.; Reed, P.C. The green turtle, Chelonia mydas, in Queensland: A preliminary description of the population structure in a coral reef feeding ground. In Biology of Australasian Frogs and Reptiles; Grigg, G., Shine, R., Ehmann, H., Eds.; Surrey Beatty & Sons: Sydney, Australia, 1985; pp. 47–52. [Google Scholar]
- Dutton, P.H. Methods for collection and preservation of samples for sea turtle genetic studies. In Proceedings of the International Symposium on Sea Turtle Conservation Genetics, NOAA Technical Memorandum NMFS-SEFSC-396, Miami, FL, USA, 12–14 September 1995; pp. 17–24. [Google Scholar]
- Taquet, C. Diversité et Différenciation Génétiques des Populations de Tortues Vertes (Chelonia Mydas) Dans les Sites de Ponte et D‘Alimentation du Sud-Ouest de L‘océan Indien: Application Aux Stratégies de Conservation de L‘espèce. Ph.D. Thesis, Université de la Réunion, La Réunion, France, 2007. (In French). [Google Scholar]
- Bourjea, J.; Lapègue, S.; Gagnevin, L.; Broderick, D.; Mortimer, J.A.; Ciccione, S.; Roos, D.; Taquet, C.; Grisel, H. Phylogeography of the green turtle, Chelonia mydas, in the Southwest Indian Ocean. Mol. Ecol. 2007, 16, 175–186. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Jensen, M.P.; FitzSimmons, N.N.; Bourjea, J.; Hamabata, T.; Reece, J.S.; Dutton, P.H. The evolutionary history and global phylogeography of the green turtle (Chelonia mydas). J. Biogeogr. 2019, 46, 860–870. [Google Scholar] [CrossRef]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987. [Google Scholar]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP v6: DNA Sequence Polymorphism Analysis of Large Datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Tempelton, A.R.; Crandall, K.A.; Sing, C.F. A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping and DNA sequence data. III. Cladogram estimation. Genetics 1992, 132, 619. [Google Scholar]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiting, S.; Macrae, I.; Thorn, R.; Murray, W.; Whiting, A. Sea turtles of the Cocos (Keeling) Islands, Indian Ocean. Raffles Bull. Zool. 2014, 30, 168–183. [Google Scholar]
- Schott, F.A.; McCreary, J.P. The monsoon circulation of the Indian Ocean. Prog. Oceanogr. 2001, 51, 1–123. [Google Scholar] [CrossRef]
- Mendonça, M.T. Movements and Feeding Ecology of Immature Green Turtles (Chelonia mydas) in a Florida Lagoon. Copeia1 1983, 4, 1013–1023. [Google Scholar] [CrossRef] [Green Version]
- Southwood, A.L.; Andrews, R.D.; Paladino, F.V.; Jones, D.R. Effects of diving and swimming behavior on body temperatures of pacific leatherback turtles in tropical seas. Physiol. Biochem. Zool. 2005, 78, 285–297. [Google Scholar] [CrossRef] [Green Version]
- Piton, B.; Magnier, Y. Les conditions favorables à la présence de thons de surface dans les parages de Madagascar. ORSTOM 1976, 47, 293–309. [Google Scholar]
- Schouten, M.W.; De Ruijter, W.P.M.; Van Leeuwen, P.J.; Ridderinkhof, H. Eddies and variability in the Mozambique Channel. Deep Sea Res. Part II Top. Stud. Oceanogr. 2003, 50, 1987–2003. [Google Scholar] [CrossRef]
- von Brandis, R.G.; Mortimer, J.A.; Van De Geer, C.; Lea, J.S.E. A Long Migratory Record for a Small Post-Pelagic Hawksbill. Mar. Turt. Newsl. 2017, 152, 13–15. [Google Scholar]
- Wallace, B.P.; DiMatteo, A.D.; Hurley, B.J.; Finkbeiner, E.M.; Bolten, A.B.; Chaloupka, M.Y.; Hutchinson, B.J.; Abreu-Grobois, F.A.; Amorocho, D.F.; Bjorndal, K.A.; et al. Regional management units for marine turtles: A novel framework for prioritizing conservation and research across multiple scales. PLoS ONE 2010, 5, e15465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteban, N.; Unsworth, R.K.F.; Gourlay, J.B.Q.; Hays, G.C. The discovery of deep-water seagrass meadows in a pristine Indian Ocean wilderness revealed by tracking green turtles. Mar. Pollut. Bull. 2018, 134, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Bourjea, J.; Dalleau, M.; Derville, S.; Beudard, F.; Marmoex, C.; M’Soili, A.; Roos, D.; Ciccione, S.; Frazier, J. Seasonality, abundance, and fifteen-year trend in green turtle nesting activity at Itsamia, Mohéli, Comoros. Endanger. Species Res. 2015, 27, 265–276. [Google Scholar] [CrossRef] [Green Version]
- Lutjeharms, J.R.E. The coastal oceans of south-eastern Africa. In The Sea; Robinson, R.A., Brink, K., Eds.; Chicago University Press: Chicago, IL, USA, 2005; pp. 781–832. [Google Scholar]
- Green, E.; Short, F.; Frederick, T. World Atlas of Seagrasses; University of California Press: California, CA, USA, 2003. [Google Scholar]
- Hughes, G.R. The Sea Turtles of South East Africa; University of Natal: Durban, South Africa, 1974. [Google Scholar]
- Frazier, J.G. Marine turtles of the Western Indian Ocean. Oryx 1975, 13, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Bourjea, J.; Ciccicone, S.; Lauret-Stepler, M.; Marmoex, C.; Jean, C. Les îles Éparses: 25 ans de recherche sur les tortues marines. Bull. Soc. Herp. Fr. 2011, 139–140, 95–111. [Google Scholar]
- Bourjea, J.; Dalleau, M.; Ciccione, S. Seasonal variability of migrating corridors and foraging areas of adult green turtles revealed by satellite tracking at the regional scale. In Proceedings of the 33rd Annual Symposium on Sea Turtle Biology and Conservation, Baltimore, Maryland, 2–8 February 2013. [Google Scholar]
- de Ruijter, W.P.M.; Ridderinkhof, H.; Lutjeharms, J.R.E.; Schouten, M.W.; Veth, C. Observations of the flow in the Mozambique Channel. Geophys. Res. Lett. 2002, 29, 140–141. [Google Scholar] [CrossRef]
- Avise, J.C. Phylogeography: The History and Formation of Species; Harvard University Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Lutjeharms, J.R.E. Three decades of research on the greater Agulhas Current. Ocean. Sci. 2007, 3, 129–147. [Google Scholar] [CrossRef] [Green Version]
- Dalleau, M.; Kramer-Schadt, S.; Gangat, Y.; Bourjea, J.; Lajoie, G.; Grimm, V. Modeling the emergence of migratory corridors and foraging hot spots of the green sea turtle. Ecol. Evol. 2019, 9, 10317–10342. [Google Scholar] [CrossRef]
- Dubois L’escadre de Perse. Sous le signe de la tortue. Voyages anciens a‘ l’île Bourbon (1611–1725); Lougnon, A., Ed.; Library Ge´rard: Saint Denis, France, 1669; pp. 75–93. [Google Scholar]
- Ciccione, S.; Bourjea, J. Nesting of green turtles in Saint Leu, Réunion Island. Mar. Turt. Newsl. 2006, 112, 1–3. [Google Scholar]
- Dalleau, M.; Benhamou, S.; Sudre, J.; Ciccione, S.; Bourjea, J. The spatial ecology of juvenile loggerhead turtles (caretta caretta) in the Indian Ocean sheds light on the lost years mystery. Mar. Biol. 2014, 161, 1835–1849. [Google Scholar] [CrossRef] [Green Version]
- Mortimer, J.A.; Von Brandis, R.G.; Liljevik, A.; Chapman, R.; Collie, J. Fall and rise of nesting green turtles (Chelonia mydas) at Aldabra Atoll, Seychelles: Positive response to four decades of protection (1968–2008). Chelonian Conserv. Biol. 2011, 10, 165–176. [Google Scholar] [CrossRef]
- Fretey, J.; Frétey, T.; Dupré, A.; Dupré, J.; André, A.A.; Meunier, A. Is the hawksbill turtle, Eretmochelys imbricata, a regular nester on Rodrigues Island (Indian Ocean)? Mar. Turt. Newsl. 2013, 138, 22–24. [Google Scholar]
- Schott, F.A.; Xie, S.P.; McCreary, J.P. Indian ocean circulation and climate variability. Rev. Geophys. 2009, 47, 1–46. [Google Scholar] [CrossRef]
- Appleyard, S.A.; Ward, R.D.; Grewe, P.M. Genetic stock structure of bigeye tuna in the Indian Ocean using mitochondrial DNA and microsatellites. J. Fish Biol. 2002, 60, 767–770. [Google Scholar] [CrossRef]
- Muths, D.; Le Couls, S.; Evano, H.; Grewe, P.; Bourjea, J. Multi-Genetic Marker Approach and Spatio-Temporal Analysis Suggest There Is a Single Panmictic Population of Swordfish Xiphias gladius in the Indian Ocean. PLoS ONE 2013, 8, e63558. [Google Scholar] [CrossRef] [PubMed]
- Obura, D.O.; Bandeira, S.O.; Bodin, N.; Burgener, V.; Braulik, G.; Chassot, E.; Gullström, M.; Kochzius, M.; Nicoll, M.; Osuka, K.; et al. The Northern Mozambique Channel. In World Seas: An Environmental Evaluation; Elsevier: Amsterdam, The Netherlands, 2019; pp. 75–99. [Google Scholar]
- Gamoyo, M.; Obura, D.; Reason, C.J.C. Estimating Connectivity Through Larval Dispersal in the Western Indian Ocean. J. Geophys. Res. Biogeosci. 2019, 124, 2446–2459. [Google Scholar] [CrossRef]
- Obura, D. The Diversity and Biogeography of Western Indian Ocean Reef-Building Corals. PLoS ONE 2012, 7, e45013. [Google Scholar] [CrossRef]
- Jahnke, M.; Gullström, M.; Larsson, J.; Asplund, M.E.; Mgeleka, S.; Silas, M.O.; Hoamby, A.; Mahafina, J.; Nordlund, L.M. Population genetic structure and connectivity of the seagrass Thalassia hemprichii in the Western Indian Ocean is influenced by predominant ocean currents. Ecol. Evol. 2019, 9, 8953–8964. [Google Scholar] [CrossRef] [Green Version]
- Visram, S.; Yang, M.C.; Pillay, R.M.; Said, S.; Henriksson, O.; Grahn, M.; Chen, C.A. Genetic connectivity and historical demography of the blue barred parrotfish (Scarus ghobban) in the western Indian Ocean. Mar. Biol. 2010, 157, 1475–1487. [Google Scholar] [CrossRef]
- Muths, D.; Tessier, E.; Gouws, G.; Craig, M.; Mwale, M.; Mwaluma, J.; Mwandya, A.; Bourjea, J. Restricted dispersal of the reef fish Myripristis berndti at the scale of the South-West Indian Ocean compared to known transoceanic species connectivity. Mar. Ecol. Prog. Ser. 2011, 443, 167–180. [Google Scholar] [CrossRef] [Green Version]
- Muths, D.; Tessier, E.; Bourjea, J. Genetic structure of the reef grouper Epinephelus merra in the West Indian Ocean appears congruent with biogeographic and oceanographic boundaries. Mar. Ecol. 2015, 36, 447–461. [Google Scholar] [CrossRef]
- Shamblin, B.M.; Bolten, A.B.; Bjorndal, K.A.; Dutton, P.H.; Nielsen, J.T.; Abreu-Grobois, F.A.; Reich, K.J.; Witherington, B.E.; Bagley, D.A.; Ehrhart, L.M.; et al. Expanded mitochondrial control region sequences increase resolution of stock structure among North Atlantic loggerhead turtle rookeries. Mar. Ecol. Prog. Ser. 2012, 469, 145–160. [Google Scholar] [CrossRef] [Green Version]
- Jensen, M.P.; Pilcher, N.J.; FitzSimmons, N.N. Genetic markers provide insight on origins of immature green turtles (Chelonia mydas) with biased sex ratios at two foraging grounds in Sabah, Malaysia. Endanger. Species Res. 2016, 31, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Shamblin, B.M.; Bjorndal, K.A.; Bolten, A.B.; Hillis-Starr, Z.M.; Lundgren, I.; Naro-maciel, E.; Nairn, C.J. Mitogenomic sequences better resolve stock structure of southern Greater Caribbean green turtle rookeries. Mol. Ecol. 2012, 21, 2330–2340. [Google Scholar] [CrossRef]
- Tikochinski, Y.; Bendelac, R.; Barash, A.; Daya, A.; Levy, Y.; Friedmann, A. Mitochondrial DNA STR analysis as a tool for studying the green sea turtle (Chelonia mydas) populations: The Mediterranean Sea case study. Mar. Genomics 2012, 6, 17–24. [Google Scholar] [CrossRef]
- Komoroske, L.M.; Jensen, M.P.; Stewart, K.R.K.R.; Shamblin, B.M.; Dutton, P.H. Advances in the Application of Genetics in Marine Turtle Biology and Conservation. Front. Mar. Sci. 2017, 4, 156. [Google Scholar] [CrossRef] [Green Version]
- Halo, I.; Backeberg, B.; Penven, P.; Ansorge, I.; Reason, C.; Ullgren, J.E. Eddy properties in the Mozambique Channel: A comparison between observations and two numerical ocean circulation models. Deep Sea Res. Part II Top. Stud. Oceanogr. 2014, 100, 38–53. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Haplotype ID | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Location | Coordinates | π | Hd | n | CM-A08 | CmP49 | CmP87 | CmP40 | CmP75 | CmP57 | CmP47 | CmP32* | CmP83* | CM-A46* | CmI11 | CmI9 | CmP176 |
| Kenya | 3°30’S–40°11’E | 0.0223 | 0.4966 | 67 | 6 | 47 | 1 | 1 | 2 | 1 | 6 | 2 | - | - | - | 1 | - |
| Aldabra | 9°41’S–46°21’E | 0.0154 | 0.3333 | 47 | - | 38 | - | - | 1 | - | 5 | - | - | - | - | 1 | 2 |
| Glorieuses | 11°58’S–47°29E | 0.0328 | 0.6874 | 31 | 3 | 17 | 1 | - | - | 1 | 7 | 1 | - | - | - | 1 | - |
| Mayotte | 12°98’S–45°19’E | 0.0241 | 0.4778 | 53 | 6 | 37 | - | - | 3 | - | 4 | 3 | - | - | - | - | - |
| west Madagascar | 18°25’S–43°55’E | 0.0285 | 0.5585 | 33 | 9 | 19 | - | - | 2 | - | 1 | 1 | - | - | 1 | - | - |
| Juan de Nova | 17°05’S–47°72’E | 0.0339 | 0.6724 | 24 | 12 | 8 | - | - | 2 | - | - | 1 | 1 | - | - | - | - |
| Europa | 22°34’S–40°37’E | 0.0326 | 0.5946 | 38 | 22 | 10 | 1 | - | 1 | - | 3 | - | - | 1 | - | - | - |
| La Réunion | 21°05’S–55°22’E | 0.0280 | 0.5005 | 65 | 6 | 44 | - | - | - | - | 13 | 1 | - | - | - | - | 1 |
| Total | 358 | 64 | 220 | 3 | 1 | 11 | 2 | 39 | 9 | 1 | 1 | 1 | 3 | 3 | |||
| North | Central | South | CKI | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Location | n | # orphan | % orphan | mean | 95% CI | mean | 95% CI | mean | 95% CI | mean | 95% CI |
| Kenya | 67 | 1(1) | 1% | 3% | (0–12) | 85% | (70–95) | 11% | (2–22) | 1% | (0–5) |
| Aldabra | 47 | 2(3) | 6% | 4% | (0–19) | 92% | (73–99) | 3% | (0–11) | 1% | (0–6) |
| Glorieuses | 31 | 1(1) | 3% | 6% | (0–28) | 79% | (45–96) | 14% | (1–31) | 2% | (0–10) |
| Mayotte | 53 | 0(0) | 0% | 4% | (0–16) | 80% | (58–94) | 15% | (3–30) | 2% | (0–7) |
| west Madagascar | 33 | 1(1) | 3% | 4% | (0–19) | 56% | (28–79) | 39% | (18–65) | 1% | (0–7) |
| Juan de Nova | 24 | 0(0) | 0% | 4% | (0–16) | 24% | (2–52) | 71% | (43–94) | 1% | (0–16) |
| Europa | 38 | 0(0) | 0% | 5% | (0–17) | 20% | (1–42) | 74% | (53–91) | 1% | (0–6) |
| La Réunion | 65 | 1(1) | 2% | 4% | (0–19) | 85% | (63–96) | 10% | (1–21) | 1% | (0–9) |
| Nesting Area | Long (°E) | Lat (°S) | Release Period |
|---|---|---|---|
| Seychelles granitics Group | 55.6 | 4.46 | August 1st to October 31 |
| Seychelles Amirantes Group | 53.4 | 5.63 | August 1st to October 31 |
| Glorieuses Island | 47.3 | 11.57 | 1st June to 31 August |
| Mayotte Island | 45.15 | 12.13 | 1st July to 31 September |
| Mohéli Island | 43.73 | 12.32 | 1st June to 31 September |
| Tromelin Island | 54.52 | 5.89 | 1st September to 31 November |
| Europa Island | 40.36 | 22.37 | 1st January–31 Marsh |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jensen, M.P.; Dalleau, M.; Gaspar, P.; Lalire, M.; Jean, C.; Ciccione, S.; Mortimer, J.A.; Quillard, M.; Taquet, C.; Wamukota, A.; et al. Seascape Genetics and the Spatial Ecology of Juvenile Green Turtles. Genes 2020, 11, 278. https://doi.org/10.3390/genes11030278
Jensen MP, Dalleau M, Gaspar P, Lalire M, Jean C, Ciccione S, Mortimer JA, Quillard M, Taquet C, Wamukota A, et al. Seascape Genetics and the Spatial Ecology of Juvenile Green Turtles. Genes. 2020; 11(3):278. https://doi.org/10.3390/genes11030278
Chicago/Turabian StyleJensen, Michael P., Mayeul Dalleau, Philippe Gaspar, Maxime Lalire, Claire Jean, Stéphane Ciccione, Jeanne A. Mortimer, Mireille Quillard, Coralie Taquet, Andrew Wamukota, and et al. 2020. "Seascape Genetics and the Spatial Ecology of Juvenile Green Turtles" Genes 11, no. 3: 278. https://doi.org/10.3390/genes11030278
APA StyleJensen, M. P., Dalleau, M., Gaspar, P., Lalire, M., Jean, C., Ciccione, S., Mortimer, J. A., Quillard, M., Taquet, C., Wamukota, A., Leroux, G., & Bourjea, J. (2020). Seascape Genetics and the Spatial Ecology of Juvenile Green Turtles. Genes, 11(3), 278. https://doi.org/10.3390/genes11030278