Genotypic and Phenotypic Characterization of Incompatibility Group FIB Positive Salmonella enterica Serovar Typhimurium Isolates from Food Animal Sources


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Phenotypic Testing
2.2.1. Antimicrobial Drug Susceptibility Testing (AST)
2.2.2. Colicin Inhibition Assay
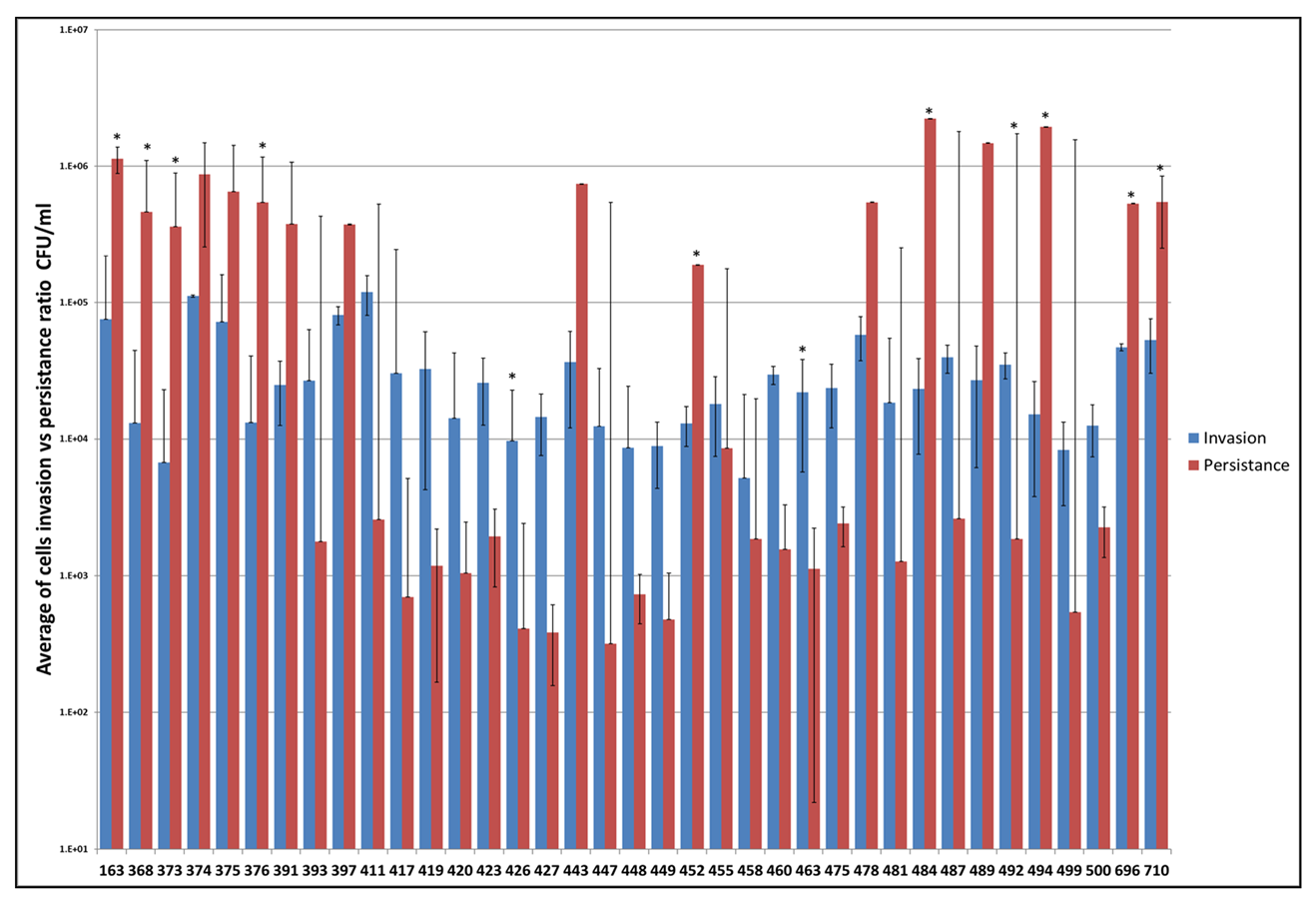
2.2.3. In Vitro Invasion and Persistence Assay
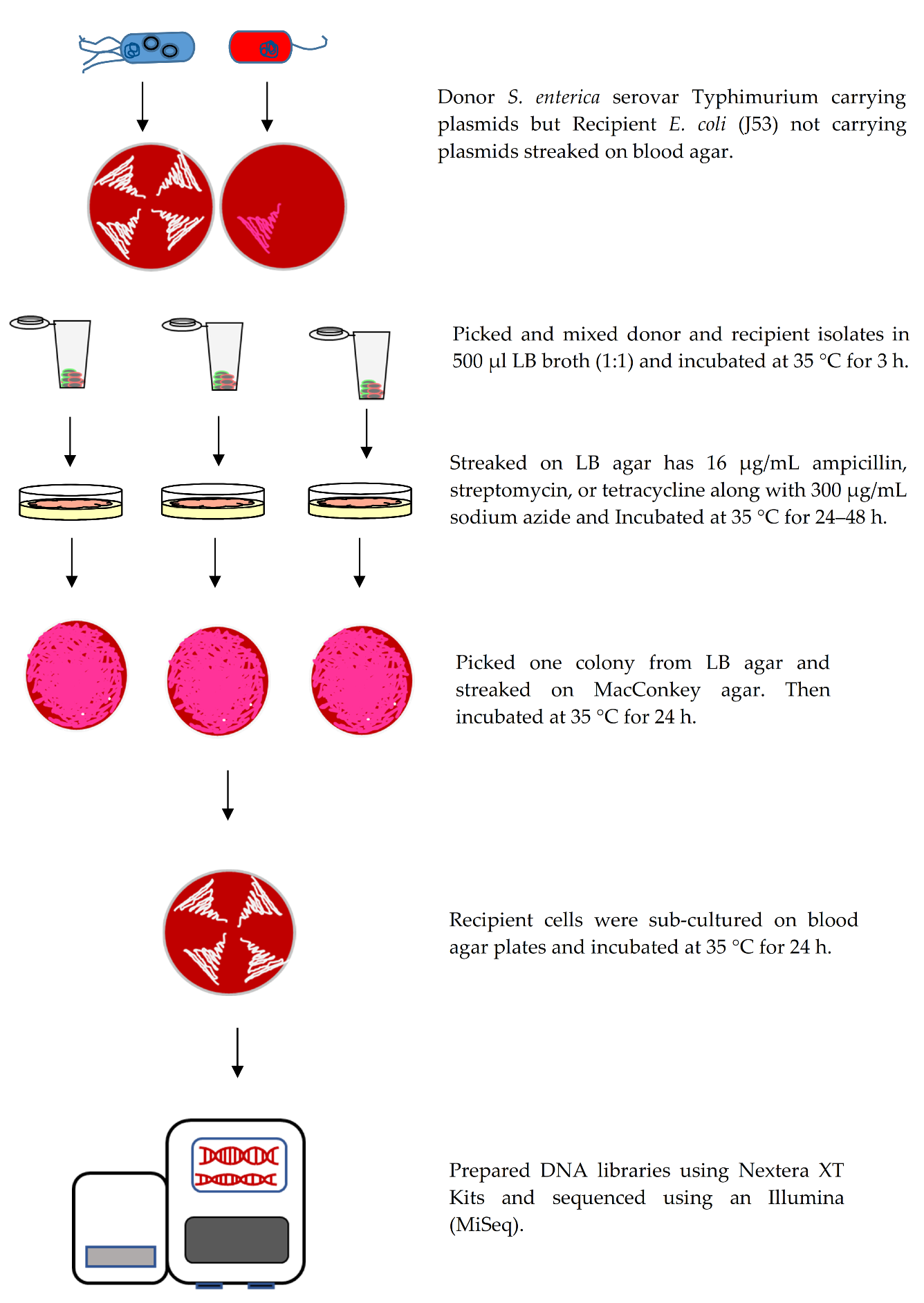
2.2.4. Conjugation Assay
2.3. Genotypic Testing
2.3.1. Whole Genome Sequencing (WGS)
2.3.2. Single Nucleotide Polymorphism (SNP) Analysis
2.3.3. Bioinformatic Analyses
2.4. Statistical Analysis
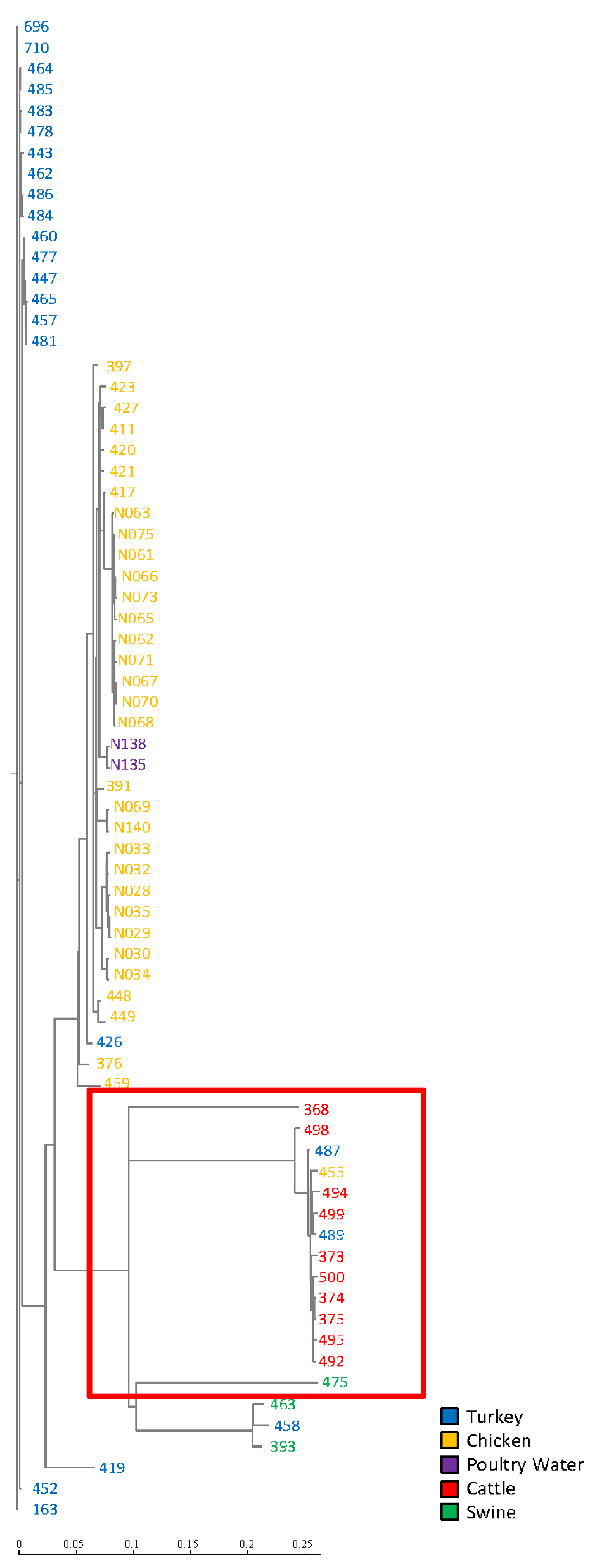
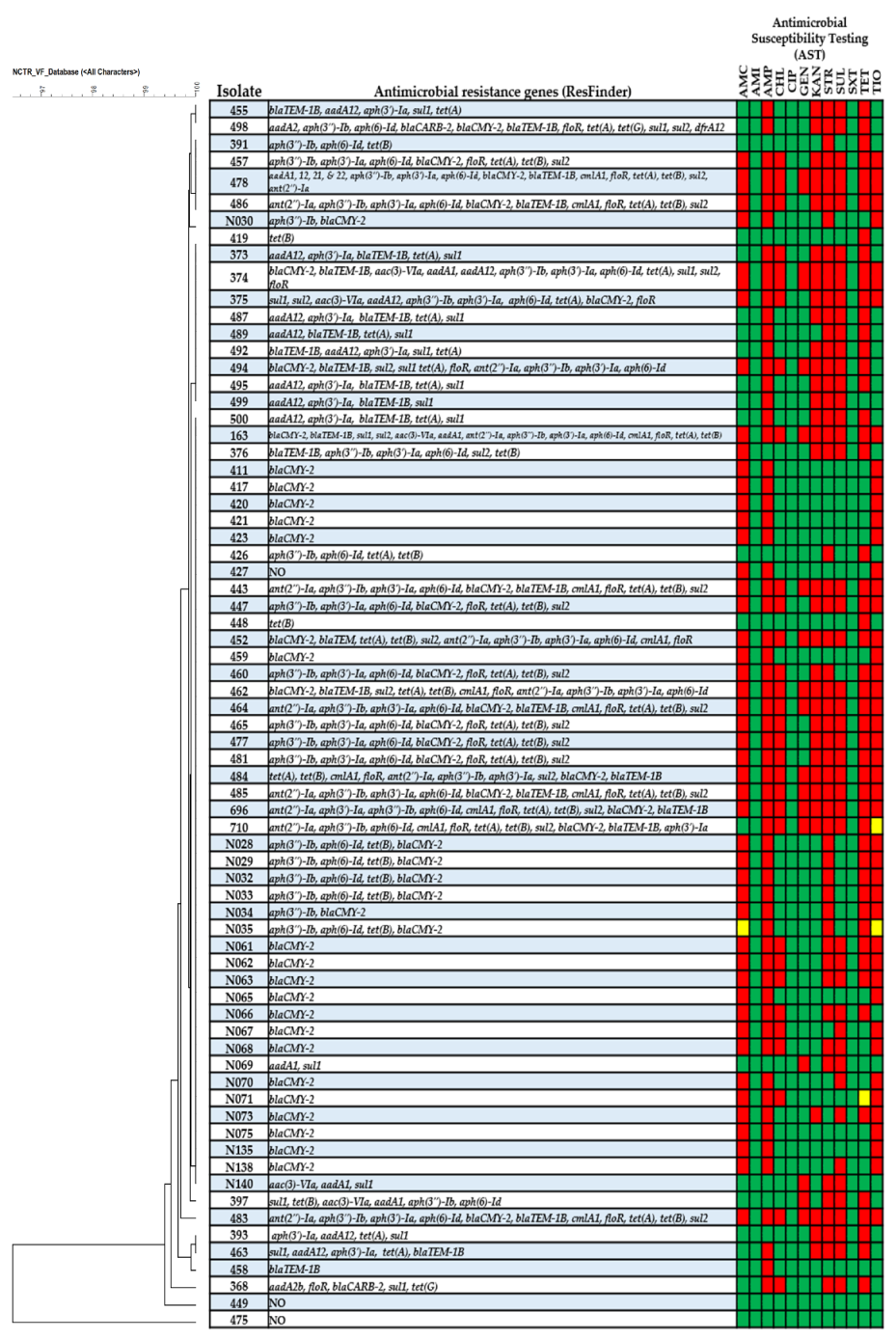
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
References
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the united states-major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- CDC. National Enteric Disease Surveillance Salmonella Annual Report. Available online: https://www.cdc.gov/salmonella/index.html (accessed on 5 March 2020).
- USDA. Cost Estimates of Foodborne Illnesses, Cost of Foodborne Illness Estimates for Salmonella (Non-typhoidal). Available online: https://www.ers.usda.gov/data-products/cost-estimates-of-foodborne-illnesses.aspx#48498 (accessed on 15 June 2020).
- Hoffman, S.; Maculloch, B.; Batz, M. Economic burden of major foodborne illnesses acquired in the United States. Econ. Res. Serv. 2015. [Google Scholar] [CrossRef]
- Foley, S.L.; Johnson, T.J.; Ricke, S.C.; Nayak, R.; Danzeisen, J. Salmonella pathogenicity and host adaptation in chicken-associated serovars. Microbiol. Mol. Biol. Rev. 2013, 77, 582–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC. National Salmonella Surveillance. Available online: https://www.cdc.gov/nationalsurveillance/salmonella-surveillance.html (accessed on 23 May 2019).
- CDC. An Altas of Salmonella in the United States, 1968–2011. Available online: https://www.cdc.gov/salmonella/pdf/salmonella-atlas-508c.pdf (accessed on 27 June 2019).
- Janakiraman, A.; Slauch, J.M. The putative iron transport system sitabcd encoded on spi1 is required for full virulence of salmonella typhimurium. Mol. Microbiol. 2000, 35, 1146–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winnen, B.; Schlumberger, M.C.; Sturm, A.; Schupbach, K.; Siebenmann, S.; Jenny, P.; Hardt, W.D. Hierarchical effector protein transport by the salmonella typhimurium spi-1 type iii secretion system. PLoS ONE 2008, 3, e2178. [Google Scholar] [CrossRef]
- Sabbagh, S.C.; Forest, C.G.; Lepage, C.; Leclerc, J.M.; Daigle, F. So similar, yet so different: Uncovering distinctive features in the genomes of salmonella enterica serovars typhimurium and typhi. FEMS Microbiol. Lett. 2010, 305, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.; Lynne, A.M.; David, D.E.; Tang, H.L.; Xu, J.S.; Nayak, R.; Kaldhone, P.; Logue, C.M.; Foley, S.L. DNA sequence analysis of plasmids from multidrug resistant salmonella enterica serotype heidelberg isolates. PLoS ONE 2012, 7, e51160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khajanchi, B.K.; Hasan, N.A.; Choi, S.Y.; Han, J.; Zhao, S.H.; Colwell, R.R.; Cerniglia, C.E.; Foley, S.L. Comparative genomic analysis and characterization of incompatibility group fib plasmid encoded virulence factors of salmonella enterica isolated from food sources. BMC Genom. 2017, 18, 14. [Google Scholar] [CrossRef] [Green Version]
- Kaldhone, P.R.; Han, J.; Deck, J.; Khajanchi, B.; Nayak, R.; Foley, S.L.; Ricke, S.C. Evaluation of the genetics and functionality of plasmids in incompatibility group i1-positive salmonella enterica. Foodborne Pathog. Dis. 2018, 15, 168–176. [Google Scholar] [CrossRef]
- Kaldhone, P.R.; Carlton, A.; Aljahdali, N.; Khajanchi, B.K.; Sanad, Y.M.; Han, J.; Deck, J.; Ricke, S.C.; Foley, S.L. Evaluation of incompatibility group i1 (inci1) plasmid-containing salmonella enterica and assessment of the plasmids in bacteriocin production and biofilm development. Front. Vet. Sci. 2019, 6, 298. [Google Scholar] [CrossRef]
- Jakociune, D.; Bisgaard, M.; Pedersen, K.; Olsen, J.E. Demonstration of persistent contamination of a cooked egg product production facility with salmonella enterica serovar Tennessee and characterization of the persistent strain. J. Appl. Microbiol. 2014, 117, 547–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szmolka, A.; Nagy, B. Multidrug resistant cornmensal escherichia coli in animals and its impact for public health. Front. Microbiol. 2013, 4, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanad, Y.M.; Johnson, K.; Park, S.H.; Han, J.; Deck, J.; Foley, S.L.; Kenney, B.; Ricke, S.; Nayak, R. Molecular characterization of salmonella enterica serovars isolated from a turkey production facility in the absence of selective antimicrobial pressure. Foodborne Pathog. Dis. 2016, 13, 80–87. [Google Scholar] [CrossRef]
- Aljahdali, N.H.; Kaldhone, P.R.; Foley, S.L.; Khajanchi, B.K. Whole-genome sequences of 35 incompatibility group i1 plasmid-carrying salmonella enterica isolates from food animal and clinical sources. Microbiol. Resour. Announc. 2019, 8, e00831-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aljahdali, N.H.; Foley, S.L.; Han, J.; Sanad, Y.M.; Nayak, R.; Khajanchi, B.K. Whole-genome sequences of 66 incompatibility group fib plasmid-carrying salmonella enterica serovar typhimurium isolates from food animal sources. Microbiol. Resour. Announc. 2020, 9. [Google Scholar] [CrossRef] [Green Version]
- Khajanchi, B.K.; Xu, J.; Grim, C.J.; Ottesen, A.R.; Ramachandran, P.; Foley, S.L. Global transcriptomic analyses of salmonella enterica in iron-depleted and iron-rich growth conditions. BMC Genom. 2019, 20, 490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porcheron, G.; Dozois, C.M. Interplay between iron homeostasis and virulence: Fur and ryhb as major regulators of bacterial pathogenicity. Vet. Microbiol. 2015, 179, 2–14. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.J.; Thorsness, J.L.; Anderson, C.P.; Lynne, A.M.; Foley, S.L.; Han, J.; Fricke, W.F.; McDermott, P.F.; White, D.G.; Khatri, M.; et al. Horizontal gene transfer of a colv plasmid has resulted in a dominant avian clonal type of salmonella enterica serovar kentucky. PLoS ONE 2010, 5, e15524. [Google Scholar] [CrossRef]
- Mnif, B.; Vimont, S.; Boyd, A.; Bourit, E.; Picard, B.; Branger, C.; Denamur, E.; Arlet, G. Molecular characterization of addiction systems of plasmids encoding extended-spectrum β-lactamases in escherichia coli. J. Antimicrob. Chemother. 2010, 65, 1599–1603. [Google Scholar] [CrossRef]
- Mnif, B.; Harhour, H.; Jdidi, J.; Mahjoubi, F.; Genel, N.; Arlet, G.; Hammami, A. Molecular epidemiology of extended-spectrum β-lactamase-producing escherichia coli in tunisia and characterization of their virulence factors and plasmid addiction systems. BMC Microbiol. 2013, 13, 147. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.J.; Johnson, S.J.; Nolan, L.K. Complete DNA sequence of a colbm plasmid from avian pathogenic escherichia coli suggests that it evolved from closely related colv virulence plasmids. J. Bacteriol. 2006, 188, 5975–5983. [Google Scholar] [CrossRef] [Green Version]
- Jacoby, G.A.; Han, P. Detection of extended-spectrum β-lactamases in clinical isolates of klebsiella pneumoniae and escherichia coli. J. Clin. Microbial. 1996, 34, 908–911. [Google Scholar] [CrossRef] [Green Version]
- Foley, S.L.; White, D.G.; McDermott, P.F.; Walker, R.D.; Rhodes, B.; Fedorka-Cray, P.J.; Simjee, S.; Zhao, S.H. Comparison of subtyping methods for differentiating salmonella enterica serovar typhimurium isolates obtained from food animal sources. J. Clin. Microbial. 2006, 44, 3569–3577. [Google Scholar] [CrossRef] [Green Version]
- Kaldhone, P.; Nayak, R.; Lynne, A.M.; David, D.E.; McDermott, P.F.; Logue, C.M.; Foley, S.L. Characterization of salmonella enterica serovar heidelberg from turkey-associated sources. Appl. Environ. Microbiol. 2008, 74, 5038–5046. [Google Scholar] [CrossRef] [Green Version]
- Lynne, A.M.; Dorsey, L.L.; David, D.E.; Foley, S.L. Characterisation of antibiotic resistance in host-adapted salmonella enterica. Int. J. Antimicrob. Agents 2009, 34, 169–172. [Google Scholar] [CrossRef]
- Lynne, A.M.; Rhodes-Clark, B.S.; Bliven, K.; Zhao, S.H.; Foley, S.L. Antimicrobial resistance genes associated with salmonella enterica serovar newport isolates from food animals. Antimicrob. Agents Chemother. 2008, 52, 353–356. [Google Scholar] [CrossRef] [Green Version]
- Gokulan, K.; Khare, S.; Rooney, A.W.; Han, J.; Lynne, A.M.; Foley, S.L. Impact of plasmids, including those encodingvirb4/d4 type iv secretion systems, on salmonella enterica serovar heidelberg virulence in macrophages and epithelial cells. PLoS ONE 2013, 8, e77866. [Google Scholar] [CrossRef] [Green Version]
- Welch, T.J.; Fricke, W.F.; McDermott, P.F.; White, D.G.; Rosso, M.L.; Rasko, D.A.; Mammel, M.K.; Eppinger, M.; Rosovitz, M.J.; Wagner, D.; et al. Multiple antimicrobial resistance in plague: An emerging public health risk. PLoS ONE 2007, 2, e309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, S.; Pettengill, J.; Lou, Y.; Payne, J.; Shpuntoff, A.; Rand, H.; Strain, E. Cfsan snp pipeline: An automated method for constructing snp matrices from next-generation sequence data. PeerJ Comput. Sci. 2015, 1, e20. [Google Scholar] [CrossRef] [Green Version]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Cech, M.; Chilton, J.; Clements, D.; Coraor, N.; Gruning, B.A.; et al. The galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. Raxml version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In silico detection and typing of plasmids using plasmidfinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Wattam, A.R.; Abraham, D.; Dalay, O.; Disz, T.L.; Driscoll, T.; Gabbard, J.L.; Gillespie, J.J.; Gough, R.; Hix, D.; Kenyon, R.; et al. Patric, the bacterial bioinformatics database and analysis resource. Nucleic Acids Res. 2014, 42, D581–D591. [Google Scholar] [CrossRef] [Green Version]
- Foley, S.L.; Lynne, A.M. Food animal-associated salmonella challenges: Pathogenicity and antimicrobial resistance. J. Anim. Sci. 2008, 86, E173–E187. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, L.; Wilcks, A.; Hammer, K.; Huys, G.; Gevers, D.; Andersen, S.R. Horizontal transfer of tet(m) and erm(b) resistance plasmids from food strains of lactobacillus plantarum to enterococcus faecalis jh2-2 in the gastrointestinal tract of gnotobiotic rats. FEMS Microbiol. Ecol. 2007, 59, 158–166. [Google Scholar] [CrossRef] [Green Version]
- Nedialkova, L.P.; Denzler, R.; Koeppel, M.B.; Diehl, M.; Ring, D.; Wille, T.; Gerlach, R.G.; Stecher, B. Inflammation fuels colicin ib-dependent competition of salmonella serovar typhimurium and e. Coli in enterobacterial blooms. PLoS Pathog. 2014, 10, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stecher, B.; Denzler, R.; Maier, L.; Bernet, F.; Sanders, M.J.; Pickard, D.J.; Barthel, M.; Westendorf, A.M.; Krogfelt, K.A.; Walker, A.W.; et al. Gut inflammation can boost horizontal gene transfer between pathogenic and commensal enterobacteriaceae. Proc. Natl. Acad. Sci. USA 2012, 109, 1269–1274. [Google Scholar] [CrossRef] [Green Version]
- Konisky, J. Characterization of colicin ia and colicin ib—Chemical studies of protein structure. J. Biol. Chem. 1972, 247, 3750–3755. [Google Scholar]
- Ernst, F.D.; Bereswill, S.; Waidner, B.; Stoof, J.; Mader, U.; Kusters, J.G.; Kuipers, E.J.; Kist, M.; van Vliet, A.H.M.; Homuth, G. Transcriptional profiling of helicobacter pylori fur- and iron-regulated gene expression. Microbiolog 2005, 151, 533–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reissbrodt, R.; Kingsley, R.; Rabsch, W.; Beer, W.; Roberts, M.; Williams, P.H. Iron-regulated excretion of α-keto acids by salmonella typhimurium. J. Bacteriol. 1997, 179, 4538–4544. [Google Scholar] [CrossRef] [Green Version]
- Baumler, A.J.; Tsolis, R.M.; vanderVelden, A.W.M.; Stojiljkovic, I.; Anic, S.; Heffron, F. Identification of a new iron regulated locus of salmonella typhi. Gene 1996, 183, 207–213. [Google Scholar] [CrossRef]
- Marchetti, M.; Bei, O.D.; Bettati, S.; Campanini, B.; Kovachka, S.; Gianquinto, E.; Spyrakis, F.; Ronda, L. Iron Metabolism at the Interface between Host and Pathogen: From Nutritional Immunity to Antibacterial Development. Int. J. Mol. Sci. 2020, 21, 2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lostroh, C.P.; Lee, C.A. The salmonella pathogenicity island-1 type iii secretion system. Microb. Infect. 2001, 3, 1281–1291. [Google Scholar] [CrossRef]
- Knodler, L.A.; Nair, V.; Steele-Mortimer, O. Quantitative assessment of cytosolic salmonella in epithelial cells. PLoS ONE 2014, 9, e84681. [Google Scholar] [CrossRef]
- Zhang, K.Y.; Griffiths, G.; Repnik, U.; Hornef, M. Seeing is understanding: Salmonella’s way to penetrate the intestinal epithelium. Int. J. Med. Microbiol. 2018, 308, 97–106. [Google Scholar] [CrossRef]
- Knodler, L.A.; Vallance, B.A.; Celli, J.; Winfree, S.; Hansen, B.; Montero, M.; Steele-Mortimer, O. Dissemination of invasive salmonella via bacterial-induced extrusion of mucosal epithelia. Proc. Natl. Acad. Sci. USA 2010, 107, 17733–17738. [Google Scholar] [CrossRef] [Green Version]
- Wrande, M.; Andrews-Polymenis, H.; Twedt, D.J.; Steele-Mortimer, O.; Porwollik, S.; McClelland, M.; Knodler, L.A. Genetic determinants of salmonella enterica serovar typhimurium proliferation in the cytosol of epithelial cells. Infect. Immun. 2016, 84, 3517–3526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salcedo, S.P.; Noursadeghi, M.; Cohen, J.; Holden, D.W. Intracellular replication of salmonella typhimurium strains in specific subsets of splenic macrophages in vivo. Cell Microbiol. 2001, 3, 587–597. [Google Scholar] [CrossRef]
- Beuzon, C.R.; Salcedo, S.P.; Holden, D.W. Growth and killing of a salmonella enterica serovar typhimurium sifa mutant strain in the cytosol of different host cell lines. Microbiology 2002, 148, 2705–2715. [Google Scholar] [CrossRef] [Green Version]
- Van Essen-Zandbergen, A.; Smith, H.; Veldman, K.; Mevius, D. In vivo transfer of an incfib plasmid harbouring a class 1 integron with gene cassettes dfra1-aada1. Vet. Microbiol. 2009, 137, 402–407. [Google Scholar] [CrossRef]
- Card, R.M.; Cawthraw, S.A.; Nunez-Garcia, J.; Ellis, R.J.; Kay, G.; Pallen, M.J.; Woodward, M.J.; Anjum, M.F. An in vitro chicken gut model demonstrates transfer of a multidrug resistance plasmid from salmonella to commensal escherichia coli. mBio 2017, 8, 15. [Google Scholar] [CrossRef] [Green Version]
- Faure, S.; Perrin-Guyomard, A.; Delmas, J.M.; Laurentie, M. Impact of therapeutic treatment with β-lactam on transfer of the bla(ctx-m-9) resistance gene from salmonella enterica serovar virchow to escherichia coli in gnotobiotic rats. Appl. Environ. Microbiol. 2009, 75, 5523–5528. [Google Scholar] [CrossRef] [Green Version]
- El-Halfawy, O.M.; Valvano, M.A. Non-genetic mechanisms communicating antibiotic resistance: Rethinking strategies for antimicrobial drug design. Expert Opin. Drug Discov. 2012, 7, 923–933. [Google Scholar] [CrossRef] [PubMed]
- Han, T.H.; Lee, J.H.; Cho, M.H.; Wood, T.K.; Lee, J. Environmental factors affecting indole production in escherichia coli. Res. Microbiol. 2011, 162, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Hirakawa, H.; Inazumi, Y.; Masaki, T.; Hirata, T.; Yamaguchi, A. Indole induces the expression of multidrug exporter genes in escherichia coli. Mol. Microbiol. 2005, 55, 1113–1126. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Molla, M.N.; Cantor, C.R.; Collins, J.J. Bacterial charity work leads to population-wide resistance. Nature 2010, 467, 82–85. [Google Scholar] [CrossRef] [Green Version]
- Nikaido, H. Preventing drug access to targets: Cell surface permeability barriers and active efflux in bacteria. Semin. Cell Dev. Biol. 2001, 12, 215–223. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Source | Isolation Location | Year | GenBank Accession |
|---|---|---|---|---|
| 163 | Turkey | OH | 1999 | LSZD00000000 |
| 368 | Cattle | GA | 1999 | PDNO00000000 |
| 373 | Cattle | MI | 1999 | PDNN00000000 |
| 374 | Cattle | MI | 1999 | VTSM00000000 |
| 375 | Cattle | MI | 1999 | VTSL00000000 |
| 376 | Chicken | GA | 1999 | PDNM00000000 |
| 391 | Chicken | USA | 1999 | VTSK00000000 |
| 393 | Swine | USA | 1999 | VSXW00000000 |
| 397 | Chicken | USA | 1999 | LYRR00000000 |
| 411 | Chicken | USA | 1999 | VSXV00000000 |
| 417 | Chicken | USA | 1999 | VSXU00000000 |
| 419 | Swine | USA | 1999 | VSXT00000000 |
| 420 | Chicken | USA | 1999 | VSXS00000000 |
| 421 | Chicken | USA | 1999 | VSXI00000000 |
| 423 | Chicken | USA | 1999 | VSXR00000000 |
| 426 | Turkey | USA | 1999 | PDNK00000000 |
| 427 | Chicken | USA | 1999 | VSXQ00000000 |
| 443 | Turkey | USA | 1999 | VSXP00000000 |
| 447 | Turkey | USA | 1999 | VSXO00000000 |
| 448 | Chicken | USA | 1999 | VSXN00000000 |
| 449 | Chicken | USA | 1999 | VSXM00000000 |
| 452 | Turkey | USA | 1999 | LYRS00000000 |
| 455 | Chicken | USA | 1999 | VSXL00000000 |
| 457 | Turkey | USA | 1999 | VSXH00000000 |
| 458 | Turkey | USA | 1999 | VSXK00000000 |
| 459 | Chicken | USA | 1999 | VTSH00000000 |
| 460 | Turkey | USA | 1999 | VTSI00000000 |
| 462 | Turkey | USA | 1999 | VTSU00000000 |
| 463 | Swine | USA | 1999 | PHGW00000000 |
| 464 | Turkey | USA | 1999 | VSXG00000000 |
| 465 | Turkey | USA | 1999 | VTSG00000000 |
| 475 | Swine | USA | 1999 | VSXJ00000000 |
| 477 | Turkey | USA | 1999 | VTSF00000000 |
| 478 | Turkey | USA | 1999 | LYRT00000000 |
| 481 | Turkey | USA | 1999 | VSXF00000000 |
| 483 | Turkey | USA | 1999 | VTSD00000000 |
| 484 | Turkey | USA | 1999 | PDOI00000000 |
| 485 | Turkey | USA | 1999 | VSXE00000000 |
| 486 | Turkey | USA | 1999 | VTSC00000000 |
| 487 | Turkey | USA | 1999 | VSXD00000000 |
| 489 | Turkey | USA | 1999 | VTSB00000000 |
| 492 | Cattle | USA | 1999 | PDOJ00000000 |
| 494 | Cattle | USA | 1999 | PDOK00000000 |
| 495 | Cattle | USA | 1999 | VTSA00000000 |
| 498 | Cattle | USA | 1999 | VSXC00000000 |
| 499 | Cattle | WV | 1999 | VTRZ00000000 |
| 500 | Cattle | USA | 1999 | VSXB00000000 |
| 696 | Turkey | USA | 1999 | LXHA00000000 |
| 710 | Turkey | ND | 1999 | LXGZ00000000 |
| N028 | Chicken | WV | 2003 | VSWY00000000 |
| N029 | Chicken | WV | 2003 | VSXA00000000 |
| N030 | Chicken | WV | 2003 | VSWX00000000 |
| N032 | Chicken | WV | 2003 | VSWZ00000000 |
| N033 | Chicken | WV | 2003 | VTRY00000000 |
| N034 | Chicken | WV | 2003 | VSWW00000000 |
| N035 | Chicken | WV | 2003 | VSWV00000000 |
| N061 | Chicken | WV | 2003 | VSWU00000000 |
| N062 | Chicken | WV | 2003 | VTRX00000000 |
| N063 | Chicken | WV | 2003 | VSWT00000000 |
| N065 | Chicken | WV | 2003 | VSWS00000000 |
| N066 | Chicken | WV | 2003 | VSWR00000000 |
| N067 | Chicken | WV | 2003 | VSWQ00000000 |
| N068 | Chicken | WV | 2003 | VSWP00000000 |
| N069 | Chicken | WV | 2003 | VSWO00000000 |
| N070 | Chicken | WV | 2003 | VSWN00000000 |
| N071 | Chicken | WV | 2003 | VSWM00000000 |
| N073 | Chicken | WV | 2003 | VTRW00000000 |
| N075 | Chicken | WV | 2003 | VSWL00000000 |
| N135 | Poultry Water | WV | 2003 | VSWK00000000 |
| N138 | Poultry Water | WV | 2003 | VSWJ00000000 |
| N140 | Chicken | WV | 2003 | VSWI00000000 |
| Isolate | Plasmid Transfer-Associated Genes | Conjugation | Colicin Genes | Colicin Inhibition | ||||
|---|---|---|---|---|---|---|---|---|
| Type IV Pilus Biogenesis Protein | Tra Genes | Colicin Ia Synthesis Protein | Colicin Ib Protein | Colicin V Synthesis Protein | Colicin Ib Immunity Protein | |||
| 163 | pilJPM | traTJ | YES | cia | cib | N | imm | NO |
| 368 | N | traT | NO | N | N | N | N | NO |
| 373 | N | traT | NO | N | N | N | N | NO |
| 374 | pilIJPM | traTJ | NO | N | N | N | N | NO |
| 375 | pilIJPM | traTJ | YES | N | N | N | N | NO |
| 376 | N | traT | NO | N | N | N | N | NO |
| 391 | N | traT | NO | N | N | cvaABC | N | NO |
| 393 | pilIJPM | traTJ | YES | N | N | N | N | NO |
| 397 | pilJPM | traJ | NO | cia | cib | N | N | NO |
| 411 | pilJPM | traTJ | NO | cia | cib | cvaBC | imm | YES |
| 417 | pilJPM | traTJ | NO | cia | cib | N | imm | NO |
| 419 | N | N | NO | N | N | N | N | NO |
| 420 | pilJPM | traTJ | NO | cia | cib | cvaBC | imm | YES |
| 421 | pilJPM | traTJ | NO | cia | cib | cvaABC | imm | YES |
| 423 | pilJPM | traTJ | NO | cia | cib | cvaABC | imm | YES |
| 426 | pilPM | traTJ | NO | N | N | cvaABC | N | YES |
| 427 | N | traT | NO | N | N | cvaABC | N | NO |
| 443 | N | traT | YES | N | N | N | N | NO |
| 447 | N | traT | YES | N | N | N | N | NO |
| 448 | N | traT | NO | N | N | N | N | NO |
| 449 | N | traT | NO | N | N | N | N | NO |
| 452 | N | traT | YES | N | N | N | N | NO |
| 455 | N | traT | NO | N | N | N | N | NO |
| 457 | N | traTJ | YES | N | N | N | N | NO |
| 458 | pilIJPM | traTJ | YES | N | N | N | N | NO |
| 459 | pilJPM | traTJ | YES | cia | cib | cvaABC | imm | YES |
| 460 | N | traT | YES | N | N | N | N | NO |
| 462 | N | traT | YES | N | N | N | N | NO |
| 463 | pilIJPM | traTJ | YES | N | N | N | N | NO |
| 464 | N | traT | YES | N | N | N | N | NO |
| 465 | N | traT | YES | N | N | N | N | NO |
| 475 | N | N | NO | N | N | N | N | NO |
| 477 | N | traT | YES | N | N | N | N | NO |
| 478 | N | traT | YES | N | N | N | N | NO |
| 481 | N | traT | NO | N | N | N | N | NO |
| 483 | N | traT | NO | N | N | N | N | NO |
| 484 | N | traT | NO | N | N | N | N | NO |
| 485 | N | traT | YES | N | N | N | N | NO |
| 486 | N | traT | YES | N | N | N | N | NO |
| 487 | N | traT | NO | N | N | N | N | NO |
| 489 | N | traT | NO | N | N | N | N | NO |
| 492 | N | traT | NO | N | N | N | N | NO |
| 494 | N | traT | YES | N | N | N | N | NO |
| 495 | N | traT | NO | N | N | N | N | NO |
| 498 | pilJPM | traTJ | NO | cia | cib | N | N | NO |
| 499 | N | traT | NO | N | N | N | N | NO |
| 500 | N | traT | NO | N | N | N | N | NO |
| 696 | N | traT | NO | N | N | N | N | NO |
| 710 | N | traT | YES | N | N | N | N | NO |
| N028 | N | traTJ | NO | cia | cib | cvaAB | imm | YES |
| N029 | N | traTJ | YES | cia | cib | cvaAB | imm | YES |
| N030 | pilJPM | traTJ | YES | cia | cib | cvaAB | imm | YES |
| N032 | N | traTJ | NO | cia | cib | cvaAB | imm | YES |
| N033 | N | traTJ | NO | cia | cib | cvaAB | imm | YES |
| N034 | pilJPM | traTJ | YES | cia | cib | cvaAB | imm | YES |
| N035 | N | traTJ | NO | cia | cib | cvaAB | imm | YES |
| N061 | pilJPM | traTJ | NO | cia | cib | N | imm | NO |
| N062 | pilJPM | traTJ | NO | cia | cib | N | imm | NO |
| N063 | pilJPM | traTJ | NO | cia | cib | N | imm | NO |
| N065 | pilJPM | traTJ | NO | cia | cib | N | imm | YES |
| N066 | pilJPM | traTJ | NO | cia | cib | N | imm | NO |
| N067 | pilJPM | traTJ | NO | cia | cib | N | imm | NO |
| N068 | pilJPM | traTJ | NO | cia | cib | N | imm | YES |
| N069 | pilJPM | traTJ | YES | cia | cib | cvaABC | imm | YES |
| N070 | pilJPM | traTJ | NO | cia | cib | N | imm | NO |
| N071 | pilJPM | traTJ | NO | cia | cib | N | imm | YES |
| N073 | pilJPM | traTJ | NO | cia | cib | N | imm | YES |
| N075 | pilJPM | traTJ | NO | cia | cib | N | imm | NO |
| N135 | pilJPM | traTJ | NO | cia | cib | cvaABC | imm | YES |
| N138 | pilJPM | traTJ | NO | cia | cib | cvaABC | imm | YES |
| N140 | pilJPM | traTJ | NO | cia | cib | cvaABC | imm | YES |
| Donor | PlasmidFinder | ResFinder (Resistance Genes) | Transconjugant | PlasmidFinder | ResFinder (Resistance Genes) |
|---|---|---|---|---|---|
| 163 | IncFIB, IncFIA, IncA/C2, IncFII(pCoo), IncX4, IncI1 | blaCMY-2, blaTEM-1B, sul1, sul2, aac(3)-VIa, aadA1, ant(2″)-Ia, aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, cmlA1, floR, tet(A), tet(B) | X163 | IncI1, IncX4 | sul1, aac(3)-VIa, aadA1 |
| 375 | IncFIB, ColpVC, IncA/C2, IncFII(S), IncI1 | sul1, sul2, aac(3)-VIa, aadA12, aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, tet(A), blaCMY-2, floR | X375 | IncA/C2 | sul2, blaCMY-2, tet(A), aph(3″)-Ib, aph(6)-Id |
| 393 | IncFIB, ColpVC, IncFIA, IncFII, IncI1 | blaTEM-1B, blaTEM, aph(3′)-Ia, aadA12, tet(A), sul1 | X393 | IncFIB, IncFIA, IncFII | tet(A), sul1, aadA12, aph(3′)-Ia |
| 443 | IncFIB, IncA/C2, IncFIA, IncFII(pCoo) | ant(2″)-Ia, aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, blaCMY-2, blaTEM-1B, cmlA1, floR, tet(A), tet(B), sul2 | X443 | IncFIB, IncFIA, IncFII(pcoo) | aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, sul2, tet(B) |
| 447 | IncFIB, ColpVC, IncA/C2, IncFIA, IncFII(pCoo) | aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, blaCMY-2, floR, tet(A), tet(B), sul2 | X447 | IncFIB, IncFIA, IncFII(pcoo) | tet(B), sul2, aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id |
| 452 | IncFIB, ColpVC, IncFIA, IncFII(pCoo), IncA/C2 | blaCMY-2, blaTEM, tet(A), tet(B), sul2, ant(2″)-Ia, aph(3″)-Ib, aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, cmlA1, floR | X452 | IncFIB, IncFIA, IncFII(pcoo) | aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, sul2, tet(B) |
| 457 | IncFIB, ColpVC, IncFIA, IncFII(pCoo) | aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, blaCMY-2, floR, tet(A), tet(B), sul2 | X457 | IncFIB, IncFIA, IncFII(pcoo) | sul2, aph(3″)-Ib, tet(B) |
| 458 | IncFIB, ColpVC, IncFIA, IncFII, IncI1 | blaTEM-1B | X458 | IncFIB, IncFIA, IncFII | blaTEM-1B |
| 459 | IncFIB, Col156, IncFIC(FII), IncI1 | blaCMY-2 | X459 | IncI1 | blaCMY-2 |
| 460 | IncFIB, ColpVC, IncA/C2, IncFIA, IncFII(pCoo) | aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, blaCMY-2, floR, tet(A), tet(B), sul2 | X460 | IncFIB, IncFIA, IncFII(pcoo) | aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, sul2, tet(B) |
| 462 | IncFIB, IncA/C2, IncFIA, IncFII(pCoo) | blaCMY-2, blaTEM-1B, sul2, tet(A), tet(B), cmlA1, floR, ant(2″)-Ia, aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id | X462 | IncFIB, IncFIA, IncFII(pcoo) | sul2, aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, tet(B) |
| 463 | IncFIB, ColpVC, IncFII, IncI1, Col156, IncFIA | sul1, aadA12, aph(3′)-Ia, tet(A), blaTEM-1B | X463 | IncFIB, IncFIA, IncFII | aadA12, aph(3′)-Ia, blaTEM-1B, sul1, tet(A) |
| 464 | IncFIB, ColpVC, IncA/C2, IncFIA, IncFII(pCoo) | ant(2″)-Ia, aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, blaCMY-2, blaTEM-1B, cmlA1, floR, tet(A), tet(B), sul2 | X464 | IncFIB, IncFIA, IncFII(pcoo) | aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, sul2, tet(B) |
| 465 | IncFIB, IncA/C2, IncFIA, IncFII(pCoo) | aac(6′)-Iaa, aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, blaCMY-2, floR, tet(A), tet(B), sul2 | X465 | IncFIB, IncFIA, IncFII(pcoo) | aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, sul2, tet(B) |
| 477 | IncFIB, ColpVC, IncA/C2, IncFIA, IncFII(pCoo) | aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, blaCMY-2, floR, tet(A), tet(B), sul2 | X477 | IncFIB, IncFIA, IncFII(pcoo) | aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, sul2 |
| 478 | IncFIB, IncA/C2, IncFIA, IncFII(pCoo) | aadA1, aadA12, aadA21, aadA22, aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, blaCMY-2, blaTEM-1B, cmlA1, floR, tet(A), tet(B), sul2 | X478 | IncFIB, IncFIA, IncFII(pcoo), IncA/C2 | cmlA1, floR, tet(A), tet(B), sul2, aph(3″)-Ib, aph(6)-Id, blaCMY-2, blaTEM-1B |
| 485 | IncFIB, IncA/C2, IncFIA, IncFII(pCoo) | ant(2″)-Ia, aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, blaCMY-2, blaTEM-1B, cmlA1, floR, tet(A), tet(B), sul2 | X485 | IncFIB, IncFIA, IncFII(pcoo) | tet(B), sul2, aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id |
| 486 | IncFIB, IncA/C2, IncFIA, IncFII(pCoo) | ant(2″)-Ia, aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, blaCMY-2, blaTEM-1B, cmlA1, floR, tet(A), tet(B), sul2 | X486 | IncFIB, IncFIA, IncFII(pcoo)) | aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, sul2, tet(B) |
| 494 | IncFIB, IncFII(S), IncA/C2 | blaCMY-2, blaTEM-1B, sul2, sul1 tet(A), floR, ant(2″)-Ia, aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id | X494 | IncA/C2 | blaCMY-2, tet(A), floR, aph(3″)-Ib, aph(6)-Id, sul2 |
| 710 | IncFIB, IncFIA, IncA/C2, IncFII(pCoo), IncX4 | ant(2″)-Ia, aph(3″)-Ib, aph(6)-Id, cmlA1, floR, tet(A), tet(B), sul2, blaCMY-2, blaTEM-1B | X710 | IncFIB, IncFIA, IncFII(pcoo) | tet(B), aph(3″)-Ib, aph(3′)-Ia, aph(6)-Id, sul2 |
| N029 | IncFIB, ColpVC, IncFIC(FII), IncHI2, IncHI2A | aph(3″)-Ib, aph(6)-Id, tet(B), blaCMY-2 | XN029 | IncFIB | aph(3″)-Ib, aph(6)-Id, tet(B) |
| N030 | IncFIB, ColpVC, IncFIC(FII), IncI1 | aph(3″)-Ib, blaCMY-2 | XN030 | ColpVC, IncI1 | aph(3″)-Ib, blaCMY-2 |
| N034 | IncFIB, ColpVC, IncFIC(FII), IncI1 | aph(3″)-Ib, blaCMY-2 | XN034 | ColpVC, IncI1 | aph(3″)-Ib, blaCMY-2 |
| N069 | IncFIB, ColpVC, IncFIC(FII), IncI1 | aadA1, sul1 | XN069 | IncI1 | aac(3)-VIa, aadA1, sul1 |
| Donor | Iron Acquisition Genes | Salmonella Plasmid Virulence (spv) | Transconjugant | Iron Acquisition Genes | Salmonella Plasmid Virulence (spv) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| iuc | iut | iro | sit | iuc | iut | iro | sit | ||||
| 163 | iucABCD | iutA | iroNB | sitA | N | X163 | N | N | N | N | N |
| 375 | N | iutA | iroNB | sitA | spv | X375 | N | N | N | N | N |
| 393 | N | iutA | iroNBC | sitA | N | X393 | N | iutA | N | N | N |
| 443 | iucABCD | iutA | iroNB | sitAB | N | X443 | iucABCD | iutA | N | sitABCD | N |
| 447 | iucABCD | iutA | iroNB | sitAB | N | X447 | iucABCD | iutA | N | sitABCD | N |
| 452 | iucABCD | iutA | iroNB | sitAB | N | X452 | iucABCD | iutA | N | sitABCD | N |
| 457 | iucABCD | iutA | iroNB | sitAB | N | X457 | iucABCD | iutA | N | sitABCD | N |
| 458 | N | iutA | iroNB | sitA | N | X458 | N | iutA | N | N | N |
| 459 | iucABCD | iutA | iroNBCD | sitAB | N | X459 | N | N | N | N | N |
| 460 | iucABCD | iutA | iroNB | sitA | N | X460 | iucABCD | N | N | sitABCD | N |
| 462 | iucABCD | iutA | iroNB | sitAB | N | X462 | iucABCD | iutA | N | sitABCD | N |
| 463 | N | iutA | iroNB | sitA | N | X463 | N | iutA | N | N | N |
| 464 | iucABCD | iutA | iroNB | sitAB | N | X464 | iucABCD | iutA | N | sitABCD | N |
| 465 | iucABCD | iutA | iroNB | sitAB | N | X465 | iucABCD | iutA | N | sitABCD | N |
| 477 | iucABCD | iutA | iroNB | sitAB | N | X477 | iucABCD | iutA | N | sitABCD | N |
| 478 | iucABCD | iutA | iroNB | sitAB | N | X478 | iucABCD | iutA | N | sitABCD | N |
| 485 | iucABCD | iutA | iroNB | sitAB | N | X485 | iucABCD | iutA | N | sitABCD | N |
| 486 | iucABCD | iutA | iroNB | sitAB | N | X486 | iucABCD | iutA | N | sitABCD | N |
| 494 | N | N | iroNB | sitA | spv | X494 | N | N | N | N | N |
| 710 | iucABCD | iutA | iroNB | sitAB | N | X710 | iucABCD | iutA | N | sitABCD | N |
| N029 | iucABCD | iutA | iroNBCD | sitAB | N | XN029 | iucACD | iutA | N | N | N |
| N030 | iucABCD | iutA | iroNBCD | sitAB | N | XN030 | N | N | N | N | N |
| N034 | iucABCD | iutA | iroNBCD | sitAB | N | XN034 | N | N | N | N | N |
| N069 | iucABCD | iutA | iroNBCD | sitAB | N | XN069 | N | N | N | N | N |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aljahdali, N.H.; Khajanchi, B.K.; Weston, K.; Deck, J.; Cox, J.; Singh, R.; Gilbert, J.; Sanad, Y.M.; Han, J.; Nayak, R.; et al. Genotypic and Phenotypic Characterization of Incompatibility Group FIB Positive Salmonella enterica Serovar Typhimurium Isolates from Food Animal Sources. Genes 2020, 11, 1307. https://doi.org/10.3390/genes11111307
Aljahdali NH, Khajanchi BK, Weston K, Deck J, Cox J, Singh R, Gilbert J, Sanad YM, Han J, Nayak R, et al. Genotypic and Phenotypic Characterization of Incompatibility Group FIB Positive Salmonella enterica Serovar Typhimurium Isolates from Food Animal Sources. Genes. 2020; 11(11):1307. https://doi.org/10.3390/genes11111307
Chicago/Turabian StyleAljahdali, Nesreen H., Bijay K. Khajanchi, Kennedi Weston, Joanna Deck, Justin Cox, Ruby Singh, Jeffrey Gilbert, Yasser M. Sanad, Jing Han, Rajesh Nayak, and et al. 2020. "Genotypic and Phenotypic Characterization of Incompatibility Group FIB Positive Salmonella enterica Serovar Typhimurium Isolates from Food Animal Sources" Genes 11, no. 11: 1307. https://doi.org/10.3390/genes11111307