A QIL1 Variant Associated with Ventricular Arrhythmias and Sudden Cardiac Death in the Juvenile Rhodesian Ridgeback Dog

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
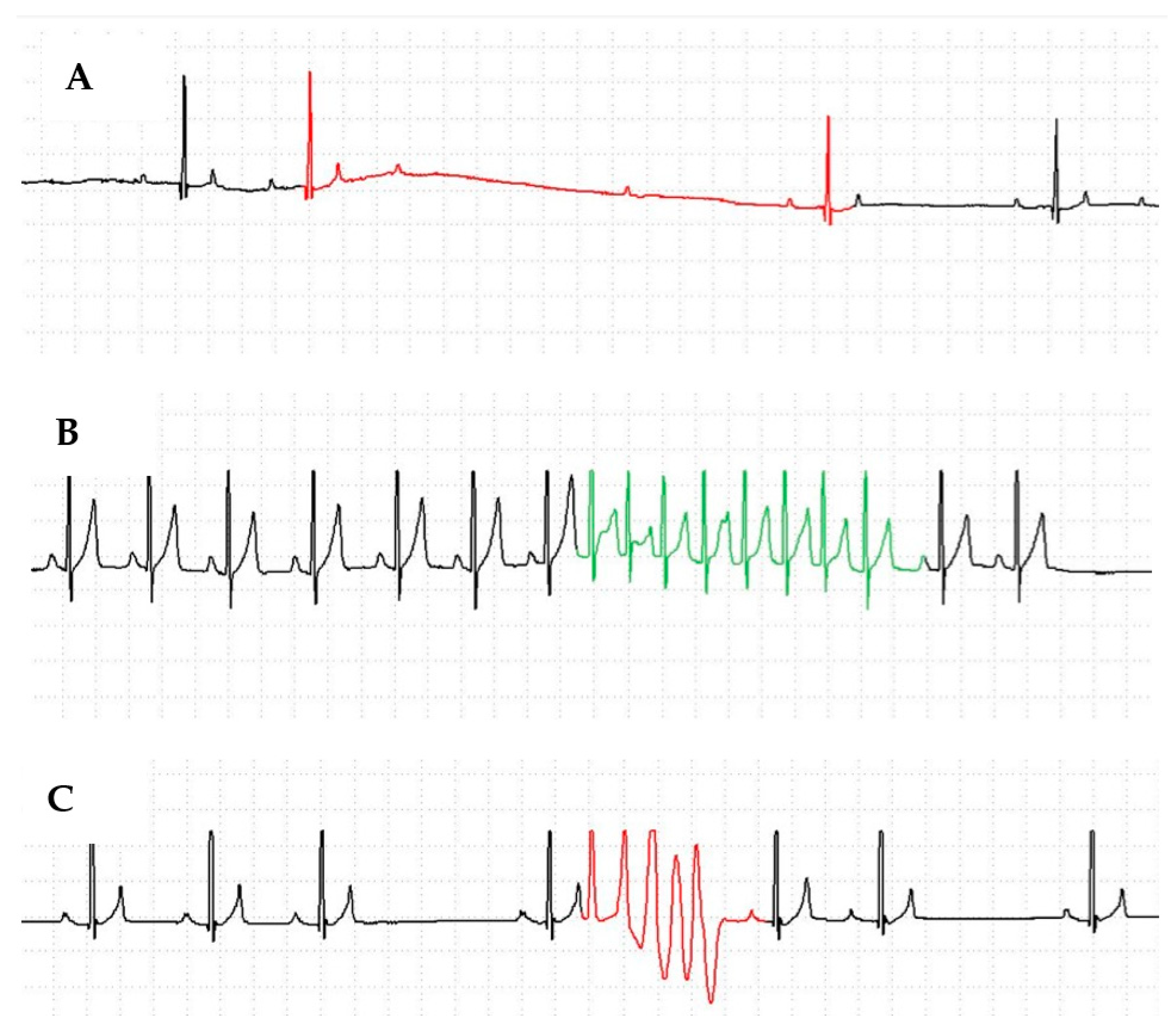
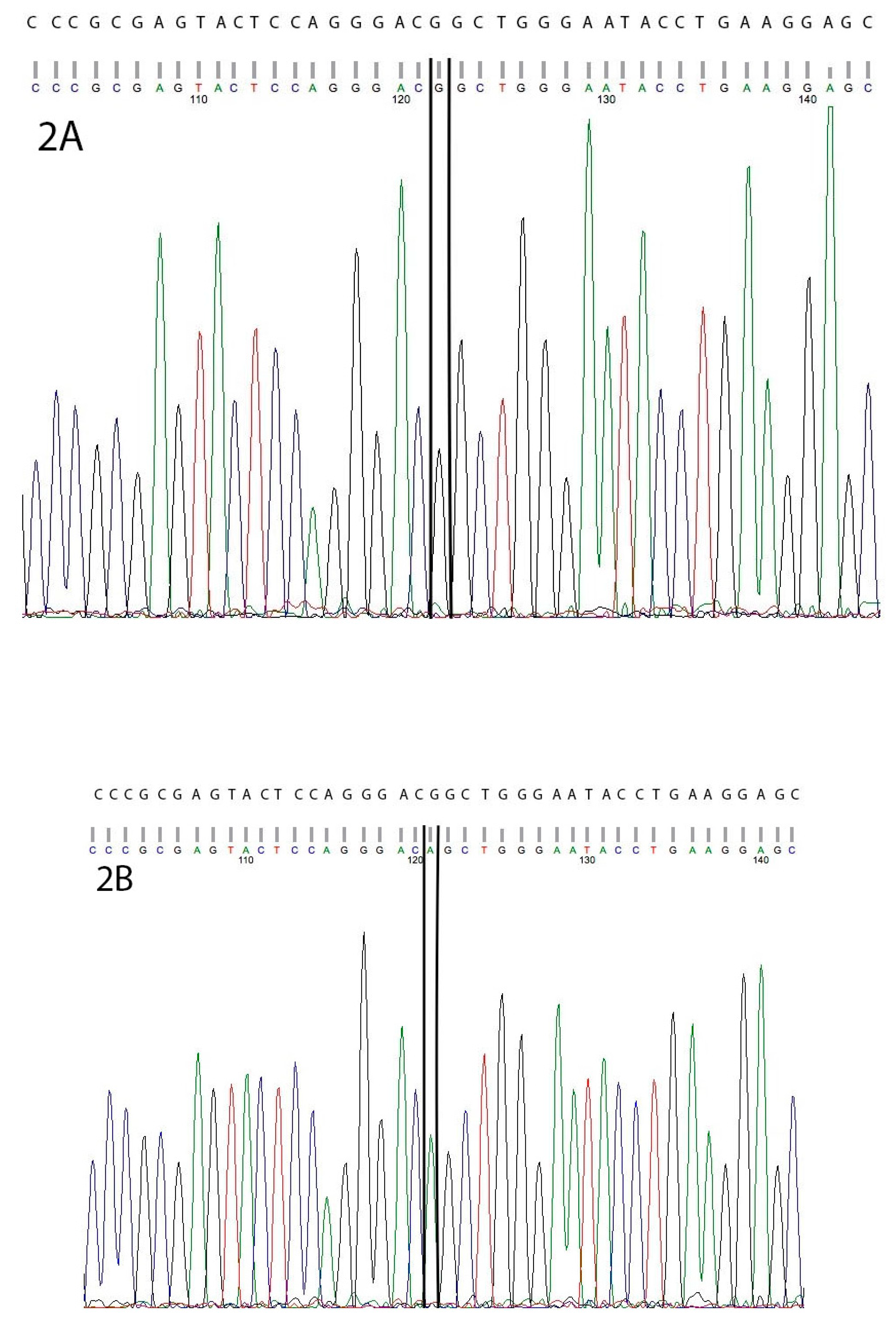
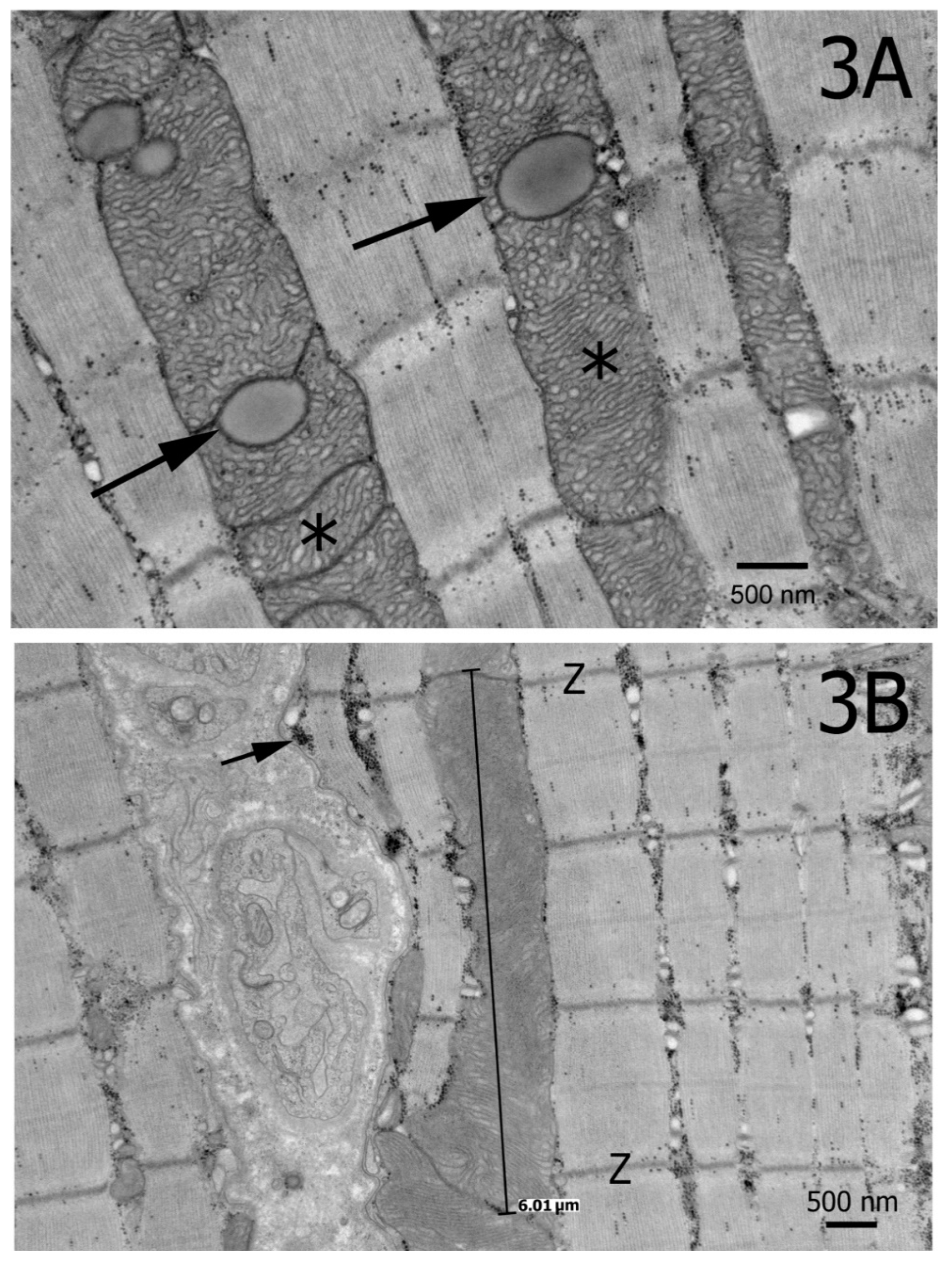
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zeharia, A.; Friedman, J.R.; Tobar, A.; Saada, A.; Konen, O.; Fellig, Y.; Shaag, A.; Nunnari, J.; Elpeleg, O. Mitochondrial hepato-encephalopathy due to deficiency of QIL1/MIC13 (C19orf70), a MICOS complex subunit. Eur. J. Hum. Genet. 2016, 24, 1778–1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerbes, R.M.; Höß, P.; Pfanner, N.; Van Der Laan, M.; Bohnert, M. Distinct Roles of Mic12 and Mic27 in the Mitochondrial Contact Site and Cristae Organizing System. J. Mol. Biol. 2016, 428, 1485–1492. [Google Scholar] [CrossRef] [PubMed]
- Vogel, F.; Bornhövd, C.; Neupert, W.; Reichert, A.S. Dynamic subcompartmentalization of the mitochondrial inner membrane. J. Cell Biol. 2006, 175, 237–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarani, V.; McNeill, E.M.; Paulo, J.A.; Huttlin, E.L.; Fröhlich, F.; Gygi, S.P.; Van Vactor, D.; Wade Harper, J. QIL1 is a novel mitochondrial protein required for MICOS complex stability and cristae morphology. Elife 2015, 4, 1–23. [Google Scholar] [CrossRef] [PubMed]
- El-Hattab, A.W.; Scaglia, F. Mitochondrial Cardiomyopathies. Front. Cardiovasc. Med. 2016, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Guarani, V.; Jardel, C.; Chretien, D.; Lombes, A.; Benit, P.; Labasse, C.; Lacene, E.; Bourillon, A.; Imbard, A.; Benoist, J.F.; et al. QIL1 mutation causes MICOS disassembly and early onset fatal mitochondrial encephalopathy with liver disease. Elife 2016, 5, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Godiker, J.; Gruneberg, M.; DuChesne, I.; Reunert, J.; Rust, S.; Westermann, C.; Wada, Y.; Classen, G.; Langhans, C.D.; Schlingmann, K.; et al. QIL1-dependent assembly of MICOS complex-lethal mutation in C19ORF70 resulting in liver disease and severe neurological retardation. J. Hum. Genet. 2018, 63, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Meurs, K.M.; Weidman, J.A.; Rosenthal, S.L.; Lahmers, K.K.; Friedenberg, S.G. Ventricular arrhythmias in Rhodesian Ridgebacks with a family history of sudden death and results of a pedigree analysis for potential inheritance patterns. J. Am. Vet. Med. Assoc. 2016, 248, 1135–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meurs, K.M.; Spier, A.W.; Wright, N.A.; Hamlin, R.L. Use of ambulatory electrocardiography for detection of ventricular premature complexes in healthy dogs. J. Am. Vet. Med. Assoc. 2001, 218, 1291–1292. [Google Scholar] [CrossRef]
- Thomas, W.P.; Gaber, C.E.; Jacobs, G.J.; Kaplan, P.M.; Lombard, C.W.; Sydney Moise, N.; Moses, B.L. Recommendations for standards in transthoracic Two-Dimensional echocardiography in the dog and cat. Vet. Radiol. Ultrasound 1994, 35, 173–178. [Google Scholar] [CrossRef]
- Friedenberg, S.G.; Meurs, K.M. Genotype imputation in the domestic dog. Mamm. Genome 2016, 27, 485–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DePristo, M.A.; Banks, E.; Poplin, R.E.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data Aaron. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Van Der Auwera, G.; Carneiro, M.O.; Hartl, C.; Poplin, R.; del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From FastQ data to high confidence varant calls: The Genonme Analysis Toolkit best practices pipeline. Curr. Protoc. Bioinform. 2014, 11, 1–33. [Google Scholar]
- Mclaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R.; Han, J. Mitochondrial mutations in cardiac disorders. Adv. Exp. Med. Biol. 2017, 982, 81–111. [Google Scholar]
- Scaglia, F.; Towbin, J.A.; Craigen, W.J.; Belmont, J.W.; Smith, E.O.; Neish, S.R.; Ware, S.M.; Hunter, J.V.; Fernbach, S.D.; Vladutiu, G.D.; et al. Clinical spectrum, morbidity, and mortality in 113 pediatric patients With mitochondrial disease. Pediatrics 2004, 114, 925–931. [Google Scholar] [CrossRef]
- Finsterer, J.; Kothari, S. Cardiac manifestations of primary mitochondrial disorders. Int. J. Cardiol. 2014, 177, 754–763. [Google Scholar] [CrossRef]
- Montaigne, D.; Pentiah, A.D. Mitochondrial cardiomyopathy and related arrhythmias. Card. Electrophysiol. Clin. 2015, 7, 293–301. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Variant | Effect | p Value |
|---|---|---|---|
| ADCY3 | ENSCAFG00000004090 | Missense/Splice site | NP |
| g.19164291 G>A | |||
| AGRN | ENSCAFG00000019342 | Missense | 0.25 |
| g.56260122 C>A | |||
| BSCL2 | ENSCAFG00000023629 | Initiator codon variant | 0.62 |
| g.53960802 A>G | |||
| FASTKD3 | ENSCAFG00000010129 | Missense | 0.63 |
| g.6105469 C>T | |||
| FAT1 | ENSCAFG00000007273 | Missense | 0.16 |
| g.44199325 G>T | |||
| HCN4 | ENSCAFG00000031809 | Frameshift | NP |
| g.36680881_36680885del | |||
| LAMA4 | ENSCAFG00000004043 | In frame deletion | NP |
| g.68553193_68553195del | |||
| MYO9B | ENSCAFG00000015532 | Missense | 0.35 |
| g.45524791 C>T | |||
| PIEZO2 | ENSCAFG00000018761 | Frameshift | NP |
| g.76508892_76508892insG | |||
| PRDM8 | ENSCAFG00000008881 | Inframe insertion | NP |
| g.4450426_4450427insG | |||
| C19orf70 | ENSCAFG00000018796 | Missense | 0.04 |
| g.54343438 G>A | |||
| SMTNL1 | ENSCAFG00000007843 | Missense | >0.99 |
| g.38627683 C>T | |||
| SORBS2 | ENSCAFG00000007475 | Missense | 0.035 |
| g.45035993 G>A |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meurs, K.M.; Friedenberg, S.G.; Olby, N.J.; Condit, J.; Weidman, J.; Rosenthal, S.; Shelton, G.D. A QIL1 Variant Associated with Ventricular Arrhythmias and Sudden Cardiac Death in the Juvenile Rhodesian Ridgeback Dog. Genes 2019, 10, 168. https://doi.org/10.3390/genes10020168
Meurs KM, Friedenberg SG, Olby NJ, Condit J, Weidman J, Rosenthal S, Shelton GD. A QIL1 Variant Associated with Ventricular Arrhythmias and Sudden Cardiac Death in the Juvenile Rhodesian Ridgeback Dog. Genes. 2019; 10(2):168. https://doi.org/10.3390/genes10020168
Chicago/Turabian StyleMeurs, Kathryn M., Steven G. Friedenberg, Natasha J. Olby, Julia Condit, Jess Weidman, Steve Rosenthal, and G. Diane Shelton. 2019. "A QIL1 Variant Associated with Ventricular Arrhythmias and Sudden Cardiac Death in the Juvenile Rhodesian Ridgeback Dog" Genes 10, no. 2: 168. https://doi.org/10.3390/genes10020168
APA StyleMeurs, K. M., Friedenberg, S. G., Olby, N. J., Condit, J., Weidman, J., Rosenthal, S., & Shelton, G. D. (2019). A QIL1 Variant Associated with Ventricular Arrhythmias and Sudden Cardiac Death in the Juvenile Rhodesian Ridgeback Dog. Genes, 10(2), 168. https://doi.org/10.3390/genes10020168