Decreased Temperature Sensitivity of Vestigial Gene Expression in Temperate Populations of Drosophila melanogaster


Abstract
1. Introduction
2. Materials and Methods
2.1. Expression Analysis
2.2. Sequence Analysis
3. Results
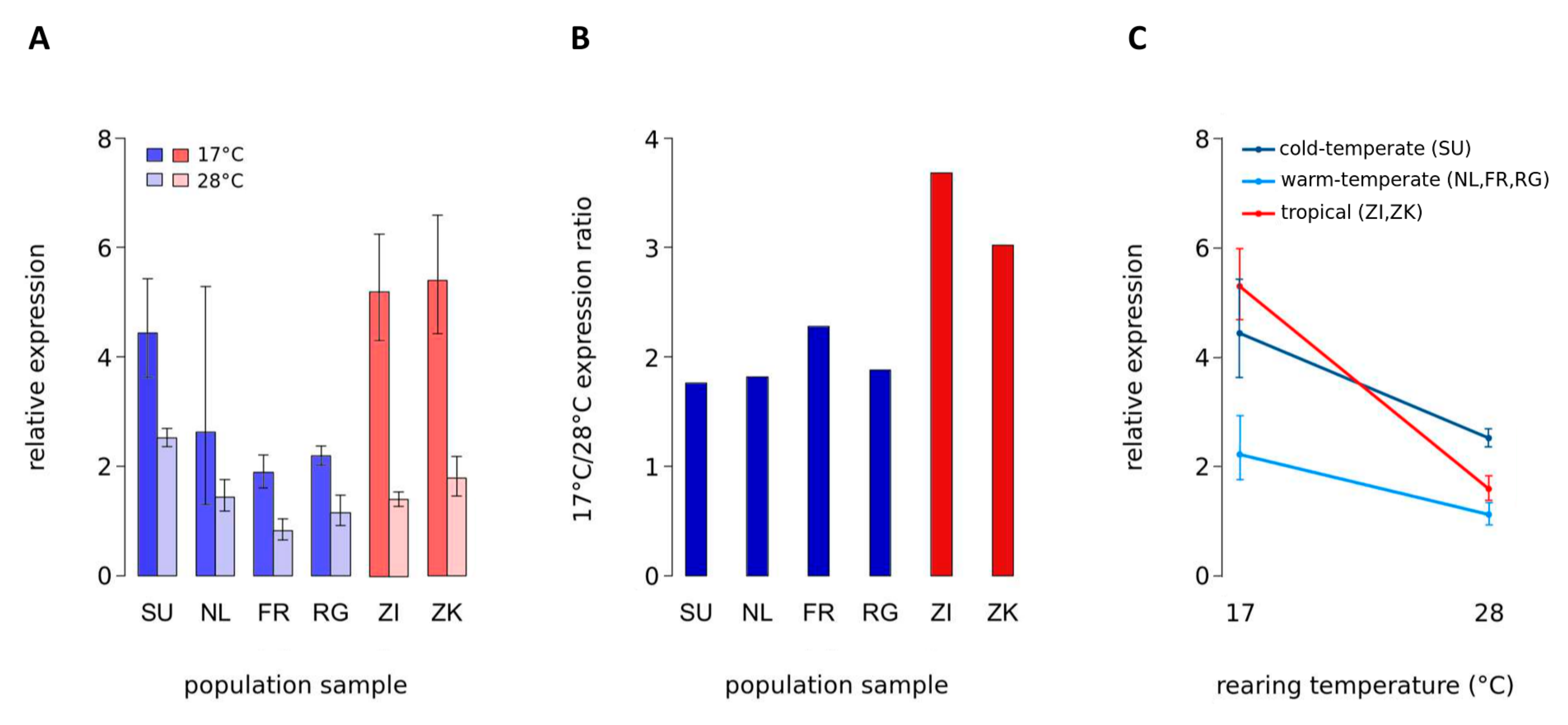
3.1. Expression Analysis in Adult D. melanogaster
3.2. Tissue-Specific Expression Analysis in Third Instar Larvae of D. melanogaster
3.3. Sequence Analysis of Genetic Differentiation at the vg Locus
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andrewartha, H.G.; Birch, L.C. The Distribution and Abundance of Animals; University Of Chicago Press: Chicago, IL, USA; p. 1954.
- Cossins, A. Temperature Biology of Animals; Springer Science & Business Media: Heidelberg, Germany, 2012; ISBN 978-94-009-3127-5. [Google Scholar]
- Hoffmann, A.A.; Sørensen, J.G.; Loeschcke, V. Adaptation of Drosophila to temperature extremes: Bringing together quantitative and molecular approaches. J. Therm. Biol. 2003, 28, 175–216. [Google Scholar] [CrossRef]
- Ayrinhac, A.; Debat, V.; Gibert, P.; Kister, A.-G.; Legout, H.; Moreteau, B.; Vergilino, R.; David, J.R. Cold adaptation in geographical populations of Drosophila melanogaster: Phenotypic plasticity is more important than genetic variability. Funct. Ecol. 2004, 18, 700–706. [Google Scholar] [CrossRef]
- Pool, J.E.; Braun, D.T.; Lack, J.B. Parallel evolution of cold tolerance within Drosophila melanogaster. Mol. Biol. Evol. 2017, 34, 349–360. [Google Scholar] [CrossRef] [PubMed]
- David, J.; Capy, P. Genetic variation of Drosophila melanogaster natural populations. Trends Genet. 1988, 4, 106–111. [Google Scholar] [CrossRef]
- Lachaise, D.; Silvain, J.-F. How Two Afrotropical Endemics Made Two Cosmopolitan Human Commensals: the Drosophila Melanogaster–D. Simulans Palaeogeographic Riddle. Genetics 2004, 120, 17–39. [Google Scholar] [CrossRef]
- Pool, J.E.; Corbett-Detig, R.B.; Sugino, R.P.; Stevens, K.A.; Cardeno, C.M.; Crepeau, M.W.; Duchen, P.; Emerson, J.J.; Saelao, P.; Begun, D.J.; et al. Population Genomics of Sub-Saharan Drosophila melanogaster: African Diversity and Non-African Admixture. PLoS Genet. 2012, 8, e1003080. [Google Scholar] [CrossRef] [PubMed]
- Stephan, W.; Li, H. The recent demographic and adaptive history of Drosophila melanogaster. Heredity 2007, 98, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Stephan, W.; Przeworski, M. Inferring the Demographic History and Rate of Adaptive Substitution in Drosophila. PLoS Genet. 2006, 2, e166. [Google Scholar] [CrossRef]
- Laurent, S.J.Y.; Werzner, A.; Excoffier, L.; Wolfgang, S. Approximate Bayesian Analysis of Drosophila melanogaster Polymorphism Data Reveals a Recent Colonization of Southeast Asia. Mol. Biol. Evol. 2011, 28, 2041–2051. [Google Scholar] [CrossRef]
- Levine, M.T.; Eckert, M.L.; Begun, D.J. Whole-genome expression plasticity across tropical and temperate Drosophila melanogaster populations from Eastern Australia. Mol. Biol. Evol. 2011, 28, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wit, J.; Svetec, N.; Begun, D.J. Parallel Gene Expression Differences between Low and High Latitude Populations of Drosophila melanogaster and D. simulans. PLoS Genet. 2015, 11, e1005184. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.T.; Begun, D.J. Evidence of Spatially Varying Selection Acting on Four Chromatin-Remodeling Loci in Drosophila melanogaster. Genetics 2008, 179, 475–485. [Google Scholar] [CrossRef][Green Version]
- Williams, J.A.; Bell, J.B.; Carroll, S.B. Control of Drosophila wing and haltere development by the nuclear vestigial gene product. Genes Dev. 1991, 5, 2481–2495. [Google Scholar] [CrossRef] [PubMed]
- Lindsley, D.L.; Zimm, G.G. The Genome of Drosophila melanogaster; Academic Press: Cambridge, MA, USA, 2012; ISBN 978-0-323-13984-7. [Google Scholar]
- Kim, J.; Sebring, A.; Esch, J.J.; Kraus, M.E.; Vorwerk, K.; Magee, J.; Carroll, S.B. Integration of positional signals and regulation of wing formation and identity by Drosophila vestigial gene. Nature 1996, 382, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Silber, J.; Flagiello, D.; Cossard, R.; Zider, A.; Becker, J.L. Temperature regulates expression of the Drosophila vestigial gene only in mutant wing discs. Mol. Genet. Genom. 1997, 256, 328–332. [Google Scholar] [CrossRef]
- Arvidsson, S.; Kwasniewski, M.; Riaño-Pachón, D.M.; Mueller-Roeber, B. QuantPrime—A flexible tool for reliable high-throughput primer design for quantitative PCR. BMC Bioinf. 2008, 9, 465. [Google Scholar] [CrossRef]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Lack, J.B.; Lange, J.D.; Tang, A.D.; Corbett-Detig, R.B.; Pool, J.E. A Thousand Fly Genomes: An Expanded Drosophila Genome Nexus. Mol. Boil. Evol. 2016, 33, 3308–3313. [Google Scholar] [CrossRef]
- Kapopoulou, A.; Kapun, M.; Pavlidis, P.; Pieper, B.; Wilches, R.; Stephan, W.; Laurent, S. Early split between African and European populations of Drosophila melanogaster. bioRxiv 2018, 340422. [Google Scholar]
- Hudson, R.R.; Slatkin, M.; Maddison, W.P. Estimation of Levels of Gene Flow from DNA Sequence Data. Genetics 1992, 132, 583–589. [Google Scholar] [PubMed]
- Nègre, N.; Brown, C.D.; Shah, P.K.; Kheradpour, P.; Morrison, C.A.; Henikoff, J.G.; Feng, X.; Ahmad, K.; Russell, S.; White, R.A.H.; et al. A Comprehensive Map of Insulator Elements for the Drosophila Genome. PLoS Genet. 2010, 6, e1000814. [Google Scholar] [CrossRef] [PubMed]
- Climate data for cities worldwide—Climate-Data.org. OpenStreetMap contributor. Available online: https://en.climate-data.org/.
- Chintapalli, V.R.; Wang, J.; Dow, J.A.T. Using FlyAtlas to identify better Drosophila melanogaster models of human disease. Nat. Genet. 2007, 39, 715–720. [Google Scholar] [CrossRef]
- Fauvarque, M.O.; Dura, J.M. polyhomeotic regulatory sequences induce developmental regulator-dependent variegation and targeted P-element insertions in Drosophila. Genes Dev. 1993, 7, 1508–1520. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.; Rastelli, L.; Pirrotta, V. A Polycomb response element in the Ubx gene that determines an epigenetically inherited state of repression. EMBO J. 1994, 13, 2553–2564. [Google Scholar] [CrossRef] [PubMed]
- Zink, D.; Paro, R. Drosophila Polycomb-group regulated chromatin inhibits the accessibility of a trans-activator to its target DNA. EMBO J. 1995, 14, 5660–5671. [Google Scholar] [CrossRef]
- Bantignies, F.; Grimaud, C.; Lavrov, S.; Gabut, M.; Cavalli, G. Inheritance of Polycomb-dependent chromosomal interactions in Drosophila. Genome Res. 2003, 17, 2406–2420. [Google Scholar] [CrossRef]
- Gibert, J.-M.; Karch, F.; Schlötterer, C. Segregating Variation in the Polycomb Group Gene cramped Alters the Effect of Temperature on Multiple Traits. PLoS Genet. 2011, 7, 1001280. [Google Scholar] [CrossRef]
- Voigt, S.; Laurent, S.; Litovchenko, M.; Stephan, W. Positive Selection at the Polyhomeotic Locus Led to Decreased Thermosensitivity of Gene Expression in Temperate Drosophila melanogaster. Genetics 2015, 200, 591–599. [Google Scholar] [CrossRef]
- Kassis, J.A.; Brown, J.L. Chapter three—Polycomb group response elements in Drosophila and vertebrates. In Advances in Genetics; Friedmann, T., Dunlap, J.C., Goodwin, S.F., Eds.; Academic Press: Cambridge, MA, USA, 2013; Volume 81, pp. 83–118. [Google Scholar]
- Sudarsan, V.; Anant, S.; Guptan, P.; VijayRaghavan, K.; Skaer, H. Myoblast Diversification and Ectodermal Signaling in Drosophila. Dev. Cell 2001, 1, 829–839. [Google Scholar] [CrossRef]
- Sigrist, C.J.A.; Pirrotta, V. Chromatin insulator elements block the silencing of a target gene by the Drosophila Polycomb response element (PRE) but allow trans interactions between PREs on different chromosomes. Genetics 1997, 147, 209–221. [Google Scholar] [PubMed]
- Comet, I.; Schuettengruber, B.; Sexton, T.; Cavalli, G. A chromatin insulator driving three-dimensional Polycomb response element (PRE) contacts and Polycomb association with the chromatin fiber. Proc. Natl. Acad. Sci. USA 2011, 108, 2294–2299. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-B.; Müller, M.; Bahechar, I.A.; Kyrchanova, O.; Ohno, K.; Georgiev, P.; Pirrotta, V. Insulators, not Polycomb response elements, are required for long-range interactions between Polycomb targets in Drosophila melanogaster. Mol. Cell. Biol. 2011, 31, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-B.; Ohno, K.; Gui, H.; Pirrotta, V. Insulators Target Active Genes to Transcription Factories and Polycomb-Repressed Genes to Polycomb Bodies. PLoS Genet. 2013, 9, e1003436. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Population Sample | Latitude | Longitude | Altitude (m) | Mean Annual Temperature (°C) | Minimum Annual Temperature (°C) | Maximum Annual Temperature (°C) | Climate |
|---|---|---|---|---|---|---|---|
| Umeå, Sweden (SU) | 63.83 | 20.26 | 12 | 2.7 | −1.6 | 7.0 | cold-temperate |
| Leiden, The Netherlands (NL) | 52.17 | 4.48 | 0 | 9.5 | 6.4 | 12.6 | warm-temperate |
| Lyon, France (FR) | 45.76 | 4.84 | 175 | 11.6 | 6.9 | 16.4 | warm-temperate |
| Gikongoro, Rwanda (RG) | −2.49 | 28.92 | 1927 | 17.6 | 12.4 | 23.0 | warm-temperate |
| Lake Kariba, Zimbabwe (ZK) | −16.52 | 28.80 | 619 | 25.5 | 18.7 | 31.9 | tropical |
| Siavonga, Zambia (ZI) | −16.54 | 28.72 | 530 | 25.2 | 18.6 | 31.9 | tropical |
| Χ2 | df | p | |
|---|---|---|---|
| Adult | |||
| Intercept | 32.6367 | 1 | <0.001 *** |
| Rearing temperature | 9.9029 | 1 | 0.001650 ** |
| Climate of origin | 19.7739 | 2 | <0.001 *** |
| Rearing temperature x climate of origin | 12.7246 | 2 | 0.001725 ** |
| wL3 wing imaginal discs | |||
| Intercept | 1.4627 | 1 | 0.2265 |
| Rearing temperature | 0.0066 | 1 | 0.9354 |
| Climate of origin | 0.3989 | 2 | 0.8192 |
| Rearing temperature x climate of origin | 1.4034 | 2 | 0.4957 |
| wL3 brains | |||
| Intercept | 19.8352 | 1 | <0.001 *** |
| Rearing temperature | 2.0164 | 1 | 0.1556 |
| Climate of origin | 1.6951 | 2 | 0.4285 |
| Rearing temperature x climate of origin | 1.0706 | 2 | 0.5855 |
| Gene | Gene Expression Ratio 17/28 °C | |||||
|---|---|---|---|---|---|---|
| Sweden (SU) | The Netherlands (NL) | France (FR) | Rwanda (RG) | Zimbabwe (ZK) | Zambia (ZI) | |
| vg | 1.76 * | 1.82 | 2.28 ** | 1.88 ** | 3.68 ** | 3.02 ** |
| Aats-asp | 1.15 | 1.52 | 0.90 | 0.92 | 1.01 | 1.17 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voigt, S.; Erpf, A.C.; Stephan, W. Decreased Temperature Sensitivity of Vestigial Gene Expression in Temperate Populations of Drosophila melanogaster. Genes 2019, 10, 498. https://doi.org/10.3390/genes10070498
Voigt S, Erpf AC, Stephan W. Decreased Temperature Sensitivity of Vestigial Gene Expression in Temperate Populations of Drosophila melanogaster. Genes. 2019; 10(7):498. https://doi.org/10.3390/genes10070498
Chicago/Turabian StyleVoigt, Susanne, Anna Christina Erpf, and Wolfgang Stephan. 2019. "Decreased Temperature Sensitivity of Vestigial Gene Expression in Temperate Populations of Drosophila melanogaster" Genes 10, no. 7: 498. https://doi.org/10.3390/genes10070498
APA StyleVoigt, S., Erpf, A. C., & Stephan, W. (2019). Decreased Temperature Sensitivity of Vestigial Gene Expression in Temperate Populations of Drosophila melanogaster. Genes, 10(7), 498. https://doi.org/10.3390/genes10070498