Gene Co-Expression Networks Restructured Gene Fusion in Rhabdomyosarcoma Cancers

 , ,
, , 
Abstract
1. Introduction
1.1. Gene Fusions in Solid Tumors
1.2. Rhabdomyosarcoma and the FOXO1-PAX3/7 Fusion
1.3. Gene Co-Expression Networks and Rhabdomyosarcoma
1.4. Study Overview
2. Materials and Methods
2.1. Data Source and Pre-Analysis Processing
2.2. Gene Co-Expression Network Analysis
2.3. Relating Co-Expression Module Expression Variance and Fusion Status
2.4. Functional Enrichment Analysis
2.5. Copy Number Variation in Rhabdomyosarcoma
3. Results
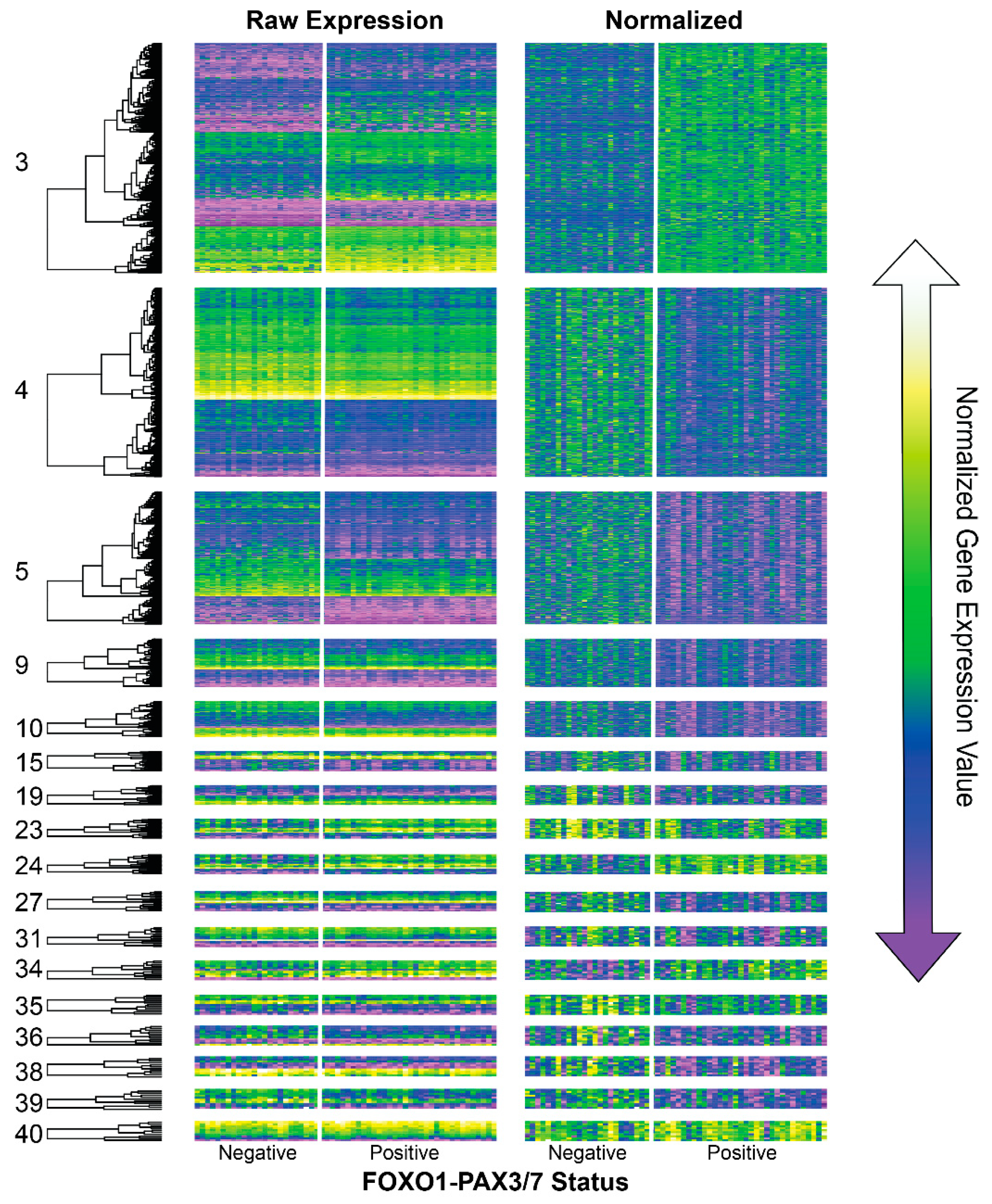
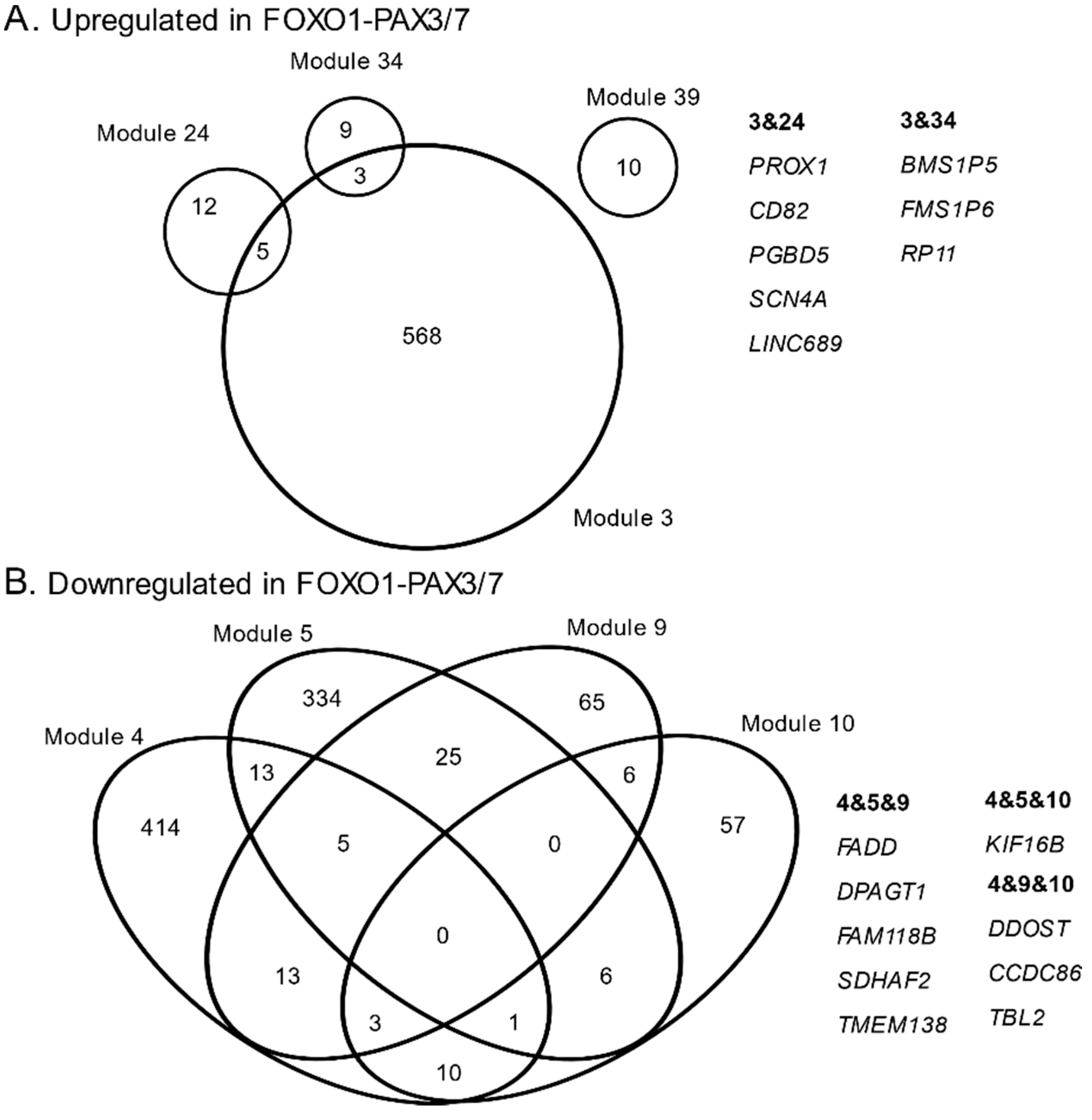
3.1. Consensus Network Analysis of Fusion-Associated Module Expression Patterns
3.2. Gene Ontology, Cytoband, and Co-Expression Atlas
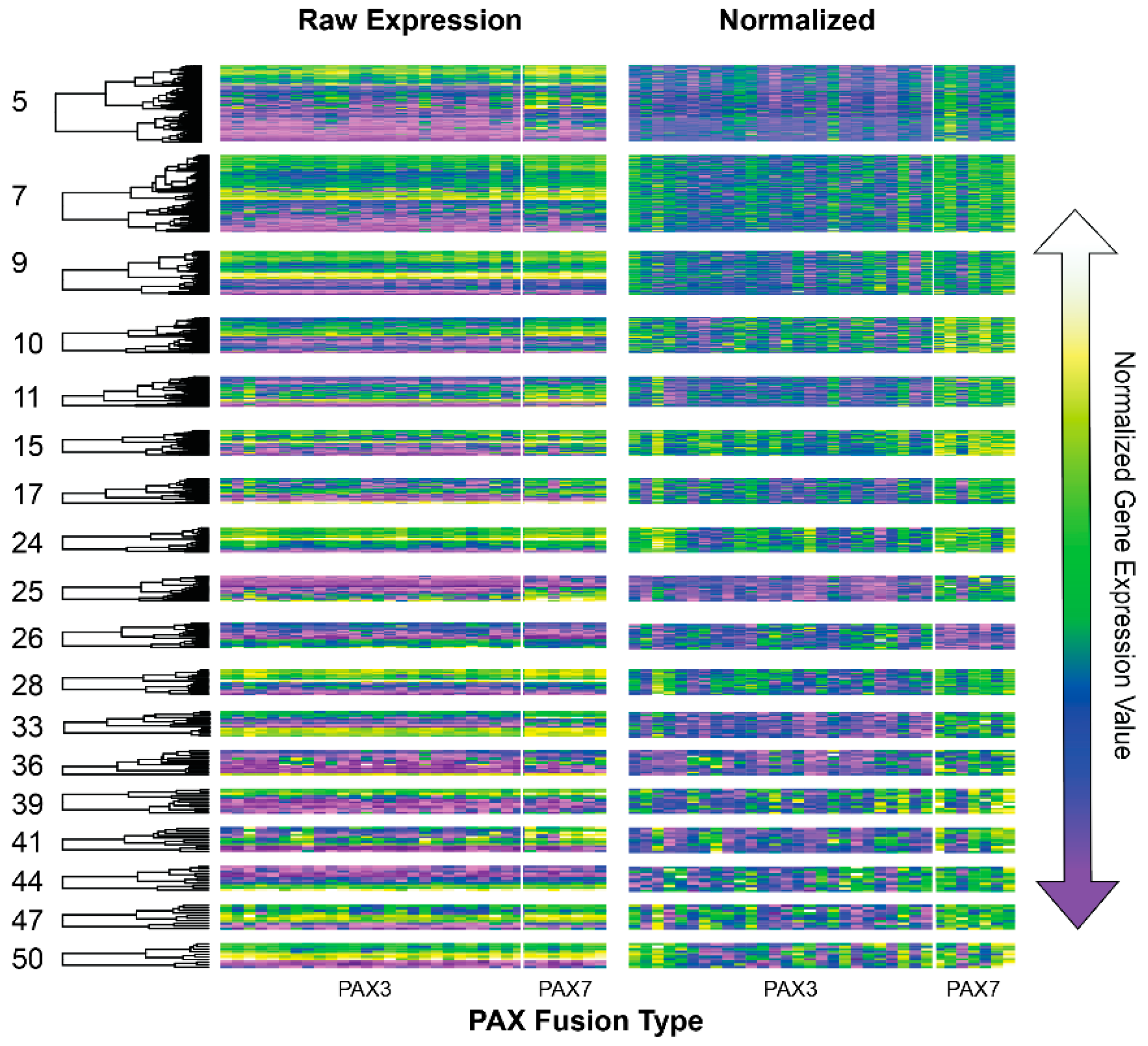
3.3. lmQCM of Fusion Negative and Fusion Positive Gene Co-Expression Networks
3.4. Copy Number Variation
3.5. Copy Number Variation in Gene Modules from Consensus Network
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mitelman, F.; Johansson, B.; Mertens, F. The impact of translocations and gene fusions on cancer causation. Nat. Rev. Cancer 2007, 7, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Barr, F.G. Therapeutic Approaches Targeting PAX3-FOXO1 and Its Regulatory and Transcriptional Pathways in Rhabdomyosarcoma. Molecules 2018, 23, 2798. [Google Scholar] [CrossRef] [PubMed]
- Shern, J.F.; Chen, L.; Chmielecki, J.; Wei, J.S.; Patidar, R.; Rosenberg, M.; Ambrogio, L.; Auclair, D.; Wang, J.; Song, Y.K.; et al. Comprehensive Genomic Analysis of Rhabdomyosarcoma Reveals a Landscape of Alterations Affecting a Common Genetic Axis in Fusion-Positive and Fusion-Negative Tumors. Cancer Discov. 2014, 4, 216–232. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Chatterjee, B.; Wang, Y.; Stevenson, H.S.; Edelman, D.C.; Meltzer, P.S.; Barr, F.G. Distinct methylation profiles characterize fusion-positive and fusion-negative rhabdomyosarcoma. Mod. Pathol. 2015, 28, 1214–1224. [Google Scholar] [CrossRef] [PubMed]
- Raney, R.B.; Anderson, J.R.; Barr, F.G.; Donaldson, S.S.; Pappo, A.S.; Qualman, S.J.; Wiener, E.S.; Maurer, H.M.; Crist, W.M. Rhabdomyosarcoma and Undifferentiated Sarcoma in the First Two Decades of Life: A Selective Review of Intergroup Rhabdomyosarcoma Study Group Experience and Rationale for Intergroup Rhabdomyosarcoma Study V. J. Pediatr. Hematol. Oncol. 2001, 23, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Seki, M.; Nishimura, R.; Yoshida, K.; Shimamura, T.; Shiraishi, Y.; Sato, Y.; Kato, M.; Chiba, K.; Tanaka, H.; Hoshino, N.; et al. Integrated genetic and epigenetic analysis defines novel molecular subgroups in rhabdomyosarcoma. Nat. Commun. 2015, 6, 1–8. [Google Scholar] [CrossRef]
- Drummond, C.J.; Hanna, J.A.; Garcia, M.R.; Devine, D.J.; Heyrana, A.J.; Finkelstein, D.; Rehg, J.E.; Hatley, M.E. Hedgehog Pathway Drives Fusion-Negative Rhabdomyosarcoma Initiated From Non-myogenic Endothelial Progenitors. Cancer Cell 2018, 33, 108–124.e5. [Google Scholar] [CrossRef]
- Skapek, S.X.; Ferrari, A.; Gupta, A.A.; Lupo, P.J.; Butler, E.; Shipley, J.; Barr, F.G.; Hawkins, D.S. Rhabdomyosarcoma. Nat. Rev. Dis. Prim. 2019, 5, 1. [Google Scholar] [CrossRef]
- Spunt, S.L.; Skapek, S.X.; Coffin, C.M. Pediatric Nonrhabdomyosarcoma Soft Tissue Sarcomas. Oncologist 2008, 13, 668–678. [Google Scholar] [CrossRef]
- Dziuba, I.; Kurzawa, P.; Dopierała, M.; Larque, A.B.; Januszkiewicz-Lewandowska, D. Rhabdomyosarcoma in children—current pathologic and molecular classification. Pol. J. Pathol. 2018, 69, 20–32. [Google Scholar] [CrossRef]
- Horn, R.C.; Enterline, H.T. Rhabdomyosarcoma: A clinicopathological study and classification of 39 cases. Cancer 1958, 11, 181–199. [Google Scholar] [CrossRef]
- Mercado, G.E.; Barr, F.G. Fusions Involving PAX and FOX Genes in the Molecular Pathogenesis of Alveolar Rhabdomyosarcoma : Recent Advances. Curr. Mol. Med. 2007, 7, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Giese, W.; Eigel, M.; Westerheide, S.; Engwer, C.; Klipp, E. Influence of cell shape, inhomogeneities and diffusion barriers in cell polarization models. Phys. Biol. 2015, 12, 0066014. [Google Scholar] [CrossRef] [PubMed]
- Cieśla, M.; Dulak, J.; Józkowicz, A. MicroRNAs and epigenetic mechanisms of rhabdomyosarcoma development. Int. J. Biochem. Cell Biol. 2014, 53, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Stewart, E.; McEvoy, J.; Wang, H.; Chen, X.; Honnell, V.; Ocarz, M.; Gordon, B.; Dapper, J.; Blankenship, K.; Yang, Y.; et al. Identification of Therapeutic Targets in Rhabdomyosarcoma through Integrated Genomic, Epigenomic, and Proteomic Analyses. Cancer Cell 2018, 34, 411–426.e19. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Chatterjee, B.; Shern, J.F.; Patidar, R.; Song, Y.; Wang, Y.; Walker, R.L.; Pawel, B.R.; Linardic, C.M.; Houghton, P.; et al. Relationship of DNA methylation to mutational changes and transcriptional organization in fusion-positive and fusion-negative rhabdomyosarcoma. Int. J. Cancer 2019, 144, 2707–2717. [Google Scholar] [CrossRef] [PubMed]
- Gryder, B.E.; Yohe, M.E.; Chou, H.C.; Zhang, X.; Marques, J.; Wachtel, M.; Schaefer, B.; Sen, N.; Song, Y.; Gualtieri, A.; et al. PAX3-FOXO1 establishes myogenic super enhancers and confers BET bromodomain vulnerability. Cancer Discov. 2017, 7, 884–899. [Google Scholar] [CrossRef]
- Vicente-García, C.; Villarejo-Balcells, B.; Irastorza-Azcárate, I.; Naranjo, S.; Acemel, R.D.; Tena, J.J.; Rigby, P.W.J.; Devos, D.P.; Gómez-Skarmeta, J.L.; Carvajal, J.J. Regulatory landscape fusion in rhabdomyosarcoma through interactions between the PAX3 promoter and FOXO1 regulatory elements. Genome Biol. 2017, 18, 106. [Google Scholar] [CrossRef]
- Reichek, J.L.; Duan, F.; Smith, L.M.; Gustafson, D.M.; O’Connor, R.S.; Zhang, C.; Dunlevy, M.J.; Gastier-Foster, J.M.; Barr, F.G. Genomic and clinical analysis of amplification of the 13q31 chromosomal region in alveolar rhabdomyosarcoma: A report from the children’s oncology group. Clin. Cancer Res. 2011, 17, 1463–1473. [Google Scholar] [CrossRef]
- Barr, F.; Duan, F.; Smith, L.M.; Gustafson, D.; Pitts, M.; Hammond, S.; Gastier-Foster, J.M. Genomic and Clinical Analysis of 2p24 and 12q13-q14 Amplification in Alveolar Rhabdomyosarcoma: A Report from the Children’s Oncology Group. Genes Chromosom. Cancer 2009, 48, 661–672. [Google Scholar] [CrossRef]
- Khan, J.; Simon, R.; Bittner, M.; Chen, Y.; Leighton, S.B.; Pohida, T.; Smith, P.D.; Jiang, Y.; Gooden, G.C.; Trent, J.M.; et al. Gene Expression Profiling of Alveolar Rhabdomyosarcoma with cDNA Microarrays. Cancer Res. 1998, 58, 5009–5013. [Google Scholar] [PubMed]
- Davicioni, E.; Finckenstein, F.G.; Shahbazian, V.; Buckley, J.D.; Triche, T.J.; Anderson, M.J. Identification of a PAX-FKHR gene expression signature that defines molecular classes and determines the prognosis of alveolar rhabdomyosarcomas. Cancer Res. 2006, 66, 6936–6946. [Google Scholar] [CrossRef] [PubMed]
- De Pittà, C.; Tombolan, L.; Albiero, G.; Sartori, F.; Romualdi, C.; Jurman, G.; Carli, M.; Furlanello, C.; Lanfranchi, G.; Rosolen, A. Gene expression profiling identifies potential relevant genes in alveolar rhabdomyosarcoma pathogenesis and discriminates PAX3-FKHR positive and negative tumors. Int. J. Cancer 2006, 118, 2772–2781. [Google Scholar] [CrossRef] [PubMed]
- Missiaglia, E.; Williamson, D.; Chisholm, J.; Wirapati, P.; Pierron, G.; Petel, F.; Concordet, J.-P.; Thway, K.; Oberlin, O.; Pritchard-Jones, K.; et al. PAX3/FOXO1 Fusion Gene Status Is the Key Prognostic Molecular Marker in Rhabdomyosarcoma and Significantly Improves Current Risk Stratification. J. Clin. Oncol. 2012, 30, 1670–1677. [Google Scholar] [CrossRef] [PubMed]
- Williamson, D.; Missiaglia, E.; De Reyniès, A.; Pierron, G.; Thuille, B.; Palenzuela, G.; Thway, K.; Orbach, D.; Laé, M.; Fréneaux, P.; et al. Fusion gene-negative alveolar rhabdomyosarcoma is clinically and molecularly indistinguishable from embryonal rhabdomyosarcoma. J. Clin. Oncol. 2010, 28, 2151–2158. [Google Scholar] [CrossRef] [PubMed]
- Missiaglia, E.; Williamson, D.; Chisholm, J.; Wirapati, P.; Pierron, G.; Petel, F.; Concordet, J.-P.; Thway, K.; Oberlin, O.; Pritchard-Jones, K.; et al. Reply to S. Stegmaier et al. J. Clin. Oncol. 2012, 30, 4040–4041. [Google Scholar] [CrossRef]
- Stegmaier, S.; Bielack, S.S.; Leuschner, I.; Klingebiel, T.; Koscielniak, E. Questionable Universal Validity of PAX3/FOXO1 Fusion Gene Status As Molecular Marker for Improvement of Risk Stratification in Rhabdomyosarcoma Therapy. J. Clin. Oncol. 2012, 30, 4039–4040. [Google Scholar] [CrossRef] [PubMed]
- Linardic, C.M. PAX3-FOXO1 fusion gene in rhabdomyosarcoma. Cancer Lett. 2008, 270, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.J.; Holder, D.D.; Pawel, B.R.; Zhang, C.; Barr, F.G. High expression of the PAX3-FKHR oncoprotein is required to promote tumorigenesis of human myoblasts. Am. J. Pathol. 2009, 175, 2600–2608. [Google Scholar] [CrossRef] [PubMed]
- Marshall, A.D.; Picchione, F.; Geltink, R.I.K.; Grosveld, G.C. PAX3-FOXO1 Induces Up-Regulation of Noxa Sensitizing Alveolar Rhabdomyosarcoma Cells to Apoptosis. Neoplasia 2013, 15, 738–748. [Google Scholar] [CrossRef]
- Schmitt-Ney, M.; Camussi, G. The PAX3-FOXO1 fusion protein present in rhabdomyosarcoma interferes with normal FOXO activity and the TGF-β pathway. Plos One 2015, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Tenente, I.M.; Hayes, M.N.; Ignatius, M.S.; McCarthy, K.; Yohe, M.; Sindiri, S.; Gryder, B.; Oliveira, M.L.; Ramakrishnan, A.; Tang, Q.; et al. Myogenic regulatory transcription factors regulate growth in rhabdomyosarcoma. Elife 2017, 6, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Davie, J.K. New insights into signalling-pathway alterations in rhabdomyosarcoma. Br. J. Cancer 2015, 112, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Yu, Y.; Bilke, S.; Walker, R.L.; Mayeenuddin, L.H.; Azorsa, D.O.; Yang, F.; Pineda, M.; Helman, L.J.; Meltzer, P.S. Genome-wide identification of PAX3-FKHR binding sites in rhabdomyosarcoma reveals candidate target genes important for development and cancer. Cancer Res. 2010, 70, 6497–6508. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.R.; Chatterjee, B.; Olanich, M.E.; Khan, J.; Miettinen, M.M.; Hewitt, S.M.; Barr, F.G. PAX3-FOXO1 is essential for tumour initiation and maintenance but not recurrence in a human myoblast model of rhabdomyosarcoma. J. Pathol. 2017, 241, 626–637. [Google Scholar] [CrossRef] [PubMed]
- Monti, E.; Fanzani, A. Uncovering metabolism in rhabdomyosarcoma. Cell Cycle 2016, 15, 184–195. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. Eigengene networks for studying the relationships between co-expression modules. Bmc Syst. Biol. 2007, 1, 54. [Google Scholar] [CrossRef]
- Zhang, B.; Genetics, H.; Zhang, B.; Horvath, S. A General Framework for Weighted Gene Co- Expression Network Analysis. Stat. Appl. Genet. Mol. Biol. 2005, 4, 1–43. [Google Scholar] [CrossRef]
- Fiscon, G.; Conte, F.; Farina, L.; Paci, P. Network-Based Approaches to Explore Complex Biological Systems towards Network Medicine. Genes 2018, 9, 437. [Google Scholar] [CrossRef]
- Xiang, Y.; Zhang, J.; Huang, K. Mining the tissue-tissue gene co-expression network for tumor microenvironment study and biomarker prediction. Bmc Genom. 2013, 14, S4. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, K.; Xiang, Y.; Islam, M.; Kotian, S.; Kais, Z.; Lee, C.; Arora, M.; Liu, H.; Parvin, J.D.; et al. Weighted Frequent Gene Co-expression Network Mining to Identify Genes Involved in Genome Stability. Plos Comput. Biol. 2012, 8, e1002656. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Huang, K. Normalized ImQCM: An Algorithm for Detecting Weak Quasi-Cliques in Weighted Graph with Applications in Gene Co-Expression Module Discovery in Cancers. Cancer Inform. 2016, 13, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Barr, F.; Smith, L.; Lynch, J.C.; Strzelecki, D.; Parham, D.; Qualman, S.; Breifeld, P. Examination of Gene Fusion Status in Archival Samples of Alveolar Rhabdomyosarcoma Entered on the Intergroup Rhabdomyosarcoma Study-III Trial: a Report from the Children’s Oncology Group. J. Mol. Diagn. 2006, 8, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.; Meltzer, P.S. GEOquery: A bridge between the Gene Expression Omnibus (GEO) and Bioconductor. Bioinformatics 2007, 23, 1846–1847. [Google Scholar] [CrossRef] [PubMed]
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating high-throughput genomic analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhang, J.; Huang, K.; Han, Z. lmQCM: An Algorithm for Gene Co-Expression Analysis. Available online: https://github.com/huangzhii/lmQCM (accessed on 15 May 2019).
- R Core Team R: A Language and Environment for Statistical Computing; R foundation for Statistical Computing: Vienna, Austria, 2018.
- Zhao, P.; Zhang, C.Q. A new clustering method and its application in social networks. Pattern Recognit. Lett. 2011, 32, 2109–2118. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. Available online: https://github.com/vegandevs/vegan (accessed on 15 May 2019).
- Seshan, V.E.; Olshen, A. DNAcopy: DNA Copy Number Data Analysis. Available online: https://github.com/veseshan/DNAcopy (accessed on 15 May 2019).
- Mermel, C.H.; Schumacher, S.E.; Hill, B.; Meyerson, M.L.; Beroukhim, R.; Getz, G. GISTIC2.0 facilitates sensitive and confident localization of the targets of focal somatic copy-number alteration in human cancers. Genome Biol. 2011, 12, R41. [Google Scholar] [CrossRef] [PubMed]
- Laddha, S.V.; Ganesan, S.; Chan, C.S.; White, E. Mutational Landscape of the Essential Autophagy Gene BECN1 in Human Cancers. Mol. Cancer Res. 2014, 12, 485–490. [Google Scholar] [CrossRef]
- Petak, I.; Douglas, L.; Tillman, D.M.; Vernes, R.; Houghton, J.A. Pediatric Rhabdomyosarcoma Cell Lines Are Resistant to Fas-induced Apoptosis and Highly Sensitive to TRAIL-induced Apoptosis. Clin. Cancer Res. 2000, 6, 4119–4127. [Google Scholar]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. Bmc Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Heinicke, U.; Haydn, T.; Kehr, S.; Vogler, M.; Fulda, S. BCL-2 selective inhibitor ABT-199 primes rhabdomyosarcoma cells to histone deacetylase inhibitor-induced apoptosis. Oncogene 2018, 37, 5325–5339. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Module | Gene Count | p |
|---|---|---|
| 3 | 576 | 0.001 |
| 4 | 459 | 0.001 |
| 5 | 334 | 0.001 |
| 9 | 118 | 0.001 |
| 10 | 84 | 0.001 |
| 15 | 33 | 0.005 |
| 19 | 24 | 0.001 |
| 23 | 17 | 0.038 |
| 24 | 17 | 0.001 |
| 27 | 15 | 0.001 |
| 31 | 13 | 0.017 |
| 34 | 12 | 0.001 |
| 35 | 11 | 0.001 |
| 36 | 11 | 0.001 |
| 38 | 10 | 0.001 |
| 39 | 10 | 0.001 |
| 40 | 10 | 0.019 |
| Module | GO ID | Biological Process | FDR B&Y | Cytoband | FDR B&Y | |
|---|---|---|---|---|---|---|
| 3 | NA | NA | NA | | | 9q21.11 | <0.0001 |
| | | 16p12.3 | <0.0001 | ||||
| | | 16p12.2 | <0.001 | ||||
| | | 10q26 | <0.001 | ||||
| 4 | 6369 | RNA Processing | <0.0001 | | | 8q24.3 | <0.0001 |
| 34660 | ncRNA metabolic process | <0.0001 | | | 11q13 | <0.0001 | |
| 34470 | ncRNA processing | <0.0001 | | | 11q21 | <0.0001 | |
| 22613 | ribonucleoprotein complex biogenesis | <0.0001 | | | | 8q24.13 | <0.0001 | |
| 70647 | protein conjugation by conjugation or removal | <0.0001 | | | | 8p21.3 | <0.001 | |
| 5 | 9790 | embryo development | <0.0001 | | | 11q13 | <0.001 |
| 35295 | tube development | <0.0001 | | | 11q21 | <0.001 | |
| 31175 | neuron projection development | <0.0001 | | | | 11p15.3 | 0.012 | |
| 22008 | neurogenesis | <0.0001 | | | 7q21 | 0.046 | |
| 45595 | regulation of cell differentiation | <0.0001 | | | | |||
| 9 | 6890 | retrograde vesicle transport | <0.0001 | | | 11q13 | <0.0001 |
| 48193 | Golgi transport | <0.0001 | | | 11p15.3 | <0.0001 | |
| 6888 | ER to Golgi vesicle transport | <0.0001 | | | 11q12.2 | <0.001 | |
| 30968 | endoplasmic reticulum unfolded protein | <0.001 | | | | 11p15.5-4 | <0.01 | |
| 6986 | response to unfolded protein | <0.001 | | | 11p12-p11 | <0.01 | |
| 10 | 6396 | RNA Processing | <0.01 | | | 19p13.3 | <0.0001 |
| 22613 | ribonucleoprotein complex biogenesis | <0.01 | | | | 19p13.11 | <0.0001 | |
| NA | NA | NA | | | 19p13.2 | <0.0001 | |
| | | 19p13.32 | <0.01 |
| Module | Input # | ID | Name | p |
|---|---|---|---|---|
| 3 | 576 | M2012 | Genes up-regulated in alveolar rhabdomyosarcoma compared to embryonic rhabdomyosarcoma | p < 0.0001 |
| 4 | 475 | 15902281—Table S1A | Human Leukemia Schoch05 | p < 0.0001 |
| 5 | 326 | M8519 | Genes down-regulated in alveolar rhabdomyosarcoma compared to embryonic rhabdomyosarcoma | p < 0.0001 |
| 9 | 121 | 15902281—Table S1B | Human Leukemia Schoch05 | p < 0.0001 |
| 10 | 80 | FacebaseRNAseq e8.5 Hind Brain Neural Epithelium top-relative-expression-ranked 2500 k-means-cluster#2 | p < 0.0001 |
| Module | Gene Count | p |
|---|---|---|
| 5 | 178 | 0.002 |
| 7 | 148 | 0.006 |
| 9 | 84 | 0.016 |
| 10 | 72 | 0.002 |
| 11 | 62 | 0.001 |
| 15 | 37 | 0.004 |
| 17 | 32 | 0.038 |
| 24 | 19 | 0.03 |
| 25 | 18 | 0.001 |
| 26 | 18 | 0.011 |
| 28 | 16 | 0.015 |
| 33 | 14 | 0.001 |
| 36 | 13 | 0.001 |
| 39 | 12 | 0.004 |
| 41 | 11 | 0.008 |
| 44 | 11 | 0.022 |
| 47 | 10 | 0.045 |
| 50 | 10 | 0.05 |
| Module | Total Gene Count | Genes with CNV | p |
|---|---|---|---|
| 3 | 576 | 37 | 0.657 |
| 4 | 459 | 157 | <0.0001 |
| 5 | 334 | 44 | <0.0001 |
| 9 | 118 | 10 | 0.241 |
| 10 | 84 | 4 | 0.819 |
| 15 | 33 | 4 | 0.132 |
| 19 | 24 | 0 | 0.398 |
| 23 | 17 | 0 | 0.621 |
| 24 | 17 | 1 | 1 |
| 27 | 15 | 0 | 1 |
| 31 | 13 | 0 | 1 |
| 34 | 12 | 0 | 1 |
| 35 | 11 | 0 | 1 |
| 36 | 11 | 0 | 1 |
| 38 | 10 | 1 | 0.46 |
| 39 | 10 | 0 | 1 |
| 40 | 10 | 1 | 0.46 |
| 41 | 10 | 0 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Helm, B.R.; Zhan, X.; Pandya, P.H.; Murray, M.E.; Pollok, K.E.; Renbarger, J.L.; Ferguson, M.J.; Han, Z.; Ni, D.; Zhang, J.; et al. Gene Co-Expression Networks Restructured Gene Fusion in Rhabdomyosarcoma Cancers. Genes 2019, 10, 665. https://doi.org/10.3390/genes10090665
Helm BR, Zhan X, Pandya PH, Murray ME, Pollok KE, Renbarger JL, Ferguson MJ, Han Z, Ni D, Zhang J, et al. Gene Co-Expression Networks Restructured Gene Fusion in Rhabdomyosarcoma Cancers. Genes. 2019; 10(9):665. https://doi.org/10.3390/genes10090665
Chicago/Turabian StyleHelm, Bryan R., Xiaohui Zhan, Pankita H. Pandya, Mary E. Murray, Karen E. Pollok, Jamie L. Renbarger, Michael J. Ferguson, Zhi Han, Dong Ni, Jie Zhang, and et al. 2019. "Gene Co-Expression Networks Restructured Gene Fusion in Rhabdomyosarcoma Cancers" Genes 10, no. 9: 665. https://doi.org/10.3390/genes10090665
APA StyleHelm, B. R., Zhan, X., Pandya, P. H., Murray, M. E., Pollok, K. E., Renbarger, J. L., Ferguson, M. J., Han, Z., Ni, D., Zhang, J., & Huang, K. (2019). Gene Co-Expression Networks Restructured Gene Fusion in Rhabdomyosarcoma Cancers. Genes, 10(9), 665. https://doi.org/10.3390/genes10090665