Transcriptome Profiling, Biochemical and Physiological Analyses Provide New Insights towards Drought Tolerance in Nicotiana tabacum L.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
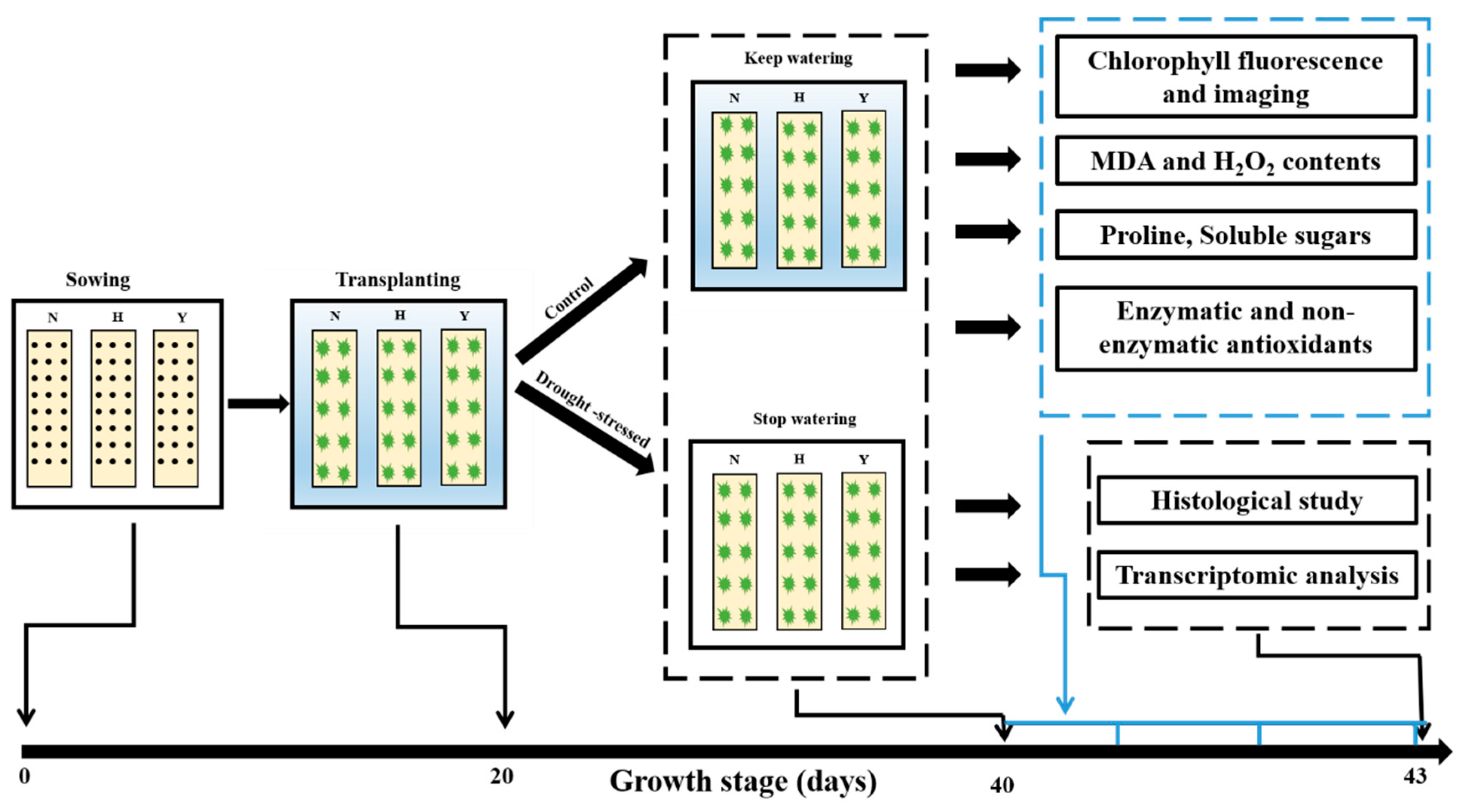
2.1. Plant Materials and Drought Stress Treatment
2.2. Determination of Fresh and Dry Biomass, Chlorophyll Content
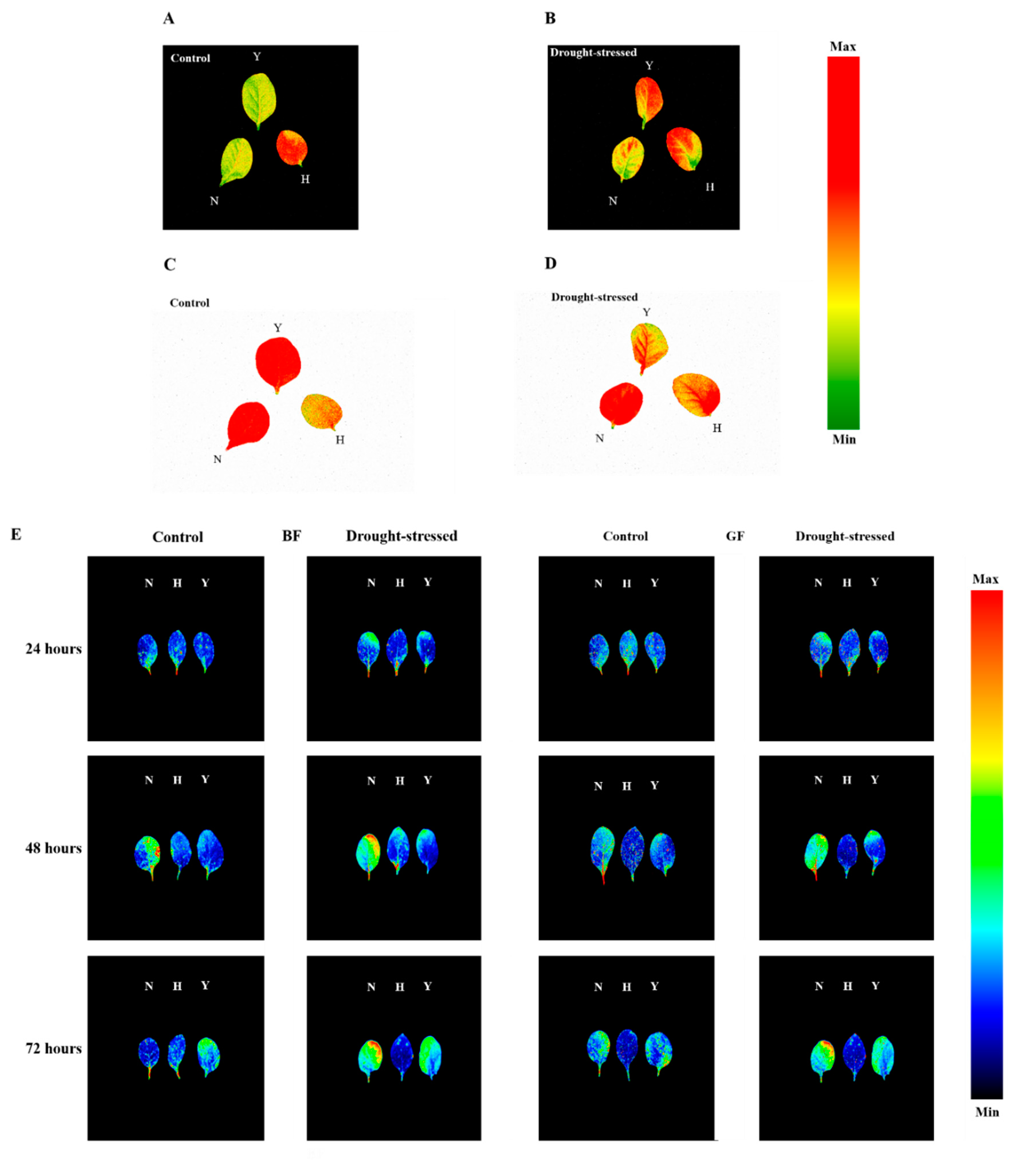
2.3. Chlorophyll Fluorescence and Multicolor Fluorescence Imaging
2.4. Determination of Leaf Water Potential
2.5. Measurements of Superoxide dismutase (SOD), Peroxidase (POD), and Catalase (CAT)
2.6. Quantification of Ascorbc acid (AsA), and Glutathione (GSH) Contents
2.7. Determination of Proline and Soluble Sugars
2.8. Determination of Malondialdehyde (MDA) and Hydrogen Peroxide (H2O2)
2.9. Histological Studies
2.10. RNA Extraction, cDNA Library Construction, and Transcriptome Sequencing
2.11. RNA Sequence Data Analysis
2.12. Validation of RNA-Seq Data through Quantitative Real-Time PCR (qRT-PCR) Analysis
2.13. Statistical Analysis
3. Results
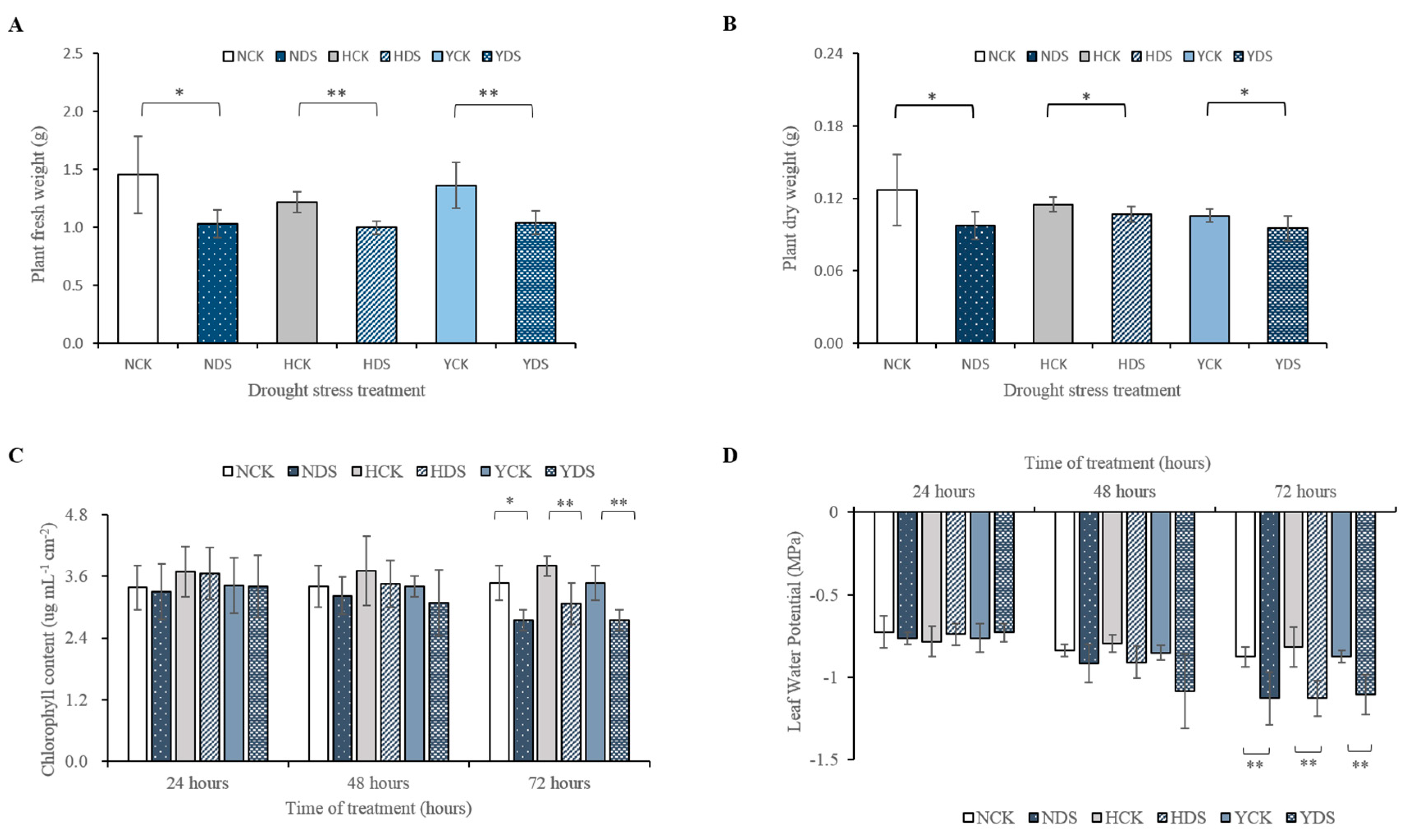
3.1. Effect of Drought Stress on Plant Growth, Chlorophyll Content and Leaf Water Potential of Three Tobacco Varieties
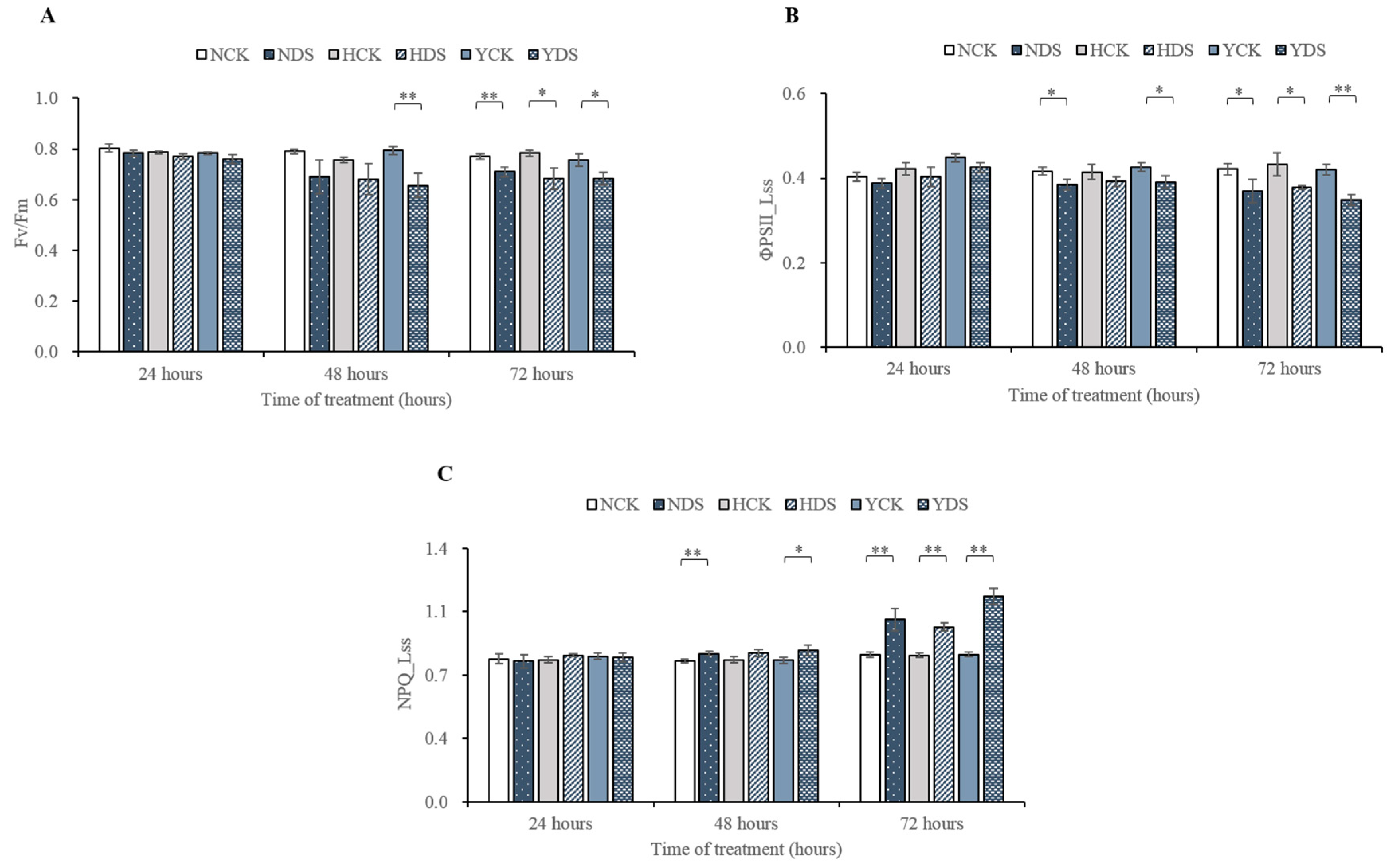
3.2. Chlorophyll Fluorescence Parameters in Response to Drought Stress
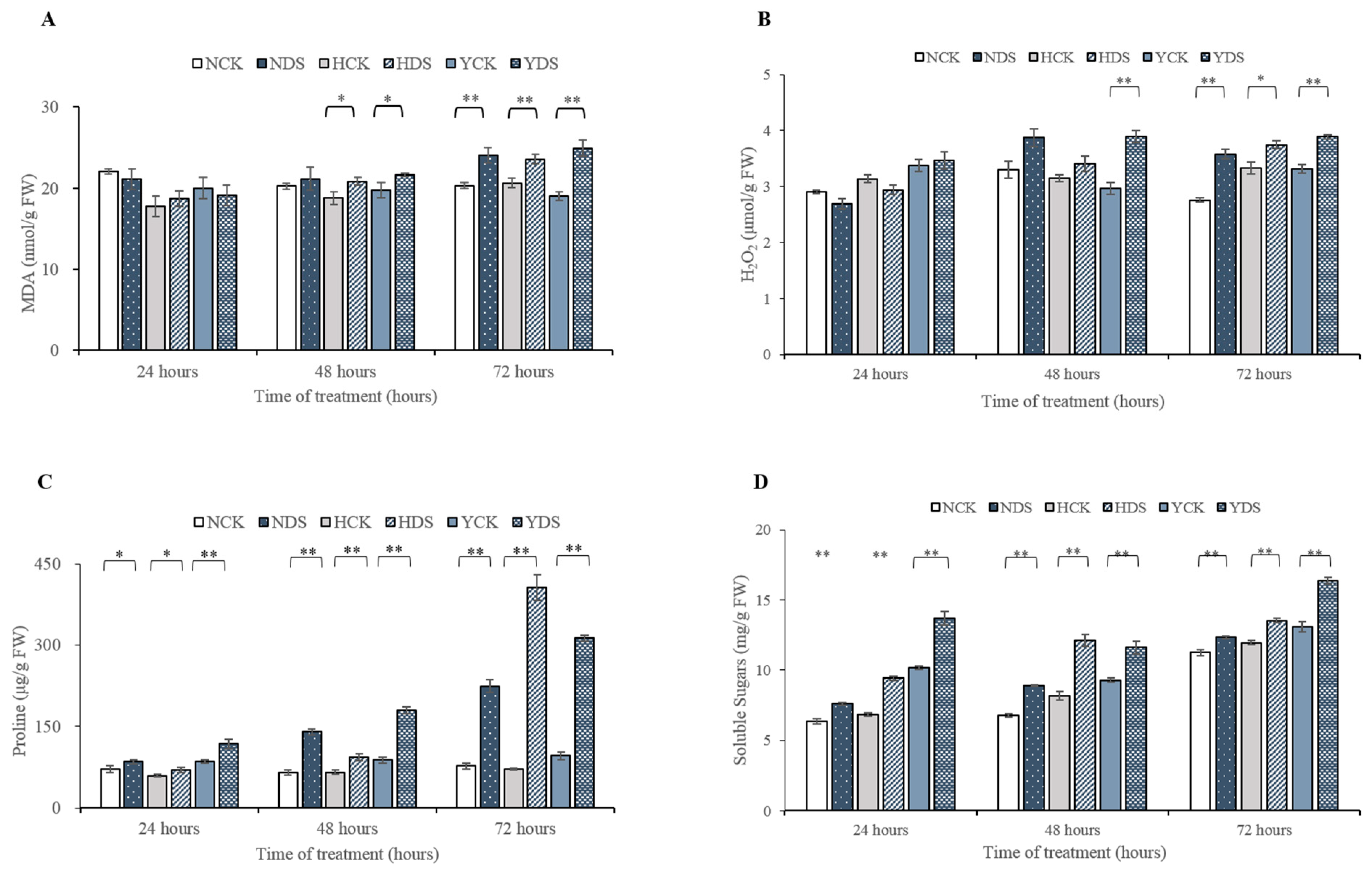
3.3. Oxidative Damage (MDA and H2O2 Contents) in Response to Drought Stress
3.4. Antioxidant Enzymes Activity in Response to Drought Stress
3.5. Non-Enzymatic Antioxidant Components in Response to Drought Stress
3.6. Proline and Soluble Sugar Contents in Response to Drought Stress
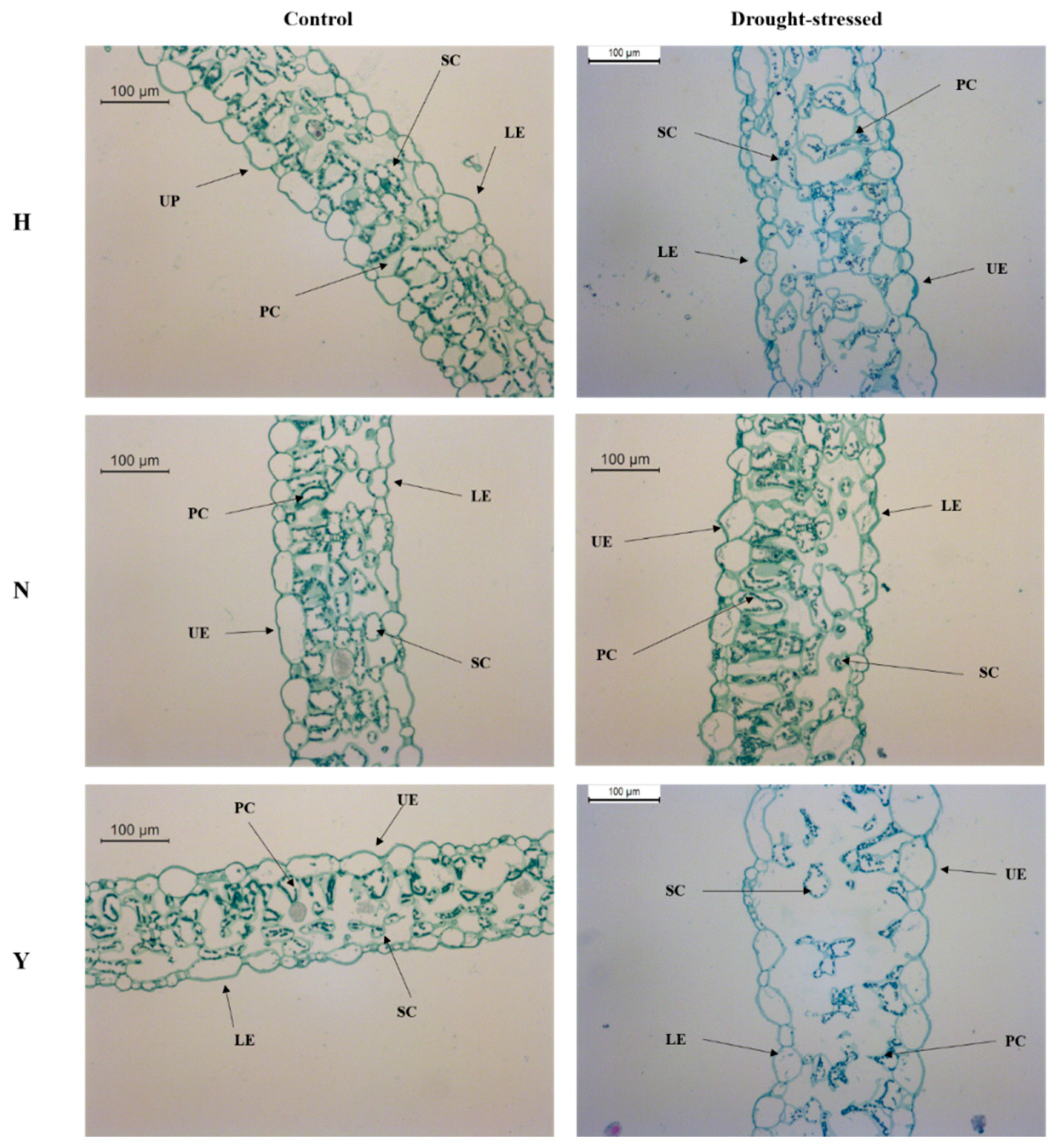
3.7. Leaf Anatomy in Response to Drought Stress
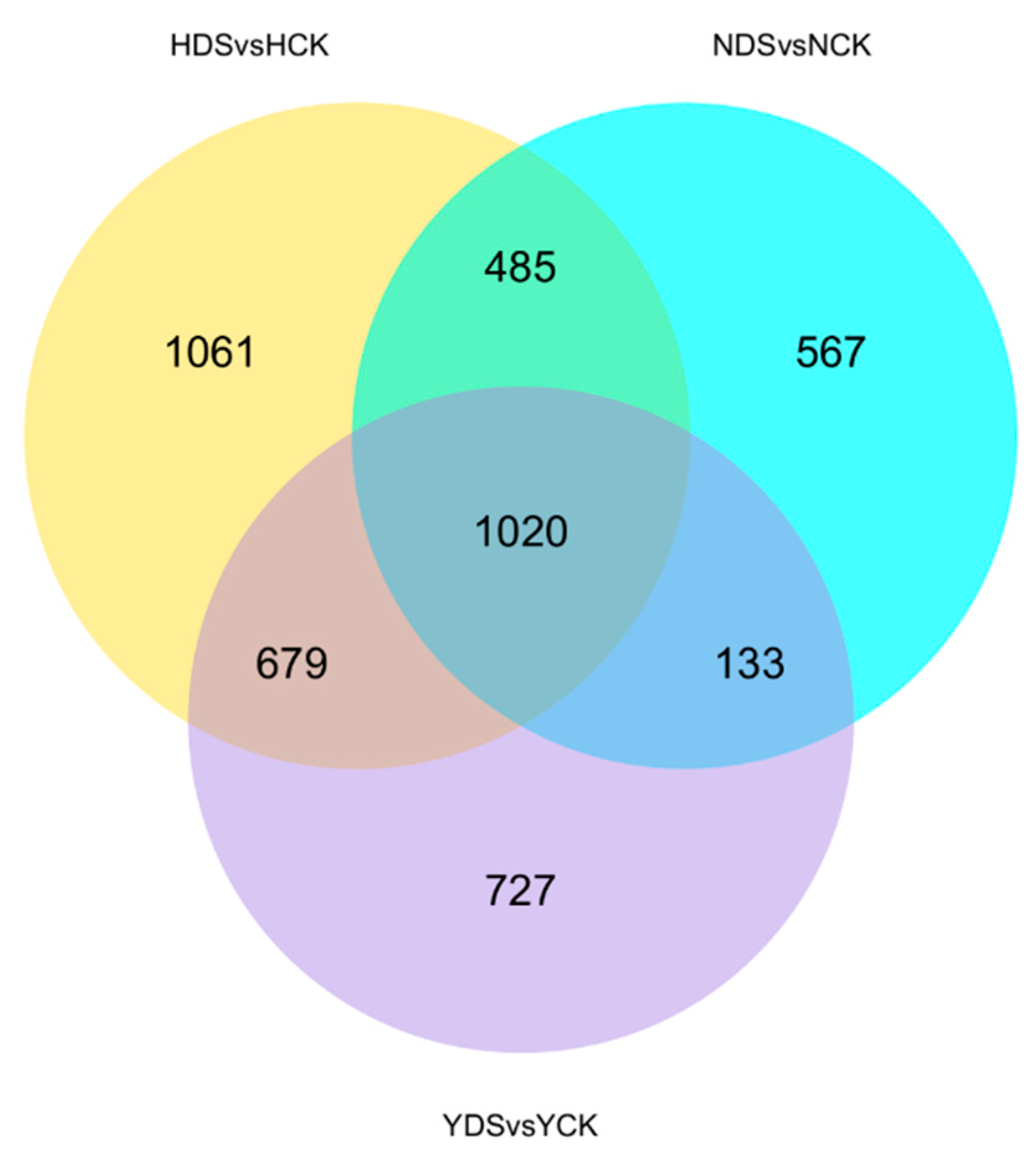
3.8. Identification of Differentially Expressed Genes in Three Tobacco Varieties in Response to Drought Stress
3.9. Gene Ontology (GO) and KEGG Enrichment Analyses of Tobacco Transcritomes
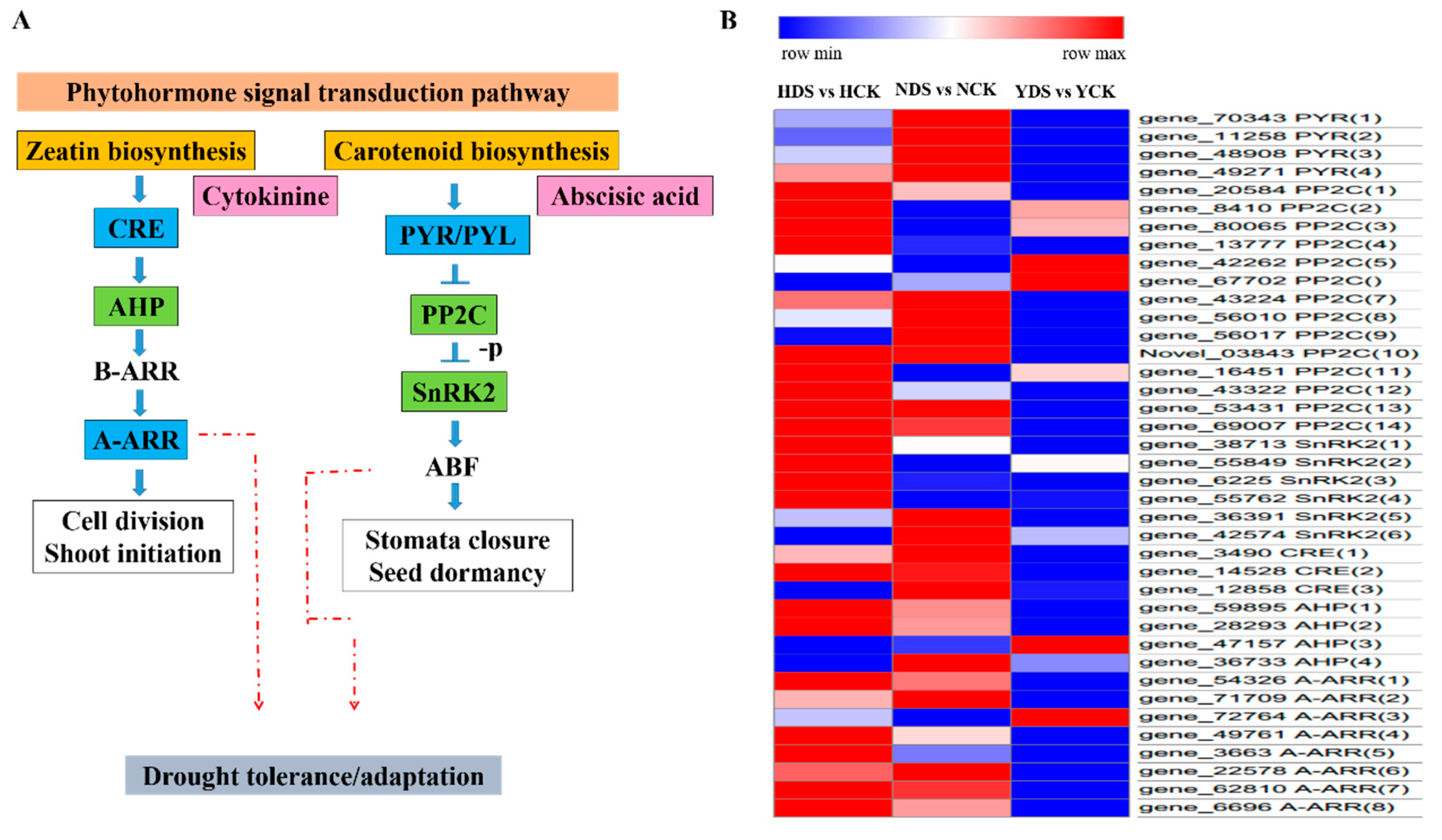
3.10. Plant Hormone Signal Transduction in Response to Drought Stress
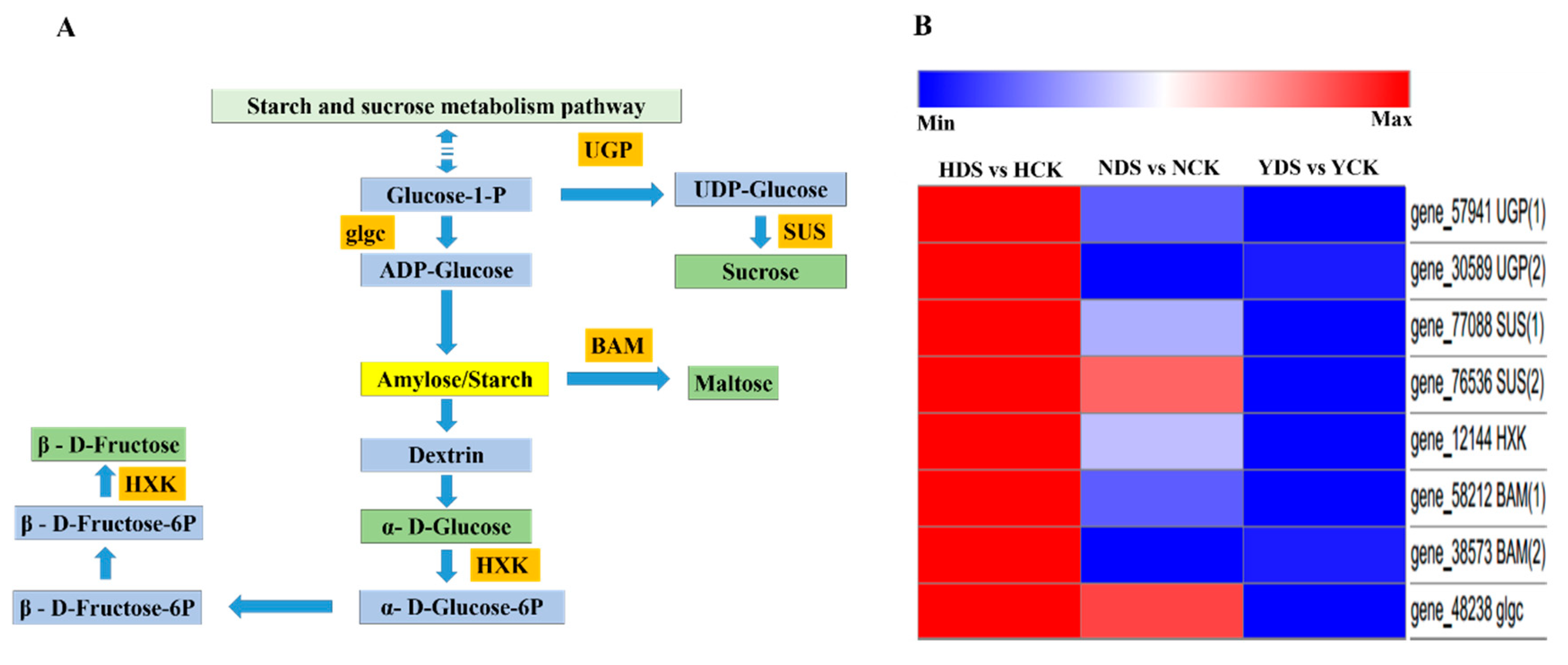
3.11. Starch and Sucrose Metabolism in Response to Drought Stress
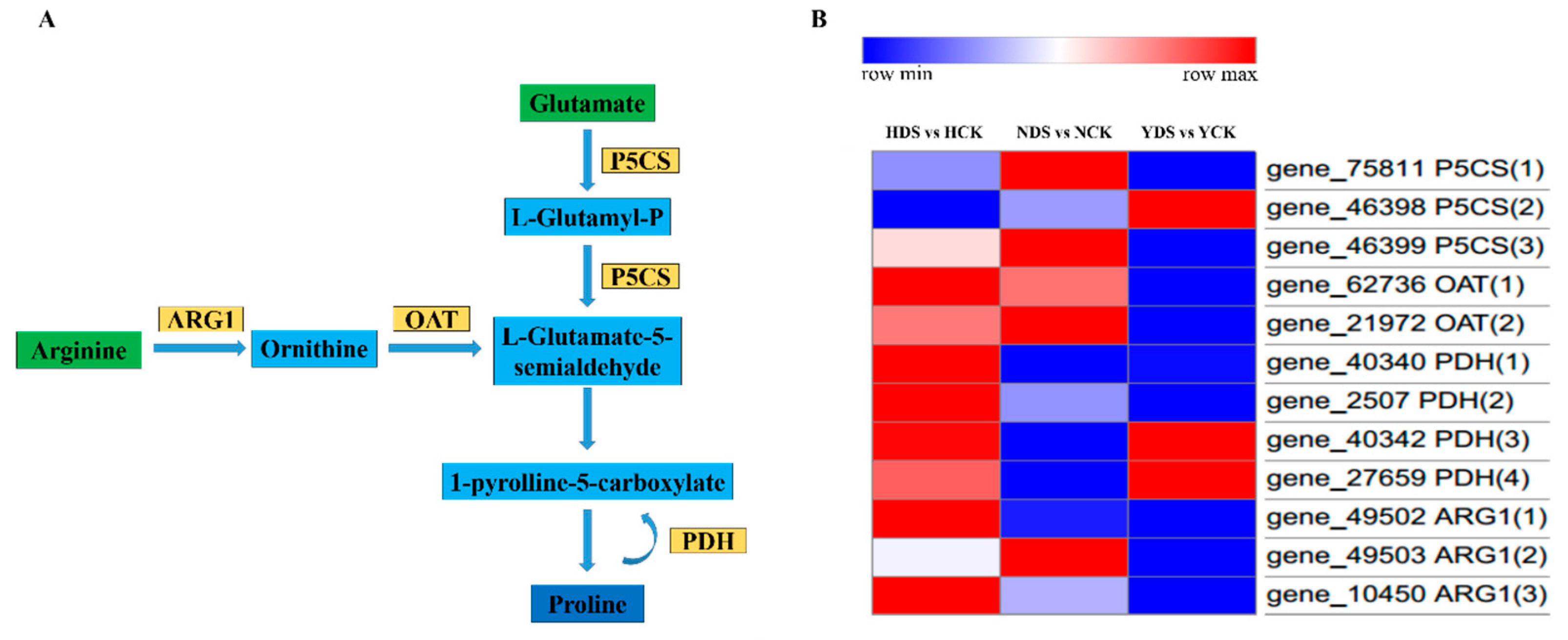
3.12. Proline Metabolism in Response to Drought Stress
4. Discussion
4.1. Growth, Chlorophyll Content and LWP of the Three Tobacco Varieties in Response to Drought Stress
4.2. Chlorophyll Fluorescence and Multicolor Analyses under Drought Stress
4.3. Oxidative Damage Caused by Drought Stress in the Three Tobacco Varieties
4.4. Enzymatic Antioxidant Defense System Alleviates the Drought Stress Response in the Three Tobacco Varieties
4.5. Non-Enzymatic Antioxidant Defense System Involved in Mitigating the Adverse Effect of Drought Stress
4.6. Proline and Soluble Sugars Improve Drought Tolerance in the Three Tobacco Varieties
4.7. Leaf Anatomical Modifications in the Three Tobacco Varieties under Drought Stress
4.8. Analysis of Plant Hormone Signal Transduction in Response to Drought Stress
4.9. Analysis of Starch and Sucrose Metabolism in Response to Drought Stress
4.10. Analysis of Proline Metabolism in Response to Drought Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lloyd-Hughes, B. The impracticality of a universal drought definition. Theor. Appl. Climatol. 2014, 117, 607–611. [Google Scholar] [CrossRef]
- Van Loon, A.F. Hydrological drought explained. Wiley Interdiscip. Rev. Water 2015, 2, 359–392. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Haider, M.S.; Kurjogi, M.M.; Khalil-ur-Rehman, M.; Pervez, T.; Songtao, J.; Fiaz, M.; Jogaiah, S.; Wang, C.; Fang, J. Drought stress revealed physiological, biochemical and gene-expressional variations in ‘yoshihime’ peach (Prunus persica L) cultivar. J. Plant Interact. 2018, 13, 83–90. [Google Scholar] [CrossRef]
- Lamaoui, M.; Jemo, M.; Datla, R.; Bekkaoui, F. Heat and drought stresses in crops and approaches for their mitigation. Front. Chem. 2018, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Tariq, A.; Pan, K.; Olatunji, O.A.; Graciano, C.; Li, Z.; Sun, F.; Zhang, L.; Wu, X.; Chen, W.; Song, D.; et al. Phosphorous fertilization alleviates drought effects on Alnus cremastogyne by regulating its antioxidant and osmotic potential. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Rushton, P.J.; Bokowiec, M.T.; Han, S.; Zhang, H.; Brannock, J.F.; Chen, X.; Laudeman, T.W.; Timko, M.P. Tobacco transcription factors: Novel insights into transcriptional regulation in the solanaceae. Plant Physiol. 2008, 147, 280–295. [Google Scholar] [CrossRef] [PubMed]
- Potopová, V.; Boroneanţ, C.; Boincean, B.; Soukup, J. Impact of agricultural drought on main crop yields in the republic of moldova. Int. J. Climatol. 2016, 36, 2063–2082. [Google Scholar] [CrossRef]
- Peng, S.Z.; Gao, X.L.; Yang, S.H.; Yang, J.; Zhang, H.X. Water requirement pattern for tobacco and its response to water deficit in guizhou province. Water Sci. Eng. 2015, 8, 96–101. [Google Scholar] [CrossRef][Green Version]
- Biglouei, M.H.; Assimi, M.H.; Akbarzadeh, A. Effect of water stress at different growth stages on quantity and quality traits of virginia (flue-cured) tobacco type. Plant Soil Environ. 2010, 56, 67–75. [Google Scholar] [CrossRef]
- de Carvalho, M.H.C. Drought stress and reactive oxygen species. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, D.; Sharma, R.; Handa, N.; Kaur, H.; Rattan, A.; Yadav, P.; Gautam, V.; Kaur, R.; Bhardwaj, R. Redox homeostasis in plants under abiotic stress: Role of electron carriers, energy metabolism mediators and proteinaceous thiols. Front. Environ. Sci. 2015, 3, 1–12. [Google Scholar] [CrossRef]
- Tiwari, S.; Lata, C.; Chauhan, P.S.; Prasad, V.; Prasad, M.A. Functional genomic perspective on drought signalling and its crosstalk with phytohormone-mediated signalling pathways in plants. Curr. Genom. 2017, 18, 469–482. [Google Scholar] [CrossRef]
- Yang, Z.; Dai, Z.; Lu, R.; Wu, B.; Tang, Q.; Xu, Y.; Cheng, C.; Su, J. Transcriptome analysis of two species of jute in response to polyethylene glycol (PEG)- induced drought stress. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Wang, M.; Jiang, B.; Peng, Q.; Liu, W.; He, X.; Liang, Z.; Lin, Y. Transcriptome analyses in different cucumber cultivars provide novel insights into drought stress responses. Int. J. Mol. Sci. 2018, 19, 2067. [Google Scholar] [CrossRef]
- Baldoni, E.; Bagnaresi, P.; Locatelli, F.; Mattana, M.; Genga, A. Comparative leaf and root transcriptomic analysis of two rice japonica cultivars reveals major differences in the root early response to osmotic stress. Rice 2016, 9, 1–20. [Google Scholar] [CrossRef]
- Zhao, X.; Li, C.; Wan, S.; Zhang, T.; Yan, C.; Shan, S. Transcriptomic analysis and discovery of genes in the response of Arachis hypogaea to drought stress. Mol. Biol. Rep. 2018, 45, 119–131. [Google Scholar] [CrossRef]
- Huang, X.; Hou, L.; Meng, J.; You, H.; Li, Z.; Gong, Z.; Yang, S.; Shi, Y. The antagonistic action of abscisic acid and cytokinin signaling mediates drought stress response in arabidopsis. Mol. Plant 2018, 11, 970–982. [Google Scholar] [CrossRef]
- Li, P.; Yang, H.; Wang, L.; Liu, H.; Huo, A.H.; Zhang, C.; Liu, A.; Zhu, A.; Hu, J.; Lin, Y.; et al. Physiological and transcriptome analyses reveal short-term responses and formation of memory under drought stress in rice. Front. Genet. 2019, 10. [Google Scholar] [CrossRef]
- Naing, A.H.; Ai, T.N.; Lee, I.J.; Lim, K.B.; Kim, C.K. Overexpression of Rosea1 from snapdragon enhances anthocyanin accumulation and abiotic stress tolerance in transgenic tobacco. Front. Plant Sci. 2019, 9, 1070. [Google Scholar] [CrossRef] [PubMed]
- Rabara, R.C.; Tripathi, P.; Rushton, P.J. Comparative metabolome profile between tobacco and soybean grown under water-stressed conditions. Biomed Res. Int. 2017. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Wei, F.; Huo, Y.; Xia, Z. Comparative physiological and molecular analyses of two contrasting flue-cured tobacco genotypes under progressive drought stress. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xiao, Y.; Wang, X.; Pei, Y. Expression of a novel small antimicrobial protein from the seeds of motherwort (Leonurus japonicus) confers disease resistance in tobacco. Appl. Environ. Microbiol. 2007, 73, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.M.U.; Ma, F.; Islam, F.; Sajid, M.; Prodhan, Z.H.; Li, F.; Shen, H.; Chen, Y.; Wang, X. Comparative Transcriptomic Analysis of Biological Process and Key Pathway in Three Cotton (Gossypium spp.) Species under Drought Stress. Int. J. Mol. Sci. 2019, 20, 2076. [Google Scholar] [CrossRef]
- You, J.; Zhang, Y.; Liu, A.; Li, D.; Wang, X.; Dossa, K.; Zhou, R.; Yu, J.; Zhang, Y.; Wang, L.; et al. Transcriptomic and metabolomic profiling of drought-tolerant and susceptible sesame genotypes in response to drought stress. BMC Plant Biol. 2019, 19, 267. [Google Scholar] [CrossRef]
- Li, Q.; Wang, G.; Guan, C.; Yang, D.; Wang, Y.; Zhang, Y.; Ji, J.; An, T. Overexpression of LcSABP, an Orthologous Gene for Salicylic Acid Binding Protein 2, Enhances Drought Stress Tolerance in Transgenic Tobacco. Front. Plant Sci. 2019, 10, 200. [Google Scholar] [CrossRef]
- Liu, S.C.; Jin, J.Q.; Ma, J.Q.; Yao, M.Z.; Ma, C.L.; Li, C.F.; Ding, Z.T.; Chen, L. Transcriptomic analysis of tea plant responding to drought stress and recovery. PLoS ONE 2016, 11, e0147306. [Google Scholar] [CrossRef]
- Shah, S.H.; Houborg, R.; McCabe, M.F. Response of chlorophyll, carotenoid and SPAD-502 measurement to salinity and nutrient stress in wheat (Triticum aestivum L.). Agronomy 2017, 7, 61. [Google Scholar] [CrossRef]
- Yao, J.; Sun, D.; Cen, H.; Xu, H.; Weng, H.; Yuan, F.; He, Y. Phenotyping of arabidopsis drought stress response using kinetic chlorophyll fluorescence and multicolor fluorescence imaging. Front. Plant Sci. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Güler, N.S.; Sağlam, A.; Demiralay, M.; Kadioğlu, A. Apoplastic and symplastic solute concentrations contribute to osmotic adjustment in bean genotypes during drought stress. Turk. J. Biol. 2012, 36, 151–160. [Google Scholar]
- Yang, L.; Tang, R.; Zhu, J.; Liu, H.; Mueller-Roeber, B.; Xia, H.; Zhang, H. Enhancement of stress tolerance in transgenic tobacco plants constitutively expressing AtIpk2β, an inositol polyphosphate 6-/3-kinase from Arabidopsis thaliana. Plant Mol. Biol. 2008, 66, 329–343. [Google Scholar] [CrossRef] [PubMed]
- Erdem, H.U.; Kaln, R.; Ozdemir, N.; Ozdemir, H. Purification and biochemical characterization of peroxidase isolated from white cabbage (Brassica Oleracea Var. Capitata f. Alba). Int. J. Food Prop. 2015, 18, 2099–2109. [Google Scholar] [CrossRef]
- Arbona, V.; Hossain, Z.; López-Climent, M.F.; Pérez-Clemente, R.M.; Gómez-Cadenas, A. Antioxidant enzymatic activity is linked to waterlogging stress tolerance in citrus. Physiol. Plant. 2008, 132, 452–466. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.M.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Trehalose-induced drought stress tolerance: A comparative study among different brassica species. Plant Omics 2014, 7, 271–283. [Google Scholar]
- Cao, Y.; Luo, Q.; Tian, Y.; Meng, F. Physiological and proteomic analyses of the drought stress response in Amygdalus Mira (Koehne) Yü et Lu roots. BMC Plant Biol. 2017, 17, 1–16. [Google Scholar] [CrossRef]
- Bistgani, Z.E.; Siadat, S.A.; Bakhshandeh, A.; Pirbalouti, A.G.; Hashemi, M. Interactive Effects of drought stress and chitosan application on physiological characteristics and essential oil yield of Thymus daenensis Celak. Crop J. 2017, 5, 407–415. [Google Scholar] [CrossRef]
- Xia, Z.; Xu, Z.; Wei, Y.; Wang, M. Overexpression of the maize sulfite oxidase increases sulfate and GSH levels and enhances drought tolerance in transgenic tobacco. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Dawood, M.G.; Taie, H.A.A.; Nassar, R.M.A.; Abdelhamid, M.T.; Schmidhalter, U. The changes induced in the physiological, biochemical and anatomical characteristics of vicia faba by the exogenous application of proline under seawater stress. S. Afr. J. Bot. 2014, 93, 54–63. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and abundance estimation from RNA-Seq reveals thousands of new transcripts and switching among isoforms. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.; Cartelat, A.; Moya, I.; Cerovic, Z.G. UV-Induced Blue-Green and Far-Red Fluorescence along Wheat Leaves: A Potential Signature of Leaf Ageing. J. Exp. Bot. 2003, 54, 757–769. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Li, L.; Wang, Y.; Chen, Q.; Zhang, W.; Jia, G.; Zhi, H.; Zhao, B.; Diao, X. Genotype-specifc physiological and transcriptomic responses to drought stress in Setaria italica (an emerging model for Panicoideae grasses). Sci. Rep. 2017, 7, 10009. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Ma, H.; Chen, S.; Gu, T.; Gong, J. Control of proline accumulation under drought via a novel pathway comprising the histone methylase CAU1 and the transcription factor ANAC055. J. Exp. Bot. 2018, 69, 579–588. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of Proline under Changing Environments: A Review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Shtereva, L.; Stoimenova, E.; Drumeva-Yoncheva, M.; Michailova, B.; Kartzeva, T.; Vassilevska-Ivanova, R. Response of Virginia (Flue-Cured) Tobacco Genotypes to Water-Deficit Stress. Arch. Biol. Sci. 2017, 69, 119–127. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Ottosen, C.O.; Rosenqvist, E.; Zhao, L.; Wang, Y.; Yu, W.; Zhao, T.; Wu, Z. Drought Stress Had a Predominant Effect over Heat Stress on Three Tomato Cultivars Subjected to Combined Stress. BMC Plant Biol. 2017, 17, 1–13. [Google Scholar] [CrossRef]
- Kamble, P.N.; Giri, S.P.; Mane, R.S.; Tiwana, A. Estimation of Chlorophyll Content in Young and Adult Leaves of Some Selected Plants. Univ. J. Environ. Res. Technol. 2015, 5, 306–310. [Google Scholar]
- Chen, D.; Wang, S.; Cao, B.; Cao, D.; Leng, G.; Li, H.; Yin, L.; Shan, L.; Deng, X. Genotypic Variation in Growth and Physiological Response to Drought Stress and Re-Watering Reveals the Critical Role of Recovery in Drought Adaptation in Maize Seedlings. Front. Plant Sci. 2016, 6, 1–15. [Google Scholar] [CrossRef]
- Batra, N.G.; Sharma, V.; Kumari, N. Drought-Induced Changes in Chlorophyll Fluorescence, Photosynthetic Pigments, and Thylakoid Membrane Proteins of Vigna radiata. J. Plant Interact. 2014, 9, 712–721. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence—A Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Kasajima, I.; Suetsugu, N.; Wada, M.; Takahara, K. Collective Calculation of Actual Values of Non-Photochemical Quenching from Their Apparent Values after Chloroplast Movement and Photoinhibition. Am. J. Plant Sci. 2015, 06, 1792–1805. [Google Scholar] [CrossRef]
- Urban, L.; Aarrouf, J.; Bidel, L.P.R. Assessing the Effects of Water Deficit on Photosynthesis Using Parameters Derived from Measurements of Leaf Gas Exchange and of Chlorophyll a Fluorescence. Front. Plant Sci. 2017, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Abid, G.; M’hamdi, M.; Mingeot, D.; Aouida, M.; Aroua, I.; Muhovski, Y.; Sassi, K.; Souissi, F.; Mannai, K.; Jebara, M. Effect of Drought Stress on Chlorophyll Fluorescence, Antioxidant Enzyme Activities and Gene Expression Patterns in Faba Bean (Vicia faba L.). Arch. Agron. Soil Sci. 2017, 63, 536–552. [Google Scholar] [CrossRef]
- Mahler, H.; Wuennenberg, P.; Linder, M.; Przybyla, D.; Zoerb, C.; Landgraf, F.; Forreiter, C. Singlet Oxygen Affects the Activity of the Thylakoid ATP Synthase and Has a Strong Impact on Its γ Subunit. Planta 2007, 225, 1073–1083. [Google Scholar] [CrossRef]
- Pineda, M.; Gáspár, L.; Morales, F.; Szigeti, Z.; Barón, M. Multicolor Fluorescence Imaging of Leaves—A Useful Tool for Visualizing Systemic Viral Infections in Plants. Photochem. Photobiol. 2008, 84, 1048–1060. [Google Scholar] [CrossRef]
- Lang, M.; Lichtenthaler, H.K.; Sowinska, M.; Heisel, F.; Miehé, J.A. Fluorescence Imaging of Water and Temperature Stress in Plant Leaves. J. Plant Physiol. 1996, 148, 613–621. [Google Scholar] [CrossRef]
- Buschmann, C.; Lichtenthaler, H.K. Principles and Characteristics of Multi-Colour Fluorescence Imaging of Plants. J. Plant Physiol. 1998, 152, 297–314. [Google Scholar] [CrossRef]
- Yonny, M.E.; Torressi, A.R.; Nazareno, M.A.; Cerutti, S. Development of a Novel, Sensitive, Selective, and Fast Methodology to Determine Malondialdehyde in Leaves of Melon Plants by Ultra-High-Performance Liquid Chromatography-Tandem Mass Spectrometry. J. Anal. Methods Chem. 2017, 9. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Fujita, M. Glutathione-Induced Drought Stress Tolerance in Mung Bean: Coordinated Roles of the Antioxidant Defence and Methylglyoxal Detoxification Systems. AoB Plants 2015, 7, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sultan, M.A.R.F.; Hui, L.; Yang, L.J.; Xian, Z.H. Assessment of Drought Tolerance of Some Triticum l. Species through Physiological Indices. Czech J. Genet. Plant Breed. 2012, 48, 178–184. [Google Scholar] [CrossRef]
- Hossain, M.A.; Bhattacharjee, S.; Armin, S.M.; Qian, P.; Xin, W.; Li, H.Y.; Burritt, D.J.; Fujita, M.; Tran, L.S.P. Hydrogen Peroxide Priming Modulates Abiotic Oxidative Stress Tolerance: Insights from ROS Detoxification and Scavenging. Front. Plant Sci. 2015, 6, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Ashraf, U.; Tanveer, M.; Khan, I.; Hussain, S.; Shahzad, B.; Zohaib, A.; Abbas, F.; Saleem, M.F.; Ali, I.; et al. Drought Induced Changes in Growth, Osmolyte Accumulation and Antioxidant Metabolism of Three Maize Hybrids. Front. Plant Sci. 2017, 8, 1–12. [Google Scholar] [CrossRef]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef]
- Dong, B.; Zheng, X.; Liu, H.; Able, J.A.; Yang, H.; Zhao, H.; Zhang, M.; Qiao, Y.; Wang, Y.; Liu, M. Effects of Drought Stress on Pollen Sterility, Grain Yield, Abscisic Acid and Protective Enzymes in Two Winter Wheat Cultivars. Front. Plant Sci. 2017, 8, 1–14. [Google Scholar] [CrossRef]
- Lee, B.R.; Kim, K.Y.; Jung, W.J.; Avice, J.C.; Ourry, A.; Kim, T.H. Peroxidases and Lignification in Relation to the Intensity of Water-Deficit Stress in White Clover (Trifolium repens L.). J. Exp. Bot. 2007, 58, 1271–1279. [Google Scholar] [CrossRef]
- Awasthi, R.; Gaur, P.; Turner, N.C.; Vadez, V.; Siddique, K.H.M.; Nayyar, H. Effects of Individual and Combined Heat and Drought Stress during Seed Filling on the Oxidative Metabolism and Yield of Chickpea (Cicer arietinum) Genotypes Differing in Heat and Drought Tolerance. Crop Pasture Sci. 2017, 68, 823–841. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones Regulate Accumulation of Osmolytes Under Abiotic Stress. Biomolecules. 2019, 9, 285. [Google Scholar] [CrossRef]
- Mwadzingeni, L.; Shimelis, H.; Tesfay, S.; Tsilo, T.J. Screening of Bread Wheat Genotypes for Drought Tolerance Using Phenotypic and Proline Analyses. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Yuanyuan, M.; Yali, Z.; Jiang, L.; Hongbo, S. Roles of Plant Soluble Sugars and Their Responses to Plant Cold Stress. Afr. J. Biotechnol. 2009, 8, 2004–2010. [Google Scholar]
- Wu, G.Q.; Feng, R.J.; Shui, Q.Z. Effect of Osmotic Stress on Growth and Osmolytes Accumulation in Sugar Beet (Beta vulgaris L.) Plants. Plant Soil Environ. 2016, 62, 189–194. [Google Scholar]
- Wu, Y.; Hong, W.; Chen, Y. Leaf Physiological and Anatomical Characteristics of Two Indicator Species in the Limestone Region of Southern China under Drought Stress. Pak. J. Bot. 2018, 50, 1335–1342. [Google Scholar]
- Das, R.; Bhagawati, K.; Boro, A.; Medhi, T.; Medhi, B.; Bhanisana, R.K. Relative Performance of Plant Cultivars under Respective Water Deficit Adaptation Strategies: A Case Study. Curr. World Environ. 2015, 10, 683–690. [Google Scholar] [CrossRef]
- Hoque, M.I.U.; Uddin, M.N.; Fakir, M.S.A.; Rasel, M. Drought and Salinity Affect Leaf and Root Anatomical Structures in Three Maize Genotypes. J. Bangladesh Agric. Univ. 2018, 16, 47–55. [Google Scholar] [CrossRef]
- Zhang, F.J.; Zhang, K.K.; Du, C.Z.; Li, J.; Xing, Y.X.; Yang, L.T.; Li, Y.R. Effect of Drought Stress on Anatomical Structure and Chloroplast Ultrastructure in Leaves of Sugarcane. Sugar Tech 2015, 17, 41–48. [Google Scholar] [CrossRef]
- Zheng, M.; Meng, Y.; Yang, C.; Zhou, Z.; Wang, Y.; Chen, B. Protein expression changes during cotton fiber elongation in response to drought stress and recovery. Proteomics 2014, 14, 1776–1795. [Google Scholar] [CrossRef]
- Ku, Y.S.; Sintaha, M.; Cheung, M.Y.; Lam, H.M. Plant Hormone Signaling Crosstalks between Biotic and Abiotic Stress Responses. Int. J. Mol. Sci. 2018, 19, 3206. [Google Scholar] [CrossRef]
- Suzuki, N. Hormone Signaling Pathways under Stress Combinations. Plant Signal. Behav. 2016, 11, 1–5. [Google Scholar] [CrossRef]
- Lee, S.C.; Luan, S. ABA Signal Transduction at the Crossroad of Biotic and Abiotic Stress Responses. Plant Cell Environ. 2012, 35, 53–60. [Google Scholar] [CrossRef]
- Zhang, L.; Li, X.; Li, D.; Sun, Y.; Li, Y.; Luo, Q.; Liu, Z.; Wang, J.; Li, X.; Zhang, H.; et al. CARK1 Mediates ABA Signaling by Phosphorylation of ABA Receptors. Cell Discov. 2018, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Cheng, C.; Zhao, L.; Yu, Y.; Tang, Q.; Xin, P.; Liu, T.; Yan, Z.; Guo, Y.; Zang, G. Genome-Wide Expression Profiles of Hemp (Cannabis sativa L.) in Response to Drought Stress. Int. J. Genom. 2018. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, Y.; Shi, Z.; Jin, Y.; Sun, H.; Xie, F.; Zhang, L. Biosynthesis and Signal Transduction of ABA, JA, and BRs in Response to Drought Stress of Kentucky Bluegrass. Int. J. Mol. Sci. 2019, 20, 1289. [Google Scholar] [CrossRef] [PubMed]
- Shinozawa, A.; Otake, R.; Takezawa, D.; Umezawa, T.; Komatsu, K.; Tanaka, K.; Amagai, A.; Ishikawa, S.; Hara, Y.; Kamisugi, Y.; et al. SnRK2 Protein Kinases Represent an Ancient System in Plants for Adaptation to a Terrestrial Environment. Commun. Biol. 2019, 2. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ge, X.; Yang, Z.; Zhang, C.; Zhao, G.; Chen, E.; Liu, J.; Zhang, X.; Li, F. Genome-Wide Identification and Characterization of SnRK2 Gene Family in Cotton (Gossypium hirsutum L.). BMC Genet. 2017, 18, 1–14. [Google Scholar] [CrossRef]
- Acharya, B.R.; Jeon, B.W.; Zhang, W.; Assmann, S.M. Open Stomata 1 (OST1) Is Limiting in Abscisic Acid Responses of Arabidopsis Guard Cells. New Phytol. 2013, 200, 1049–1063. [Google Scholar] [CrossRef]
- Tran, L.P.; Urao, T.; Qin, F.; Maruyama, K.; Kakimoto, T.; Shinozaki, K.; Yamaguchi-shinozaki, K. Functional Analysis of AHK1/ATHK1 and Cytokinin Receptor Histidine Kinases in Response to Abscisic Acid, Drought, and Salt Stress in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 20623–20628. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, M.; Kong, L.; Lv, Y.; Zou, M.; Lu, G.; Cao, J.; Yu, X. Genome-Wide Identification, Phylogeny, Duplication, and Expression Analyses of Two-Component System Genes in Chinese Cabbage (Brassica rapa ssp. pekinensis). DNA Res. 2014, 21, 379–396. [Google Scholar] [CrossRef]
- O’Brien, J.A.; Benková, E. Cytokinin Cross-Talking during Biotic and Abiotic Stress Responses. Front. Plant Sci. 2013, 4, 1–11. [Google Scholar] [CrossRef]
- Thalmann, M.; Santelia, D. Starch as a Determinant of Plant Fitness under Abiotic Stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef]
- Granot, D.; Kelly, G.; Stein, O.; David-Schwartz, R. Substantial Roles of Hexokinase and Fructokinase in the Effects of Sugars on Plant Physiology and Development. J. Exp. Bot. 2014, 65, 809–819. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, F.; Guy, C.L. Β-Amylase Induction and the Protective Role of Maltose During Temperature Shock. Plant Physiol. 2004, 135, 1674–1684. [Google Scholar] [CrossRef] [PubMed]
- Nemati, F.; Ghanati, F.; Ahmadi Gavlighi, H.; Sharifi, M. Comparison of Sucrose Metabolism in Wheat Seedlings during Drought Stress and Subsequent Recovery. Biol. Plant. 2018, 62, 595–599. [Google Scholar] [CrossRef]
- Hayano-Kanashiro, C.; Calderón-Vásquez, C.; Ibarra-Laclette, E.; Herrera-Estrella, L.; Simpson, J. Analysis of Gene Expression and Physiological Responses in Three Mexican Maize Landraces under Drought Stress and Recovery Irrigation. PLoS ONE 2009, 4. [Google Scholar] [CrossRef]
- Wani, S.H.; Singh, N.B.; Haribhushan, A.; Mir, J.I. Compatible Solute Engineering in Plants for Abiotic Stress Tolerance—Role of Glycine Betaine. Curr. Genom. 2013, 14, 157–165. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A Multifunctional Amino Acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Fichman, Y.; Gerdes, S.Y.; Kovács, H.; Szabados, L.; Zilberstein, A.; Csonka, L.N. Evolution of Proline Biosynthesis: Enzymology, Bioinformatics, Genetics, and Transcriptional Regulation. Biol. Rev. 2015, 90, 1065–1099. [Google Scholar] [CrossRef]
- Muthusamy, M.; Uma, S.; Backiyarani, S.; Saraswathi, M.S.; Chandrasekar, A. Transcriptomic Changes of Drought-Tolerant and Sensitive Banana Cultivars Exposed to Drought Stress. Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef]
- Ji-Bao, C.; Yuan-nan, C.; Zhao-Yuan, Z.; Shu-Min, W.; Jing, W.; Lan-Fan, W. Cloning of the OAT Gene and the Correlation between Its Expression and Drought Tolerance in Phaseolus vulgaris L. J. Integr. Agric. 2016, 15, 973–982. [Google Scholar]
- Ribarits, A.; Abdullaev, A.; Tashpulatov, A.; Richter, A.; Heberle-Bors, E.; Touraev, A. Two Tobacco Proline Dehydrogenases Are Differentially Regulated and Play a Role in Early Plant Development. Planta 2007, 225, 1313–1324. [Google Scholar] [CrossRef]
- Bhaskara, G.B.; Yang, T.-H.; Verslues, P.E. Dynamic Proline Metabolism: Importance and Regulation in Water-Limited Environments. Front. Plant Sci. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Ye, T.; Chen, F.; Cheng, Z.; Wang, Y.; Yang, P.; Zhang, Y.; Chan, Z. Manipulation of Arginase Expression Modulates Abiotic Stress Tolerance in Arabidopsis: Effect on Arginine Metabolism and ROS Accumulation. J. Exp. Bot. 2013, 64, 1367–1379. [Google Scholar] [CrossRef] [PubMed]











© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, R.; Zhou, P.; Ma, X.; Zhou, L.; Wu, Y.; Ullah, Z.; Wang, S. Transcriptome Profiling, Biochemical and Physiological Analyses Provide New Insights towards Drought Tolerance in Nicotiana tabacum L. Genes 2019, 10, 1041. https://doi.org/10.3390/genes10121041
Khan R, Zhou P, Ma X, Zhou L, Wu Y, Ullah Z, Wang S. Transcriptome Profiling, Biochemical and Physiological Analyses Provide New Insights towards Drought Tolerance in Nicotiana tabacum L. Genes. 2019; 10(12):1041. https://doi.org/10.3390/genes10121041
Chicago/Turabian StyleKhan, Rayyan, Peilu Zhou, Xinghua Ma, Lei Zhou, Yuanhua Wu, Zia Ullah, and Shusheng Wang. 2019. "Transcriptome Profiling, Biochemical and Physiological Analyses Provide New Insights towards Drought Tolerance in Nicotiana tabacum L." Genes 10, no. 12: 1041. https://doi.org/10.3390/genes10121041
APA StyleKhan, R., Zhou, P., Ma, X., Zhou, L., Wu, Y., Ullah, Z., & Wang, S. (2019). Transcriptome Profiling, Biochemical and Physiological Analyses Provide New Insights towards Drought Tolerance in Nicotiana tabacum L. Genes, 10(12), 1041. https://doi.org/10.3390/genes10121041