Insight into the B3Transcription Factor Superfamily and Expression Profiling of B3 Genes in Axillary Buds after Topping in Tobacco (Nicotiana tabacum L.)

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Classification of B3TF Family Members in Tobacco
2.2. Multiple Sequence Alignments and Phylogenetic Analysis
2.3. Chromosomal Location
2.4. Gene Structure and Conserved Motif Analysis
2.5. Expression Pattern Analysis of Tobacco B3 Genes
2.6. Plant Growth, Topping Treatment, and Quantitative Real-Time PCR Analysis
3. Results
3.1. Identification of B3 Genes in the Tobacco Genome
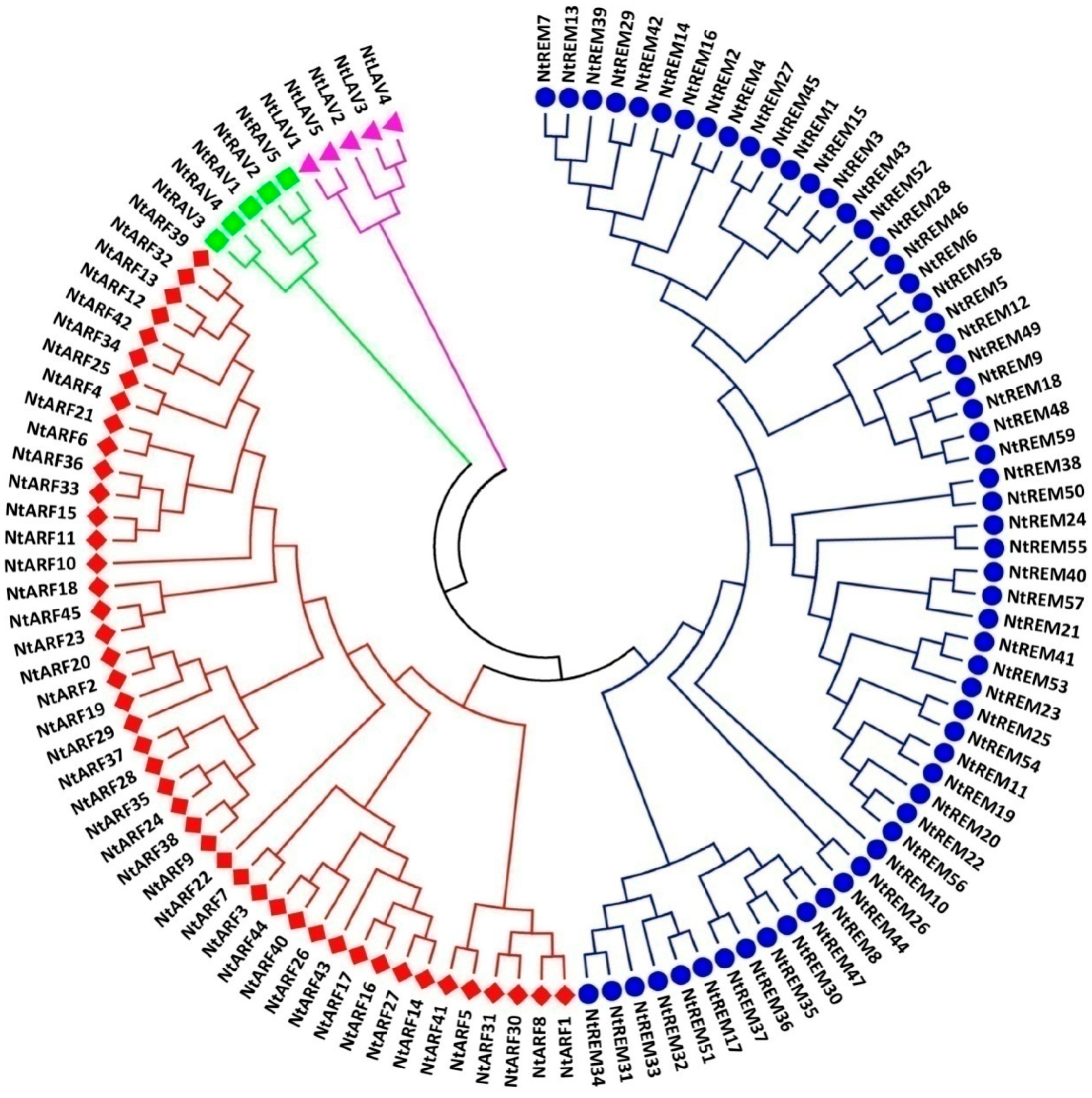
3.2. Phylogenetic Relationship Analysis of B3 Proteins in Tobacco
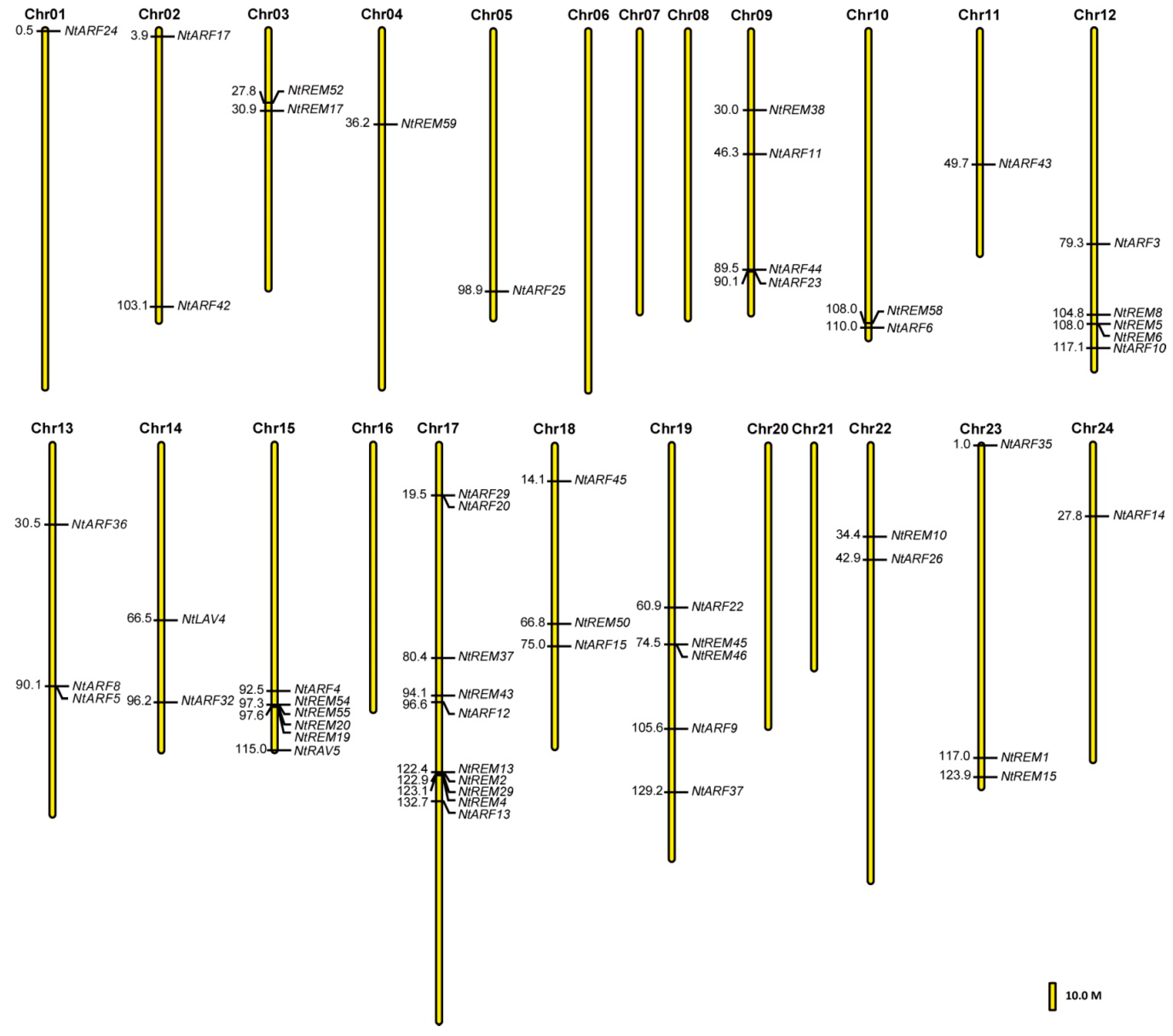
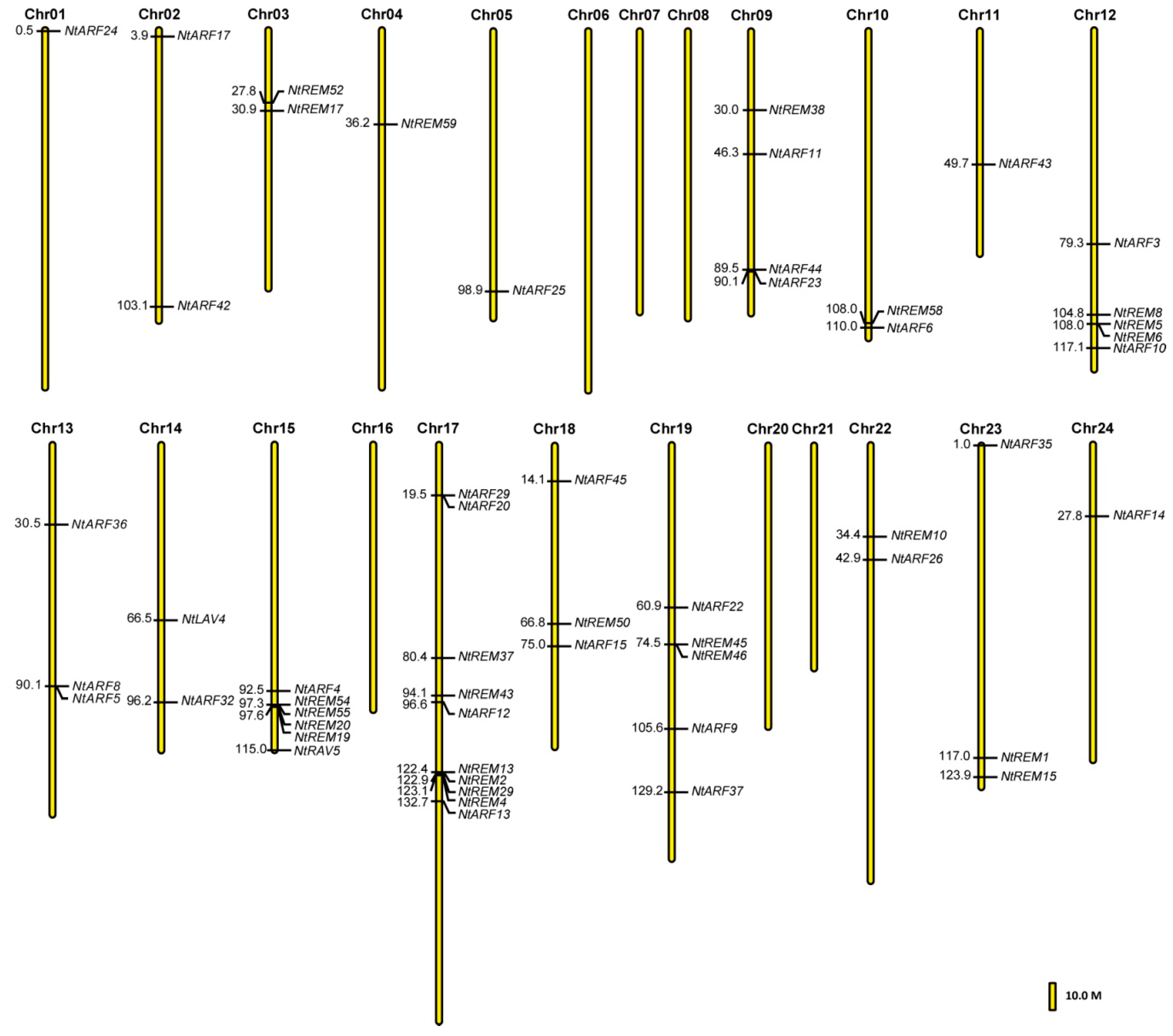
3.3. Chromosomal Location of B3 Genes in the Tobacco Genome
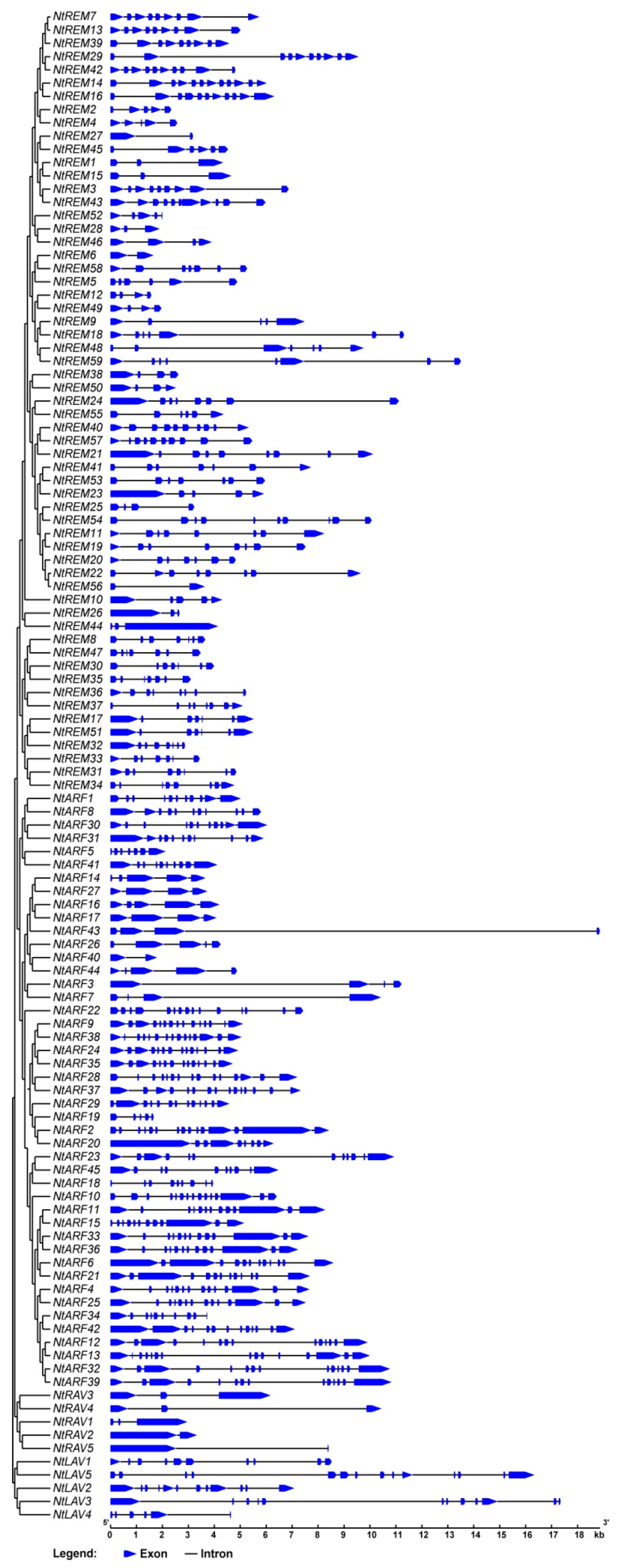
3.4. Gene Structure and Conserved Motif Analysis of B3 Superfamily Proteins
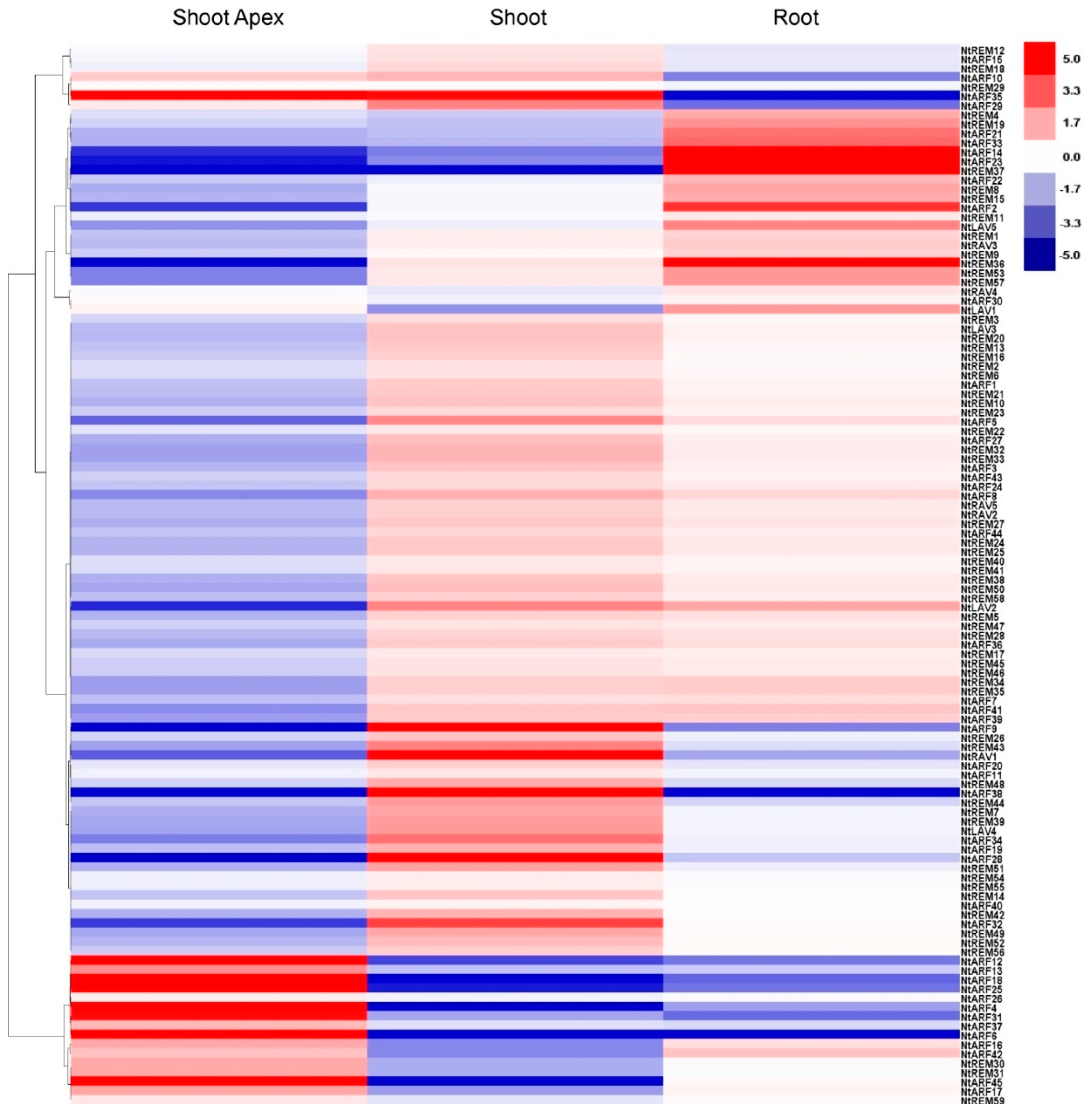
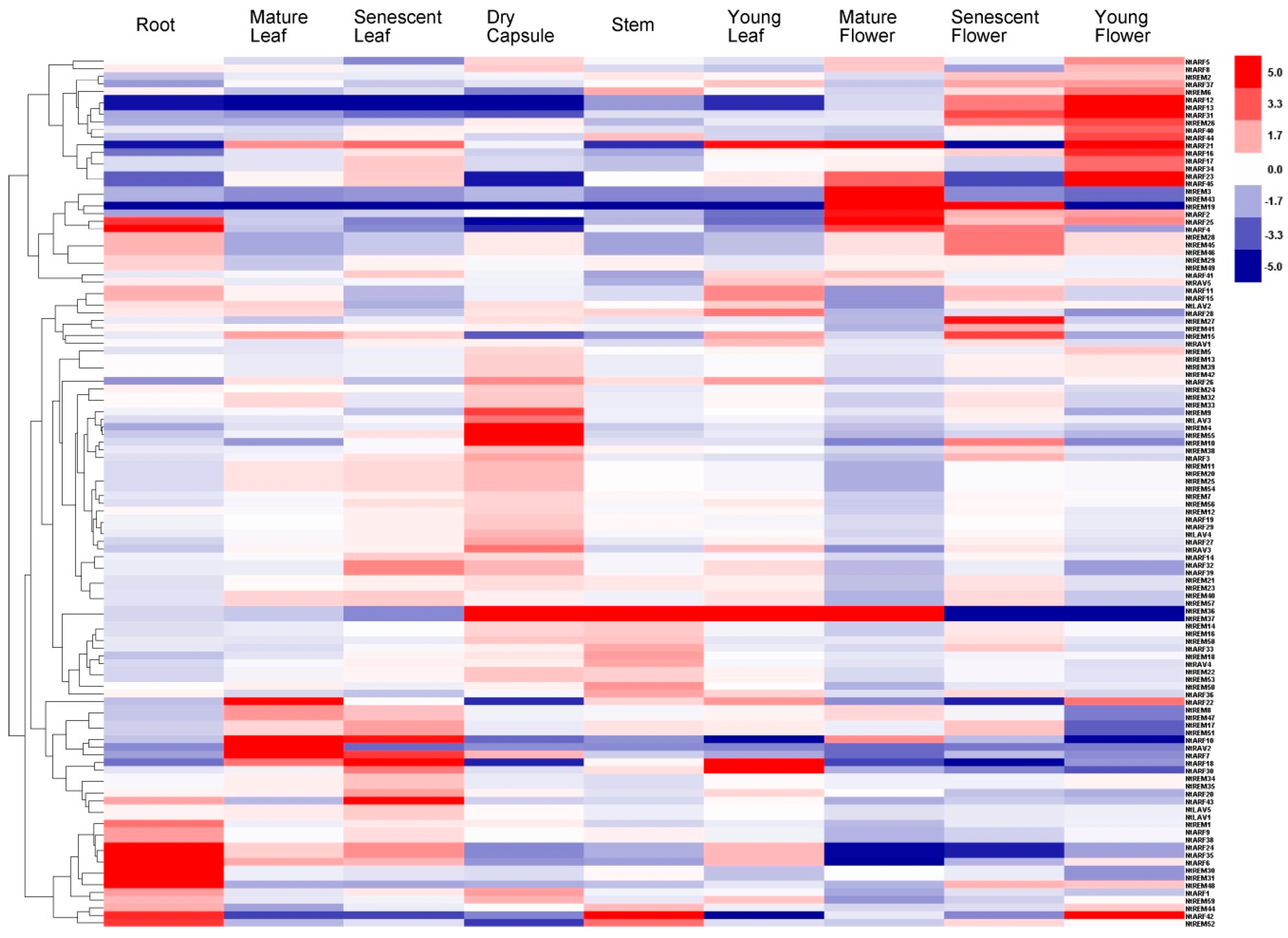
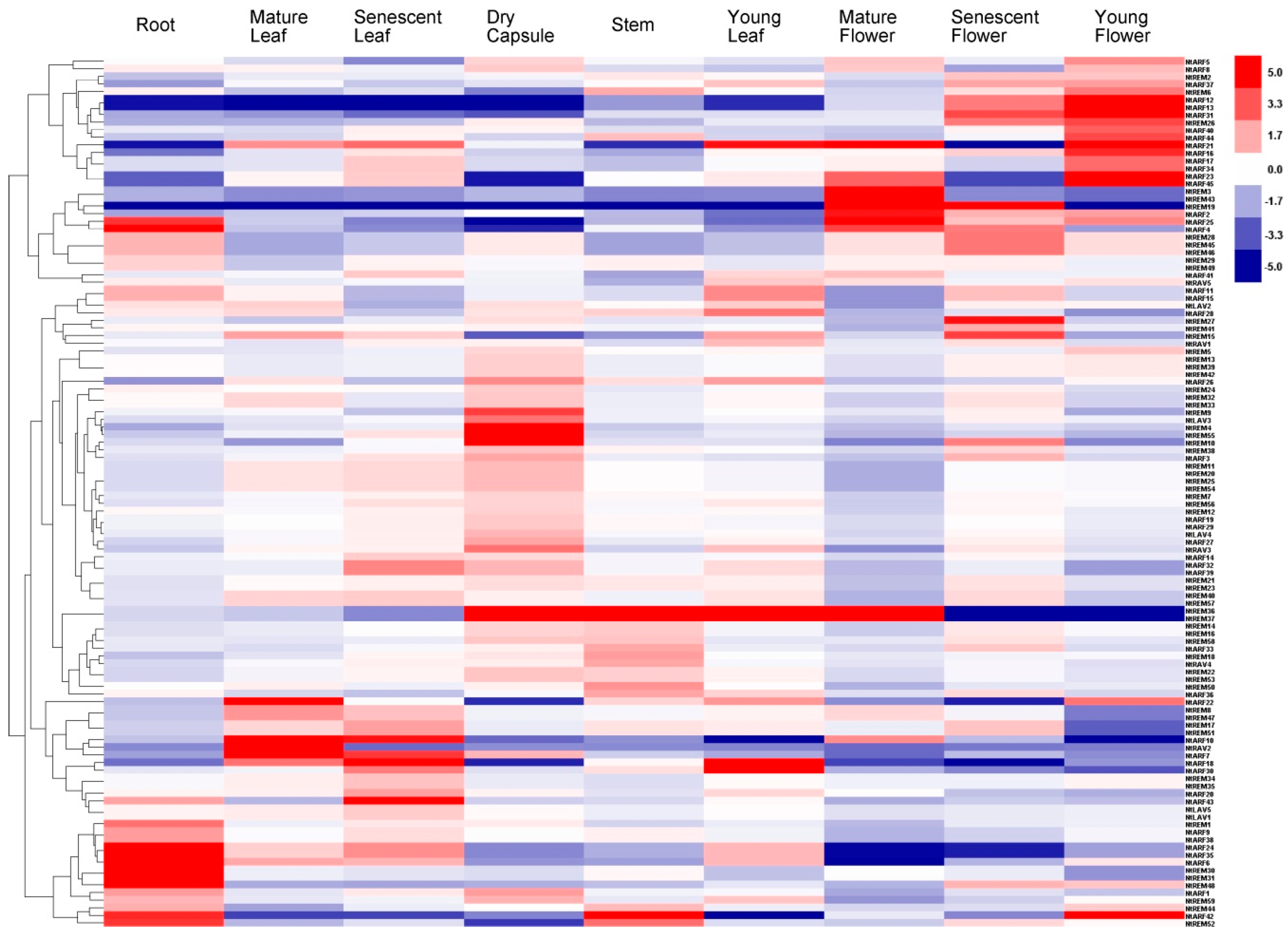
3.5. Tissue-Specific Expression Profiling of the B3 Genes in Tobacco
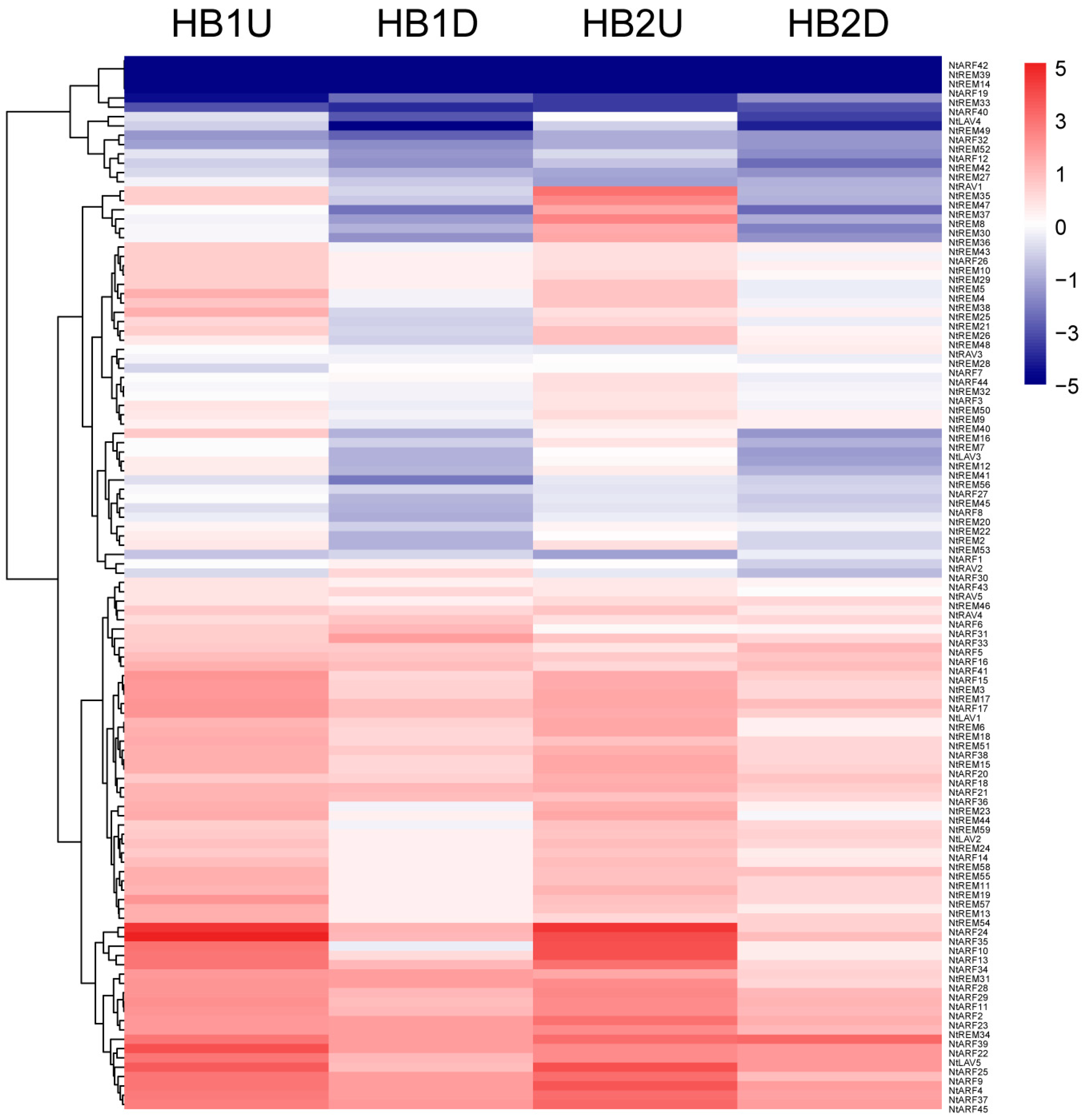
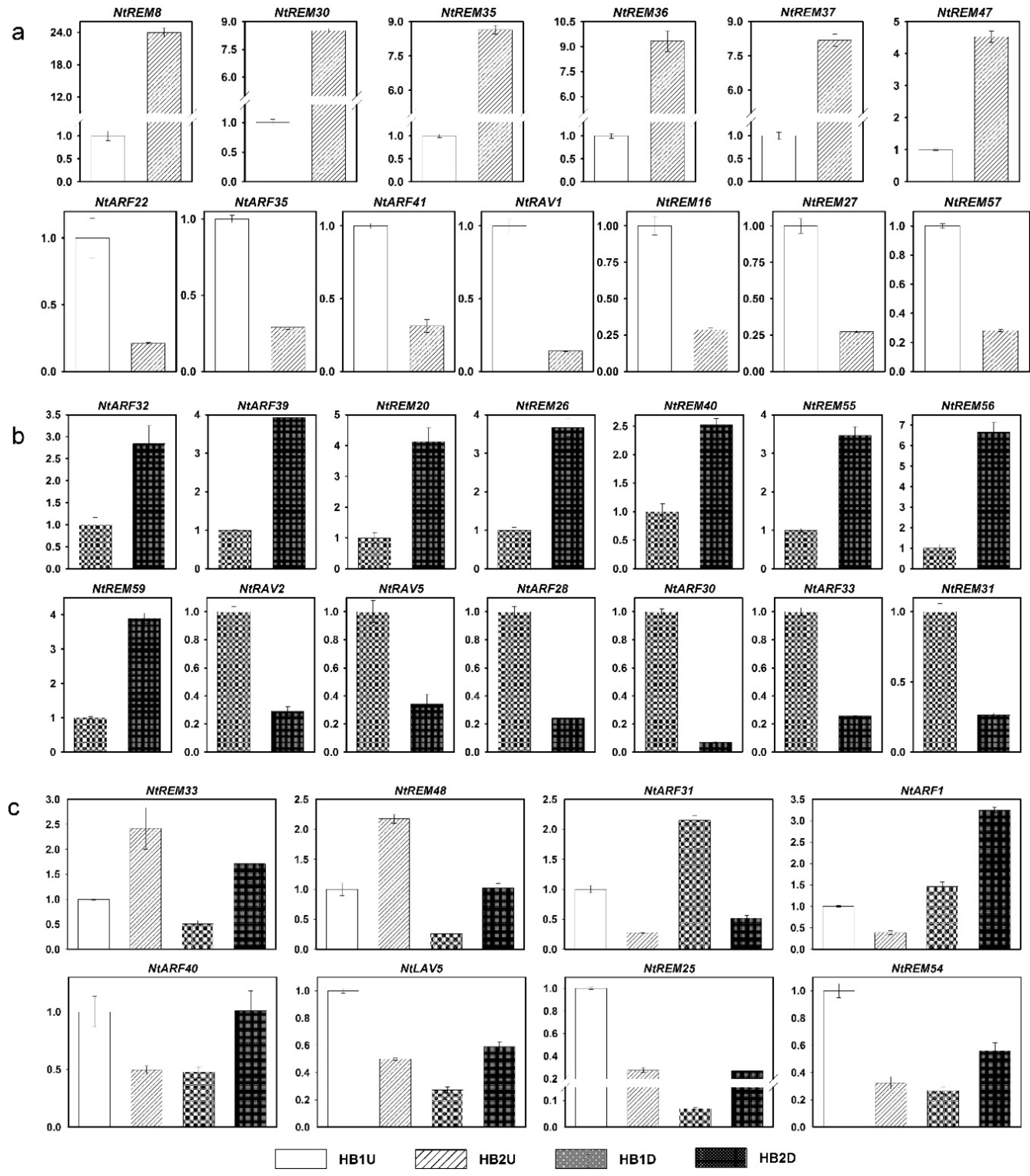
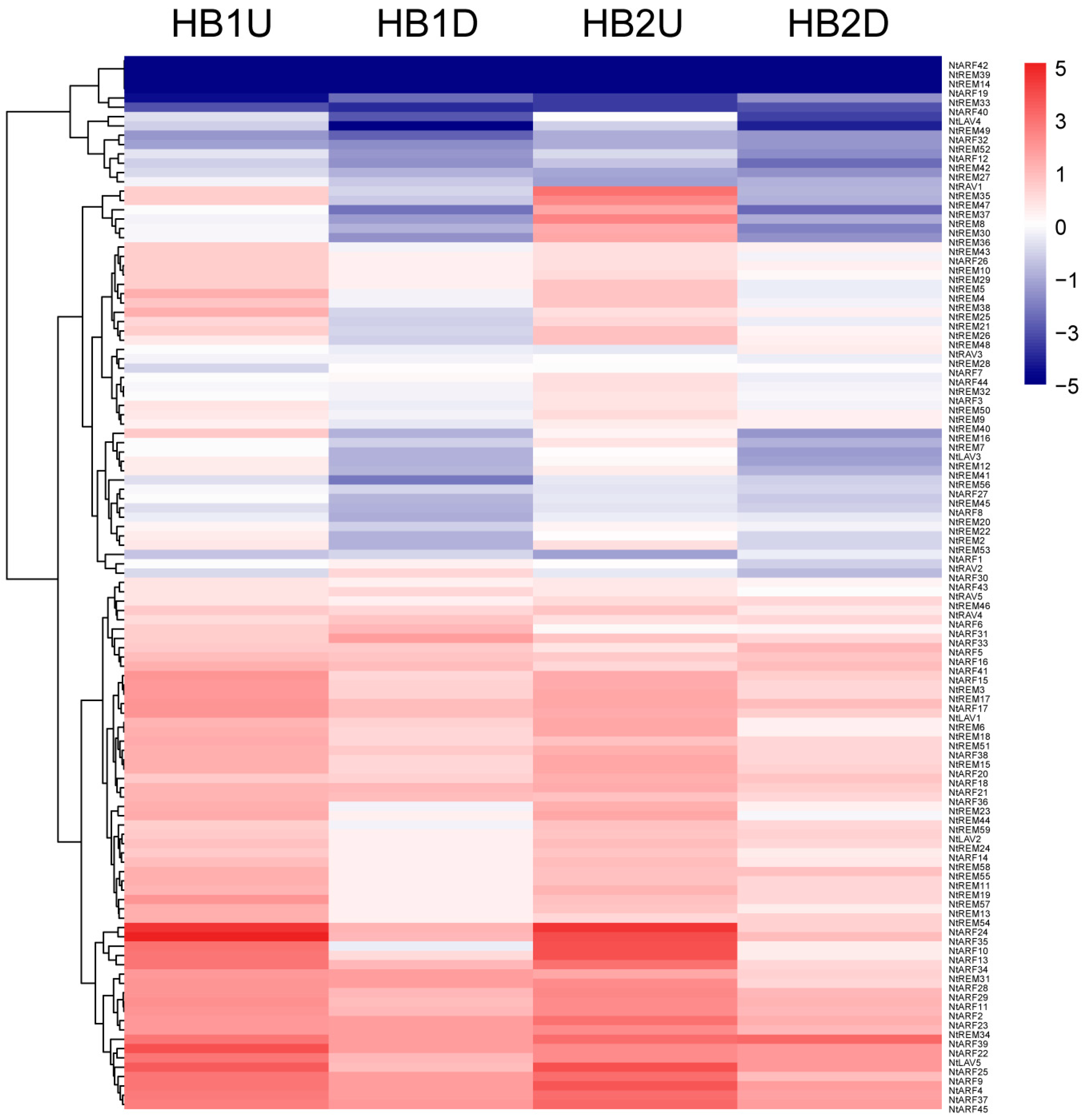
3.6. Expression Profiles of B3 Genes in Tobacco Axillary Buds in Response to Topping
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.Z.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; et al. Arabidopsis Transcription Factors: Genome-Wide Comparative Analysis Among Eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Zhang, H.; Kong, L.; Gao, G.; Luo, J. PlantTFDB 3.0: A portal for the functional and evolutionary study of plant transcription factors. Nucleic Acids Res. 2014, 42, D1182–D1187. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Liu, T.; Tian, C.; Sun, S.; Li, J.; Chen, M. Transcription factors in rice: A genome-wide comparative analysis between monocots and eudicots. Plant Mol. Biol. 2005, 59, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, K.; Peterson, K.; Jack, T. The plant B3 superfamily. Trends Plant Sci. 2008, 13, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Kigawa, T.; Seki, M.; Shinozaki, K.; Yokoyama, S. DNA-binding domains of plant-specific transcription factors: Structure, function, and evolution. Trends Plant Sci. 2013, 18, 267–276. [Google Scholar] [CrossRef] [PubMed]
- McCarty, D.R.; Hattori, T.; Carson, C.B.; Vasil, V.; Lazar, M.; Vasil, I.K. The Viviparous-1 developmental gene of maize encodes a novel transcriptional activator. Cell 1991, 66, 895–905. [Google Scholar] [CrossRef]
- Giraudat, J.; Hauge, B.M.; Valon, C.; Smalle, J.; Parcy, F.; Goodman, H.M. Isolation of the Arabidopsis ABI3 gene by positional cloning. Plant Cell 1992, 4, 1251–1261. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Kao, C.Y.; McCarty, D.R. The conserved B3 domain of VIVIPAROUS1 has a cooperative DNA binding activity. Plant Cell 1997, 9, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.Y.; Weselake, R.J. Genome-wide identification and analysis of the B3 superfamily of transcription factors in Brassicaceae and major crop plants. TAG. Theoretical and applied genetics. Theoretische und AngewandteGenetik 2013, 126, 1305–1319. [Google Scholar] [CrossRef]
- Suzuki, M.; McCarty, D.R. Functional symmetry of the B3 network controlling seed development. Curr. Opin. Plant Biol. 2008, 11, 548–553. [Google Scholar] [CrossRef] [PubMed]
- To, A.; Valon, C.; Savino, G.; Guilleminot, J.; Devic, M.; Giraudat, J.; Parcy, F. A network of local and redundant gene regulation governs Arabidopsis seed maturation. Plant Cell 2006, 18, 1642–1651. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, L.; Van Wuytswinkel, O.; Castelain, M.; Bellini, C. Combined networks regulating seed maturation. Trends Plant Sci. 2007, 12, 294–300. [Google Scholar] [CrossRef]
- Braybrook, S.A.; Harada, J.J. LECs go crazy in embryo development. Trends Plant Sci. 2008, 13, 624–630. [Google Scholar] [CrossRef]
- Santos-Mendoza, M.; Dubreucq, B.; Baud, S.; Parcy, F.; Caboche, M.; Lepiniec, L. Deciphering gene regulatory networks that control seed development and maturation in Arabidopsis. Plant J. Cell Mol. Biol. 2008, 54, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.; Kapoor, S.; Tyagi, A.K. Transcription factors regulating the progression of monocot and dicot seed development. BioEssays News Rev. Mol. Cell. Dev. Biol. 2011, 33, 189–202. [Google Scholar] [CrossRef]
- Boulard, C.; Fatihi, A.; Lepiniec, L.; Dubreucq, B. Regulation and evolution of the interaction of the seed B3 transcription factors with NF-Y subunits. Biochim. Biophys. Acta 2017, 1860, 1069–1078. [Google Scholar] [CrossRef]
- Carbonero, P.; Iglesias-Fernandez, R.; Vicente-Carbajosa, J. The AFL subfamily of B3 transcription factors: Evolution and function in angiosperm seeds. J. Exp. Bot. 2017, 68, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Horstman, A.; Li, M.; Heidmann, I.; Weemen, M.; Chen, B.; Muino, J.M.; Angenent, G.C.; Boutilier, K. The BABY BOOM Transcription Factor Activates the LEC1-ABI3-FUS3-LEC2 Network to Induce Somatic Embryogenesis. Plant Physiol. 2017, 175, 848–857. [Google Scholar] [CrossRef]
- Tsukagoshi, H.; Saijo, T.; Shibata, D.; Morikami, A.; Nakamura, K. Analysis of a sugar response mutant of Arabidopsis identified a novel B3 domain protein that functions as an active transcriptional repressor. Plant Physiol. 2005, 138, 675–685. [Google Scholar] [CrossRef]
- Tsukagoshi, H.; Morikami, A.; Nakamura, K. Two B3 domain transcriptional repressors prevent sugar-inducible expression of seed maturation genes in Arabidopsis seedlings. Proc. Natl. Acad. Sci. USA 2007, 104, 2543–2547. [Google Scholar] [CrossRef]
- Suzuki, M.; Wang, H.H.; McCarty, D.R. Repression of the Leafy Cotyledon 1/B3 regulatory network in plant embryo development by VP1/ABSCISIC Acid Insensitive 3-LIKE B3 genes. Plant Physiol. 2007, 143, 902–911. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; McCarty, D.R.; Suzuki, M. Distinct roles of LAFL network genes in promoting the embryonic seedling fate in the absence of VAL repression. Plant Physiol. 2013, 163, 1293–1305. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Bratzel, F.; Hohmann, N.; Koch, M.; Turck, F.; Calonje, M. VAL- and AtBMI1-mediated H2Aub initiate the switch from embryonic to postgerminative growth in Arabidopsis. Curr. Biol. CB 2013, 23, 1324–1329. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Tan, B.; Luo, M.; Li, Y.; Liu, C.; Chen, C.; Yu, C.W.; Yang, S.; Dong, S.; Ruan, J.; et al. HISTONE DEACETYLASE19 interacts with HSL1 and participates in the repression of seed maturation genes in Arabidopsis seedlings. Plant Cell 2013, 25, 134–148. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Cui, Y. A DNA element that remembers winter. Nat. Genet. 2016, 48, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Questa, J.I.; Song, J.; Geraldo, N.; An, H.; Dean, C. Arabidopsis transcriptional repressor VAL1 triggers Polycomb silencing at FLC during vernalization. Science 2016, 353, 485–488. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Luo, X.; Li, Z.; Yang, W.; Wang, Y.; Liu, R.; Du, J.; He, Y. A cis cold memory element and a trans epigenome reader mediate Polycomb silencing of FLC by vernalization in Arabidopsis. Nat. Genet. 2016, 48, 1527–1534. [Google Scholar] [CrossRef] [PubMed]
- Magnani, E.; Sjolander, K.; Hake, S. From endonucleases to transcription factors: Evolution of the AP2 DNA binding domain in plants. Plant Cell 2004, 16, 2265–2277. [Google Scholar] [CrossRef]
- Kim, S.; Soltis, P.S.; Wall, K.; Soltis, D.E. Phylogeny and domain evolution in the APETALA2-like gene family. Mol. Biol. Evol. 2006, 23, 107–120. [Google Scholar] [CrossRef]
- Romanel, E.A.; Schrago, C.G.; Couñago, R.M.; Russo, C.A.; Alves-Ferreira, M. Evolution of the B3 DNA binding superfamily: New insights into REM family gene diversification. PLoS ONE 2009, 4, e5791. [Google Scholar] [CrossRef]
- Wang, Y.J.; Wang, S.X.; Deng, D.X.; Bian, Y.L.; Zhang, R.; Yin, Z.T. Systematic analysis of plant-specific B3 domain-containing proteins based on the genome resources of 11 sequenced species. Mol. Biol. Rep. 2012, 39, 16. [Google Scholar] [CrossRef] [PubMed]
- Kagaya, Y.; Ohmiya, K.; Hattori, T. RAV1, a novel DNA-binding protein, binds to bipartite recognition sequence through two distinct DNA-binding domains uniquely found in higher plants. Nucleic Acids Res. 1999, 27, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Castillejo, C.; Pelaz, S. The balance between Constans and Tempranillo activities determines FT expression to trigger flowering. Curr. Biol. CB 2008, 18, 1338–1343. [Google Scholar] [CrossRef] [PubMed]
- Osnato, M.; Castillejo, C.; Matías-Hernández, L.; Pelaz, S. Tempranillo genes link photoperiod and gibberellin pathways to control flowering in Arabidopsis. Nat. Commun. 2012, 3, 808. [Google Scholar] [CrossRef] [PubMed]
- Matías-Hernández, L.; Aguilar-Jaramillo, A.E.; Marín-González, E.; Suárez-López, P.; Pelaz, S. RAV genes: Regulationof floral induction and beyond. Ann. Bot. 2014, 114, 1459–1470. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Kang, H.K.; Son, S.H.; Kim, S.K.; Nam, K.H. A Subset of Arabidopsis RAV Transcription Factors Modulates Drought and Salt Stress Responses Independent of ABA. Plant Cell Physiol. 2014, 55, 1892–1904. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Li, P.F.; Chen, M.K.; Lee, Y.I.; Yang, C.H. Forever Young Flower Negatively Regulates Ethylene Response DNA-Binding Factors by Activating an Ethylene-Responsive Factor to Control Arabidopsis Floral Organ Senescence and Abscission. Plant Physiol. 2015, 168, 1666–1683. [Google Scholar] [CrossRef]
- Woo, H.R.; Kim, J.H.; Kim, J.; Kim, J.; Lee, U.; Song, I.J.; Kim, J.H.; Lee, H.Y.; Nam, H.G.; Lim, P.O. The RAV1 transcription factor positively regulates leaf senescence in Arabidopsis. J. Exp. Bot. 2010, 61, 3947–3957. [Google Scholar] [CrossRef]
- Feng, C.Z.; Chen, Y.; Wang, C.; Kong, Y.H.; Wu, W.H.; Chen, Y.F. Arabidopsis RAV1 transcription factor, phosphorylated by SnRK2 kinases, regulates the expression of ABI3, ABI4, and ABI5 during seed germination and early seedling development. Plant J. Cell Mol. Biol. 2014, 80, 654–668. [Google Scholar] [CrossRef]
- Alvarez, J.P.; Pekker, I.; Goldshmidt, A.; Blum, E.; Amsellem, Z.; Eshed, Y. Endogenous and synthetic microRNAs stimulate simultaneous, efficient, and localized regulation of multiple targets in diverse species. Plant Cell 2006, 18, 1134–1151. [Google Scholar] [CrossRef]
- Alvarez, J.P.; Goldshmidt, A.; Efroni, I.; Bowman, J.L.; Eshed, Y. The NGATHA distal organ development genes are essential for style specification in Arabidopsis. Plant Cell 2009, 21, 1373–1393. [Google Scholar] [CrossRef] [PubMed]
- Scacchi, E.; Osmont, K.S.; Beuchat, J.; Salinas, P.; Navarrete-Gomez, M.; Trigueros, M.; Ferrandiz, C.; Hardtke, C.S. Dynamic, auxin-responsive plasma membrane-to-nucleus movement of Arabidopsis BRX. Development 2009, 136, 2059–2067. [Google Scholar] [CrossRef] [PubMed]
- Trigueros, M.; Navarrete-Gómez, M.; Sato, S.; Christensen, S.K.; Pelaz, S.; Weigel, D.; Yanofsky, M.F.; Ferrándiz, C. The NGATHA genes direct style development in the Arabidopsis gynoecium. Plant Cell 2009, 21, 1394–1409. [Google Scholar] [CrossRef]
- Shao, J.X.; Liu, X.Y.; Wang, R.; Zhang, G.S.; Yu, F. The Over-Expression of an Arabidopsis B3 Transcription Factor, ABS2/NGAL1, Leads to the Loss of Flower Petals. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Zhang, Y.; Du, L.; Xu, R.; Cui, R.; Hao, J.; Sun, C.; Li, Y. Transcription factors SOD7/NGAL2 and DPA4/NGAL3 act redundantly to regulate seed size by directly repressing KLU expression in Arabidopsis thaliana. Plant Cell 2015, 27, 620–632. [Google Scholar] [CrossRef] [PubMed]
- Je, B.I.; Piao, H.L.; Park, S.J.; Park, S.H.; Kim, C.M.; Xuan, Y.H.; Park, S.H.; Huang, J.; Do Choi, Y.; An, G.; et al. RAV-Like1 maintains brassinosteroid homeostasis via the coordinated activation of BRI1 and biosynthetic genes in rice. Plant Cell 2010, 22, 1777–1791. [Google Scholar] [CrossRef] [PubMed]
- Je, B.I.; Han, C.D. Brassinosteroid homeostasis via coordinate regulation of signaling and synthetic pathways. Plant Signal. Behav. 2010, 5, 2. [Google Scholar] [CrossRef]
- Xuan, Y.H.; Duan, F.Y.; Je, B.I.; Kim, C.M.; Li, T.Y.; Liu, J.M.; Park, S.J.; Cho, J.H.; Kim, T.H.; von Wiren, N.; et al. Related to ABI3/VP1-Like 1 (RAVL1) regulates brassinosteroid-mediated activation of AMT1;2 in rice (Oryza sativa). J. Exp. Bot. 2017, 68, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Cortés, A.; Hernández-Verdeja, T.; Sánchez-Jiménez, P.; González-Melendi, P.; Aragoncillo, C.; Allona, I. CsRAV1 induces sylleptic branching in hybrid poplar. New Phytol. 2012, 194, 83–90. [Google Scholar] [CrossRef]
- Zhao, L.; Hao, D.; Chen, L.; Lu, Q.; Zhang, Y.; Li, Y.; Duan, Y.; Li, W. Roles for a soybean RAV-like orthologue in shoot regeneration and photoperiodicity inferred from transgenic plants. J. Exp. Bot. 2012, 63, 3257–3270. [Google Scholar] [CrossRef]
- Kirch, T. The dornroschen/enhancer of shoot regeneration1 Gene of Arabidopsis Acts in the Control of Meristem Cell Fate and Lateral Organ Development. Plant Cell Online 2003, 15, 694–705. [Google Scholar] [CrossRef]
- Kwon, S.H.; Lee, B.H.; Kim, E.Y.; Seo, Y.S.; Lee, S.; Kim, W.T.; Song, J.T.; Kim, J.H. Overexpression of a Brassica rapa NGATHA gene in Arabidopsis thaliana negatively affects cell proliferation during lateral organ and root growth. Plant Cell Physiol. 2009, 50, 2162–2173. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Luo, Q.; Yang, C.; Han, Y.; Li, W. A RAV-like transcription factor controls photosynthesis and senescence in soybean. Planta 2008, 227, 1389–1399. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.Y.; Zhao, L.; Li, D.M.; Hao, D.Q.; Zhan, Y.; Li, W.B. A GmRAV Ortholog Is Involved in Photoperiod and Sucrose Control of Flowering Time in Soybean. PLoS ONE 2014, 9, 89145. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, L.; Zhang, Y.; Cao, C.; Liu, F.; Huang, F.; Qiu, Y.; Li, R.; Lou, X. Map-based cloning and characterization of BPH29, a B3 domain-containing recessive gene conferring brown planthopper resistance in rice. J. Exp. Bot. 2015, 66, 6035–6045. [Google Scholar] [CrossRef] [PubMed]
- Li, C.W.; Su, R.C.; Cheng, C.P.; Sanjaya; You, S.J.; Hsieh, T.H.; Chao, T.C.; Chan, M.T. Tomato RAV Transcription Factor Is a Pivotal Modulator Involved in the AP2/EREBP-Mediated Defense Pathway. Plant Physiol. 2011, 156, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.B.; Li, J.; Qin, R.Y.; Xu, R.F.; Li, H.; Yang, Y.C.; Ma, H.; Li, L.; Wei, P.C.; Yang, J.B. Identification of a regulatory element responsible for salt induction of rice OsRAV2 through ex situ and in situ promoter analysis. Plant Mol. Biol. 2015. [Google Scholar] [CrossRef]
- Chandler, J.W. Auxin response factors. Plant Cell Environ. 2016, 39, 1014–1028. [Google Scholar] [CrossRef]
- Liu, Z.-H.; Yu, Y.-C.; Xiang, F.-N. Auxin response factors and plant growth and development. Hereditas 2011, 33, 1335–1346. [Google Scholar] [CrossRef]
- Li, S.B.; Xie, Z.Z.; Hu, C.G.; Zhang, J.Z. A Review of Auxin Response Factors (ARFs) in Plants. Front. Plant Sci. 2016, 7, 47. [Google Scholar] [CrossRef]
- Ulmasov, T.; Hagen, G.; Guilfoyle, T.J. Activation and repression of transcription by auxin-response factors. Proc. Natl. Acad. Sci. USA 1999, 96, 5844–5849. [Google Scholar] [CrossRef] [PubMed]
- Ulmasov, T.; Hagen, G.; Guilfoyle, T.J. ARF1, a Transcription Factor That Binds to Auxin Response Elements. Science 1997, 276, 1865–1868. [Google Scholar] [CrossRef]
- Roosjen, M.; Paque, S.; Weijers, D. Auxin Response Factors: Output control in auxin biology. J. Exp. Bot. 2018, 69, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wu, X.; Li, T.; Jia, M.; Liu, X.; Zou, Y.; Liu, Z.; Wen, F. Identification, characterization, and expression analysis of auxin response factor (ARF) gene family in Brachypodium distachyon. Funct. Integr. Genomics 2018. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; He, P.; Zhao, P.; Liu, H.; Zhang, L.; Pang, C.; Yu, J. Genome-wide identification of the GhARF gene family reveals that GhARF2 and GhARF18 are involved in cotton fibre cell initiation. J. Exp. Bot. 2018, 69, 4323–4337. [Google Scholar] [CrossRef]
- Tang, Y.; Bao, X.; Liu, K.; Wang, J.; Zhang, J.; Feng, Y.; Wang, Y.; Lin, L.; Feng, J.; Li, C. Genome-wide identification and expression profiling of the auxin response factor (ARF) gene family in physic nut. PLoS ONE 2018, 13, e0201024. [Google Scholar] [CrossRef]
- Die, J.V.; Gil, J.; Millan, T. Genome-wide identification of the auxin response factor gene family in Cicer arietinum. BMC Genom. 2018, 19, 301. [Google Scholar] [CrossRef]
- Singh, V.K.; Rajkumar, M.S.; Garg, R.; Jain, M. Genome-wide identification and co-expression network analysis provide insights into the roles of auxin response factor gene family in chickpea. Sci. Rep. 2017, 7, 10895. [Google Scholar] [CrossRef]
- Wang, W.; Gu, L.; Ye, S.; Zhang, H.; Cai, C.; Xiang, M.; Gao, Y.; Wang, Q.; Lin, C.; Zhu, Q. Genome-wide analysis and transcriptomic profiling of the auxin biosynthesis, transport and signaling family genes in moso bamboo (Phyllostachys heterocycla). BMC Genomics 2017, 18, 870. [Google Scholar] [CrossRef]
- Guilfoyle, T.J.; Hagen, G. Auxin response factors. Curr. Opin. Plant Biol. 2007, 10, 453–460. [Google Scholar] [CrossRef]
- Rademacher, E.H.; Lokerse, A.S.; Schlereth, A.; Llavata-Peris, C.I.; Bayer, M.; Kientz, M.; Freire Rios, A.; Borst, J.W.; Lukowitz, W.; Jurgens, G.; et al. Different auxin response machineries control distinct cell fates in the early plant embryo. Dev. Cell 2012, 22, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Hardtke, C.S.; Berleth, T. The Arabidopsis gene MONOPTEROS encodes a transcription factor mediating embryo axis formation and vascular development. EMBO J. 1998, 17, 1405–1411. [Google Scholar] [CrossRef] [PubMed]
- Hardtke, C.S.; Ckurshumova, W.; Vidaurre, D.P.; Singh, S.A.; Stamatiou, G.; Tiwari, S.B.; Hagen, G.; Guilfoyle, T.J.; Berleth, T. Overlapping and non-redundant functions of the Arabidopsis auxin response factors Monopteros and Nonphototropic Hypocotyl 4. Development 2004, 131, 1089–1100. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.R.; Arreola, A.; Gallagher, T.L.; Gasser, C.S. ETTIN (ARF3) physically interacts with KANADI proteins to form a functional complex essential for integument development and polarity determination in Arabidopsis. Development 2012, 139, 1105–1109. [Google Scholar] [CrossRef]
- Pekker, I.; Alvarez, J.P.; Eshed, Y. Auxin response factors mediate Arabidopsis organ asymmetry via modulation of KANADI activity. Plant Cell 2005, 17, 2899–2910. [Google Scholar] [CrossRef] [PubMed]
- Lim, P.O.; Lee, I.C.; Kim, J.; Kim, H.J.; Ryu, J.S.; Woo, H.R.; Nam, H.G. Auxin response factor 2 (ARF2) plays a major role in regulating auxin-mediated leaf longevity. J. Exp. Bot. 2010, 61, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- Ellis, C.M.; Nagpal, P.; Young, J.C.; Hagen, G.; Guilfoyle, T.J.; Reed, J.W. Auxin response factor1 and auxin response factor2 regulate senescence and floral organ abscission in Arabidopsis thaliana. Development 2005, 132, 4563–4574. [Google Scholar] [CrossRef]
- Wang, Y.; Li, K.; Chen, L.; Zou, Y.; Liu, H.; Tian, Y.; Li, D.; Wang, R.; Zhao, F.; Ferguson, B.J.; et al. MicroRNA167-Directed Regulation of the Auxin Response Factors GmARF8a and GmARF8b Is Required for Soybean Nodulation and Lateral Root Development. Plant Physiol. 2015, 168, 984–999. [Google Scholar] [CrossRef]
- Wilmoth, J.C.; Wang, S.; Tiwari, S.B.; Joshi, A.D.; Hagen, G.; Guilfoyle, T.J.; Alonso, J.M.; Ecker, J.R.; Reed, J.W. NPH4/ARF7 and ARF19 promote leaf expansion and auxin-induced lateral root formation. Plant J. Cell Mol. Biol. 2005, 43, 118–130. [Google Scholar] [CrossRef]
- Cho, H.; Ryu, H.; Rho, S.; Hill, K.; Smith, S.; Audenaert, D.; Park, J.; Han, S.; Beeckman, T.; Bennett, M.J.; et al. A secreted peptide acts on BIN2-mediated phosphorylation of ARFs to potentiate auxin response during lateral root development. Nat. Cell Biol. 2014, 16, 66–76. [Google Scholar] [CrossRef]
- Liu, X.; Dinh, T.T.; Li, D.; Shi, B.; Li, Y.; Cao, X.; Guo, L.; Pan, Y.; Jiao, Y.; Chen, X. Auxin response factor 3 integrates the functions of AGAMOUS and APETALA2 in floral meristem determinacy. Plant J. Cell Mol. Biol. 2014, 80, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Wu, S.; Van Houten, J.; Wang, Y.; Ding, B.; Fei, Z.; Clarke, T.H.; Reed, J.W.; van der Knaap, E. Down-regulation of Auxin Response Factors 6 and 8 by microRNA 167 leads to floral development defects and female sterility in tomato. J. Exp. Bot. 2014, 65, 2507–2520. [Google Scholar] [CrossRef] [PubMed]
- Goetz, M.; Vivian-Smith, A.; Johnson, S.D.; Koltunow, A.M. Auxin Response Factor8 is a negative regulator of fruit initiation in Arabidopsis. Plant Cell 2006, 18, 1873–1886. [Google Scholar] [CrossRef] [PubMed]
- Sagar, M.; Chervin, C.; Mila, I.; Hao, Y.; Roustan, J.P.; Benichou, M.; Gibon, Y.; Biais, B.; Maury, P.; Latche, A.; et al. SlARF4, an auxin response factor involved in the control of sugar metabolism during tomato fruit development. Plant Physiol. 2013, 161, 1362–1374. [Google Scholar] [CrossRef] [PubMed]
- de Jong, M.; Wolters-Arts, M.; Garcia-Martinez, J.L.; Mariani, C.; Vriezen, W.H. The Solanum lycopersicum Auxin Response Factor 7 (SlARF7) mediates cross-talk between auxin and gibberellin signalling during tomato fruit set and development. J. Exp. Bot. 2011, 62, 617–626. [Google Scholar] [CrossRef] [PubMed]
- de Jong, M.; Wolters-Arts, M.; Feron, R.; Mariani, C.; Vriezen, W.H. The Solanum lycopersicum auxin response factor 7 (SlARF7) regulates auxin signaling during tomato fruit set and development. Plant J. Cell Mol. Biol. 2009, 57, 160–170. [Google Scholar] [CrossRef] [PubMed]
- de Jong, M.; Wolters-Arts, M.; Schimmel, B.C.; Stultiens, C.L.; de Groot, P.F.; Powers, S.J.; Tikunov, Y.M.; Bovy, A.G.; Mariani, C.; Vriezen, W.H.; et al. Solanum lycopersicum AUXIN RESPONSE FACTOR 9 regulates cell division activity during early tomato fruit development. J. Exp. Bot. 2015, 66, 3405–3416. [Google Scholar] [CrossRef]
- Ha, C.V.; Le, D.T.; Nishiyama, R.; Watanabe, Y.; Sulieman, S.; Tran, U.T.; Mochida, K.; Dong, N.V.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; et al. The auxin response factor transcription factor family in soybean: Genome-wide identification and expression analyses during development and water stress. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2013, 20, 511–524. [Google Scholar] [CrossRef]
- Hu, W.; Zuo, J.; Hou, X.; Yan, Y.; Wei, Y.; Liu, J.; Li, M.; Xu, B.; Jin, Z. The auxin response factor gene family in banana: Genome-wide identification and expression analyses during development, ripening, and abiotic stress. Front. Plant Sci. 2015, 6, 742. [Google Scholar] [CrossRef]
- Jain, M.; Khurana, J.P. Transcript profiling reveals diverse roles of auxin-responsive genes during reproductive development and abiotic stress in rice. FEBS J. 2009, 276, 3148–3162. [Google Scholar] [CrossRef]
- Wang, S.K.; Bai, Y.H.; Shen, C.J.; Wu, Y.R.; Zhang, S.N.; Jiang, D.A.; Guilfoyle, T.J.; Chen, M.; Qi, Y.H. Auxin-related gene families in abiotic stress response in Sorghum bicolor. Funct. Integr. Genomics 2010, 10, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dai, X.; Zhao, Y. A role for auxin response factor 19 in auxin and ethylene signaling in Arabidopsis. Plant Physiol. 2006, 140, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Zouine, M.; Fu, Y.; Chateigner-Boutin, A.L.; Mila, I.; Frasse, P.; Wang, H.; Audran, C.; Roustan, J.P.; Bouzayen, M. Characterization of the tomato ARF gene family uncovers a multi-levels post-transcriptional regulation including alternative splicing. PLoS ONE 2014, 9, e84203. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.; Maharjan, P.M.; Lee, O.; Fujioka, S.; Jang, S.; Kim, B.; Takatsuto, S.; Tsujimoto, M.; Kim, H.; Cho, S.; et al. Auxin stimulates DWARF4 expression and brassinosteroid biosynthesis in Arabidopsis. Plant J. Cell Mol. Biol. 2011, 66, 564–578. [Google Scholar] [CrossRef] [PubMed]
- Walcher, C.L.; Nemhauser, J.L. Bipartite promoter element required for auxin response. Plant Physiol. 2012, 158, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Vert, G.; Walcher, C.L.; Chory, J.; Nemhauser, J.L. Integration of auxin and brassinosteroid pathways by Auxin Response Factor 2. Proc. Natl. Acad. Sci. USA 2008, 105, 9829–9834. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.; Zhu, J.Y.; Bai, M.Y.; Arenhart, R.A.; Sun, Y.; Wang, Z.Y. Cell elongation is regulated through a central circuit of interacting transcription factors in the Arabidopsis hypocotyl. eLife 2014, 3. [Google Scholar] [CrossRef] [PubMed]
- Richter, R.; Behringer, C.; Zourelidou, M.; Schwechheimer, C. Convergence of auxin and gibberellin signaling on the regulation of the GATA transcription factors GNC and GNL in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2013, 110, 13192–13197. [Google Scholar] [CrossRef] [PubMed]
- King, G.J.; Chanson, A.H.; McCallum, E.J.; Ohme-Takagi, M.; Byriel, K.; Hill, J.M.; Martin, J.L.; Mylne, J.S. The Arabidopsis B3 domain protein VERNALIZATION1 (VRN1) is involved in processes essential for development, with structural and mutational studies revealing its DNA-binding surface. J. Biol. Chem. 2013, 288, 3198–3207. [Google Scholar] [CrossRef] [PubMed]
- Mantegazza, O.; Gregis, V.; Mendes, M.A.; Morandini, P.; Alves-Ferreira, M.; Patreze, C.M.; Nardeli, S.M.; Kater, M.M.; Colombo, L. Analysis of the arabidopsis REM gene family predicts functions during flower development. Ann. Bot. 2014, 114, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Franco-Zorrilla, J.M.; Fernandez-Calvin, B.; Madueno, F.; Cruz-Alvarez, M.; Salinas, J.; Martinez-Zapater, J.M. Identification of genes specifically expressed in cauliflower reproductive meristems. Mol. Charact. BoREM1 Plant Mol. Biol. 1999, 39, 427–436. [Google Scholar] [CrossRef]
- Franco-Zorrilla, J.M.; Cubas, P.; Jarillo, J.A.; Fernandez-Calvin, B.; Salinas, J.; Martinez-Zapater, J.M. AtREM1, a member of a new family of B3 domain-containing genes, is preferentially expressed in reproductive meristems. Plant Physiol. 2002, 128, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Levy, Y.Y.; Mesnage, S.; Mylne, J.S.; Gendall, A.R.; Dean, C. Multiple roles of Arabidopsis VRN1 in vernalization and flowering time control. Science 2002, 297, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Sung, S.; Amasino, R.M. Vernalization in Arabidopsis thaliana is mediated by the PHD finger protein VIN3. Nature 2004, 427, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Mylne, J.S.; Barrett, L.; Tessadori, F.; Mesnage, S.; Johnson, L.; Bernatavichute, Y.V.; Jacobsen, S.E.; Fransz, P.; Dean, C. LHP1, the Arabidopsis homologue of Heterochromatin Protein1, is required for epigenetic silencing of FLC. Proc. Natl. Acad. Sci. USA 2006, 103, 5012–5017. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.; Wilson, A.; Dean, C. Arabidopsis mutants showing an altered response to vernalization. Plant J. Cell Mol. Biol. 1996, 10, 637–644. [Google Scholar] [CrossRef]
- Favaro, R.; Pinyopich, A.; Battaglia, R.; Kooiker, M.; Borghi, L.; Ditta, G.; Yanofsky, M.F.; Kater, M.M.; Colombo, L. MADS-box protein complexes control carpel and ovule development in Arabidopsis. Plant Cell 2003, 15, 2603–2611. [Google Scholar] [CrossRef]
- Mendes, M.A.; Guerra, R.F.; Berns, M.C.; Manzo, C.; Masiero, S.; Finzi, L.; Kater, M.M.; Colombo, L. MADS domain transcription factors mediate short-range DNA looping that is essential for target gene expression in Arabidopsis. Plant Cell 2013, 25, 2560–2572. [Google Scholar] [CrossRef]
- Matias-Hernandez, L.; Battaglia, R.; Galbiati, F.; Rubes, M.; Eichenberger, C.; Grossniklaus, U.; Kater, M.M.; Colombo, L. VERDANDI is a direct target of the MADS domain ovule identity complex and affects embryo sac differentiation in Arabidopsis. Plant Cell 2010, 22, 1702–1715. [Google Scholar] [CrossRef]
- Villarino, G.H.; Hu, Q.; Manrique, S.; Flores-Vergara, M.; Sehra, B.; Robles, L.; Brumos, J.; Stepanova, A.N.; Colombo, L.; Sundberg, E.; et al. Transcriptomic Signature of the SHATTERPROOF2 Expression Domain Reveals the Meristematic Nature of Arabidopsis Gynoecial Medial Domain. Plant Physiol. 2016, 171, 42–61. [Google Scholar] [CrossRef]
- Gomez-Mena, C.; de Folter, S.; Costa, M.M.; Angenent, G.C.; Sablowski, R. Transcriptional program controlled by the floral homeotic gene AGAMOUS during early organogenesis. Development 2005, 132, 429–438. [Google Scholar] [CrossRef]
- Andrianov, V.; Borisjuk, N.; Pogrebnyak, N.; Brinker, A.; Dixon, J.; Spitsin, S.; Flynn, J.; Matyszczuk, P.; Andryszak, K.; Laurelli, M.; et al. Tobacco as a production platform for biofuel: Overexpression of Arabidopsis DGAT and LEC2 genes increases accumulation and shifts the composition of lipids in green biomass. Plant Biotechnol. J. 2010, 8, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, J.; Neuberger, T.; Rolletschek, H.; Schiebold, S.; Nguyen, T.H.; Borisjuk, N.; Borner, A.; Melkus, G.; Jakob, P.; Borisjuk, L. A noninvasive platform for imaging and quantifying oil storage in submillimeter tobacco seed. Plant Physiol. 2013, 161, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Vanhercke, T.; El Tahchy, A.; Liu, Q.; Zhou, X.R.; Shrestha, P.; Divi, U.K.; Ral, J.P.; Mansour, M.P.; Nichols, P.D.; James, C.N.; et al. Metabolic engineering of biomass for high energy density: Oilseed-like triacylglycerol yields from plant leaves. Plant Biotechnol. J. 2014, 12, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Sierro, N.; Battey, J.N.; Ouadi, S.; Bakaher, N.; Bovet, L.; Willig, A.; Goepfert, S.; Peitsch, M.C.; Ivanov, N.V. The tobacco genome sequence and its comparison with those of tomato and potato. Nat. Commun. 2014, 5, 3833. [Google Scholar] [CrossRef]
- Bombarely, A.; Rosli, H.G.; Vrebalov, J.; Moffett, P.; Mueller, L.A.; Martin, G.B. A draft genome sequence of Nicotiana benthamiana to enhance molecular plant-microbe biology research. Mol. Plant-Microbe Interact. MPMI 2012, 25, 1523–1530. [Google Scholar] [CrossRef] [PubMed]
- Sierro, N.; Battey, J.N.; Ouadi, S.; Bovet, L.; Goepfert, S.; Bakaher, N.; Peitsch, M.C.; Ivanov, N.V. Reference genomes and transcriptomes of Nicotiana sylvestris and Nicotiana tomentosiformis. Genome Biol. 2013, 14, R60. [Google Scholar] [CrossRef]
- Edwards, K.D.; Fernandez-Pozo, N.; Drake-Stowe, K.; Humphry, M.; Evans, A.D.; Bombarely, A.; Allen, F.; Hurst, R.; White, B.; Kernodle, S.P.; et al. A reference genome for Nicotiana tabacum enables map-based cloning of homeologous loci implicated in nitrogen utilization efficiency. BMC Genomics 2017, 18. [Google Scholar] [CrossRef]
- Wang, D.; Wang, S.; Chao, J.; Wu, X.; Sun, Y.; Li, F.; Lv, J.; Gao, X.; Liu, G.; Wang, Y. Morphological phenotyping and genetic analyses of a new chemical-mutagenized population of tobacco (Nicotiana tabacum L.). Planta 2017, 246, 149–163. [Google Scholar] [CrossRef]
- Liu, F.; Gong, D.; Zhang, Q.; Wang, D.; Cui, M.; Zhang, Z.; Liu, G.; Wu, J.; Wang, Y. High-throughput generation of an activation-tagged mutant library for functional genomic analyses in tobacco. Planta 2015, 241, 629–640. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. CABIOS 1992, 8, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.H.; Meng, J.L. [MapDraw: A microsoft excel macro for drawing genetic linkage maps based on given genetic linkage data]. Yi chuan = Hereditas/Zhongguoyichuanxue hui bian ji 2003, 25, 317–321. [Google Scholar]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Kao, C.Y.; Cocciolone, S.; McCarty, D.R. Maize VP1 complements Arabidopsis abi3 and confers a novel ABA/auxin interaction in roots. Plant J. Cell Mol. Biol. 2001, 28, 409–418. [Google Scholar] [CrossRef]
- Shen, C.; Wang, S.; Bai, Y.; Wu, Y.; Zhang, S.; Chen, M.; Guilfoyle, T.J.; Wu, P.; Qi, Y. Functional analysis of the structural domain of ARF proteins in rice (Oryza sativa L.). J. Exp. Bot. 2010, 61, 3971–3981. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.H.; Ok, S.H.; Yoo, K.S.; Jung, K.W.; Yoo, S.D.; Shin, J.S. An Arabidopsis cell growth defect factor-related protein, CRS, promotes plant senescence by increasing the production of hydrogen peroxide. Plant Cell Physiol. 2013, 54, 155–167. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Locus Name | Gene Length | Exon Number | Amino Acid Length | Molecular Weight | Theoretical pI | WoLF PAORT |
|---|---|---|---|---|---|---|---|
| NtARF1 | mRNA_8768_cds | 5017 | 11 | 673 | 76,403.29 | 5.5 | Nuclear |
| NtARF2 | mRNA_123344_cds | 8417 | 15 | 827 | 92,057.7 | 6.34 | Nuclear |
| NtARF3 | mRNA_123920_cds | 10585 | 4 | 585 | 64,124.39 | 5.55 | Nuclear |
| NtARF4 | mRNA_136897_cds | 7668 | 14 | 752 | 84,000.46 | 6.32 | Nuclear |
| NtARF5 | mRNA_10429_cds | 2123 | 7 | 560 | 62,854.56 | 5.99 | Nuclear |
| NtARF6 | mRNA_138033_cds | 8596 | 13 | 1090 | 122,010.84 | 6.14 | Nuclear |
| NtARF7 | mRNA_143247_cds | 10421 | 4 | 585 | 64,156.31 | 5.64 | Nuclear |
| NtARF8 | mRNA_10432_cds | 5799 | 11 | 663 | 75,220.24 | 5.61 | Nuclear |
| NtARF9 | mRNA_13156_cds | 5103 | 15 | 671 | 75,146.84 | 5.99 | Nuclear |
| NtARF10 | mRNA_20404_cds | 6406 | 15 | 926 | 102,414.84 | 5.42 | Nuclear |
| NtARF11 | mRNA_21030_cds | 8276 | 13 | 1097 | 121,044.53 | 6.1 | Nuclear |
| NtARF12 | mRNA_828_cds | 9910 | 14 | 839 | 93,884.44 | 5.91 | Nuclear |
| NtARF13 | mRNA_28726_cds | 9993 | 15 | 841 | 94,136.7 | 5.96 | Nuclear |
| NtARF14 | mRNA_38282_cds | 3657 | 5 | 645 | 71,194.66 | 5.97 | Nuclear |
| NtARF15 | mRNA_39086_cds | 5147 | 12 | 1036 | 114,794.48 | 6.12 | Nuclear |
| NtARF16 | mRNA_2133_cds | 4199 | 5 | 581 | 63,946.73 | 8.57 | Nuclear |
| NtARF17 | mRNA_46303_cds | 4080 | 4 | 697 | 76,601.62 | 7 | Nuclear |
| NtARF18 | mRNA_51744_cds | 3949 | 9 | 228 | 25,791.46 | 9.8 | Chloroplast |
| NtARF19 | mRNA_56246_cds | 1662 | 5 | 120 | 13,615.58 | 6.28 | Cytoplasmic |
| NtARF20 | mRNA_56267_cds | 6287 | 9 | 659 | 73,065.38 | 6.82 | Chloroplast |
| NtARF21 | mRNA_58268_cds | 7685 | 13 | 1080 | 120,714.47 | 6.24 | Nuclear |
| NtARF22 | mRNA_61599_cds | 7427 | 15 | 586 | 66,727.98 | 9.17 | Nuclear |
| NtARF23 | mRNA_63531_cds | 10932 | 12 | 813 | 90,516.88 | 6.01 | Nuclear |
| NtARF24 | mRNA_4624_cds | 4923 | 14 | 371 | 41,847.43 | 7.23 | Nuclear |
| NtARF25 | mRNA_64415_cds | 7527 | 14 | 891 | 98,491.78 | 6.01 | Nuclear |
| NtARF26 | mRNA_67201_cds | 4245 | 5 | 669 | 73,902.26 | 6.94 | Nuclear |
| NtARF27 | mRNA_67540_cds | 3030 | 4 | 645 | 71,452.8 | 5.82 | Nuclear |
| NtARF28 | mRNA_72472_cds | 7201 | 15 | 653 | 72,750.24 | 6.19 | Nuclear |
| NtARF29 | mRNA_74528_cds | 4581 | 13 | 777 | 86,319.1 | 8.21 | Nuclear |
| NtARF30 | mRNA_77950_cds | 6035 | 11 | 648 | 73,746.25 | 5.47 | Nuclear |
| NtARF31 | mRNA_84970_cds | 5899 | 11 | 651 | 74,131.64 | 5.43 | Nuclear |
| NtARF32 | mRNA_86031_cds | 10773 | 14 | 843 | 94,349.21 | 6.15 | Nuclear |
| NtARF33 | mRNA_88384_cds | 7628 | 13 | 1106 | 122,429.8 | 5.99 | Nuclear |
| NtARF34 | mRNA_91665_cds | 3726 | 10 | 322 | 35,652.78 | 8.7 | Chloroplast |
| NtARF35 | mRNA_92678_cds | 4707 | 14 | 669 | 74,989.83 | 6.36 | Nuclear |
| NtARF36 | mRNA_93898_cds | 7230 | 13 | 1098 | 121,609.98 | 5.97 | Nuclear |
| NtARF37 | mRNA_106346_cds | 7331 | 15 | 654 | 72,964.4 | 6.06 | Nuclear |
| NtARF38 | mRNA_108249_cds | 5049 | 15 | 678 | 75,608.24 | 5.98 | Nuclear |
| NtARF39 | mRNA_110567_cds | 10827 | 14 | 845 | 94,574.25 | 6.04 | Nuclear |
| NtARF40 | mRNA_114883_cds | 1793 | 2 | 170 | 18,801.42 | 5.46 | Cytoplasmic |
| NtARF41 | mRNA_115491_cds | 4118 | 10 | 555 | 62,458.06 | 5.48 | Nuclear |
| NtARF42 | mRNA_115527_cds | 7098 | 13 | 791 | 87,193.17 | 6.68 | Nuclear |
| NtARF43 | mRNA_117690_cds | 18861 | 4 | 696 | 77,106.71 | 6.99 | Nuclear |
| NtARF44 | mRNA_119154_cds | 4871 | 5 | 696 | 77,152.17 | 7.25 | Nuclear |
| NtARF45 | mRNA_120244_cds | 6469 | 10 | 696 | 77,178.19 | 8.2 | Nuclear |
| NtLAV1 | mRNA_123857_cds | 8524 | 10 | 575 | 62,891.09 | 8.38 | Nuclear |
| NtLAV2 | mRNA_45860_cds | 7079 | 12 | 910 | 100,620.7 | 6.58 | Nuclear |
| NtLAV3 | mRNA_47387_cds | 17347 | 12 | 762 | 83,983.75 | 6.26 | Nuclear |
| NtLAV4 | mRNA_91460_cds | 4643 | 6 | 363 | 40,056.98 | 6.16 | Nuclear |
| NtLAV5 | mRNA_113848_cds | 16337 | 14 | 908 | 99,224.65 | 6.38 | Nuclear |
| NtRAV1 | mRNA_143107_cds | 2955 | 3 | 419 | 47,086.53 | 7.32 | Nuclear |
| NtRAV2 | mRNA_64217_cds | 3327 | 2 | 432 | 48,228.39 | 8.21 | Nuclear |
| NtRAV3 | mRNA_89347_cds | 6164 | 3 | 303 | 34,119.48 | 7.79 | Nuclear |
| NtRAV4 | mRNA_91850_cds | 10441 | 3 | 325 | 36,682.32 | 6.97 | Nuclear |
| NtRAV5 | mRNA_121756_cds | 8408 | 2 | 427 | 47,396.55 | 8.21 | Nuclear |
| NtREM1 | mRNA_174_cds | 4337 | 3 | 204 | 23,183.87 | 4.5 | Nuclear |
| NtREM2 | mRNA_4145_cds | 2340 | 5 | 462 | 52,478.62 | 8.33 | Nuclear |
| NtREM3 | mRNA_5127_cds | 6868 | 9 | 858 | 101,612.96 | 9.37 | Nuclear |
| NtREM4 | mRNA_6438_cds | 2573 | 5 | 246 | 55,365.5 | 6.09 | Nuclear |
| NtREM5 | mRNA_7338_cds | 4885 | 6 | 454 | 55,397.2 | 8.44 | Nuclear |
| NtREM6 | mRNA_7342_cds | 1659 | 2 | 99 | 32,215.88 | 9.74 | Cytoplasmic |
| NtREM7 | mRNA_12898_cds | 5729 | 9 | 835 | 49,752.96 | 9.17 | Nuclear |
| NtREM8 | mRNA_15098_cds | 3650 | 7 | 234 | 28,827.84 | 5.46 | Nuclear |
| NtREM9 | mRNA_17560_cds | 7480 | 5 | 389 | 23,183.87 | 4.5 | Nuclear |
| NtREM10 | mRNA_27980_cds | 3762 | 5 | 254 | 26,819.91 | 9.56 | Cytoplasmic |
| NtREM11 | mRNA_30353_cds | 8238 | 8 | 429 | 93,438.97 | 9.45 | Cytoplasmic |
| NtREM12 | mRNA_35402_cds | 1570 | 4 | 279 | 11,220.96 | 8.26 | Cytoplasmic |
| NtREM13 | mRNA_37366_cds | 5008 | 9 | 782 | 62,632.89 | 6.84 | Nuclear |
| NtREM14 | mRNA_39822_cds | 6017 | 12 | 1081 | 43,996.37 | 9.02 | Nuclear |
| NtREM15 | mRNA_41054_cds | 4658 | 3 | 247 | 69,342.61 | 6.66 | Nuclear |
| NtREM16 | mRNA_41763_cds | 6313 | 11 | 888 | 19,209.82 | 5.76 | Nuclear |
| NtREM17 | mRNA_42571_cds | 5521 | 7 | 526 | 31,624.25 | 9.76 | Nuclear |
| NtREM18 | mRNA_46958_cds | 11,311 | 7 | 488 | 45,134.83 | 9.47 | Nuclear |
| NtREM19 | mRNA_48978_cds | 7520 | 8 | 425 | 41,583.82 | 8.89 | Cytoplasmic |
| NtREM20 | mRNA_49562_cds | 4826 | 7 | 356 | 59,256.58 | 7.59 | Cytoplasmic |
| NtREM21 | mRNA_54810_cds | 10,122 | 9 | 269 | 33,651.85 | 9.28 | Nuclear |
| NtREM22 | mRNA_54825_cds | 9646 | 8 | 483 | 50,571.69 | 8.89 | Cytoplasmic |
| NtREM23 | mRNA_54831_cds | 5913 | 5 | 253 | 32,265.84 | 9.63 | Cytoplasmic |
| NtREM24 | mRNA_54839_cds | 11,118 | 8 | 445 | 59,248.32 | 8.1 | Nuclear |
| NtREM25 | mRNA_54844_cds | 3229 | 4 | 250 | 26,849.94 | 9.54 | Mitochondrial |
| NtREM26 | mRNA_56616_cds | 2658 | 3 | 169 | 43,796.4 | 5.24 | Nuclear |
| NtREM27 | mRNA_57793_cds | 3180 | 2 | 347 | 72,565.57 | 8.76 | Nuclear |
| NtREM28 | mRNA_57802_cds | 1893 | 3 | 320 | 17,770.98 | 4.93 | Nuclear |
| NtREM29 | mRNA_68307_cds | 9559 | 10 | 853 | 99,049.56 | 8.89 | Nuclear |
| NtREM30 | mRNA_68657_cds | 3991 | 7 | 225 | 41,069.05 | 8.82 | Nuclear |
| NtREM31 | mRNA_68659_cds | 4851 | 8 | 335 | 59,213.86 | 6.36 | Nuclear |
| NtREM32 | mRNA_68660_cds | 2871 | 8 | 491 | 28,195.38 | 4.96 | Chloroplast |
| NtREM33 | mRNA_68662_cds | 3438 | 7 | 303 | 94,586.78 | 9.44 | Nuclear |
| NtREM34 | mRNA_70050_cds | 4777 | 8 | 339 | 28,277.8 | 9.48 | Nuclear |
| NtREM35 | mRNA_70052_cds | 3095 | 7 | 225 | 33,265.54 | 9.38 | Nuclear |
| NtREM36 | mRNA_71850_cds | 5227 | 8 | 293 | 33,240.55 | 9.5 | Nuclear |
| NtREM37 | mRNA_75299_cds | 5098 | 8 | 293 | 26,167.04 | 9.13 | Nuclear |
| NtREM38 | mRNA_80332_cds | 2619 | 4 | 250 | 38,446.93 | 9.44 | Cytoplasmic |
| NtREM39 | mRNA_82188_cds | 4574 | 9 | 838 | 34,953.11 | 9.33 | Nuclear |
| NtREM40 | mRNA_84203_cds | 5328 | 10 | 515 | 38,349.91 | 9.56 | Cytoplasmic |
| NtREM41 | mRNA_84209_cds | 7730 | 7 | 357 | 26,187.97 | 8.99 | Cytoplasmic |
| NtREM42 | mRNA_87132_cds | 4813 | 10 | 898 | 97,709.52 | 8.82 | Nuclear |
| NtREM43 | mRNA_88675_cds | 5421 | 11 | 868 | 95,633.22 | 9.04 | Nuclear |
| NtREM44 | mRNA_88789_cds | 4139 | 3 | 156 | 37,223.68 | 4.99 | Nuclear |
| NtREM45 | mRNA_91785_cds | 4529 | 6 | 639 | 39,751.05 | 6.87 | Nuclear |
| NtREM46 | mRNA_91786_cds | 3902 | 4 | 375 | 19,071.44 | 4.6 | Nuclear |
| NtREM47 | mRNA_97289_cds | 3476 | 7 | 234 | 29,559.15 | 9.52 | Nuclear |
| NtREM48 | mRNA_98542_cds | 9751 | 7 | 524 | 50,779.57 | 8.2 | Nuclear |
| NtREM49 | mRNA_99366_cds | 1958 | 4 | 282 | 29,188.64 | 9.2 | Cytoplasmic |
| NtREM50 | mRNA_107285_cds | 2517 | 4 | 296 | 31,029.21 | 9.14 | Chloroplast |
| NtREM51 | mRNA_119576_cds | 5507 | 7 | 528 | 41,119.08 | 8.88 | Nuclear |
| NtREM52 | mRNA_124977_cds | 2000 | 5 | 338 | 52,478.62 | 8.33 | Nuclear |
| NtREM53 | mRNA_136475_cds | 5957 | 7 | 358 | 49,339.11 | 8.34 | Cytoplasmic |
| NtREM54 | mRNA_136480_cds | 10,066 | 10 | 390 | 54,683.67 | 7.31 | Cytoplasmic |
| NtREM55 | mRNA_136483_cds | 4361 | 6 | 274 | 59,060.4 | 8.38 | Cytoplasmic |
| NtREM56 | mRNA_136485_cds | 3637 | 2 | 165 | 100,682.42 | 9.17 | Cytoplasmic |
| NtREM57 | mRNA_136491_cds | 5465 | 10 | 602 | 27,678.15 | 4.61 | Cytoplasmic |
| NtREM58 | mRNA_140712_cds | 5265 | 7 | 394 | 122,461.5 | 9.28 | Nuclear |
| NtREM59 | mRNA_140992_cds | 13,507 | 8 | 552 | 88,099.21 | 9.45 | Nuclear |
| Species | ARF | LAV | RAV | REM | Total |
|---|---|---|---|---|---|
| Arabidopsis (A. thaliana) | 23 | 6 | 13 | 76 | 118 |
| Rice (Oryza sativa) | 28 | 7 | 16 | 40 | 91 |
| Tomato (Solanum lycopersicum) | 22 | 4 | 9 | 62 | 97 |
| Tobacco (N. tabacum) | 45 | 5 | 5 | 59 | 114 |
| Tobacco (Nicotiana sylvestris) | 23 | 4 | 4 | 44 | 75 |
| Tobacco (Nicotiana tomentosiformis) | 23 | 4 | 4 | 15 | 46 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, F.; Sun, T.; Yang, S.; Wang, X.; Chao, J.; Li, X.; Hu, J.; Cui, M.; Liu, G.; Wang, D.; et al. Insight into the B3Transcription Factor Superfamily and Expression Profiling of B3 Genes in Axillary Buds after Topping in Tobacco (Nicotiana tabacum L.). Genes 2019, 10, 164. https://doi.org/10.3390/genes10020164
Xia F, Sun T, Yang S, Wang X, Chao J, Li X, Hu J, Cui M, Liu G, Wang D, et al. Insight into the B3Transcription Factor Superfamily and Expression Profiling of B3 Genes in Axillary Buds after Topping in Tobacco (Nicotiana tabacum L.). Genes. 2019; 10(2):164. https://doi.org/10.3390/genes10020164
Chicago/Turabian StyleXia, Fei, Tingting Sun, Shuangjuan Yang, Xiao Wang, Jiangtao Chao, Xiaoxu Li, Junhua Hu, Mengmeng Cui, Guanshan Liu, Dawei Wang, and et al. 2019. "Insight into the B3Transcription Factor Superfamily and Expression Profiling of B3 Genes in Axillary Buds after Topping in Tobacco (Nicotiana tabacum L.)" Genes 10, no. 2: 164. https://doi.org/10.3390/genes10020164
APA StyleXia, F., Sun, T., Yang, S., Wang, X., Chao, J., Li, X., Hu, J., Cui, M., Liu, G., Wang, D., & Sun, Y. (2019). Insight into the B3Transcription Factor Superfamily and Expression Profiling of B3 Genes in Axillary Buds after Topping in Tobacco (Nicotiana tabacum L.). Genes, 10(2), 164. https://doi.org/10.3390/genes10020164