Structural and Functional Dissection of the 5′ Region of the Notch Gene in Drosophila melanogaster

Abstract
1. Introduction
2. Materials and Methods
2.1. Fly Stocks
2.2. DNA Clones
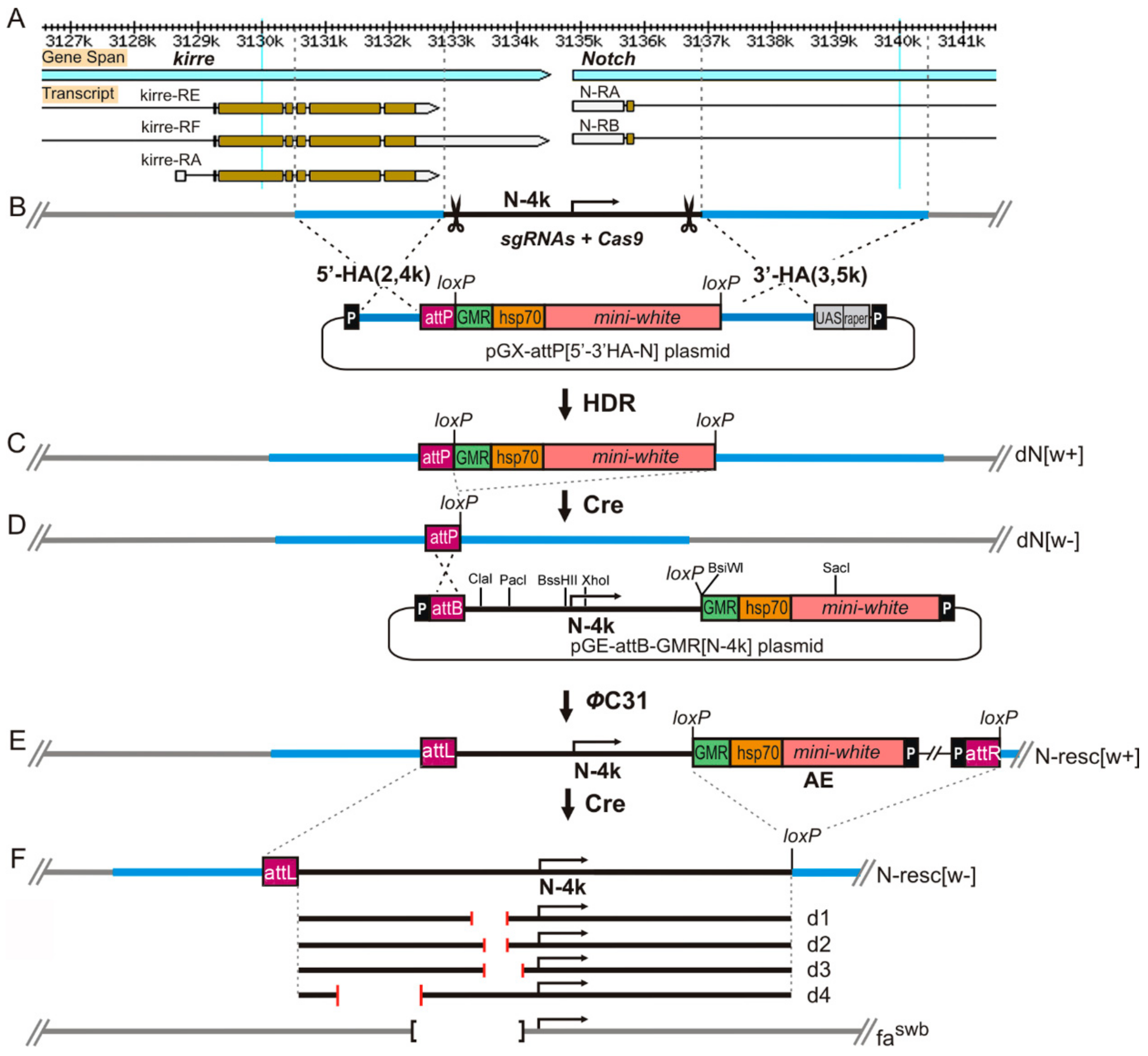
2.3. Donor Construction for Homologous Recombination (HR)
2.4. CRISPR/Cas9 Mediated Homology Directed Reparation and Synthesis of a Founder Line
2.5. Notch Transgenic Constructs
2.6. Immunostaining of Polytene Chromosomes
2.7. Electron Microscopy
3. Results
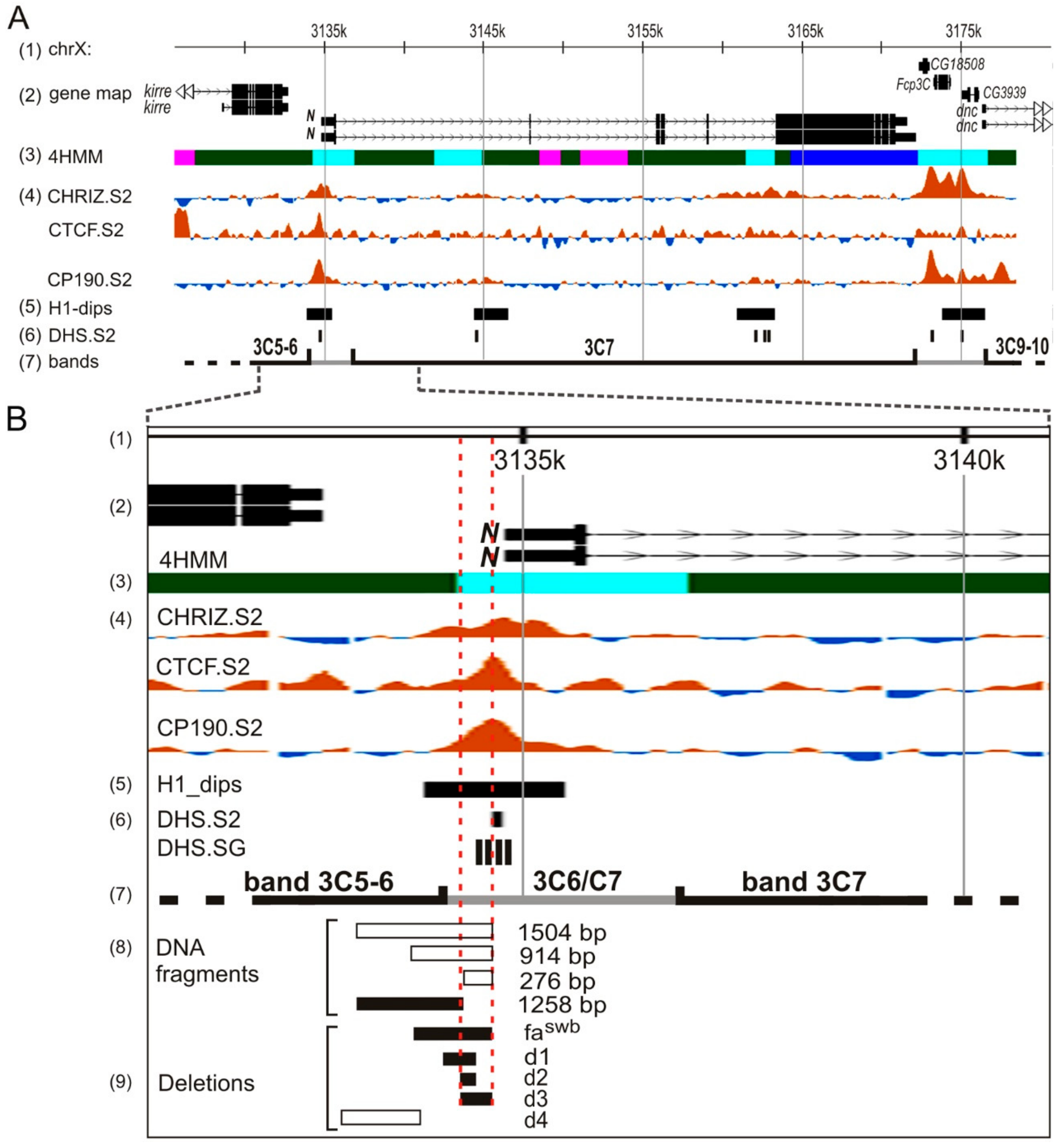
3.1. Custom Deletions of the 5′ Region of the Notch Gene
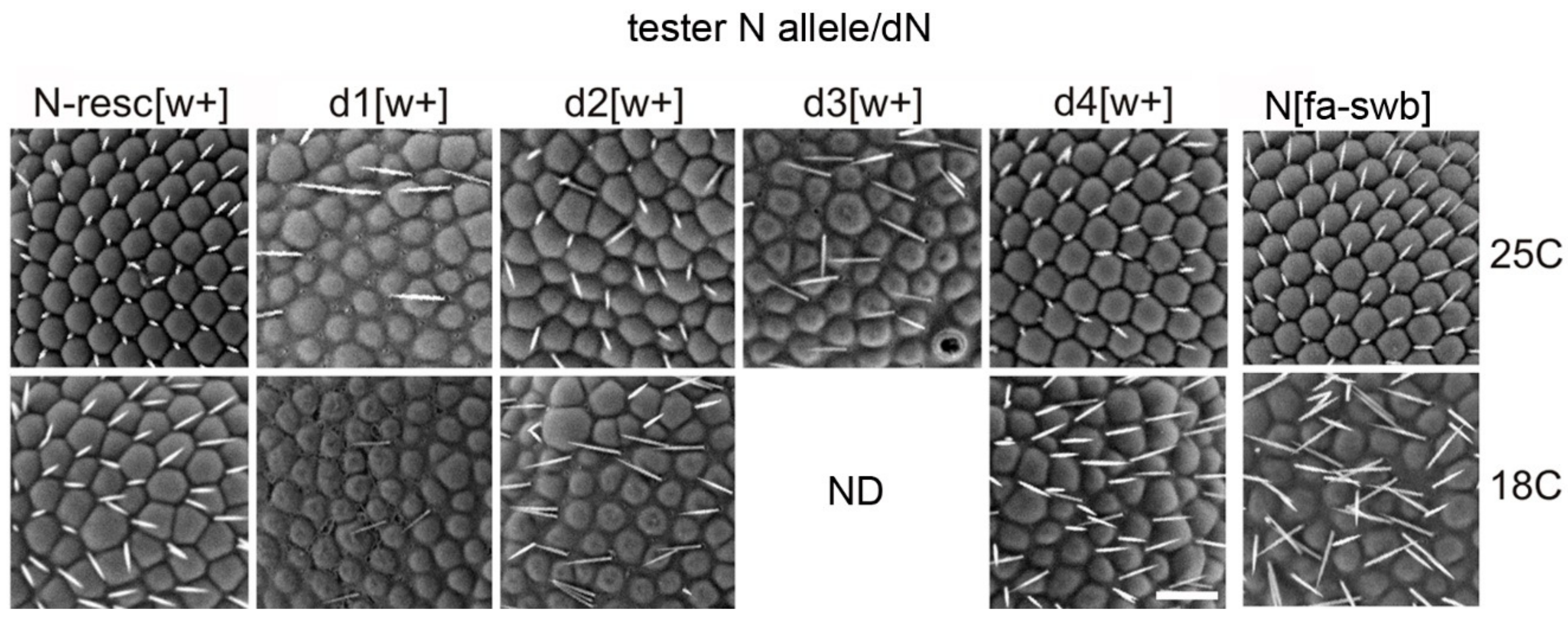
3.2. Flies with Homozygous Deletions Display Mutant Phenotypes Only in the Presence of the AEs in the First Intron of Notch
3.3. In the Absence of a Normal Copy of the Notch Gene the Deletions with AEs Demonstrate Multiple Phenotype Disorders
3.4. Modeling the fa-swb Deletion
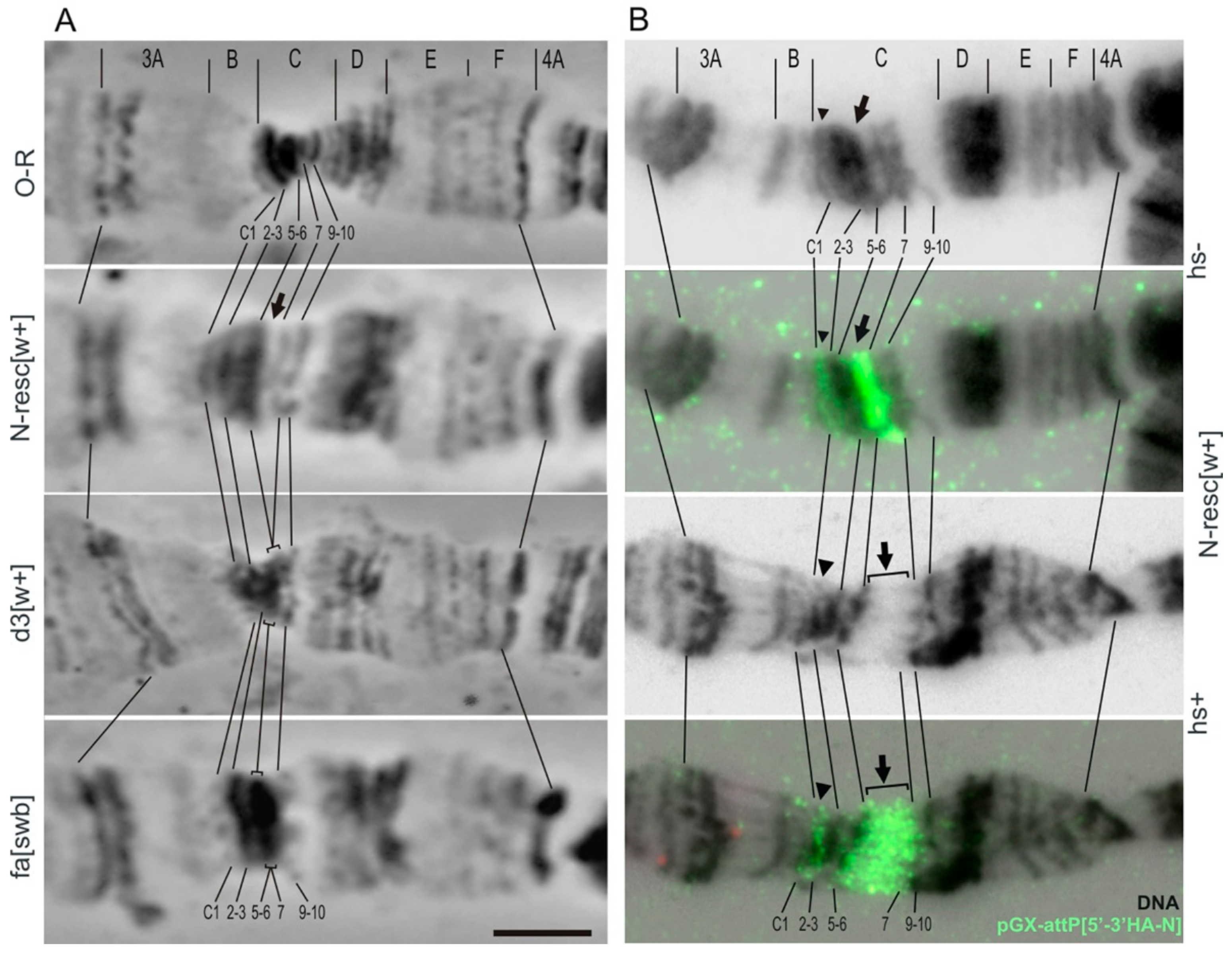
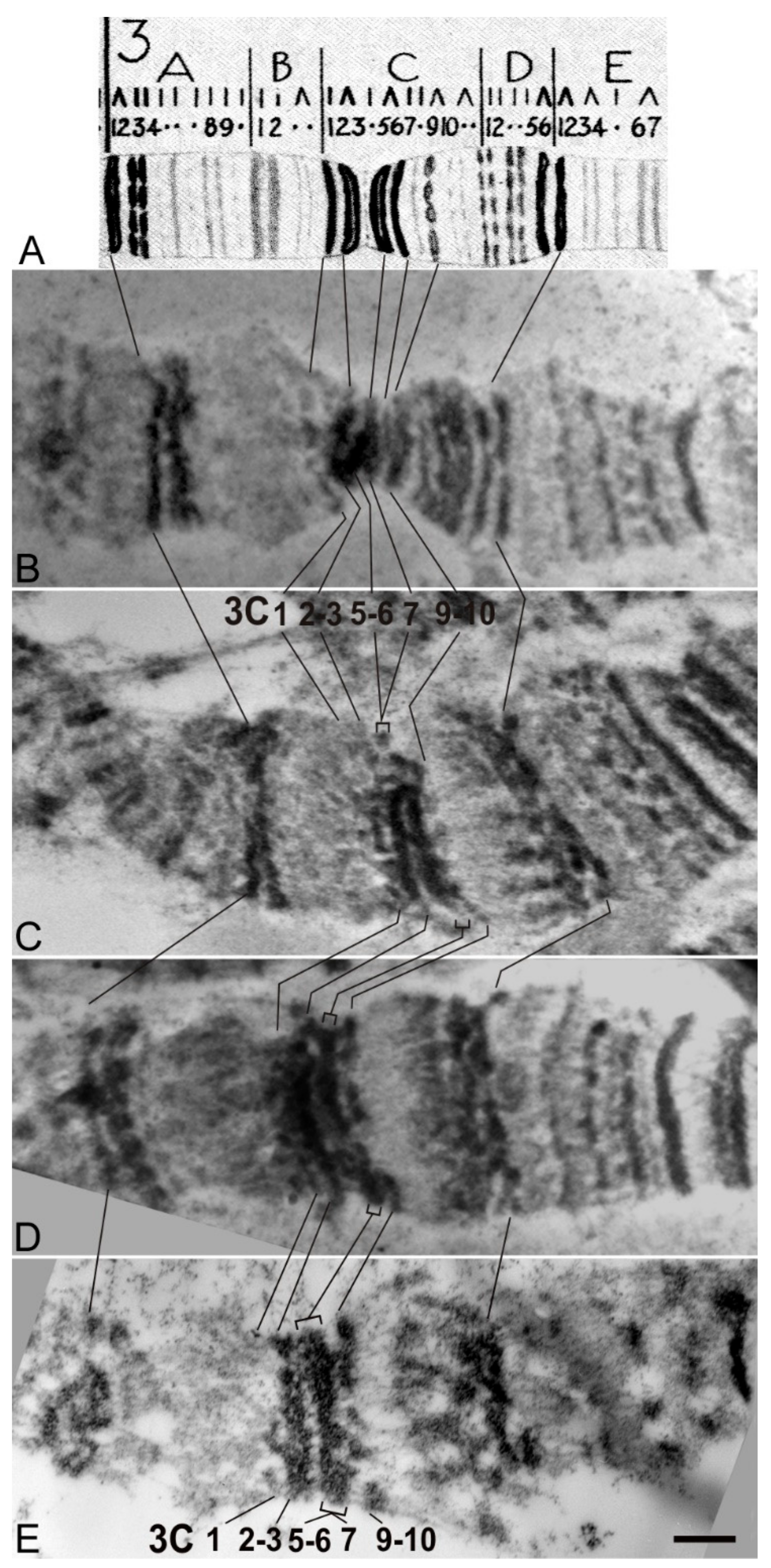
3.5. Cytological Analysis of Mutations in the 5′ Region of the Notch Gene
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fortini, M.E. Notch signaling: The core pathway and its posttranslational regulation. Dev. Cell 2009, 16, 633–647. [Google Scholar] [CrossRef] [PubMed]
- Kopan, R.; Ilagan, M.X. The canonical Notch signaling pathway: Unfolding the activation mechanism. Cell 2009, 137, 216–233. [Google Scholar] [CrossRef] [PubMed]
- Tien, A.C.; Rajan, A.; Bellen, H.J. A Notch updated. J. Cell Biol. 2009, 184, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Artavanis-Tsakonas, S.; Matsuno, K.; Fortini, M.E. Notch signaling. Science 1995, 268, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Portin, P. General outlines of the molecular genetics of the Notch signalling pathway in Drosophila melanogaster: A review. Hereditas 2002, 136, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Heitzler, P.; Simpson, P. The choice of cell fate in the epidermis of Drosophila. Cell 1991, 64, 1083–1092. [Google Scholar] [CrossRef]
- Campuzano, S.; Modolell, J. Patterning of the Drosophila nervous system: The achaete-scute system. Trends Genet. 1992, 8, 202–208. [Google Scholar] [CrossRef]
- Cagan, R.L.; Ready, D.F. Notch is required for successive cell decisions in the developing Drosophila retina. Genes Dev. 1989, 3, 1099–1112. [Google Scholar] [CrossRef]
- Baonza, A.; Freeman, M. Notch signalling and the initiation of neural development in the Drosophila eye. Development 2001, 128, 3889–3898. [Google Scholar]
- Gho, M.; Bellaiche, Y.; Schweisguth, F. Revisitingthe Drosophila microchaete lineage: A novel intrinsically asymmetric cell division generates a glial cell. Development 1999, 126, 3573–3584. [Google Scholar]
- Jordan, K.; Schaeffer, V.; Fischer, K.A.; Gray, E.E.; Ruohola-Baker, H. Notch signaling through Tramtrack bypasses the mitosis promoting activity of the JNK pathway in the mitotic-to-endocycle transition of Drosophila follicle cells. BMC Dev. Biol. 2006, 6, 16. [Google Scholar] [CrossRef]
- López-Schier, H.; St. Johnston, D. Delta signaling from the germ line controls the proliferation and differentiationof the somatic follicle cells during Drosophila oogenesis. Genes Dev. 2001, 15, 1393–1405. [Google Scholar]
- Rauskolb, C.; Irvine, K.D. Notch-mediated segmentation and growth control of the Drosophila leg. Dev. Biol. 1999, 210, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Welshons, W.J.; Keppy, D.O. Intragenic deletions and salivary band relationships in Drosophila. Genetics 1975, 80, 143–155. [Google Scholar] [PubMed]
- Keppy, D.O.; Welshons, W.J. The cytogenetics of a recessive visible mutant associated with a deficiency adjacent to the notch locus in Drosophila melanogaster. Genetics 1977, 85, 497–506. [Google Scholar]
- Rykowski, M.C.; Parmelee, S.J.; Agard, D.A.; Sedat, J.W. Precise determination of the molecular limits of a polytene chromosome band: Regulatory sequences for the Notch gene are in the interband. Cell 1988, 54, 461–472. [Google Scholar] [CrossRef]
- Ramos, R.G.; Grimwade, B.G.; Wharton, K.A.; Scottgale, T.N.; Artavanis-Tsakonas, S. Physical and functional definition of the Drosophila Notch locus by P element transformation. Genetics 1989, 123, 337–348. [Google Scholar]
- Markopoulou, K.; Welshons, W.J.; Artavanis-Tsakonas, S. Phenotypic and molecular analysis of the facets, a group of intronic mutations at the Notch locus of Drosophila melanogaster which affect postembryonic development. Genetics 1989, 122, 417–428. [Google Scholar]
- Welshons, W.J.; Welshons, H.J. Suppression of the facet-strawberry position effect in Drosophila by lesions adjacent to notch. Genetics 1985, 110, 465–477. [Google Scholar]
- Welshons, W.J.; Welshons, H.J. Enhancement and Suppression of a Euchromatic Position Effect at NOTCH in Drosophila. Genetics 1986, 113, 337–354. [Google Scholar]
- Vazquez, J.; Schedl, P. Deletion of an insulator element by the mutation facet-strawberry in Drosophila melanogaster. Genetics 2000, 155, 1297–1311. [Google Scholar] [PubMed]
- Vatolina, T.Y.; Boldyreva, L.V.; Demakova, O.V.; Demakov, S.A.; Kokoza, E.B.; Semeshin, V.F.; Babenko, V.N.; Goncharov, F.P.; Belyaeva, E.S.; Zhimulev, I.F. Identical Functional Organization of Nonpolytene and Polytene Chromosomes in Drosophila melanogaster. PLoS ONE 2011, 6, 1–12. [Google Scholar] [CrossRef]
- Hart, C.M.; Zhao, K.; Laemmli, U.K. The scs’ boundary element: Characterization of boundary element-associated factors. Mol. Cell Biol. 1997, 17, 999–1009. [Google Scholar] [CrossRef] [PubMed]
- Gaszner, M.; Vazquez, J.; Schedl, P. The Zw5 protein, a component of the scs chromatin domain boundary, is able to block enhancer-promoter interaction. Genes Dev. 1999, 13, 2098–2107. [Google Scholar] [CrossRef]
- Braunschweig, U.; Hogan, G.J.; Pagie, L.; van Steensel, B. Histone H1 binding isinhibited by histone variant H3.3. EMBO J. 2009, 28, 3635–3645. [Google Scholar] [CrossRef] [PubMed]
- Filion, G.J.; van Bemmel, J.G.; Braunschweig, U.; Talhout, W.; Kind, J.; Ward, L.D.; Brugman, W.; de Castro, I.J.; Kerkhoven, R.M.; Bussemaker, H.J.; et al. Systematic protein location mapping reveals five principal chromatin types in Drosophila cells. Cell 2010, 143, 212–224. [Google Scholar] [CrossRef]
- Kharchenko, P.V.; Alekseyenko, A.A.; Schwartz, Y.B.; Minoda, A.; Riddle, N.C.; Ernst, J.; Sabo, P.J.; Larschan, E.; Gorchakov, A.A.; Gu, T.; et al. Comprehensive analysis of the chromatin landscape in Drosophila melanogaster. Nature 2011, 471, 480–485. [Google Scholar] [CrossRef]
- Demakov, S.A.; Vatolina, T.Y.; Babenko, V.N.; Semeshin, V.F.; Belyaeva, E.S.; Zhimulev, I.F. Protein composition of interband regions in polytene and cell line chromosomes of Drosophila melanogaster. BMC Genom. 2011, 12, 566. [Google Scholar] [CrossRef]
- Hou, C.; Li, L.; Qin, Z.S.; Corces, V.G. Gene density, transcription, and insulators contribute to the partition of the Drosophila genome into physical domains. Mol. Cell 2012, 48, 471–484. [Google Scholar] [CrossRef]
- Sexton, T.; Yaffe, E.; Kenigsberg, E.; Bantignies, F.; Leblanc, B.; Hoichman, M.; Parrinello, H.; Tanay, A.; Cavalli, G. Three-dimensional folding and functional organization principles of the Drosophila genome. Cell 2012, 148, 458–472. [Google Scholar] [CrossRef]
- Stadler, M.R.; Haines, J.E.; Eisen, M.B. Convergence of topological domain boundaries, insulators, and polytene interbands revealed by high-resolution mapping of chromatin contacts in the early Drosophila melanogaster embryo. eLife 2017, 6, e29550. [Google Scholar] [CrossRef] [PubMed]
- Gratz, S.J.; Ukken, F.P.; Rubinstein, C.D.; Thiede, G.; Donohue, L.K.; Cummings, A.M.; O’Connor-Giles, K.M. Highly specific and efficient CRISPR/Cas9-catalyzed homology-directed repair in Drosophila. Genetics 2014, 196, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Laktionov, P.P.; White-Cooper, H.; Maksimov, D.A.; Belyakin, S.N. Transcription factor COMR acts as a direct activator in the genetic program controlling spermatogenesis in D. melanogaster. Mol. Biol. 2014, 48, 153–165. [Google Scholar] [CrossRef]
- Huang, J.; Zhou, W.; Watson, A.M.; Jan, Y.N.; Hong, Y. Efficient ends-out gene targeting in Drosophila. Genetics 2008, 180, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Port, F.; Chen, H.M.; Lee, T.; Bullock, S.L. Optimized CRISPR/Cas tools for efficient germline and somatic genome engineering in Drosophila. Proc. Natl. Acad. Sci. USA 2014, 111, E2967–E2976. [Google Scholar] [CrossRef] [PubMed]
- Kolesnikova, T.D.; Posukh, O.V.; Andreyeva, E.N.; Bebyakina, D.S.; Ivankin, A.V.; Zhimulev, I.F. Drosophila SUUR protein associates with PCNA and binds chromatin in a cell cycle-dependent manner. Chromosoma 2013, 122, 55–66. [Google Scholar] [CrossRef]
- Semeshin, V.F.; Baricheva, E.M.; Belyaeva, E.S.; Zhimulev, I.F. Electron microscopical analysis of Drosophila Polytene chromosomes. I. Mapping of the 87A and 87C heat shock puffs in development. Chromosoma 1982, 87, 229–237. [Google Scholar] [CrossRef]
- Andreenkov, O.V.; Volkova, E.I.; Semeshin, V.F.; Zhimulev, I.F.; Demakov, S.A. Structural features of chromatin organization 3C6/C7 interband in Drosophila melanogaster polytene chromosome. Tsitologiya 2013, 55, 198–203. [Google Scholar] [CrossRef]
- Kroeger, H. Hypo- and hyperdevelopment of the male genital apparatus and the Bd-M combination in Drosophila melanogaster. J. Morphol. 1960, 107, 227–232. [Google Scholar] [CrossRef]
- Lefevre, G.; Kelley, J. Strawberry vs. facet-glossy, a locus correction. Drosophila Inf. Serv. 1972, 48, 146–147. [Google Scholar]
- Demakov, S.A.; Gortchakov, A.; Schwartz, Y.; Semeshin, V.F.; Campuzano, S.; Modolell, J.; Zhimulev, I. Molecular and genetic organization of Drosophila melanogaster polytene chromosomes: Evidence for two types of interband regions. Genetica 2004, 122, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Vogelmann, J.; Le Gall, A.; Dejardin, S.; Allemand, F.; Gamot, A.; Labesse, G.; Cuvier, O.; Nègre, N.; Cohen-Gonsaud, M.; Margeat, E.; et al. Chromatin insulator factors involved in long-range DNA interactions and their role in the folding of the Drosophila genome. PLoS Genet. 2014, 10, e1004544. [Google Scholar] [CrossRef]
- Celniker, S.E.; Dillon, L.A.; Gerstein, M.B.; Gunsalus, K.C.; Henikoff, S.; Karpen, G.H.; Kellis, M.; Lai, E.C.; Lieb, J.D.; MacAlpine, D.M.; et al. modENCODE Consortium. Unlocking the secrets of the genome. Nature 2009, 18, 927–930. [Google Scholar] [CrossRef] [PubMed]
- Karagodin, D.A.; Battulina, N.V.; Merkulova, T.I.; Baricheva, E.M. The reasons of Trithorax-like expression disturbance in Trl 3609 allele of Drosophila melanogaster. Dokl. Biochem. Biophys. 2016, 471, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Eagen, K.P.; Hartl, T.A.; Kornberg, R.D. Stable Chromosome Condensation Revealed by Chromosome Conformation Capture. Cell 2015, 163, 934–946. [Google Scholar] [CrossRef] [PubMed]
- Zhimulev, I.F.; Zykova, T.Y.; Goncharov, F.P.; Khoroshko, V.A.; Demakova, O.V.; Semeshin, V.F.; Pokholkova, G.V.; Boldyreva, L.V.; Demidova, D.S.; Babenko, V.N.; et al. Genetic organization of interphase chromosome bands and interbands in Drosophila melanogaster. PLoS ONE 2014, 9, e101631. [Google Scholar] [CrossRef] [PubMed]
- Andreyenkov, O.V.; Volkova, E.I.; Demakov, S.A.; Semeshin, V.F.; Zhimulev, I.F. The decompact state of interchromomeric chromatin from the 3C6/C7 region of Drosophila melanogaster is determined by short DNA sequence. Dokl. Biochem. Biophys. 2010, 431, 57–59. [Google Scholar] [CrossRef]
- Keppy, D.O. An Analysis of a Cytologically Aberrant Recessive Visible Mutation at the Notch Locus in Drosophila Melanogaster. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 1975. [Google Scholar]
- Kidd, S.; Kelley, M.R.; Young, M.W. Sequence of the Notch locus of Drosophila melanogaster: Relationship of the encoded protein to mammalian clotting and growth factors. Mol. Cell. Biol. 1986, 6, 3094–3108. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tester N allele | dN[w–] | ||||
|---|---|---|---|---|---|
| Viability(N allele/dNotch:N allele/FM0) | Eyes | Wings | Bristles | Legs | |
| rescue[w+] | 18:366 | R | VTN | Ma | + |
| rescue[w–] | 186:203 | + | + | + | + |
| d3 [w+] | 7:230 | RG | VTN | Ma | C |
| d3 [w–] | 183:175 | R- | + | + | + |
| Nfa-swb | 42:425 | R | VN | H, Ma | C |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volkova, E.I.; Andreyenkova, N.G.; Andreyenkov, O.V.; Sidorenko, D.S.; Zhimulev, I.F.; Demakov, S.A. Structural and Functional Dissection of the 5′ Region of the Notch Gene in Drosophila melanogaster. Genes 2019, 10, 1037. https://doi.org/10.3390/genes10121037
Volkova EI, Andreyenkova NG, Andreyenkov OV, Sidorenko DS, Zhimulev IF, Demakov SA. Structural and Functional Dissection of the 5′ Region of the Notch Gene in Drosophila melanogaster. Genes. 2019; 10(12):1037. https://doi.org/10.3390/genes10121037
Chicago/Turabian StyleVolkova, Elena I., Natalya G. Andreyenkova, Oleg V. Andreyenkov, Darya S. Sidorenko, Igor F. Zhimulev, and Sergey A. Demakov. 2019. "Structural and Functional Dissection of the 5′ Region of the Notch Gene in Drosophila melanogaster" Genes 10, no. 12: 1037. https://doi.org/10.3390/genes10121037
APA StyleVolkova, E. I., Andreyenkova, N. G., Andreyenkov, O. V., Sidorenko, D. S., Zhimulev, I. F., & Demakov, S. A. (2019). Structural and Functional Dissection of the 5′ Region of the Notch Gene in Drosophila melanogaster. Genes, 10(12), 1037. https://doi.org/10.3390/genes10121037