Present and Future Salmonid Cytogenetics

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Studied Material
2.2. Chromosome Preparation and Staining
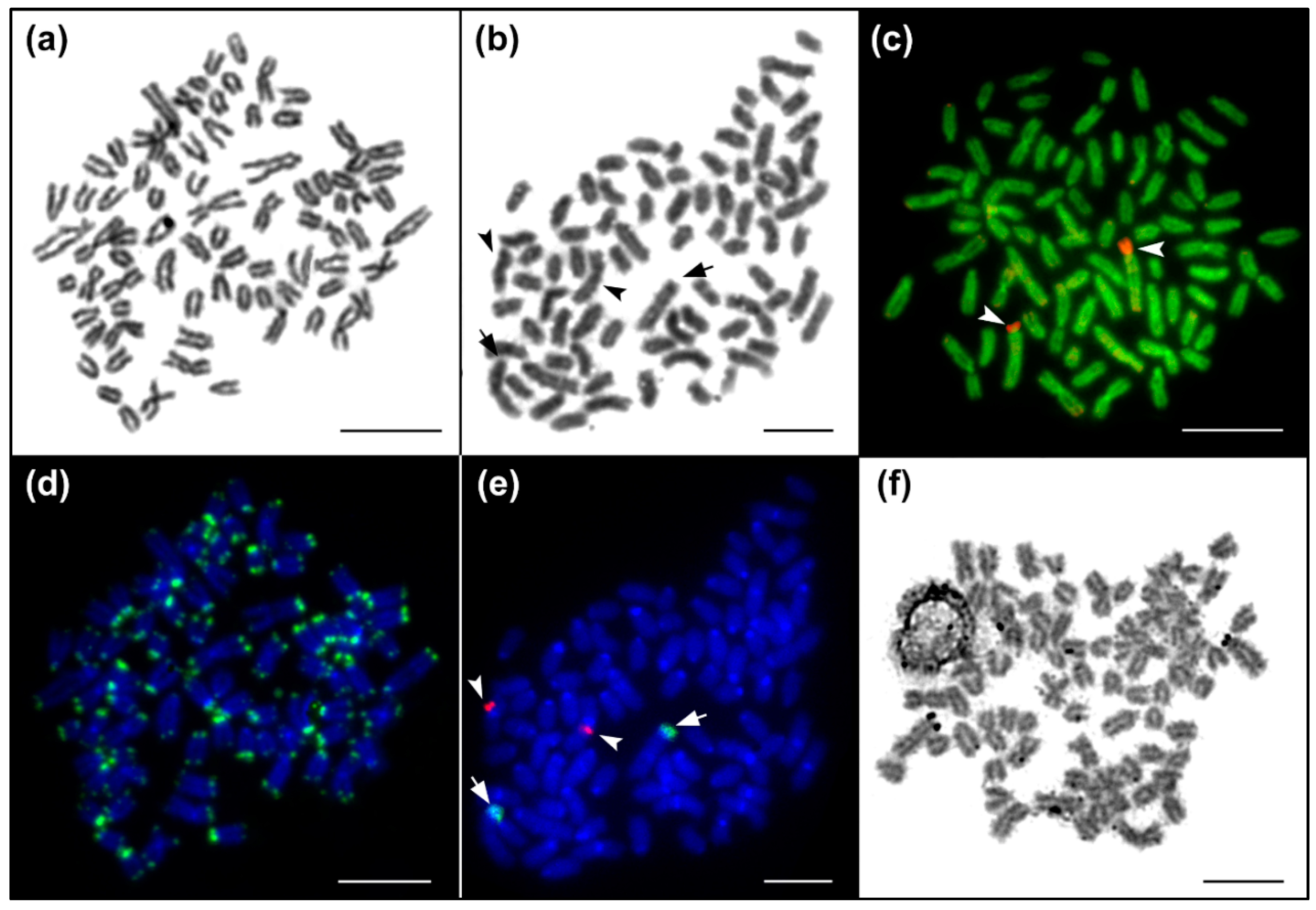
2.3. Fluorescence In Situ Hybridization (FISH) with Telomeric and rRNA Genes Probes
2.4. Microscopy and Image Analyses
2.5. Cytogenomic Analyses
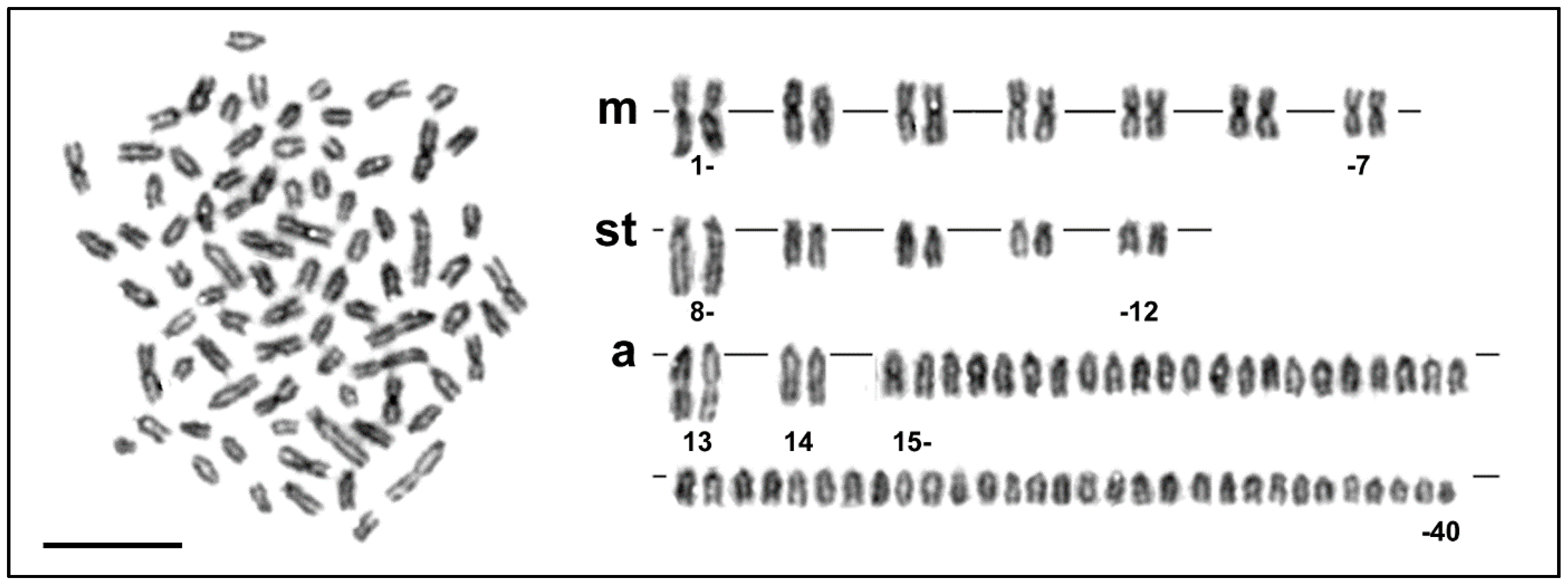
3. Results
3.1. Karyotypes and Molecular Cytogenetic Traits
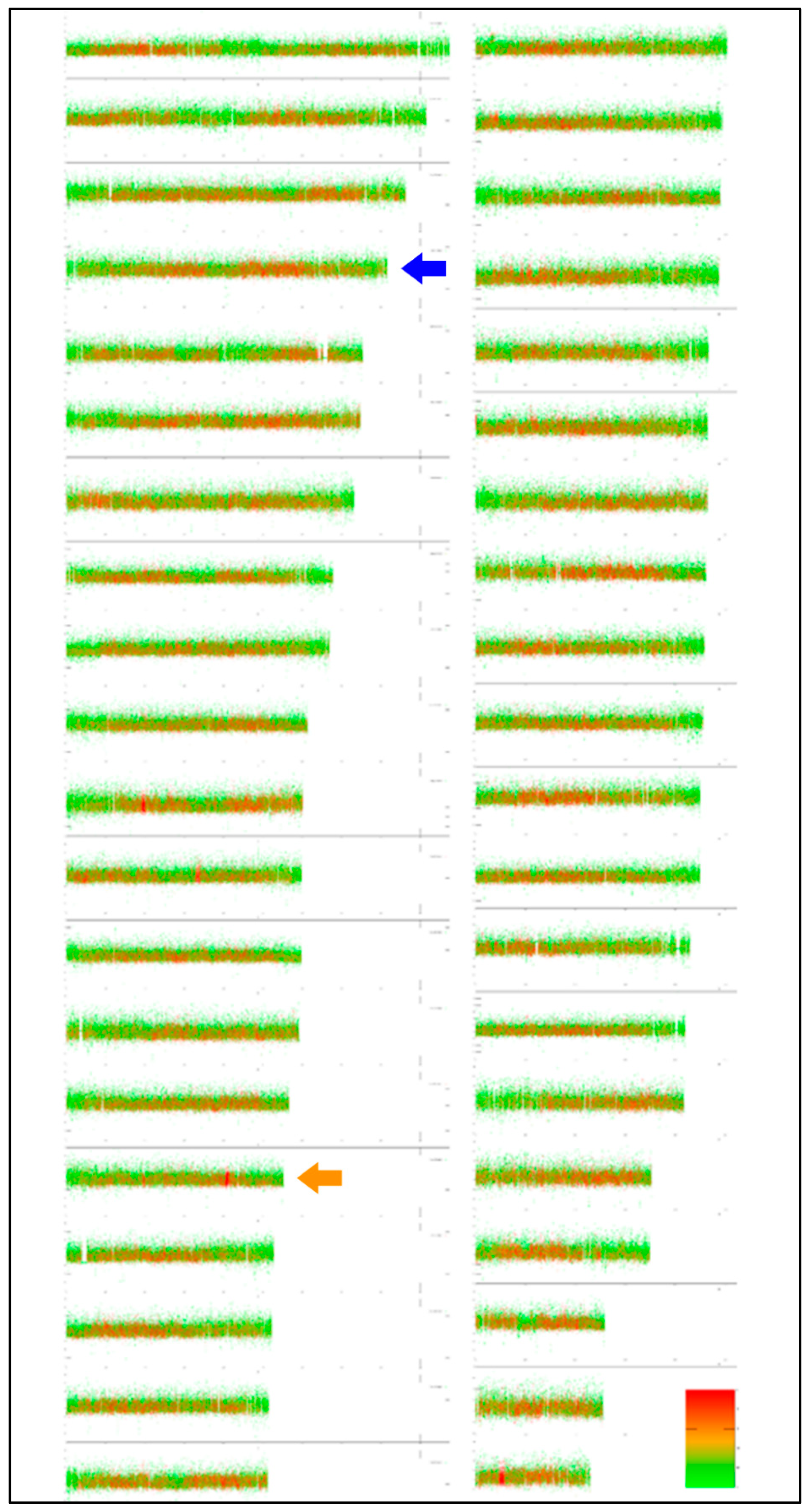
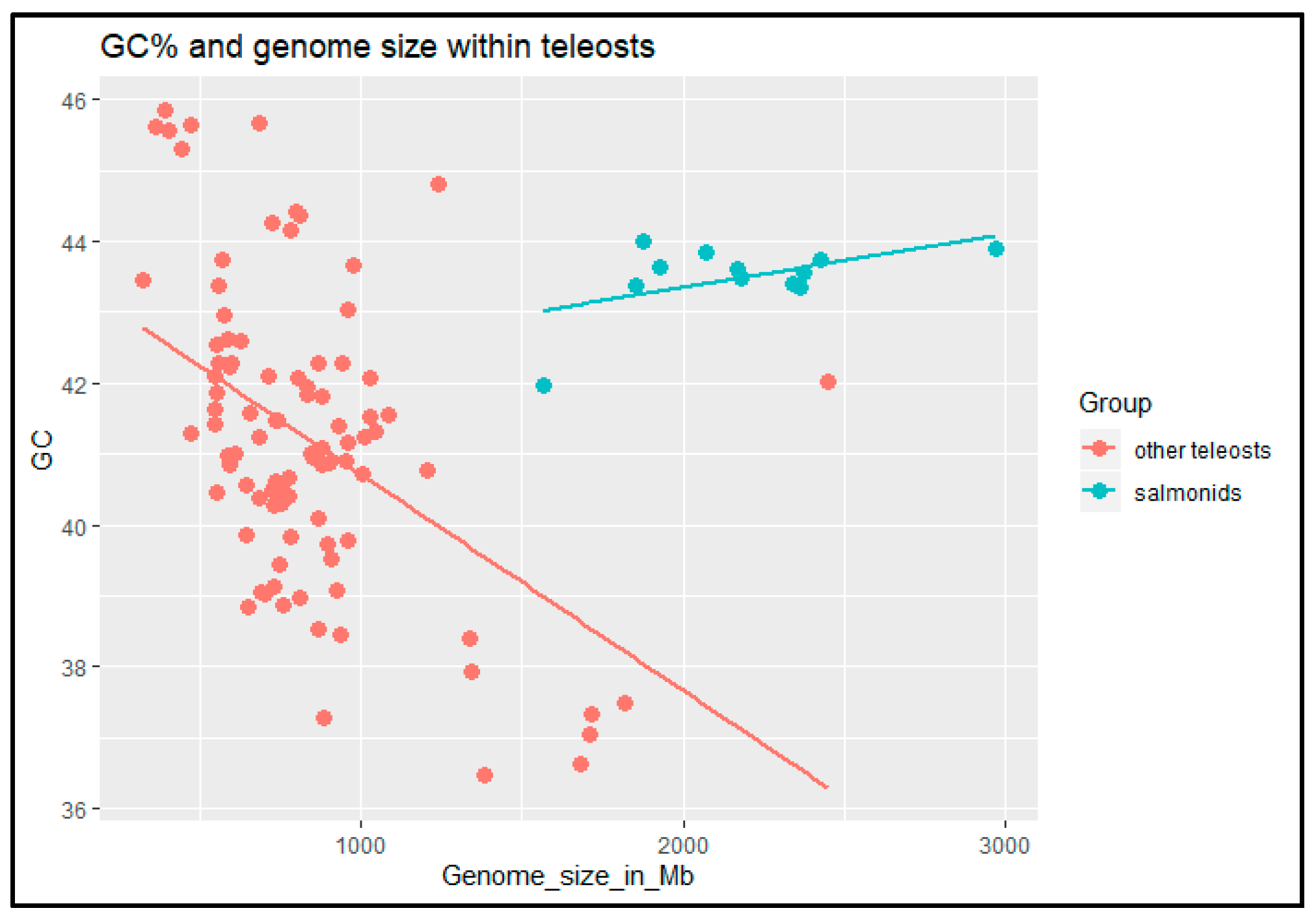
3.2. Virtual Karyotype and Cytogenomics in Salmonids
4. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/Form | Locality | Country | Examined Individuals | 2n | Karyotype Composition | NF | Ref. | Notes | |
|---|---|---|---|---|---|---|---|---|---|
| m/sm | st/a | ||||||||
| Aral Sea Basin | |||||||||
| S. oxianus | Kyzylsu R. (Amu Darya basin) | KAZ | 3 | 80 | 18 | 62 | 98 | [53] | 1; 2 |
| S. oxianus | Alamedin R. (Chu basin) | KG | 5 | 80 | 18 | 62 | 98 | [53] | 1; 2 |
| S. oxianus | Bech–Tach (Talas basin) | KG | 11 | 80 | 18 | 62 | 98 | [53] | 1; 2 |
| Balkans and Mediterranean Sea Basin | |||||||||
| S. carpio | Garda L. | IT | embryos | 80 | 20 | 60 | 100 | [54] | 1 |
| S. cenerinus | Monti Sibillini | IT | 57 † | 80 | 14/8 | 58 | 102 | [55] | 2; 5 |
| S. farioides | Drosopigi R. | GR | / | 80 | 20 | 60 | 100 | [56] | 1; 2 |
| S. lourosensis | Louros R. | GR | / | 80 | 20 | 60 | 100 | [56] | 1; 2 |
| S. letnica | Ochrid L. | MK | embryos | 80 | 104 | [57] | 1 | ||
| S. marmoratus | Socha R. | SI | / | 80 | 22 | 58 | 102 | [58] | 1 |
| S. marmoratus | Socha R. | SI | 1 | 80 | 22 | 58 | 102 | [59] | 1 |
| S. marmoratus | Idrijca R. | SI | 2 | 80 | 22 | 58 | 102 | [59] | 1 |
| S. marmoratus | Friuli–Venezia | IT | 57 † | 80 | 14/8 | 58 | 102 | [55] | 2; 5 |
| S. obtusirostris | Buna R. (Neretva R. basin) | BIH | / | 82 | 12 | 70 | 94 | [60] | 1 |
| S. peristericus | Aigos Germanos | GR | / | 80 | 20 | 60 | 100 | [56] | 1; 2 |
| S. trutta * | Buni, Krupica, Bistrica R. | RU | 17 | 80 | 18–20 | 62–60 | 100 | [61] | 1; 2 |
| S. trutta * | Klinje L. | BIH | 17 | 80 | 20 | 60 | 100 | [61] | 1; 2 |
| S. trutta * | Pschata R. | SI | / | 80 | 20 | 60 | 100 | [58] | 1 |
| S. trutta * | Tripotamos R. | GR | / | 76 | 16 | 60 | 92 | [56] | 1; 2 |
| Baltic Sea Basin | |||||||||
| S. trutta | Ropsha | RU | / | 78 | 20 | 58 | 98 | [62] | 3 |
| S. trutta (anadromous) | Vistula R. | PL | 23 | 80 | 14/6 | 60 | 100 | [63] | |
| S. trutta (anadromous) | Vistula R. | PL | 21 | 80 | 22 | 58 | 102 | [64] | |
| S. trutta | Vistula R. | PL | 18 | 80 | 22 | 58 | 102 | [65] | |
| S. trutta (lacustrine) | Wdzydze L. | PL | 13 | 80 | 22 | 58 | 102 | [65] | |
| S. trutta | Gawrych Ruda Hatchery | PL | 21 | 80 | 22 | 58 | 102 | [66] | |
| Black Sea Basin | |||||||||
| S. labrax | Local hatchery | GE | embryos | 80 | 18 | 62 | 98 | [67] | 1 |
| S. labrax | Local hatchery | GE | 6 | 80 | 22 | 58 | 102 | [68] | |
| S. trutta * | Black R. | GE | 8 | 80–82 | 20–22 | 60 | 100–104 | [68] | 3 |
| S. trutta * | Bzyb R. | GE | 9 | 82 | 22 | 60 | 104 | [68] | 1 |
| S. trutta * | Gumista R. | GE | 9 | 82 | 22 | 60 | 104 | [68] | 1 |
| S. trutta * | Kodori R. | GE | 8 | 80–82 | 20–22 | 60 | 100–104 | [68] | 1; 2 |
| S. trutta * | Bicaz, Prejmer, Azuga | RO | / | 80 | 24 | 56 | 104 | [69] | 1; 2 |
| S. trutta * | Western–Middle Carpathians | RO | / | 80 | 24 | 56 | 104 | [69] | 1; 2 |
| Caspian Sea Basin | |||||||||
| S. caspius | embryos | 80 | 18 | 62 | 98 | [70] | 1 | ||
| S. caspius | Kura R. | AZ | 2 | 82 | 20 | 62 | 102 | [68] | 1; 2 |
| S. ischchan “winter ischchan” | Sevan L. | AR | 11 | 80 | 16 | 64 | 96 | [67,71] | 1; 2 |
| S. ischchan “gegarkuni” | Sevan L. | AR | 17 | 80 | 18 | 62 | 98 | [67,71] | 1; 2 |
| S. ischchan “summer ischchan” | Sevan L. | AR | 23 | 82 | 18 | 64 | 100 | [67,71] | 1; 2 |
| S. ischchan “bodjak” | Sevan L. | AR | 7 | 82 | 16 | 66 | 98 | [67,71] | 1; 2 |
| S. trutta “alabalach” * | Argichi R. | AR | 8 | 80 | 16 | 64 | 96 | [72] | 1 |
| S. trutta | Marmarik R. | AR | / | 82 | 16 | 66 | 98 | [73] | 1 |
| S. trutta | Vedi R. | AR | / | 78 | 20 | 58 | 98 | [73] | 1 |
| S. trutta * | Azat R. | AR | 8 | 78 | 20 | 58 | 98 | [68] | 1 |
| S. trutta * | Arindg R. | 7 | 80 | 18 | 62 | 98 | [68] | 1 | |
| S. trutta * | Vedi R. | AR | 18 | 78 | 20 | 58 | 98 | [68] | 1 |
| S. trutta * | Korotan R. | 15 | 80 | 20 | 60 | 100 | [68] | 1 | |
| S. trutta * | Dzeoraget R. | AR | 8 | 80 | 20 | 60 | 100 | [68] | 1 |
| S. trutta * | Kcia R. | 3 | 82 | 20 | 62 | 102 | [68] | 1 | |
| S. trutta * | Kyuretchai R. | 9 | 84 | 16 | 68 | 100 | [68] | 1 | |
| S. trutta * | Marmarik R. | AR | 7 | 82 | 16 | 66 | 98 | [68] | 1 |
| S. trutta * | Ochtchi R. | 8 | 82 | 20 | 62 | 102 | [68] | 1 | |
| S. trutta * | Chatchen R. | 7 | 80 | 20 | 60 | 100 | [68] | 1 | |
| S. trutta * | Tchaki R. | 8 | 82 | 18 | 64 | 100 | [68] | 1 | |
| S. trutta * | Goygol L. | AZ | 7 | 80 | 20 | 60 | 100 | [68] | 1 |
| S. trutta * | Tabackuri L. | GE | 15 | 80 | 20 | 60 | 100 | [68] | 1 |
| Northern Sea Basin, European Atlantic coast | |||||||||
| S. trutta | Cares R. | ES | 49 | 80 | 22–23 | 57–58 | 102–103 | [74] | 1 |
| S. trutta | Pyrenees hatchery | ES | 44 | 81 | 22–24 | 57–59 | 103–105 | [74] | 1 |
| S. trutta (anadromous) | Galicia | ES | 14 | 80 | 20 | 60 | 100 | [75] | 4 |
| S. trutta (local hatchery strain) | Galicia | ES | 19 | 80 | 20 | 60 | 100 | [75] | 4 |
| S. trutta | Pšovka Cr. | CZ | 10 | 80 | 14/4 | 62 | 98 | [76] | 4 |
| S. trutta | Navia, Tambre, Umia, Mino R. | ES | 133 | 78–80 | 20 | 58–60 | 98–100 | [77,78] | 2; 4; 5 |
| S. trutta | Galicia | ES | 15 | 80 | 20 | 60 | 100 | [79] | 3 |
| S. trutta | Hatchery stock AT lineage | IT | 20 | 80 | 14/8 | 58 | 102 | [55] | 2; 5 |
| S. trutta | Loch Lomond | SCT | 6 | 79–80 | 21–22 | 58–59 | 100–102 | [80] | 3 |
| S. trutta | Norway (migratory) | NO | / | 80 | 14 | 66 | 94 | [81] | 3 |
| S. trutta | Germany | DE | 6 | 78–82 | 20–26 | 52–62 | 102–104 | [82] | 2 |
| S. trutta | 10 localities across all Sweden | SW | 14 | 80 | 20 | 60 | 100 | [83] | 1 |
4.1. Cytotaxonomy and Diversity of Eurasian Trouts
4.2. Cytogenomics in Salmonids
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Delling, B.; Doario, I. Systematics of the trouts endemic to Moroccan lakes, with description of a new species (Teleostei: Salmonidae). Ichthyol. Explor. Freshw. 2005, 16, 49–64. [Google Scholar]
- Doadrio, I.; Perea, S.; Yahyaoui, A. Two new species of atlantic trout (Actynopterygii, Salmonidae) from Morocco. Graellsia 2015, 71, e031. [Google Scholar] [CrossRef]
- Doadrio, I. Ictiofauna Continental Española: Bases Para Su Seguimiento; Ministerio de Medio Ambiente y Medio Rural y Marino, Centro de Publicaciones: Madrid, Spain, 2011; ISBN 84-491-1158-7. [Google Scholar]
- Kottelat, M.; Freyhof, J. Handbook of European Freshwater Fishes; Publications Kottelat: Cornol, Switzerland, 2007; ISBN 2-8399-0298-2. [Google Scholar]
- Delling, B. Diversity of western and southern Balkan trouts, with the description of a new species from the Louros River, Greece (Teleostei: Salmonidae). Ichthyol. Explor. Freshw. 2010, 21, 331. [Google Scholar]
- Turan, D.; Kottelat, M.; Engin, S. Two new species of trouts, resident and migratory, sympatric in streams of northern Anatolia (Salmoniformes: Salmonidae). Ichthyol. Explor. Freshw. 2009, 20, 333–364. [Google Scholar]
- Turan, D.; Kottelat, M.; Bektas, Y. Salmo tigridis, a new species of trout from the Tigris River, Turkey (Teleostei: Salmonidae). Zootaxa 2011, 2993, 23–33. [Google Scholar] [CrossRef]
- Turan, D.; Kottelat, M.; Engin, S. The trouts of the Mediterranean drainages of southern Anatolia, Turkey, with description of three new species (Teleostei: Salmonidae). Ichthyol. Explor. Freshw. 2012, 23, 219. [Google Scholar]
- Turan, D.; Doğan, E.; Kaya, C.; Kanyılmaz, M. Salmo kottelati, a new species of trout from Alakır Stream, draining to the Mediterranean in southern Anatolia, Turkey (Teleostei, Salmonidae). ZooKeys 2014, 135–151. [Google Scholar] [CrossRef]
- Turan, D.; Kottelat, M.; Kaya, C. Salmo munzuricus, a new species of trout from the Euphrates River drainage, Turkey (Teleostei: Salmonidae). Ichthyol. Explor. Freshw. 2017, 28, 55–63. [Google Scholar]
- Çiçek, E.; Fricke, R.; Sungur, S.; Eagderi, S. Endemic freshwater fishes of Turkey. FishTaxa 2018, 3, 1–39. [Google Scholar]
- Lobón-Cerviá, J.; Sanz, N. Brown trout: Biology, ecology and management; John Wiley & Sons: Hoboken, NJ, USA, 2017; ISBN 1-119-26831-1. [Google Scholar]
- Lobón-Cerviá, J.; Esteve, M.; Berrebi, P.; Duchi, A.; Lorenzoni, M.; Young, K.A. Trout and char of central and Southern Europe and Northern Africa. In Trout and char of the world; American Fisheries Society: Bethesda, MA, USA, 2019. [Google Scholar]
- Behnke, R.J. A new subgenus and species of trout, Salmo (Platysalmo) platycephalus, from southcentral Turkey, with comments on the classification of the subfamily Salmoninae. Mitt. Hamburg. Zool. Mus. Inst. 1968, 66, 1–15. [Google Scholar]
- Kara, C.; Alp, A.; Gürlek, M.E. Morphological variations of the trouts (Salmo trutta and Salmo platycephalus) in the rivers of Ceyhan, Seyhan and Euphrates, Turkey. Turk. J. Fish. Aquat. Sci. 2011, 11, 77–85. [Google Scholar]
- Kara, C.; Alp, A.; Can, M.F. Growth and reproductive properties of flathead trout (Salmo platycephalus Bhenke, 1968) population from Zamantı Stream, Seyhan River, Turkey. Turk. J. Fish. Aquat. Sci. 2011, 11, 367–375. [Google Scholar]
- Tarkan, A.N.; Tarkan, A.S.; Bilge, G.; Gaygusuz, Ö.; Gürsoy, Ç. Threatened fishes of the world: Salmo platycephalus Behnke, 1968 (Salmonidae). Environ. Biol. Fishes 2008, 81, 371–372. [Google Scholar] [CrossRef]
- Phillips, R.; Ráb, P. Chromosome evolution in the Salmonidae (Pisces): An update. Biol. Rev. Camb. Philos. Soc. 2001, 76, 1–25. [Google Scholar] [CrossRef]
- Mazzuchelli, J.; Kocher, T.D.; Yang, F.; Martins, C. Integrating cytogenetics and genomics in comparative evolutionary studies of cichlid fish. BMC Genom. 2012, 13, 463. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, E.A.; Bertollo, L.A.C.; Ráb, P.; Ezaz, T.; Yano, C.F.; Hatanaka, T.; Jegede, O.I.; Tanomtong, A.; Liehr, T.; Sember, A.; et al. Cytogenetics, genomics and biodiversity of the South American and African Arapaimidae fish family (Teleostei, Osteoglossiformes). PLoS ONE 2019, 14, e0214225. [Google Scholar] [CrossRef]
- Pennell, W.; Prouzet, K. Salmonid fish: Bology, conservation status, and economic importance of wild and cultured stocks. Fish. Aquac. P Safran Enciclopedia Life Suport Syst. 2009, 3, 42–65. [Google Scholar]
- Gregory, T.R. Animal Genome Size Database. 2020. Available online: http://www.genomesize.com (accessed on 6 December 2020).
- Koop, B.F.; Davidson, W.S. Genomics and the genome duplication in Salmonids. In Fisheries for Global Welfare and Environment; Tsukamoto, K., Kawamura, R., Takeuchi, T., Beard, T.D., Jr., Kaiser, M.J., Eds.; 5th World Fisheries Congress: Tokyo, Japan, 2008; pp. 77–86. [Google Scholar]
- Davidson, W.S.; Koop, B.F.; Jones, S.J.; Iturra, P.; Vidal, R.; Maass, A.; Jonassen, I.; Lien, S.; Omholt, S.W. Sequencing the genome of the Atlantic salmon (Salmo salar). Genome Biol. 2010, 11, 403. [Google Scholar] [CrossRef]
- Hunt, S.E.; McLaren, W.; Gil, L.; Thormann, A.; Schuilenburg, H.; Sheppard, D.; Parton, A.; Armean, I.M.; Trevanion, S.J.; Flicek, P.; et al. Ensembl variation resources. Database 2018, 2018. [Google Scholar] [CrossRef]
- Matoulek, D.; Borůvková, V.; Ocalewicz, K.; Symonová, R. GC and repeats profiling along chromosomes — The future of fish compositional cytogenomics. Genes 2020, 11. this special issue. [Google Scholar]
- Canapa, A.; Barucca, M.; Biscotti, M.A.; Forconi, M.; Olmo, E. Transposons, genome size, and evolutionary insights in animals. Cytogenet. Genome Res. 2015, 147, 217–239. [Google Scholar] [CrossRef] [PubMed]
- Lien, S.; Koop, B.F.; Sandve, S.R.; Miller, J.R.; Kent, M.P.; Nome, T.; Hvidsten, T.R.; Leong, J.S.; Minkley, D.R.; Zimin, A.; et al. The Atlantic salmon genome provides insights into rediploidization. Nature 2016, 533, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Symonová, R.; Suh, A. Nucleotide composition of transposable elements likely contributes to AT/GC compositional homogeneity of teleost fish genomes. Mobile DNA 2019, 10, 49. [Google Scholar] [CrossRef] [PubMed]
- Borůvková, V.; Howell, W.; Matoulek, D.; Symonová, R. Quantitative approach to fish cytogenetics in the context of vertebrate genome evolution. Genes 2020, 11. this special issue. [Google Scholar]
- Ráb, P.; Roth, P. Cold-blooded vertebrates. In Methods of Chromosome Analysis; Balíček, P., Forejt, J., Rubeš, J., Eds.; Cytogenetic Section of Czechoslovakian Biological Society Publishers: Brno, Czech Republic, 1988; pp. 115–124. [Google Scholar]
- Sola, L.; Rossi, A.R.; Iaselli, V.; Rasch, E.M.; Monaco, P.J. Cytogenetics of bisexual/unisexual species of Poecilia. II. Analysis of heterochromatin and nucleolar organizer regions in Poecilia mexicana mexicana by C-banding and DAPI, quinacrine, chromomycin A3, and silver staining. Cytogen. Cell Genet. 1992, 60, 229–235. [Google Scholar] [CrossRef]
- Howell, W.M.; Black, D.A. Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: A 1-step method. Experientia 1980, 36, 1014–1015. [Google Scholar] [CrossRef]
- Rábová, M.; Völker, M.; Pelikánová, Š.; Ráb, P. Sequential chromosome banding in fishes. In Fish Cytogenetic Techniques; Ozouf-Costaz, C., Pisano, E., Foresti, F., de Almeida Toledo, L.F., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 92–102. ISBN 978-1-4822-1198-6. [Google Scholar]
- Komiya, H.; Takemura, S. Nucleotide sequence of 5S ribosomal RNA from rainbow trout (Salmo gairdnerii) liver. J. Biochem. 1979, 86, 1067–1080. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, J., Sninsky, J., White, T., Eds.; Academic Press: Orlando, FL, USA, 1990; pp. 315–322. [Google Scholar]
- Symonová, R.; Sember, A.; Majtánová, Z.; Ráb, P. Characterization of fish genomes by GISH and CGH. In Fish Cytogenetic Techniques; Ozouf-Costaz, C., Pisano, E., Foresti, F., de Almeida, L., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 118–131. ISBN 978-1-4822-1198-6. [Google Scholar]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Matthey, R. L’évolution de la formule chromosomiale chez les vertébrés. Experientia 1945, 1, 78–86. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing, version 2.6.2; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Sušnik, S.; Schöffmann, J.; Snoj, A. Phylogenetic position of Salmo (Platysalmo) platycephalus Behnke 1968 from south-central Turkey, evidenced by genetic data. J. Fish. Biol. 2004, 64, 947–960. [Google Scholar] [CrossRef]
- Bănărescu, P. Zoogeography of Fresh Waters. Volume 2: Distribution and Dispersal of Freshwater Animals in North America and Eurasia; Aula-Verlag: Wiebelsheim, Germany, 1991; ISBN 3-89104-482-8. [Google Scholar]
- Budy, P.; Gaeta, J.W. Brown trout as an invader: A Synthesis of problems and perspectives in North America. In Brown trout: Biology, ecology, and management; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 2018; pp. 525–534. [Google Scholar]
- Jones, P.; Closs, G. The introduction of brown trout to New Zealand and their impact on native fish communities. In Brown Trout: Biology, Ecology, and Management; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 2018; pp. 545–567. [Google Scholar]
- Jellyman, P.G.; McHugh, P.A.; Simon, K.S.; Thompson, R.M.; McIntosh, A.R. The effects of brown trout on the trophic webs of New Zealand streams. In Brown Trout: Biology, Ecology and Management; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 569–598. [Google Scholar]
- Casalinuovo, M.A.; Alonso, M.F.; Macchi, P.J.; Kuroda, J.A. Brown trout in Argentina: History, interactions, and perspectives. In Brown Trout: Biology, Ecology, and Management; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 2018; pp. 599–622. [Google Scholar]
- Weyl, O.L.; Ellender, B.R.; Ivey, P.; Jackson, M.C.; Tweddle, D.; Wasserman, R.J.; Woodford, D.J.; Zengeya, T.A. Africa: Brown trout introductions, establishment, current status, impacts and conflicts. In Brown trout: Biology, ecology, and management; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 2018; pp. 623–639. [Google Scholar]
- FAO Fisheries and Aquaculture—Statistics—Introduction. Available online: http://www.fao.org/fishery/statistics/en (accessed on 6 November 2020).
- GAs, M. Revue bibliographique sur la caryologie des Téléostéens. Etude critique des méthodes employées et des résultats obtenus. Biologie Médical 1970, 54, 54–81. [Google Scholar]
- Dettai, A.; Pruvost, P. Storage of Karyotyped Voucher Specimens and their Molecular Identification. In Fish Cytogenetic Techniques: Ray-Fin Fishes and Chondrichthyans; CRC Press Taylor and Francis Group: London, UK, 2015; p. 11. [Google Scholar]
- Kohout, J.; Jašková, I.; Papoušek, I.; Šedivá, A.; Šlechta, V. Effects of stocking on the genetic structure of brown trout, Salmo trutta, in Central Europe inferred from mitochondrial and nuclear DNA markers. Fish Manag. Ecol. 2012, 19, 252–263. [Google Scholar] [CrossRef]
- Leitwein, M.; Gagnaire, P.A.; Desmarais, E.; Guendouz, S.; Rohmer, M.; Berrebi, P.; Guinand, B. Genome-wide nucleotide diversity of hatchery-reared Atlantic and Mediterranean strains of brown trout Salmo trutta compared to wild M editerranean populations. J. Fish. Biol. 2016, 89, 2717–2734. [Google Scholar] [CrossRef] [PubMed]
- Mazik, E.J.; Toktosunov, A.T. Karyotype of the Amu Darya trout Salmo trutta oxianus (Salmoniformes, Salmonidae) from Kyzylsu River. J. Zool. 1986, 65, 1582–1586. [Google Scholar]
- Merlo, S. Osservazioni cariologiche su Salmo carpio: (Con 1 tavola fuori testo). Ital. J. Zool. 1957, 24, 253–258. [Google Scholar]
- Caputo, V.; Giovannotti, M.; Cerioni, P.N.; Splendiani, A.; Olmo, E. Chromosomal study of native and hatchery trouts from Italy (Salmo trutta complex, Salmonidae): Conventional and FISH analysis. Cytogen. Gen. Res. 2009, 124, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Karakousis, Y.; Paschos, J.; Triantaphyllidis, C. Chromosomal studies in brown trout (Salmo trutta L.) populations. Cytobios 1992, 72, 117–124. [Google Scholar]
- Dimovska, A. Chromosome complement of Ochrid trout (Salmo letnica Karaman). Godisen zb. Prirodno-matem. Fak. Univ. Skopje 1959, 12, 115–135. [Google Scholar]
- Al-Sabti, K. Karyotypical studies in three Salmonidaae in Slovenia using leukocyte culture technique. Ichthyologia 1983, 15, 41–46. [Google Scholar]
- Al-Sabti, K. Chromosomal studies by blood leukocyte culture technique on three salmonids from Yugoslavian waters. J. Fish. Biol. 1985, 26, 5–12. [Google Scholar] [CrossRef]
- Berberovic, L.; Curic, M.; Hadziselimovic, R.; Sofradzija, A. Chromosome complement of Salmothymus obtusirostris oxyrhynchus (Steindachner). Acta Biol. Jug. Genet. 1970, 2, 55–63. [Google Scholar]
- Sofradzija, A. The chromosomes of the trout Salmo trutta m. fario and Salmo gairdneri. Godisnajak Bioloskog Instituta Univerziteta u Sarajevu 1982, 35, 117–128. [Google Scholar]
- Kaidanova, T.I. Karyotype study of brown trout Salmo trutta morpha fario from Ropsha population. Voprosy Ichtiologii 1975, 15, 1124–1128. [Google Scholar]
- Jankun, M. Standard karyotype of sea trout (Salmo trutta morpha trutta) based on replication banding patterns. Cytobios 2000, 103, 79–89. [Google Scholar] [PubMed]
- Woznicki, P.; Jankun, M.; Kucharczyk, D.; Boron, A.; Luczynski, M. Cytogenetic characterization of sea trout (Salmo trutta) from Poland. Copeia 1999, 1999, 501–505. [Google Scholar] [CrossRef]
- Woznicki, P.; Sanchez, L.; Martinez, P.; Pardo, B.G.; Jankun, M. A population analysis of the structure and variability of NOR in Salmo trutta by Ag, CMA3 and ISH. Genetica 2000, 108, 113–118. [Google Scholar] [CrossRef]
- Woznicki, P.; Jankun, M.; Luczynski, M. Chromosome studies in brown trout (Salmo trutta m. fario) from Poland: Hypothetical evolution of the 11th, 12th and 14th chromosome pairs in the Salmo karyotype. Cytobios 1997, 91, 2017–2214. [Google Scholar]
- Dorofeeva, E.A.; Ruhkjan, R.G. Divergence of Salmo ischchan Kessler in light of karyological and morphological characteristics. Voprosy Ichtiologii 1982, 22, 36–48. [Google Scholar]
- Ruhkjan, R.G. Karyology and Origin of the Transcaucasian Trouts; Academy of Sciences Armenian SSR Press: Yerevan, Armenia, 1989; p. 166. (In Russian) [Google Scholar]
- Raicu, P.; Taisescu, E. Cytogenetic study in Salmo irideus and S. trutta fario. Cytologia 1977, 42, 311–314. [Google Scholar] [CrossRef]
- Dorofeeva, E.A. Karyology and systematic status of Caspian and Black Sea salmons (Salmo trutta caspius Kessler, Salmo trutta labrax Pallas). Voprosy Ichtiologii 1965, 5, 28–45. [Google Scholar]
- Ruhkjan, R.G. A comparative analysis of the karyotypes of the Sevan trout Salmo ischchan Kessler. Citologia 1982, 24, 66–77. [Google Scholar]
- Ruhkjan, R.G. On the origin and species identity of alabalach trout (genus Salmo, Salmonidae) based on its karyological characteristics). Voprosy Ichtiologi 1984, 23, 368–373. [Google Scholar]
- Ruhkjan, R.G. Karyotypes of brown trouts of Armenia Salmo trutta m. fario. Biol. J. Armenii 1981, 34, 412–417. [Google Scholar]
- Mořan, P.; Pendas, A.M.; García-Vázquez, E.; Linde, A.R. Chromosomal and morphological analysis of two populations of Salmo trutta sbp. fario employed in repopulation. J. Fish Biol. 1989, 35, 839–843. [Google Scholar] [CrossRef]
- Martinez, P.; Vinas, A.; Bouza, C.; Arias, J.; Amaro, R.; Sánchez, L. Cytogenetical characterization of hatchery stocks and natural populations of sea and brown trout from northwestern Spain. Heredity 1991, 66, 9–17. [Google Scholar] [CrossRef]
- Mayr, B.; Ráb, P.; Kalat, M. Localisation of NORs and counterstain-enhanced fluorescence studies in Salmo gairdneri and Salmo trutta (Pisces, Salmonidae). Theoret. Appl. Genet. 1986, 71, 703–707. [Google Scholar] [CrossRef]
- Castro, J.; Rodríguez, S.; Arias, J.; Sánchez, L.; Martínez, P. A population analysis of Robertsonian and Ag-NOR polymorphisms in brown trout (Salmo trutta). Theoret. Appl. Genetics 1994, 89, 105–111. [Google Scholar] [CrossRef]
- Castro, J.; Viñas, A.; Sánchez, L.; Martínez, P. Characterization of an atypical NOR site polymorphism in brown trout (Salmo trutta) with Ag- and CMA3-staining, and fluorescent in situ hybridization. Cytogenet. Genome Res. 1996, 75, 234–239. [Google Scholar] [CrossRef]
- Sánchez, L.; Martínez, P.; Bouza, C.; Viñas, A. Chromosomal heterochromatin differentiation in Salmo trutta with restriction enzymes. Heredity 1991, 66, 241–249. [Google Scholar] [CrossRef][Green Version]
- Hartley, S.E.; Horne, M.T. Chromosome relationships in the genus Salmo. Chromosoma 1984, 90, 229–237. [Google Scholar] [CrossRef]
- Gjedrem, T.; Eggum, Å.; Refstie, T. Chromosomes of some salmonids and salmonid hybrids. Aquaculture 1977, 11, 335–348. [Google Scholar] [CrossRef]
- Zenzes, M.T.; Voiculescu, I. C-banding patterns in Salmo trutta, a species of tetraploid origin. Genetica 1975, 45, 531–536. [Google Scholar] [CrossRef]
- Nygren, A.; Nilsson, B.; Jahnke, M. Cytological studies in Salmo trutta and Salmo alpinus. Hereditas 1971, 67, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Pendás, A.M.; Morán, P.; García-Vázquez, E. Multi-chromosomal location of ribosomal RNA genes and heterochromatin association in brown trout. Chromosome Res. 1993, 1, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Gornung, E. Twenty Years of Physical Mapping of Major Ribosomal RNA Genes across the Teleosts: A Review of Research. Cytogenet. Genome Res. 2013. [Google Scholar] [CrossRef] [PubMed]
- Sember, A.; Bohlen, J.; Šlechtová, V.; Altmanová, M.; Symonová, R.; Ráb, P. Karyotype differentiation in 19 species of river loach fishes (Nemacheilidae, Teleostei): Extensive variability associated with rDNA and heterochromatin distribution and its phylogenetic and ecological interpretation. BMC Evol. Biol. 2015, 15, 251. [Google Scholar] [CrossRef]
- Majtánová, Z.; Unmack, P.J.; Prasongmaneerut, T.; Shams, F.; Srikulnath, K.; Ráb, P.; Ezaz, T. Evidence of Interspecific Chromosomal Diversification in Rainbowfishes (Melanotaeniidae, Teleostei). Genes 2020, 11, 818. [Google Scholar] [CrossRef]
- Ráb, P.; Rábová, M.; Pereira, C.S.; Collares-Pereira, M.J.; Pelikánová, Š. Chromosome studies of European cyprinid fishes: Interspecific homology of leuciscine cytotaxonomic marker—the largest subtelocentric chromosome pair as revealed by cross-species painting. Chromosome Res. 2008, 16, 863–873. [Google Scholar] [CrossRef]
- Sušnik, S.; Snoj, A.; Wilson, I.F.; Mrdak, D.; Weiss, S. Historical demography of brown trout (Salmo trutta) in the Adriatic drainage including the putative S. letnica endemic to Lake Ohrid. Mol. Phylogen. Evol. 2007, 44, 63–76. [Google Scholar] [CrossRef]
- Phillips, R.B.; Matsuoka, M.P.; Konon, I.; Reed, K.M.; McEachran, M. Phylogenetic Analysis of Mitochondrial and Nuclear Sequences Supports Inclusion of Acantholingua ohridana in the Genus Salmo. Copeia 2000, 2000, 546–550. [Google Scholar] [CrossRef]
- Kottelat, M. Freshwater fishes of western and central Europe. Biologia 1997, 52, 1–271. [Google Scholar]
- Behnke, R.J. Native trout of western North America. Am. Fish. Soc. Monogr. USA 1992, 6, 233–256. [Google Scholar]
- FishBase. A Global Information System on Fishes. Available online: https://www.fishbase.se/home.htm (accessed on 6 November 2020).
- Cortey, M.; Pla, C.; García-Marín, J.-L. Historical biogeography of Mediterranean trout. Mol. Phylogenetics Evol. 2004, 33, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Marić, S.; Sušnik, S.; Simonović, P.; Snoj, A. Phylogeographic study of brown trout from Serbia, based on mitochondrial DNA control region analysis. Genet. Sel. Evol. 2006, 38, 411. [Google Scholar] [CrossRef][Green Version]
- McKeown, N.J.; Hynes, R.A.; Duguid, R.A.; Ferguson, A.; Prodöhl, P.A. Phylogeographic structure of brown trout Salmo trutta in Britain and Ireland: Glacial refugia, postglacial colonization and origins of sympatric populations. J. Fish. Biol. 2010, 76, 319–347. [Google Scholar] [CrossRef]
- Snoj, A.; Marić, S.; Bajec, S.S.; Berrebi, P.; Janjani, S.; Schöffmann, J. Phylogeographic structure and demographic patterns of brown trout in North-West Africa. Mol. Phylogenetics Evol. 2011, 61, 203–211. [Google Scholar] [CrossRef]
- Ninua, L.; Tarkhnishvili, D.; Gvazava, E. Phylogeography and taxonomic status of trout and salmon from the Ponto-Caspian drainages, with inferences on European Brown Trout evolution and taxonomy. Ecol. Evol. 2018, 8, 2645–2658. [Google Scholar] [CrossRef]
- Tougard, C.; Justy, F.; Guinand, B.; Douzery, E.J.P.; Berrebi, P. Salmo macrostigma (Teleostei, Salmonidae): Nothing more than a brown trout (S. trutta) lineage? J. Fish. Biol. 2018, 93, 302–310. [Google Scholar] [CrossRef]
- Schöffmann, J.; Sušnik, S.; Snoj, A. Phylogenetic origin of Salmo trutta L. 1758 from Sicily, based on mitochondrial and nuclear DNA analyses. Hydrobiologia 2007, 575, 51–55. [Google Scholar] [CrossRef]
- Apostolidis, A.P.; Loukovitis, D.; Tsigenopoulos, C.S. Genetic characterization of brown trout (Salmo trutta) populations from the Southern Balkans using mtDNA sequencing and RFLP analysis. Hydrobiologia 2008, 600, 169–176. [Google Scholar] [CrossRef]
- Gratton, P.; Allegrucci, G.; Gandolfi, A.; Sbordoni, V. Genetic differentiation and hybridization in two naturally occurring sympatric trout Salmo spp. forms from a small karstic lake. J. Fish. Biol. 2013, 82, 637–657. [Google Scholar] [CrossRef] [PubMed]
- Berrebi, P.; Retif, X.; Bouhbouh, S. Genetic evidence of unisexual reproduction in the Moroccan hexaploid barbel Labeobarbus fritschi. Folia Zoo. 2013, 62, 257–263. [Google Scholar] [CrossRef]
- Delling, B.; Sabatini, A.; Muracciole, S.; Tougard, C.; Berrebi, P. Morphologic and genetic characterisation of Corsican and Sardinian trout with comments on Salmo taxonomy. Knowl. Manag. Aquat. Ecosyst. 2020, 21. [Google Scholar] [CrossRef]
- Dion-Côté, A.-M.; Symonová, R.; Lamaze, F.C.; Pelikánová, Š.; Ráb, P.; Bernatchez, L. Standing chromosomal variation in Lake Whitefish species pairs: The role of historical contingency and relevance for speciation. Mol. Ecol. 2017, 26, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Mandáková, T.; Heenan, P.B.; Lysak, M.A. Island species radiation and karyotypic stasis in Pachycladon allopolyploids. BMC Evol. Biol. 2010, 10, 367. [Google Scholar] [CrossRef]
- Bomfleur, B.; McLoughlin, S.; Vajda, V. Fossilized nuclei and chromosomes reveal 180 million years of genomic stasis in royal ferns. Science 2014, 343, 1376–1377. [Google Scholar] [CrossRef]
- Samad, N.A.; Dagher-Kharrat, M.B.; Hidalgo, O.; Zein, R.E.; Douaihy, B.; Siljak-Yakovlev, S. Unlocking the karyological and cytogenetic diversity of Iris from Lebanon: Oncocyclus section shows a distinctive profile and relative stasis during its continental radiation. PLoS ONE 2016, 11, e0160816. [Google Scholar] [CrossRef]
- Sessions, S.K.; Kezer, J. Chapter 5—Evolutionary cytogenetics of Bolitoglossine Salamanders (Family Plethodontidae). In Amphibian Cytogenetics and Evolution; Green, D.M., Sessions, S.K., Eds.; Academic Press: San Diego, CA, USA, 1991; pp. 89–130. ISBN 978-0-12-297880-7. [Google Scholar]
- Odierna, G.; Aprea, G.; Capriglion, T.; Castellano, S.; Balletto, E. Evidence for chromosome and Pst I satellite DNA family evolutionary stasis in the Bufo viridis group (Amphibia, Anura). Chromosome Res. 2004, 12, 671–681. [Google Scholar] [CrossRef]
- Ellegren, H. Evolutionary stasis: The stable chromosomes of birds. Trends Ecol. Evol. 2010, 25, 283–291. [Google Scholar] [CrossRef]
- Ráb, P.; Crossman, E.J. Chromosomal NOR phenotypes in North American pikes and pickerels, genus Esox, with notes on the Umbridae (Euteleostei: Esocae). Canadian J. Zool. 2011. [Google Scholar] [CrossRef]
- Symonová, R.; Ocalewicz, K.; Kirtiklis, L.; Delmastro, G.B.; Pelikánová, Š.; Garcia, S.; Kovařík, A. Higher-order organisation of extremely amplified, potentially functional and massively methylated 5S rDNA in European pikes (Esox sp.). BMC Genom. 2017, 18, 391. [Google Scholar] [CrossRef]
- Ráb, P.; Collares-Pereira, M.J. Chromosomes of European cyprinid fishes (Cyprinidae, Cypriniformes) Review. Folia Zool. 1995, 44, 193–214. [Google Scholar]
- Bianco, P.G.; Aprea, G.; Balletto, E.; Capriglione, T.; Fulgione, D.; Odierna, G. The karyology of the cyprinid genera Scardinius and Rutilus in southern Europe. Ichthyol. Res. 2004, 51, 274–278. [Google Scholar] [CrossRef]
- Pereira, C.S.A.; Ráb, P.; Collares-Pereira, M.J. Chromosomes of European cyprinid fishes: Comparative cytogenetics and chromosomal characteristics of ribosomal DNAs in nine Iberian chondrostomine species (Leuciscinae). Genetica 2012, 140, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Armbruster, J.W. Phylogenetic classification of extant genera of fishes of the order Cypriniformes (Teleostei: Ostariophysi). Zootaxa 2018, 4476, 6–39. [Google Scholar] [CrossRef] [PubMed]
- Sember, A.; Pelikánová, Š.; de Bello Cioffi, M.; Šlechtová, V.; Hatanaka, T.; Do Doan, H.; Knytl, M.; Ráb, P. Taxonomic diversity not associated with gross karyotype differentiation: The case of bighead carps, Genus Hypophthalmichthys (Teleostei, Cypriniformes, Xenocyprididae). Genes 2020, 11, 479. [Google Scholar] [CrossRef] [PubMed]
- Molina, W.F. Chromosomal changes and stasis in marine fish groups. In Fish Cytogenetics; Pisano, E., Ozouf-Costaz, C., Foresti, F., Kapoor, B.G., Eds.; Science Publishers: Enfield, NH, USA, 2007; pp. 69–110. ISBN 1-57808-330-3. [Google Scholar]
- Motta Neto, C.C.; Cioffi, M.D.B.; Costa, G.W.W.F.; Amorim, K.D.J.; Bertollo, L.A.C.; Artoni, R.F.; Molina, W.F. Overview on karyotype stasis in atlantic grunts (Eupercaria, Haemulidae) and the Evolutionary Extensions for Other Marine Fish Groups. Front. Mar. Sci. 2019, 6. [Google Scholar] [CrossRef]
- Neto, C.C.M.; Cioffi, M.B.; Bertollo, L.A.C.; Molina, W.F. Extensive chromosomal homologies and evidence of karyotypic stasis in Atlantic grunts of the genus Haemulon (Perciformes). J. Exp. Mar. Biol. Ecol. 2011, 401, 75–79. [Google Scholar] [CrossRef]
- Neto, C.C.M.; Cioffi, M.B.; Bertollo, L.A.C.; Molina, W.F. Molecular cytogenetic analysis of Haemulidae fish (Perciformes): Evidence of evolutionary conservation. J. Exp. Mar. Biol. Ecol. 2011, 407, 97–100. [Google Scholar] [CrossRef]
- da Costa, G.W.W.F.; de Bello Cioffi, M.; Bertollo, L.A.C.; Molina, W.F. The Evolutionary Dynamics of ribosomal genes, histone H3, and transposable Rex elements in the genome of Atlantic snappers. J. Hered. 2016, 107, 173–180. [Google Scholar] [CrossRef]
- Majtánová, Z.; Indermaur, A.; Bitja Nyom, A.R.; Ráb, P.; Musilová, Z. Adaptive radiation from a chromosomal perspective: Evidence of chromosome set stability in cichlid fishes (Cichlidae: Teleostei) from the Barombi Mbo Lake, Cameroon. Int. J. Mol. Sci. 2019, 20, 4994. [Google Scholar] [CrossRef] [PubMed]
- Razin, S.V.; Gavrilov, A.A.; Vassetzky, Y.S.; Ulianov, S.V. Topologically-associating domains: Gene warehouses adapted to serve transcriptional regulation. Transcription 2016, 7, 84–90. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rosin, L.F.; Crocker, O.; Isenhart, R.L.; Nguyen, S.C.; Xu, Z.; Joyce, E.F. Chromosome territory formation attenuates the translocation potential of cells. eLife 2019, 8, e49553. [Google Scholar] [CrossRef] [PubMed]
- Symonová, R.; Howell, W.M. Vertebrate genome evolution in the light of fish cytogenomics and rDNAomics. Genes 2018, 9, 96. [Google Scholar] [CrossRef]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; Wiley: Hoboken, NJ, USA, 2016; ISBN 978-1-118-34233-6. [Google Scholar]
- Fujiwara, A.; Abe, S.; Yamaha, E.; Yamazaki, F.; Yoshida, M.C. Chromosomal localization and heterochromatin association of ribosomal RNA gene loci and silver-stained nucleolar organizer regions in salmonid fishes. Chrom. Res. 1998, 6, 463–471. [Google Scholar] [CrossRef]
- Symonová, R.; Majtánová, Z.; Sember, A.; Staaks, G.B.O.; Bohlen, J.; Freyhof, J.; Rábová, M.; Ráb, P. Genome differentiation in a species pair of coregonine fishes: An extremely rapid speciation driven by stress-activated retrotransposons mediating extensive ribosomal DNA multiplications. BMC Evol. Biol. 2013, 13, 42. [Google Scholar] [CrossRef]
- Dion-Côté, A.-M.; Symonová, R.; Ráb, P.; Bernatchez, L. Reproductive isolation in a nascent species pair is associated with aneuploidy in hybrid offspring. Proc. Royal Soc. B 2015, 282, 20142862. [Google Scholar] [CrossRef]
- Bernardi, G. Structural and Evolutionary Genomics: Natural Selection in Genome Evolution; Elsevier: Amsterdam, The Netherlands, 2005; ISBN 978-0-08-046187-8. [Google Scholar]
- Symonová, R. Integrative rDNAomics—Importance of the oldest repetitive fraction of the Eukaryote genome. Genes 2019, 10, 345. [Google Scholar] [CrossRef]
- Sochorová, J.; Garcia, S.; Gálvez, F.; Symonová, R.; Kovařík, A. Evolutionary trends in animal ribosomal DNA loci: Introduction to a new online database. Chromosoma 2018, 127, 141–150. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaffaroglu, M.; Majtánová, Z.; Symonová, R.; Pelikánová, Š.; Unal, S.; Lajbner, Z.; Ráb, P. Present and Future Salmonid Cytogenetics. Genes 2020, 11, 1462. https://doi.org/10.3390/genes11121462
Gaffaroglu M, Majtánová Z, Symonová R, Pelikánová Š, Unal S, Lajbner Z, Ráb P. Present and Future Salmonid Cytogenetics. Genes. 2020; 11(12):1462. https://doi.org/10.3390/genes11121462
Chicago/Turabian StyleGaffaroglu, Muhammet, Zuzana Majtánová, Radka Symonová, Šárka Pelikánová, Sevgi Unal, Zdeněk Lajbner, and Petr Ráb. 2020. "Present and Future Salmonid Cytogenetics" Genes 11, no. 12: 1462. https://doi.org/10.3390/genes11121462
APA StyleGaffaroglu, M., Majtánová, Z., Symonová, R., Pelikánová, Š., Unal, S., Lajbner, Z., & Ráb, P. (2020). Present and Future Salmonid Cytogenetics. Genes, 11(12), 1462. https://doi.org/10.3390/genes11121462