Was the Cinta Senese Pig Already a Luxury Food in the Late Middle Ages? Ancient DNA and Archaeozoological Evidence from Central Italy

 , ,
, ,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Archeological Excavation
2.2. Archaeozoological Materials and Methods
2.3. Sample Selection
2.4. DNA Extraction
2.5. Amplification and Sequencing
2.6. Genotyping with PCR-RFLP
3. Results
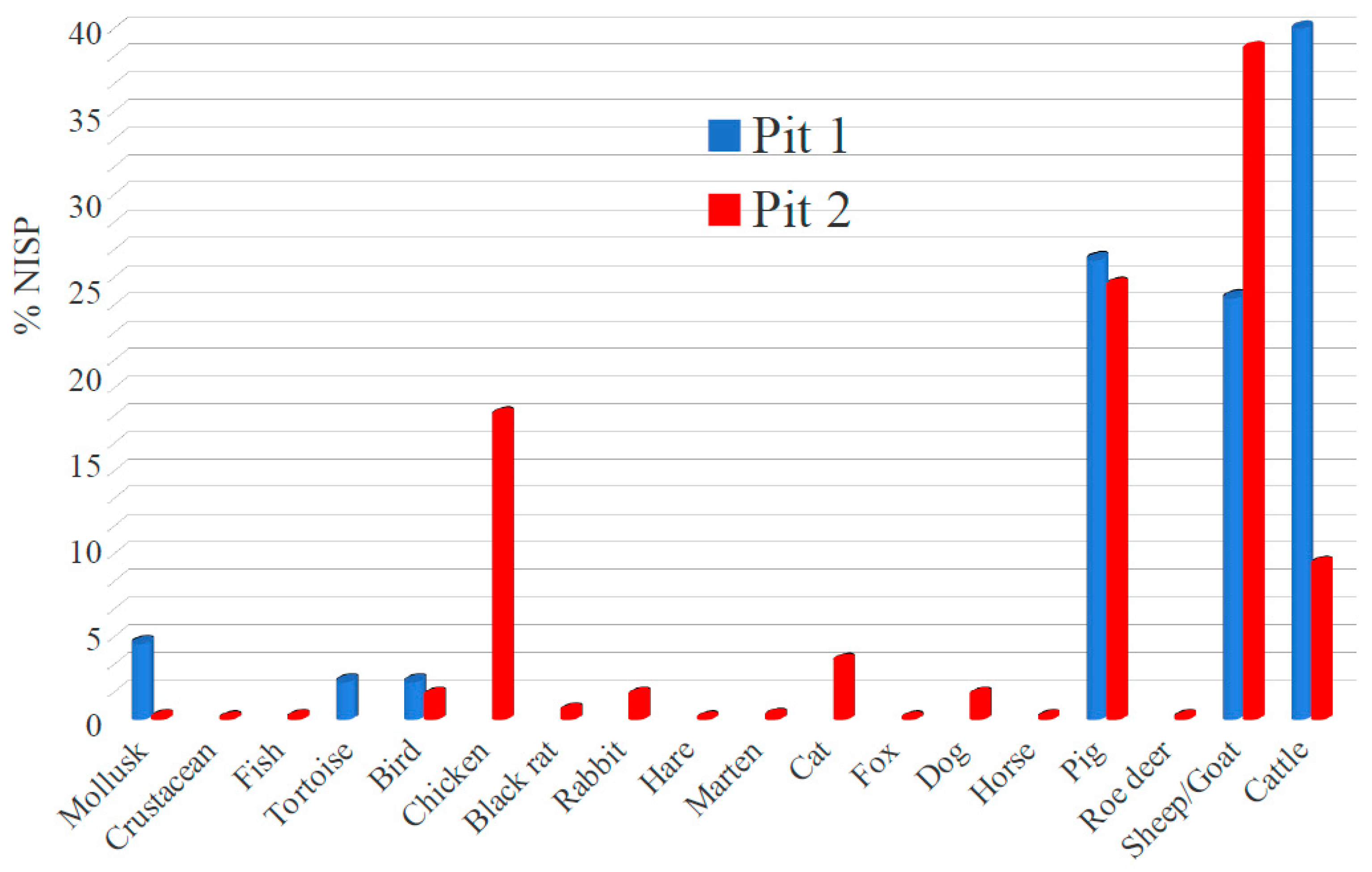
3.1. Archaeozoological Analysis
3.2. Genetic Analysis
3.2.1. Assessing the Presence of Cinta Senese Breed
3.2.2. Detection of Possible Cross-Breeding
3.2.3. Sexing of the Bones
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Larson, G.; Piperno, D.R.; Allaby, R.G.; Purugganan, M.D.; Andersson, L.; Arroyo-Kalin, M.; Barton, L.; Climer Vigueira, C.; Denham, T.; Dobney, K.; et al. Current perspectives and the future of domestication studies. Proc. Natl. Acad. Sci. USA 2014, 111, 6139–6146. [Google Scholar] [CrossRef]
- Larson, G.; Burger, J. A population genetics view of animal domestication. Trends Genet. 2013, 29, 197–205. [Google Scholar] [CrossRef]
- Caliebe, A.; Nebel, A.; Makarewicz, C.; Krawczak, M.; Krause-Kyora, B. Insights into early pig domestication provided by ancient DNA analysis. Sci. Rep. 2017, 7, 44550. [Google Scholar] [CrossRef]
- Zeder, M.A. Domestication and early agriculture in the Mediterranean Basin: Origins, diffusion, and impact. Proc. Natl. Acad. Sci. USA 2008, 105, 11597–11604. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, M.R.; Chen, W.; Lenstra, J.A.; Goderie, C.R.J.; MacHugh, D.E.; Park, S.D.E.; Magee, D.A.; Matassino, D.; Ciani, F.; Megens, H.-J.; et al. Data from: Genetic Origin, Admixture and Population History of Aurochs (Bos Primigenius) and Primitive European Cattle. Heredity 2016, 118, 169–176. [Google Scholar]
- Wang, G.-D.; Peng, M.-S.; Yang, H.-C.; Savolainen, P.; Zhang, Y.-P. Questioning the evidence for a Central Asian domestication origin of dogs. Proc. Natl. Acad. Sci. USA 2016, 113, E2554–E2555. [Google Scholar] [CrossRef]
- Amills, M.; Capote, J.; Tosser-Klopp, G. Goat domestication and breeding: A jigsaw of historical, biological and molecular data with missing pieces. Anim. Genet. 2017, 48, 631–644. [Google Scholar] [CrossRef]
- MacHugh, D.E.; Larson, G.; Orlando, L. Taming the Past: Ancient DNA and the Study of Animal Domestication. Annu. Rev. Anim. Biosci. 2017, 5, 329–351. [Google Scholar] [CrossRef]
- Conolly, J.; Colledge, S.; Dobney, K.; Vigne, J.-D.; Peters, J.; Stopp, B.; Manning, K.; Shennan, S. Meta-analysis of zooarchaeological data from SW Asia and SE Europe provides insight into the origins and spread of animal husbandry. J. Archaeol. Sci. 2011, 38, 538–545. [Google Scholar] [CrossRef]
- Larson, G. Worldwide Phylogeography of Wild Boar Reveals Multiple Centers of Pig Domestication. Science 2005, 307, 1618–1621. [Google Scholar] [CrossRef]
- McHugo, G.P.; Dover, M.J.; MacHugh, D.E. Unlocking the origins and biology of domestic animals using ancient DNA and paleogenomics. BMC Biol. 2019, 17, 98. [Google Scholar] [CrossRef]
- Ottoni, C.; Girdland Flink, L.; Evin, A.; Geörg, C.; De Cupere, B.; Van Neer, W.; Bartosiewicz, L.; Linderholm, A.; Barnett, R.; Peters, J.; et al. Pig Domestication and Human-Mediated Dispersal in Western Eurasia Revealed through Ancient DNA and Geometric Morphometrics. Mol. Biol. Evol. 2013, 30, 824–832. [Google Scholar] [CrossRef]
- Frantz, L.A.F.; Haile, J.; Lin, A.T.; Scheu, A.; Geörg, C.; Benecke, N.; Alexander, M.; Linderholm, A.; Mullin, V.E.; Daly, K.G.; et al. Ancient pigs reveal a near-complete genomic turnover following their introduction to Europe. Proc. Natl. Acad. Sci. USA 2019, 116, 17231–17238. [Google Scholar] [CrossRef]
- Albarella, U.; Dobney, K.; Rowley-Conwy, P. Size and shape of the Eurasian wild boar (Sus scrofa), with a view to the reconstruction of its Holocene history. Environ. Archaeol. 2009, 14, 103–136. [Google Scholar] [CrossRef]
- Erwynck, A.; Lentacker, A.; Muldner, G.; Richards, M.; Dobney, K. An investigation into the transition from forest dwelling pigs to farm animals in Medieval Flanders. In Pigs and Humans: 10,000 Years of Interaction; Albarella, U., Dobney, K.K., Erwynck, A., Eds.; Ofxord University Press: Oxford, UK, 2007; pp. 171–196. [Google Scholar]
- White, S. From Globalized Pig Breeds to Capitalist Pigs: A Study in Animal Cultures and Evolutionary History. Environ. Hist. 2011, 16, 94–120. [Google Scholar] [CrossRef]
- Giuffra, E.; Kijas, J.M.; Amarger, V.; Carlborg, O.; Jeon, J.T.; Andersson, L. The origin of the domestic pig: Independent domestication and subsequent introgression. Genetics 2000, 154, 1785–1791. [Google Scholar]
- Felius, M.; Theunissen, B.; Lenstra, J.A. Conservation of cattle genetic resources: The role of breeds. J. Agric. Sci. 2015, 153, 152–162. [Google Scholar] [CrossRef]
- Bosse, M. A Genomics Perspective on Pig Domestication. In Animal Domestication; Teletchea, F., Ed.; IntechOpen: London, UK, 2019; ISBN 978-1-83880-174-8. [Google Scholar]
- van Asch, B.; Pereira, F.; Santos, L.S.; Carneiro, J.; Santos, N.; Amorim, A. Mitochondrial lineages reveal intense gene flow between Iberian wild boars and South Iberian pig breeds: mtDNA lineages of Iberian wild boars and pigs. Anim. Genet. 2012, 43, 35–41. [Google Scholar] [CrossRef]
- Amaral, A.J.; Ferretti, L.; Megens, H.-J.; Crooijmans, R.P.M.A.; Nie, H.; Ramos-Onsins, S.E.; Perez-Enciso, M.; Schook, L.B.; Groenen, M.A.M. Genome-Wide Footprints of Pig Domestication and Selection Revealed through Massive Parallel Sequencing of Pooled DNA. PLoS ONE 2011, 6, e14782. [Google Scholar] [CrossRef]
- Larson, G.; Liu, R.; Zhao, X.; Yuan, J.; Fuller, D.; Barton, L.; Dobney, K.; Fan, Q.; Gu, Z.; Liu, X.-H.; et al. Patterns of East Asian pig domestication, migration, and turnover revealed by modern and ancient DNA. Proc. Natl. Acad. Sci. USA 2010, 107, 7686–7691. [Google Scholar] [CrossRef]
- Groeneveld, L.F.; Lenstra, J.A.; Eding, H.; Toro, M.A.; Scherf, B.; Pilling, D.; Negrini, R.; Finlay, E.K.; Jianlin, H.; Groeneveld, E.; et al. Genetic diversity in farm animals—A review. Anim. Genet. 2010, 41, 6–31. [Google Scholar] [CrossRef]
- Orozco-terWengel, P.; Barbato, M.; Nicolazzi, E.; Biscarini, F.; Milanesi, M.; Davies, W.; Williams, D.; Stella, A.; Ajmone-Marsan, P.; Bruford, M.W. Revisiting demographic processes in cattle with genome-wide population genetic analysis. Front. Genet. 2015, 6, 191. [Google Scholar] [CrossRef]
- Pellecchia, M.; Negrini, R.; Colli, L.; Patrini, M.; Milanesi, E.; Achilli, A.; Bertorelle, G.; Cavalli-Sforza, L.L.; Piazza, A.; Torroni, A.; et al. The mystery of Etruscan origins: Novel clues from Bos taurus mitochondrial DNA. Proc. R. Soc. B Biol. Sci. 2007, 274, 1175–1179. [Google Scholar] [CrossRef]
- Evin, A.; Cucchi, T.; Escarguel, G.; Owen, J.; Larson, G.; Strand Vidarsdottir, U.; Dobney, K. Using traditional biometrical data to distinguish West Palearctic wild boar and domestic pigs in the archaeological record: New methods and standards. J. Archaeol. Sci. 2014, 43, 1–8. [Google Scholar] [CrossRef]
- Kohut, M. Assessment of external appearance of Black-Motley cows. Acta Sci. Pol. Zootech. 2015, 14, 101–108. [Google Scholar]
- Ramírez, O.; Burgos-Paz, W.; Casas, E.; Ballester, M.; Bianco, E.; Olalde, I.; Santpere, G.; Novella, V.; Gut, M.; Lalueza-Fox, C.; et al. Genome data from a sixteenth century pig illuminate modern breed relationships. Heredity 2015, 114, 175–184. [Google Scholar] [CrossRef]
- Hughes, S.; Fernández, H.; Cucchi, T.; Duffraisse, M.; Casabianca, F.; Istria, D.; Pompanon, F.; Vigne, J.-D.; Hänni, C.; Taberlet, P. A Dig into the Past Mitochondrial Diversity of Corsican Goats Reveals the Influence of Secular Herding Practices. PLoS ONE 2012, 7, e30272. [Google Scholar] [CrossRef]
- Gabbianelli, F.; Gargani, M.; Pariset, L.; Mariotti, M.; Alhaique, F.; De Minicis, E.; Barelli, L.; Ciammetti, E.; Redi, F.; Valentini, A. Mitochondrial DNA analysis of medieval sheep (Ovis aries) in central Italy reveals the predominance of haplogroup B already in the Middle Ages. Anim. Genet. 2015, 46, 329–332. [Google Scholar] [CrossRef]
- Gargani, M.; Pariset, L.; Lenstra, J.A.; De Minicis, E.; European Cattle Genetic Diversity Consortium; Valentini, A. Microsatellite genotyping of medieval cattle from central Italy suggests an old origin of Chianina and Romagnola cattle. Front. Genet. 2015, 6, 68. [Google Scholar] [CrossRef]
- Iemura, A.; Tsai, M.; Ando, A.; Wershil, B.K.; Galli, S.J. The c-kit ligand, stem cell factor, promotes mast cell survival by suppressing apoptosis. Am. J. Pathol. 1994, 144, 321–328. [Google Scholar]
- Besmer, P.; Murphy, J.E.; George, P.C.; Qiu, F.; Bergold, P.J.; Lederman, L.; Snyder, H.W.; Brodeur, D.; Zuckerman, E.E.; Hardy, W.D. A new acute transforming feline retrovirus and relationship of its oncogene v-kit with the protein kinase gene family. Nature 1986, 320, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Rodríguez, A.; Estellé, J.; Blin, A.; Muñoz, M.; Créchet, F.; Demenais, F.; Vincent-Naulleau, S.; Bourneuf, E. KIT and melanoma predisposition in pigs: Sequence variants and association analysis. Anim. Genet. 2014, 45, 445–448. [Google Scholar] [CrossRef] [PubMed]
- Alhaique, F.; Piermartini, L.; Romagnoli, G. Un castello della valle tiberina tra la fine del medioevo e l’inizio dell’età moderna. Stratigrafia e materiali di un pozzo da butto nella rocca di Graffignano (Viterbo). In Proceedings of the Atti VIII Congresso Nazionale di Archeologia Medievale, Matera, Italy, 12–15 September 2018. [Google Scholar]
- Romagnoli, G.; Alhaique, F.; Brancazi, L.; Piermartini, L. Smaltimento e gestione dei rifiuti in una residenza signorile medievale. Lo scavo degli scarichi domestici del Castello Baglioni di Graffignano (VT). Archeol. Mediev. 2020, in press. [Google Scholar]
- Barone, R. Anatomia Comparata dei Mammiferi Domestici; Edagricole: Bologna, Italy, 1981; Volume 1. [Google Scholar]
- Barone, R. Anatomia Comparata dei Mammiferi Domestici; Edagricole: Bologna, Italy, 1995; Volume 3. [Google Scholar]
- Bull, S.; Payne, G. Tooth eruption and epiphyseal fusion in pigs and wild boar. In Ageing and Sexing Animal Bones from Archaeological Sites; Wilson, B., Grigson, C., Payne, S., Eds.; British Archaeological Reports: London, UK, 1982; Volume 109, pp. 55–71. [Google Scholar]
- Grigson, C. Sex and age determination of some bones and teeth of domestic cattle: A review of the literature. In Ageing and Sexing Animal Bones from Archaeological Sites; Wilson, B., Grigson, C., Payne, S., Eds.; BAR British Series; British Archaeological Reports: Oxford, UK, 1982; pp. 7–24. [Google Scholar]
- Payne, S. Kill-off Patterns in Sheep and Goats: The Mandibles from Aşvan Kale. Anatol. Stud. 1973, 23, 281–303. [Google Scholar] [CrossRef]
- Silver, I. The Ageing of Domestic Animals. In Science in Archaeology: A Survey of Progress and Research; Brothwell, D.R., Higgs, E.S., Clark, G., Eds.; Thames and Hudson: London, UK, 1969; pp. 283–302. ISBN 0-500-05011-2. [Google Scholar]
- Teichert, M. Osteometrische Untersuchungen zur Berechnung der Widerristhöhe bei vor- und Frühgeschichtlichen Schweinen; Kühn-Archiv; Akad.-Verlag: Berlin, Germany, 1969. [Google Scholar]
- Gabbianelli, F. A Simple and Robust Method for Sexing Ancient Bovine Bones. Open Forensic Sci. J. 2012, 5, 9–12. [Google Scholar] [CrossRef]
- Gilbert, M.T.P.; Bandelt, H.-J.; Hofreiter, M.; Barnes, I. Assessing ancient DNA studies. Trends Ecol. Evol. 2005, 20, 541–544. [Google Scholar] [CrossRef]
- Yang, D.Y.; Eng, B.; Waye, J.S.; Dudar, J.C.; Saunders, S.R. Technical note: Improved DNA extraction from ancient bones using silica-based spin columns. Am. J. Phys. Anthropol. 1998, 105, 539–543. [Google Scholar] [CrossRef]
- Dabney, J.; Knapp, M.; Glocke, I.; Gansauge, M.-T.; Weihmann, A.; Nickel, B.; Valdiosera, C.; Garcia, N.; Paabo, S.; Arsuaga, J.-L.; et al. Complete mitochondrial genome sequence of a Middle Pleistocene cave bear reconstructed from ultrashort DNA fragments. Proc. Natl. Acad. Sci. USA 2013, 110, 15758–15763. [Google Scholar] [CrossRef]
- Stahlschmidt, M.C.; Collin, T.C.; Fernandes, D.M.; Bar-Oz, G.; Belfer-Cohen, A.; Gao, Z.; Jakeli, N.; Matskevich, Z.; Meshveliani, T.; Pritchard, J.K.; et al. Ancient Mammalian and Plant DNA from Late Quaternary Stalagmite Layers at Solkota Cave, Georgia. Sci. Rep. 2019, 9, 6628. [Google Scholar] [CrossRef]
- Fontanesi, L.; D’Alessandro, E.; Scotti, E.; Liotta, L.; Crovetti, A.; Chiofalo, V.; Russo, V. Genetic heterogeneity and selection signature at the KIT gene in pigs showing different coat colours and patterns: Pig KIT polymorphisms and coat colours. Anim. Genet. 2010, 41, 478–492. [Google Scholar] [CrossRef]
- Fontanesi, L.; Scotti, E.; Gallo, M.; Nanni Costa, L.; Dall’Olio, S. Authentication of “mono-breed” pork products: Identification of a coat colour gene marker in Cinta Senese pigs useful to this purpose. Livest. Sci. 2016, 184, 71–77. [Google Scholar] [CrossRef]
- Beugin, M.-P.; Baubet, E.; Dufaure De Citres, C.; Kaerle, C.; Muselet, L.; Klein, F.; Queney, G. A set of 20 multiplexed single nucleotide polymorphism (SNP) markers specifically selected for the identification of the wild boar (Sus scrofa scrofa) and the domestic pig (Sus scrofa domesticus). Conserv. Genet. Resour. 2017, 9, 671–675. [Google Scholar] [CrossRef]
- Reed, C.A. A Natural History of Domesticated Mammals. Juliet Clutton-Brock. Q. Rev. Biol. 1990, 65, 232. [Google Scholar] [CrossRef]
- Marshall, F.B.; Dobney, K.; Denham, T.; Capriles, J.M. Evaluating the roles of directed breeding and gene flow in animal domestication. Proc. Natl. Acad. Sci. USA 2014, 111, 6153–6158. [Google Scholar] [CrossRef]
- Larson, G.; Albarella, U.; Dobney, K.; Rowley-Conwy, P.; Schibler, J.; Tresset, A.; Vigne, J.-D.; Edwards, C.J.; Schlumbaum, A.; Dinu, A.; et al. Ancient DNA, pig domestication, and the spread of the Neolithic into Europe. Proc. Natl. Acad. Sci. USA 2007, 104, 15276–15281. [Google Scholar] [CrossRef]
- Godinho, R.; López-Bao, J.V.; Castro, D.; Llaneza, L.; Lopes, S.; Silva, P.; Ferrand, N. Real-time assessment of hybridization between wolves and dogs: Combining noninvasive samples with ancestry informative markers. Mol. Ecol. Resour. 2015, 15, 317–328. [Google Scholar] [CrossRef]
- Frantz, L.A.F.; Schraiber, J.G.; Madsen, O.; Megens, H.-J.; Cagan, A.; Bosse, M.; Paudel, Y.; Crooijmans, R.P.M.A.; Larson, G.; Groenen, M.A.M. Evidence of long-term gene flow and selection during domestication from analyses of Eurasian wild and domestic pig genomes. Nat. Genet. 2015, 47, 1141–1148. [Google Scholar] [CrossRef]
- Larson, G.; Karlsson, E.K.; Perri, A.; Webster, M.T.; Ho, S.Y.; Peters, J.; Stahl, P.W.; Piper, P.J.; Lingaas, F.; Fredholm, M.; et al. Rethinking dog domestication by integrating genetics, archeology, and biogeography. Proc. Natl. Acad. Sci. USA 2012, 109, 8878–8883. [Google Scholar] [CrossRef]
- Wang, Z.; Uzun, M.; Dione, M.M.; Olsaker, I.; Holm, L.E.; Saarma, U.; Ahmad, S.; Marzanov, N.; Eythorsdottir, E.; Holland, M.J.; et al. Revealing the history of sheep domestication using retrovirus integrations. Science 2009, 324, 532–536. [Google Scholar]
- Evin, A.; Owen, J.; Larson, G.; Debiais-Thibaud, M.; Cucchi, T.; Vidarsdottir, U.S.; Dobney, K. A test for paedomorphism in domestic pig cranial morphology. Biol. Lett. 2017, 13, 20170321. [Google Scholar] [CrossRef]
- Goedbloed, D.J.; Megens, H.J.; Van Hooft, P.; Herrero-Medrano, J.M.; Lutz, W.; Alexandri, P.; Crooijmans, R.P.M.A.; Groenen, M.; Van Wieren, S.E.; Ydenberg, R.C.; et al. Genome-wide single nucleotide polymorphism analysis reveals recent genetic introgression from domestic pigs into Northwest European wild boar populations: INTROGRESSION IN WILD BOAR. Mol. Ecol. 2013, 22, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Giuffra, E.; Evans, G.; Törnsten, A.; Wales, R.; Day, A.; Looft, H.; Plastow, G.; Andersson, L. The Belt mutation in pigs is an allele at the Dominant white (I/KIT) locus. Mamm. Genome 1999, 10, 1132–1136. [Google Scholar] [CrossRef] [PubMed]
- Virgili, R.; Degni, M.; Schivazappa, C.; Faeti, V.; Poletti, E.; Marchetto, G.; Pacchioli, M.T.; Mordenti, A. Effect of age at slaughter on carcass traits and meat quality of Italian heavy pigs1. J. Anim. Sci. 2003, 81, 2448–2456. [Google Scholar] [CrossRef] [PubMed]
- Latorre, M.A.; Lázaro, R.; Valencia, D.G.; Medel, P.; Mateos, G.G. The effects of gender and slaughter weight on the growth performance, carcass traits, and meat quality characteristics of heavy pigs1. J. Anim. Sci. 2004, 82, 526–533. [Google Scholar] [CrossRef]
- MacKinnon, M. High on the Hog: Linking Zooarchaeological, Literary, and Artistic Data for Pig Breeds in Roman Italy. Am. J. Archaeol. 2001, 105, 649. [Google Scholar] [CrossRef]
- Crescentio, P. de Agricultura Vulgare; Bindoni, A., Ed.; Venetiis: Venice, Italy, 1519. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Pit Number | Chronology | SU | MNI | Sampled |
|---|---|---|---|---|
| 1 | ca.1400–1425 CE | 1004 | 1 | 1 |
| 1006 | 1 | - | ||
| 1007 | 1 | 1 | ||
| 2 | ca. 1450–1525 CE | 2000 | 2 | - |
| 2002 | 3 | 1 | ||
| 2003 | 6 | 3 | ||
| 2004 | 6 | 4 | ||
| 2005 | 5 | 4 | ||
| ca. 1425–1450 CE | 2006-2007-2008 | 5 | 5 | |
| 2009-2010-2011 | 3 | 2 |
| Pit Number | Sample ID | SU | Skeletal Element | Age | DE1 a (ng/µL) | DE2 b (ng/µL) | PCR1 c | PCR2 d | PCR3 e |
|---|---|---|---|---|---|---|---|---|---|
| 1 | G8 | 1004 | 4th metatarsal | <24 months | 0 | 6 | DP(C/C) | ||
| 1 | G9 | 1007 | Mandible, right | 31–35 months | 6 | DP(C/C) | DP(T/T) | F | |
| 2 | G21 | 2008 | Maxilla, left | 7–11 months | 0 | 2 | DP(C/C) | ||
| 2 | G24 | 2008 | Mandible, right | 2–4 months | 0 | 3 | CINTA(T/T) | DP(T/T) | |
| 2 | G161 | 2002 | Maxilla, right | >4 years | 3 | 7 | CINTA(T/T) | ||
| 2 | G182 | 2003 | Cranial fragment, right | Undetectable | 0 | 3 | DP(C/C) | WBO(T/T) | |
| 2 | G192 | 2003 | Mandible, left | 31–35 months | 3 | DP(C/C) | |||
| 2 | G206 | 2003 | Mandible, left | 0–4 months | 4 | DP(C/C) | DP(T/T) | ||
| 2 | GB | 2004 | 3rd metatarsal, left | >24 months | 3 | DP(C/C) | |||
| 2 | G262 | 2004 | Maxilla, right | 7–11 months | 4 | DP(C/C) | DP(T/T) | M | |
| 2 | G264 | 2004 | Maxilla, left | 7–11 months | 4 | DP(C/C) | WBO(C/C) | F | |
| 2 | G286 | 2004 | Maxilla, right | 3–4 years | 2 | DP(C/C) | DP(T/T) | F | |
| 2 | GC | 2005 | Femur, left | 1–2 years | 11 | DP(C/C) | DP(T/T) | F | |
| 2 | GD | 2005 | Mandible, left | 0–4 months | 4.5 | 7 | CINTA(C/T) | ||
| 2 | G133 | 2005 | Mandible, left | 3–4 years | 3 | 4 | CINTA (C/T) | WBO(C/C) | F |
| 2 | G198 | 2005 | Mandible, left | 0–4 months | 2 | DP(C/C) | DP(T/T) | F | |
| 2 | G184 | 2006 | Mandible, left | 0–4 months | 1 | DP(C/C) | DP(T/T) | F | |
| 2 | G23 | 2008 | Mandible, right | 0–4 months | 4.5 | DP(C/C) | WBO(C/C) | M | |
| 2 | G27 | 2008 | Mandible, left | >4 years | 5.5 | DP(C/C) | DP(T/T) | F | |
| 2 | G4 | 2009 | Mandible, left | 7–11 months | 4 | DP(C/C) | DP(T/T) | F | |
| 2 | G1 | 2011 | Mandible, right | 0–4 months | 4.5 | DP(C/C) | DP(T/T) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gabbianelli, F.; Alhaique, F.; Romagnoli, G.; Brancazi, L.; Piermartini, L.; Ottoni, C.; Valentini, A.; Chillemi, G. Was the Cinta Senese Pig Already a Luxury Food in the Late Middle Ages? Ancient DNA and Archaeozoological Evidence from Central Italy. Genes 2020, 11, 85. https://doi.org/10.3390/genes11010085
Gabbianelli F, Alhaique F, Romagnoli G, Brancazi L, Piermartini L, Ottoni C, Valentini A, Chillemi G. Was the Cinta Senese Pig Already a Luxury Food in the Late Middle Ages? Ancient DNA and Archaeozoological Evidence from Central Italy. Genes. 2020; 11(1):85. https://doi.org/10.3390/genes11010085
Chicago/Turabian StyleGabbianelli, Federica, Francesca Alhaique, Giuseppe Romagnoli, Luca Brancazi, Lavinia Piermartini, Claudio Ottoni, Alessio Valentini, and Giovanni Chillemi. 2020. "Was the Cinta Senese Pig Already a Luxury Food in the Late Middle Ages? Ancient DNA and Archaeozoological Evidence from Central Italy" Genes 11, no. 1: 85. https://doi.org/10.3390/genes11010085
APA StyleGabbianelli, F., Alhaique, F., Romagnoli, G., Brancazi, L., Piermartini, L., Ottoni, C., Valentini, A., & Chillemi, G. (2020). Was the Cinta Senese Pig Already a Luxury Food in the Late Middle Ages? Ancient DNA and Archaeozoological Evidence from Central Italy. Genes, 11(1), 85. https://doi.org/10.3390/genes11010085