Characterization and Functional Divergence of a Novel DUF668 Gene Family in Rice Based on Comprehensive Expression Patterns

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of the DUF668 Genes
2.2. Phylogenetic Analysis, Gene Structure, and Conserved Motifs
2.3. Chromosomal Locations, Gene Duplication Events, and Cis-acting Elements Analysis
2.4. Plant Materials and Treatments
2.5. RNA Isolation and RT-PCR Analysis
3. Results
3.1. Genome-Wide Identification and Characterization of DUF668 Genes in Rice
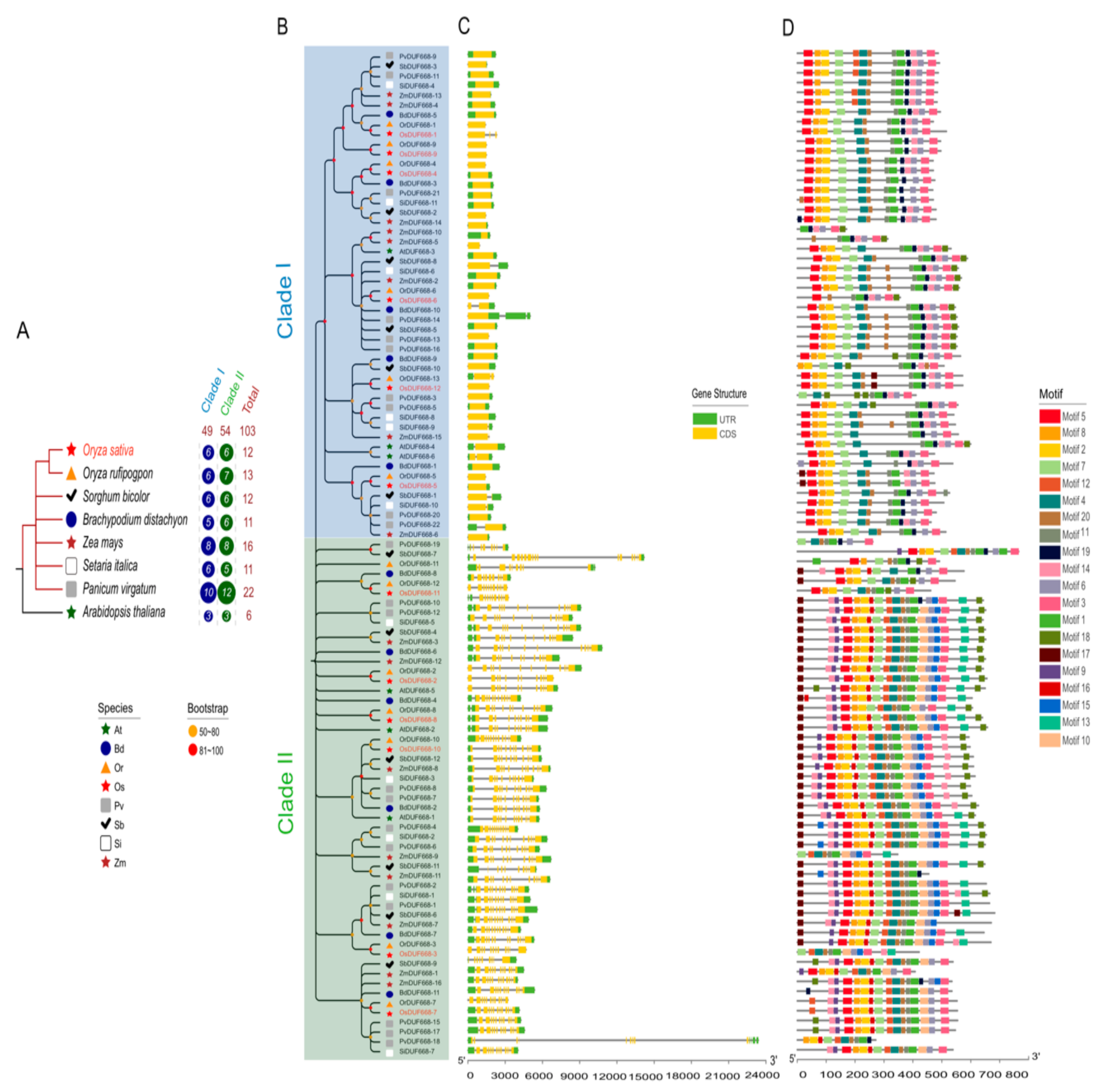
3.2. Phylogenetic Analysis, Gene Structure, and Motif Composition of the DUF668s in Eight Plants
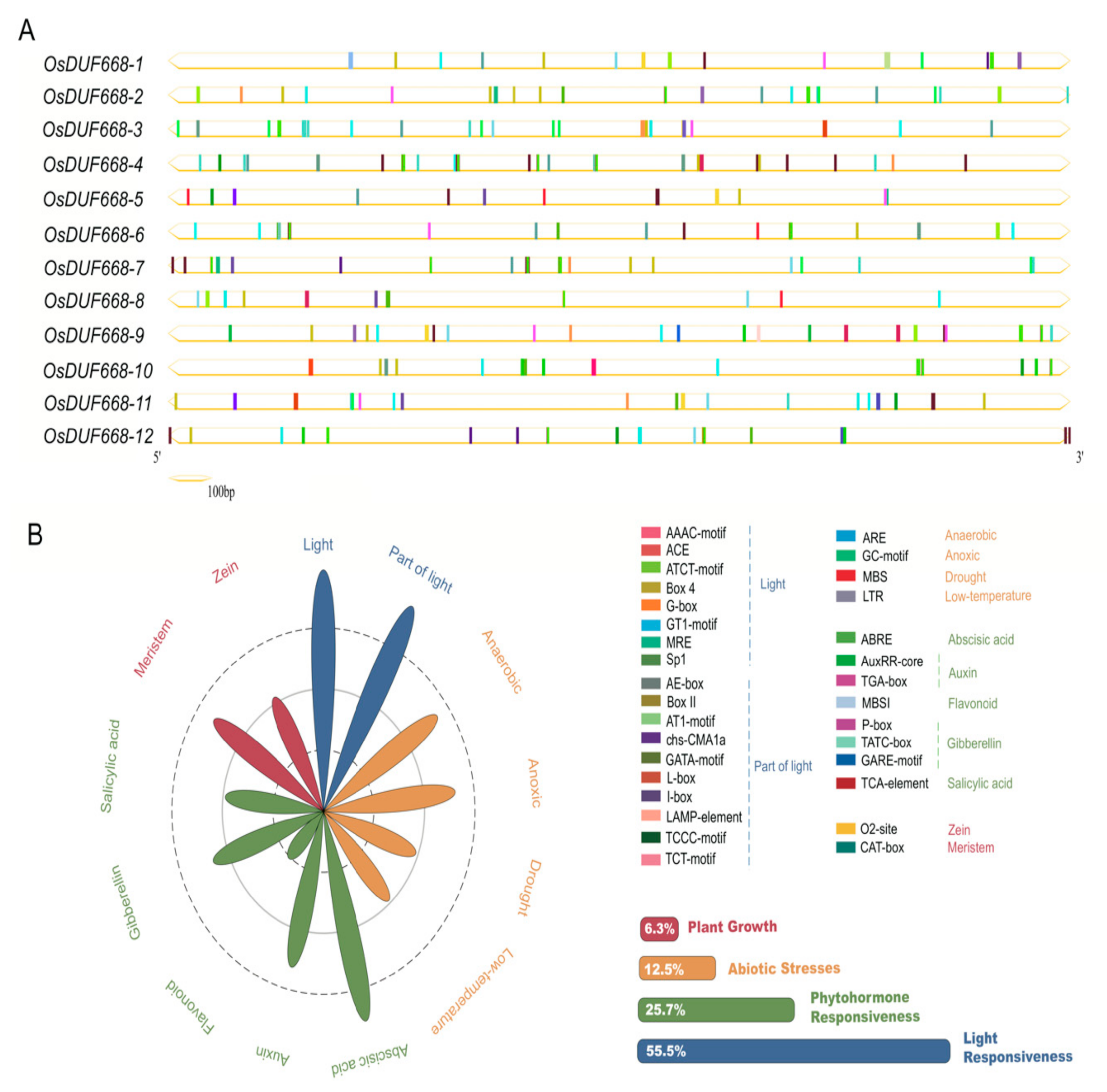
3.3. Cis-acting Elements Prediction of DUF668 Genes in Rice
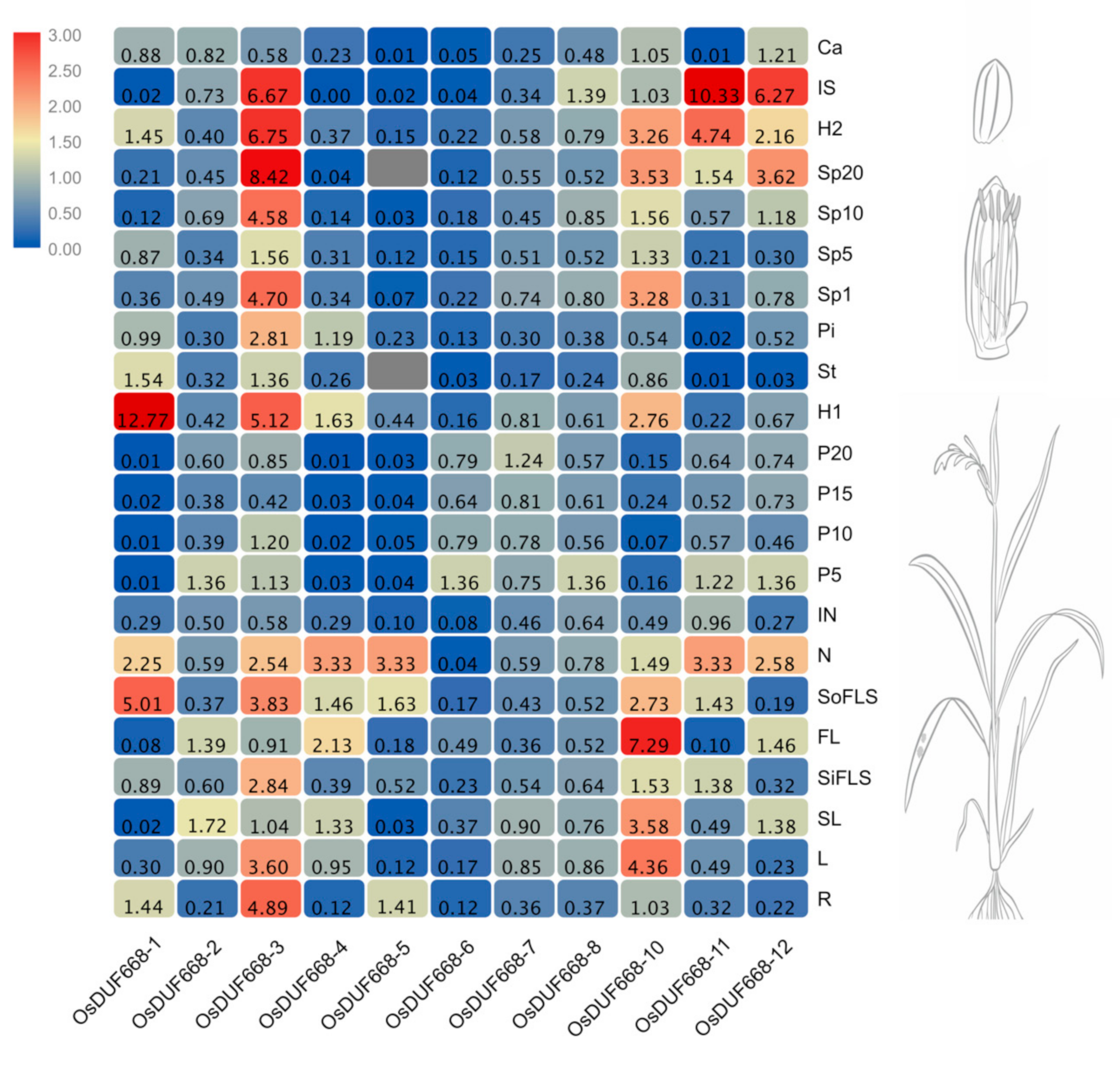
3.4. Tissue-Specific Expression Patterns of OsDUF668s
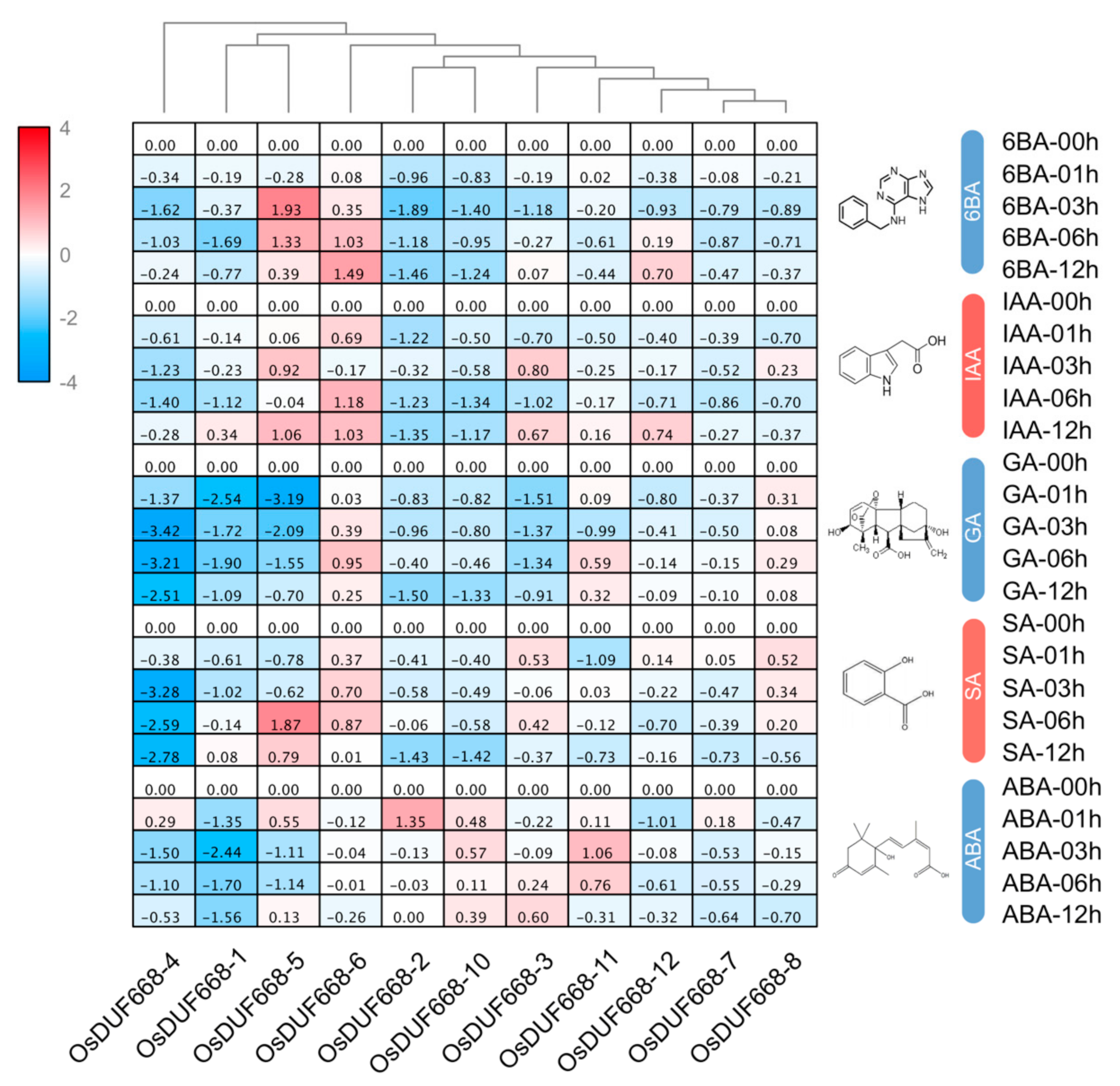
3.5. Expression Profiles of OsDUF668s in Response to Plant Hormone
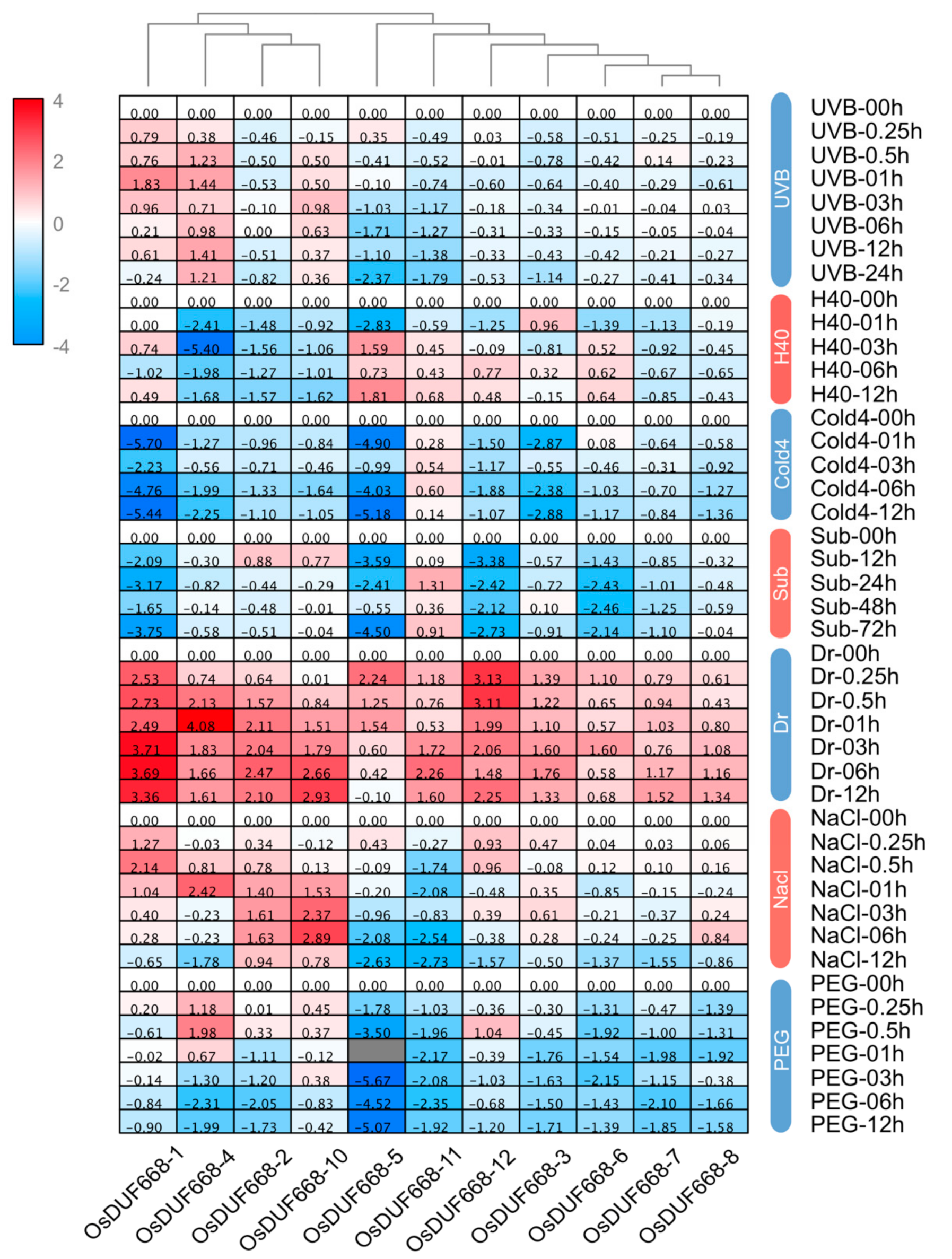
3.6. Transcriptional Responses of OsDUF668s Facing Environmental Stresses
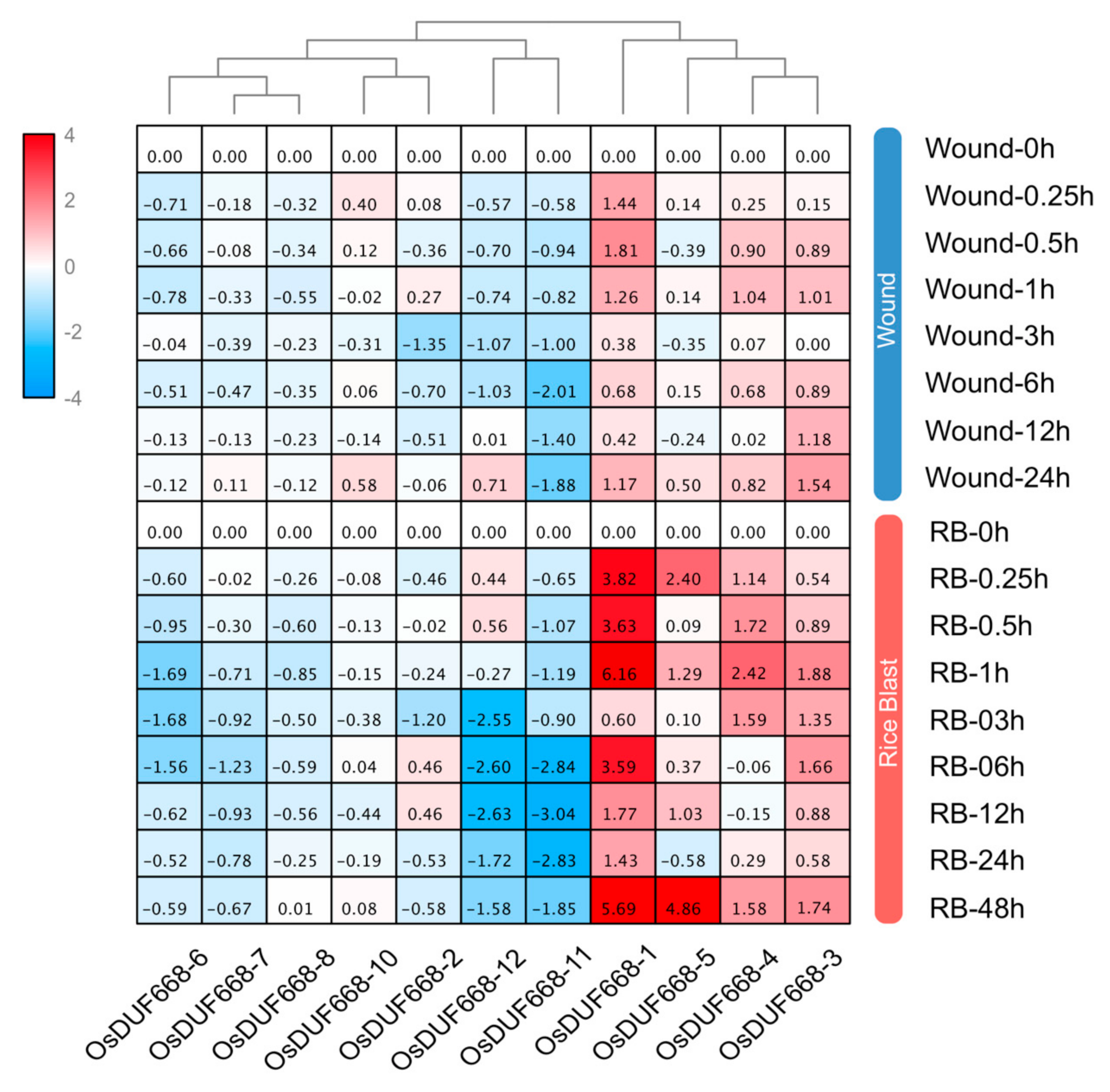
3.7. Transcriptional Responses of OsDUF668s Facing Pathogen-Defense Related Stresses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ohta, T. Evolution of gene families. Gene 2000, 259, 45–52. [Google Scholar] [CrossRef]
- Bateman, A.; Coggill, P.; Finn, R.D. DUFs: Families in search of function. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 1148–1152. [Google Scholar] [CrossRef]
- Simm, R.; Morr, M.; Kader, A.; Nimtz, M.; Römling, U. GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessibility to motility. Mol. Microbiol. 2004, 53, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Karras, G.I.; Kustatscher, G.; Buhecha, H.R.; Allen, M.D.; Pugieux, C.; Sait, F.; Bycroft, M.; Ladurner, A.G. The macro domain is an ADP-ribose binding module. EMBO J. 2005, 24, 1911–1920. [Google Scholar] [CrossRef] [PubMed]
- Dlakić, M. DUF283 domain of dicer proteins has a double-stranded RNA-binding fold. Bioinformatics 2006, 22, 2711–2714. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Chen, F.; Huan, X.; Machida, S.; Song, J.; Yuan, Y.A. Structure of the Arabidopsis thaliana DCL4 DUF283 domain reveals a noncanonical double-stranded RNA-binding fold for protein—Protein interaction. Rna 2010, 16, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Gholizadeh, A. DUF538 protein superfamily is predicted to be chlorophyll hydrolyzing enzymes in plants. Physiol. Mol. Biol. Plants 2016, 22, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Gholizadeh, A. Chlorophyll binding ability of non-chloroplastic DUF538 protein superfamily in plants. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2018, 88, 967–976. [Google Scholar] [CrossRef]
- Palmeros-Suárez, P.A.; Massange-Sánchez, J.A.; Sánchez-Segura, L.; Martínez-Gallardo, N.A.; Espitia Rangel, E.; Gómez-Leyva, J.F.; Délano-Frier, J.P. AhDGR2, an amaranth abiotic stress-induced DUF642 protein gene, modifies cell wall structure and composition and causes salt and ABA hyper-sensibility in transgenic Arabidopsis. Planta 2017, 245, 623–640. [Google Scholar] [CrossRef]
- Harada, K.; Yamashita, E.; Inoue, K.; Yamaguchi, K.; Fujiwara, T.; Nakagawa, A.; Kawasaki, T.; Kojima, C. Plant-specific DUF1110 protein from Oryza sativa: Expression, purification and crystallization. Acta Crystallogr. Sect. Struct. Biol. Commun. 2016, 72, 480–484. [Google Scholar] [CrossRef]
- Bakolitsa, C.; Bateman, A.; Jin, K.K.; McMullan, D.; Krishna, S.S.; Miller, M.D.; Abdubek, P.; Acosta, C.; Astakhova, T.; Axelrod, H.L.; et al. The structure of Jann-2411 (DUF1470) from Jannaschia sp. at 1.45 Å resolution reveals a new fold (the ABATE domain) and suggests its possible role as a transcription regulator. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 1198–1204. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shen, R.; Chen, L.T.; Liu, Y.G. Characterization of a novel DUF1618 gene family in rice. J. Integr. Plant Biol. 2014, 56, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.B.; Bogdanove, A.J.; Sessa, G. Understanding the functions of plant disease resistance proteins. Annu. Rev. Plant Biol. 2003, 54, 23–61. [Google Scholar] [CrossRef] [PubMed]
- Heese, A.; Ludwig, A.A.; Jones, J.D.G. Rapid phosphorylation of a syntaxin during the Avr9/Cf-9-race-specific signaling pathway. Plant Physiol. 2005, 138, 2406–2416. [Google Scholar] [CrossRef] [PubMed]
- Durrant, W.E.; Rowland, O.; Piedras, P.; Hammond-Kosack, K.E.; Jones, J.D.G. cDNA-AFLP reveals a striking overlap in race-specific resistance and wound response gene expression profiles. Plant Cell 2000, 12, 963–977. [Google Scholar] [CrossRef]
- Van Den Burg, H.A.; Tsitsigiannis, D.I.; Rowland, O.; Lo, J.; Rallapalli, G.; MacLean, D.; Takken, F.L.W.; Jones, J.D.G. The F-box protein ACRE189/ACIF1 regulates cell death and defense responses activated during pathogen recognition in tobacco and tomato. Plant Cell 2008, 20, 697–719. [Google Scholar] [CrossRef]
- Rowland, O.; Ludwig, A.A.; Merrick, C.J.; Baillieul, F.; Tracy, F.E.; Durrant, W.E.; Fritz-Laylin, L.; Nekrasov, V.; Sjölander, K.; Yoshioka, H.; et al. Functional analysis of Avr9/Cf-9 rapidly elicited genes identifies a protein kinase, ACIK1, that is essential for full Cf-9-dependent disease resistance in tomato. Plant Cell 2005, 17, 295–310. [Google Scholar] [CrossRef]
- Savatin, D.V.; Gramegna, G.; Modesti, V.; Cervone, F. Wounding in the plant tissue: The defense of a dangerous passage. Front. Plant Sci. 2014, 5, 1–11. [Google Scholar] [CrossRef]
- Walley, J.W.; Coughlan, S.; Hudson, M.E.; Covington, M.F.; Kaspi, R.; Banu, G.; Harmer, S.L.; Dehesh, K. Mechanical stress induces biotic and abiotic stress responses via a novel cis-element. PLoS Genet. 2007, 3, 1800–1812. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Chou, K.C.; Shen, H. Bin Cell-PLoc: A package of web servers for predicting subcellular localization of proteins in various organisms. Nat. Protoc. 2008, 3, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a toolkit for biologists integrating various HTS-data handling tools with a user-friendly interface. bioRxiv 2018. [Google Scholar] [CrossRef]
- He, Z.; Zhang, H.; Gao, S.; Lercher, M.J.; Chen, W.H.; Hu, S. Evolview v2: An online visualization and management tool for customized and annotated phylogenetic trees. Nucleic Acids Res. 2016, 44, W236–W241. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, 1–14. [Google Scholar] [CrossRef]
- Lescot, M. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.; Yang, J.; Hu, K.; An, B.; Deng, X.; Li, Y. Reliable selection and holistic stability evaluation of reference genes for rice under 22 different experimental conditions. Appl. Biochem. Biotechnol. 2016, 179, 753–775. [Google Scholar] [CrossRef]
- Jain, M.; Nijhawan, A.; Tyagi, A.K.; Khurana, J.P. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2006, 345, 646–651. [Google Scholar] [CrossRef]
- Song, Y.; Wang, L.; Xiong, L. Comprehensive expression profiling analysis of OsIAA gene family in developmental processes and in response to phytohormone and stress treatments. Planta 2009, 229, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Hazman, M.; Sühnel, M.; Schäfer, S.; Zumsteg, J.; Lesot, A.; Beltran, F.; Marquis, V.; Herrgott, L.; Miesch, L.; Riemann, M.; et al. Characterization of jasmonoyl-isoleucine (JA-Ile) hormonal catabolic pathways in rice upon wounding and salt stress. Rice 2019, 12, 45. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Wang, L.; Pan, Q. Identification and characterization of a new blast resistance gene located on rice chromosome 1 through linkage and differential analyses. Phytopathology 2004, 94, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Peterson, P.M. Poaceae (Gramineae). Encycl. Life Sci. 2013. [Google Scholar] [CrossRef]
- Soltani, B.M.; Ehlting, J.; Hamberger, B.; Douglas, C.J. Multiple cis-regulatory elements regulate distinct and complex patterns of developmental and wound-induced expression of Arabidopsis thaliana 4CL gene family members. Planta 2006, 224, 1226–1238. [Google Scholar] [CrossRef]
- Walther, D.; Brunnemann, R.; Selbig, J. The regulatory code for transcriptional response diversity and its relation to genome structural properties in A. thaliana. PLoS Genet. 2007, 3, 216–229. [Google Scholar] [CrossRef]
- Abdullah, M.; Cheng, X.; Cao, Y.; Su, X.; Manzoor, M.A.; Gao, J.; Cai, Y.; Lin, Y. Zinc finger-homeodomain transcriptional factors (ZHDs) in upland cotton (Gossypium hirsutum): Genome-wide identification and expression analysis in fiber development. Front. Genet. 2018, 9, 357. [Google Scholar] [CrossRef]
- Marciniak, K.; Przedniczek, K. Comprehensive insight into gibberellin-and jasmonate-mediated stamen development. Genes 2019, 10, 811. [Google Scholar] [CrossRef]
- Deng, X.; An, B.; Zhong, H.; Yang, J.; Kong, W.; Li, Y. A novel insight into functional divergence of the MST gene family in rice based on comprehensive expression patterns. Genes 2019, 10, 239. [Google Scholar] [CrossRef]
- Zhong, H.; Kong, W.; Gong, Z.; Fang, X.; Deng, X.; Liu, C.; Li, Y. Evolutionary analyses reveal diverged patterns of SQUAMOSA promoter binding protein-like (Spl) gene family in Oryza genus. Front. Plant Sci. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Guo, M.; Chen, K.; Zhang, P. Transcriptome profile analysis of resistance induced by burdock fructooligosaccharide in tobacco. J. Plant Physiol. 2012, 169, 1511–1519. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Clade | Chr | mRNA Length (bp) | Protein (aa) | MW (Da) | Theoretical pI | Predicted Location | Annotation |
|---|---|---|---|---|---|---|---|---|---|
| LOC_Os01g62670 | OsDUF668-1 | clade-I | Chr1 | 2317 | 516 | 55952.4 | 8.31 | Chloroplast | avr9/Cf-9 rapidly elicited protein |
| LOC_Os01g65330 | OsDUF668-2 | clade-II | Chr1 | 7233 | 656 | 73065.2 | 9.86 | Nucleus | expressed protein |
| LOC_Os02g34650 | OsDUF668-3 | clade-II | Chr2 | 3899 | 422 | 46119.6 | 10.36 | Chloroplast | expressed protein |
| LOC_Os03g16390 | OsDUF668-4 | clade-I | Chr3 | 1936 | 471 | 52064.3 | 10.07 | Peroxisome | avr9/Cf-9 rapidly elicited protein |
| LOC_Os03g64130 | OsDUF668-5 | clade-I | Chr3 | 1757 | 476 | 52051.3 | 10.31 | Chloroplast | avr9/Cf-9 rapidly elicited protein |
| LOC_Os04g08764 | OsDUF668-6 | clade-I | Chr4 | 2151 | 357 | 38783 | 7.39 | Chloroplast | avr9/Cf-9 rapidly elicited protein |
| LOC_Os04g35410 | OsDUF668-7 | clade-II | Chr4 | 4154 | 554 | 61523.2 | 6.37 | Chloroplast | expressed protein |
| LOC_Os05g35530 | OsDUF668-8 | clade-II | Chr5 | 6418 | 641 | 71108.8 | 9.98 | Cytoplasm | expressed protein |
| LOC_Os05g38320 | OsDUF668-9 | clade-I | Chr5 | 1490 | 497 | 53685 | 10.21 | Chloroplast | avr9/Cf-9 rapidly elicited protein |
| LOC_Os06g50220 | OsDUF668-10 | clade-II | Chr6 | 5930 | 598 | 67106.3 | 9.14 | Nucleus | expressed protein |
| LOC_Os11g07840 | OsDUF668-11 | clade-II | Chr11 | 3264 | 462 | 51730.7 | 7.38 | Mitochondrion | expressed protein |
| LOC_Os12g05180 | OsDUF668-12 | clade-I | Chr12 | 1970 | 573 | 62417.4 | 10.5 | Chloroplast | avr9/Cf-9 rapidly elicited protein |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, H.; Zhang, H.; Guo, R.; Wang, Q.; Huang, X.; Liao, J.; Li, Y.; Huang, Y.; Wang, Z. Characterization and Functional Divergence of a Novel DUF668 Gene Family in Rice Based on Comprehensive Expression Patterns. Genes 2019, 10, 980. https://doi.org/10.3390/genes10120980
Zhong H, Zhang H, Guo R, Wang Q, Huang X, Liao J, Li Y, Huang Y, Wang Z. Characterization and Functional Divergence of a Novel DUF668 Gene Family in Rice Based on Comprehensive Expression Patterns. Genes. 2019; 10(12):980. https://doi.org/10.3390/genes10120980
Chicago/Turabian StyleZhong, Hua, Hongyu Zhang, Rong Guo, Qiang Wang, Xiaoping Huang, Jianglin Liao, Yangsheng Li, Yingjin Huang, and Zhaohai Wang. 2019. "Characterization and Functional Divergence of a Novel DUF668 Gene Family in Rice Based on Comprehensive Expression Patterns" Genes 10, no. 12: 980. https://doi.org/10.3390/genes10120980
APA StyleZhong, H., Zhang, H., Guo, R., Wang, Q., Huang, X., Liao, J., Li, Y., Huang, Y., & Wang, Z. (2019). Characterization and Functional Divergence of a Novel DUF668 Gene Family in Rice Based on Comprehensive Expression Patterns. Genes, 10(12), 980. https://doi.org/10.3390/genes10120980