Genome-Wide Association Study of Body Mass Index and Body Fat in Mexican-Mestizo Children

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef]
- Encuesta Nacional de Salud y Nutrición de Medio Camino 2016. Inst. Nac. Salud. Pública 2016, 2016, 151. [CrossRef]
- Costa-Urrutia, P.; Vizuet-Gámez, A.; Ramirez-Alcántara, M.; Guillen-González, M.A.; Medina-Contreras, O.; Valdes-Moreno, M.; Musalem-Younes, C.; Solares-Tlapechco, J.; Granados, J.; Franco-Trecu, V.; et al. Obesity measured as percent body fat, relationship with body mass index, and percentile curves for Mexican pediatric population. PLoS ONE 2019, 14, e0212792. [Google Scholar] [CrossRef]
- Locke, A.; Kahali, B.; Berndt, S.; Justice, A.; Pers, T. Genetic studies of body mass index yield new insights for obesity biology. Nature 2015, 518, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Day, F.R.; Loos, R.J.F. Developments in Obesity Genetics in the Era of Genome-Wide Association Studies. Lifestyle Genom 2011, 4, 222–238. [Google Scholar] [CrossRef]
- Bradfield, J.P.; Taal, H.R.; Timpson, N.J.; Scherag, A.; Lecoeur, C.; Warrington, N.M.; Hypponen, E.; Holst, C.; Valcarcel, B.; Thiering, E.; et al. A genome-wide association meta-analysis identifies new childhood obesity loci. Nat. Genet. 2012, 44, 526–531. [Google Scholar] [CrossRef][Green Version]
- Felix, J.F.; Bradfield, J.P.; Monnereau, C.; van Der Valk, R.J.P.; Stergiakouli, E.; Chesi, A.; Gaillard, R.; Feenstra, B.; Thiering, E.; Kreiner-Møller, E.; et al. Genome-wide association analysis identifies three new susceptibility loci for childhood body mass index. Hum. Mol. Genet. 2016, 25, 389–403. [Google Scholar] [CrossRef]
- Geng, T.; Smith, C.E.; Li, C.; Huang, T. Childhood BMI and adult type 2 diabetes, coronary artery diseases, chronic kidney disease, and cardiometabolic traits: A Mendelian randomization analysis. Diabetes Care 2018, 41, 1089–1096. [Google Scholar] [CrossRef]
- Villalobos-Comparán, M.; Villamil-Ramírez, H.; Villarreal-Molina, T.; Larrieta-Carrasco, E.; Leon-Mimila, P.; Romero-Hidalgo, S.; Jacobo-Albavera, L.; Liceaga-Fuentes, A.E.; Campos-Pérez, F.J.; López-Contreras, B.E.; et al. PCSK1 rs6232 is associated with childhood and adult class III obesity in the Mexican population. PLoS ONE 2012, 7, e39037. [Google Scholar] [CrossRef]
- Mejía-Benítez, A.; Klünder-Klünder, M.; Yengo, L.; Meyre, D.; Aradillas, C.; Cruz, E.; Pérez-Luque, E.; Malacara, J.M.; Garay, M.E.; Peralta-Romero, J.; et al. Analysis of the contribution of FTO, NPC1, ENPP1, NEGR1, GNPDA2 and MC4R genes to obesity in Mexican children. BMC Med. Genet. 2013, 14, 1–6. [Google Scholar] [CrossRef]
- Aradillas-García, C.; Cruz, M.; Pérez-Luque, E.; Garay-Sevilla, M.E.; Malacara, J.M.R.A.; Peralta, J.; Burguete-García, A.; Alegría-Torres, J.A. Obesity is associated with the Arg389Gly ADRB1 but not with the Trp64Arg ADRB3 polymorphism in children from San Luis Potosí and León, México. J. Biomed. Res. 2017, 31, 40. [Google Scholar] [CrossRef]
- Abadi, A.R.K.A.N.; Jesus, J.; De Romero, P.; Gomez, J.; Peralta-romero, J.; Suarez, F.; Gomez-zamudio, J.; Burguete-Garcia, A.I.; Cruz, M.; Meyre, D. Assessing the effects of 35 European-derived BMI-associated SNPs in Mexican children: Effects of European BMI SNPs in M Assessing the Effects of 35 European-Derived BMI-Associated SNPs in Mexican Children. Obesity 2016, 24, 1989–1995. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y.; Alyass, A.; Abadi, A.; Peralta-romero, J. Fine-mapping of 98 obesity loci in Mexican children. Int. J. Obes. 2018, 43, 23. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Osorio, A.S.; Aguilar-Lucio, A.O.; Cárdenas-Hernández, H.; Musalem-Younes, C.; Solares-Tlapechco, J.; Costa-Urrutia, P.; Medina-Contreras, O.; Granados, J.; Rodríguez-Arellano, M.E. Polymorphisms in adipokines in mexican children with obesity. Int. J. Endocrinol. 2019, 2019, 4764751. [Google Scholar] [CrossRef]
- Fomon, S.J.; Haschke, F.; Ziegler, E.E.; Nelson, S.E. Body composition of reference children from birth to age 10 years. Am. J. Clin. Nutr. 1982, 35, 1169–1175. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Loh, P.; Danecek, P.; Palamara, P.F.; Fuchsberger, C.; Reshef, A.; Finucane, H.K. Reference-based phasing using the Haplotype Reference Consortium panel. Nat. Genet. 2017, 48, 1443–1448. [Google Scholar] [CrossRef]
- Sayantan, D.; Forer, L.; Sebastian, S.; Carlo Sidore, C.; Locke, A.E.; Alan Kwong, A.; Scott, I.; Vrieze, S.J.; Chew, E.Y.; Levy, S.; et al. Next-generation genotype imputation service and methods. Nat. Genet. 2016, 48, 1284–1287. [Google Scholar] [CrossRef]
- Patterson, N.; Price, A.L.; Reich, D. Population structure and eigenanalysis. PLoS Genet. 2006, 2, 2074–2093. [Google Scholar] [CrossRef]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef]
- Gauderman, W.J.; Morrison, J.M. Quanto 1.1: A Computer Program for Power and Sample Size Calculations for Genetic-Epidemiology Studies. 2006. Available online: http://hydra.usc.edu/gxe (accessed on 15 September 2019).
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed]
- Silva-Zolezzi, I.; Hidalgo-Miranda, A.; Estrada-Gil, J.; Fernandez-Lopez, J.C.; Uribe-Figueroa, L.; Contreras, A.V.; Balam-Ortiz, E.; Del Bosque-Plata, L.; Velazquez-Fernandez, D.; Lara, C.; et al. Analysis of genomic diversity in Mexican Mestizo populations to develop genomic medicine in Mexico. Proc. Natl. Acad. Sci. USA 2009, 106, 8611–8616. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Estrada, A.; Gignoux, C.R.; Fernández-López, J.C.; Zakharia, F.; Sikora, M.; Contreras, A.V.; Acuña-Alonzo, V.; Sandoval, K.; Eng, C.; Romero-Hidalgo, S.; et al. The genetics of Mexico recapitulates Native American substructure and affects biomedical traits. Science 2014, 344, 1280–1285. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Villalobos, H.; Muñoz-Valle, J.F.; González-Martín, A.; Gorostiza, A.; Magaña, M.T.; Páez-Riberos, L.A. Genetic admixture, relatedness, and structure patterns among Mexican populations revealed by the Y-chromosome. Am. J. Phys. Anthropol. 2008, 135, 448–461. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Marignac, V.L.; Valladares, A.; Cameron, E.; Chan, A.; Perera, A.; Globus-Goldberg, R.; Shriver, M.D.; Wacher, N.; Kumate, J.; McKeigue, P.; et al. Admixture in Mexico City: Implications for admixture mapping of Type 2 diabetes genetic risk factors. Hum. Genet. 2007, 120, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Barquera, R.; Martínez-Álvarez, J.C.; Hernández-Zaragoza, D.I.; Bravo-Acevedo, A.; Juárez-Nicolás, F.; Arriaga-Perea, A.J.; Vega-Martínez, M.D.R.; Ortega-Yáñez, A.; Benítez-Arvizu, G.; Arrieta-Bolaños, E.; et al. Genetic diversity of HLA system in six populations from Mexico City Metropolitan Area, Mexico: Mexico City North, Mexico City South, Mexico City East, Mexico City West, Mexico City Center and rural Mexico City. Hum. Immunol. 2019, 5, 1111. [Google Scholar] [CrossRef] [PubMed]
- Costa-Urrutia, P.; Abud, C.; Franco-Trecu, V.; Colistro, V.; Rodríguez-Arellano, M.E.; Vázquez-Pérez, J.; Granados, J.; Seelaender, M. Genetic Obesity Risk and Attenuation Effect of Physical Fitness in Mexican-Mestizo Population: A Case-Control Study. Ann. Hum. Genet. 2017, 81, 106–116. [Google Scholar] [CrossRef]
- Lu, Y.; Loos, R.J.F. Obesity genomics: Assessing the transferability of susceptibility loci across diverse populations. Genome Med. 2013, 5, 55. [Google Scholar] [CrossRef]
- Dulloo, A.G.; Jacquet, J.; Solinas, G.; Montani, J.P.; Schutz, Y. Body composition phenotypes in pathways to obesity and the metabolic syndrome. Int. J. Obes. 2010, 34, S4–S17. [Google Scholar] [CrossRef]
- Azeloglu, E.U.; Iyengar, R. Signaling networks: Information flow, computation, and decision making. Cold Spring Harb. Perspect. Biol. 2015, 7, 1–13. [Google Scholar] [CrossRef]
- Larsen, P.J.; Tennagels, N. On ceramides, other sphingolipids and impaired glucose homeostasis. Mol. Metab. 2014, 3, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.; Futerman, A.H. Mammalian ceramide synthases. IUBMB Life 2010, 62, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Fucho, R.; Casals, Ń.; Serra, D.; Herrero, L. Ceramides and mitochondrial fatty acid oxidation in obesity. FASEB J. 2017, 31, 1263–1272. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Kasumov, T.; Gatmaitan, P.; Heneghan, H.M.; Kashyap, S.R.; Schauer, P.R.; Brethauer, S.A.; Kirwan, J.P. Gastric bypass surgery reduces plasma ceramide subspecies and improves insulin sensitivity in severely obese patients. Obesity 2011, 19, 2235–2240. [Google Scholar] [CrossRef]
- Hulpiau, P.; van Roy, F. Molecular evolution of the cadherin superfamily. Int. J. Biochem. Cell Biol. 2009, 41, 349–369. [Google Scholar] [CrossRef]
- Huertas-Vazquez, A.; Plaisier, C.L.; Geng, R.; Haas, B.E.; Lee, J.; Greevenbroek, M.M.; Pajukanta, P.; Taskinen, M.R.; Alagramam, K.N.; Pajukanta, P.; et al. A nonsynonymous SNP within PCDH15 is associated with lipid traits in familial combined hyperlipidemia. Hum. Genet. 2010, 127, 83–89. [Google Scholar] [CrossRef]
- Nazarenko, M.S.; Sleptcov, A.A.; Lebedev, I.N.; Skryabin, N.A.; Markov, A.V.; Golubenko, M.V.; Koroleva, I.A.; Kazancev, A.N.; Barbarash, O.L.; Puzyrev, V.P.; et al. Genomic structural variations for cardiovascular and metabolic comorbidity. Sci. Rep. 2017, 7, 41268. [Google Scholar] [CrossRef]
- Xie, G.; Myint, P.K.; Voora, D.; Laskowitz, D.T.; Shi, P.; Ren, F.; Wang, H.; Yang, Y.; Huo, Y.; Gao, W.; et al. Genome-wide association study on progression of carotid artery intima media thickness over 10 years in a Chinese cohort. Atherosclerosis 2015, 243, 30–37. [Google Scholar] [CrossRef]
- Zong, H.; Armoni, M.; Harel, C.; Karnieli, E.; Pessin, J.E. Cytochrome P-450 CYP2E1 knockout mice are protected against high-fat diet-induced obesity and insulin resistance. Am. J. 2012, 302, 532–539. [Google Scholar] [CrossRef]
- Gade, C.; Dalhoff, K.; Petersen, T.S.; Riis, T.; Schmeltz, C.; Chabanova, E.; Christensen, H.R.; Mikus, G.; Burhenne, J.; Holm, J.C.; et al. Higher chlorzoxazone clearance in obese children compared with nonobese peers. Br. J. Clin. Pharmacol. 2018, 84, 1738–1747. [Google Scholar] [CrossRef]


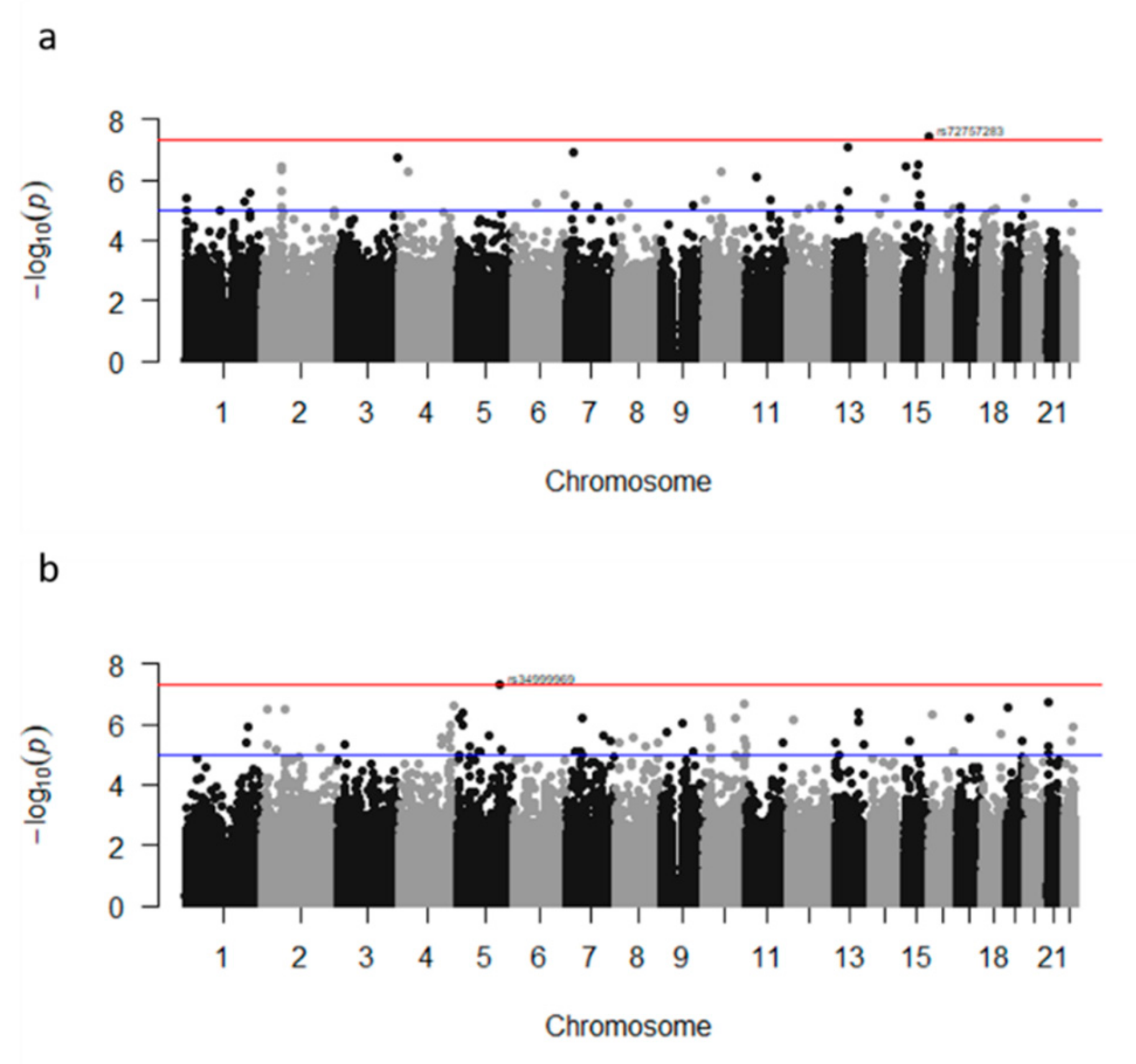{kind=link}
| Age | N | BMI (SD) | %BF (SD) |
|---|---|---|---|
| Girls | |||
| 3 | 22 | 16.6 (1.7) | 0.30 (0.13) |
| 4 | 28 | 15.9 (1.4) | 0.26 (0.14) |
| 5 | 30 | 17.3 (3.4) | 0.23 (0.15) |
| 6 | 30 | 16.3 (1.7) | 0.23 (0.04) |
| 7 | 39 | 17.8 (3.6) | 0.25 (0.13) |
| 8 | 26 | 17.3 (3.0) | 0.26 (0.08) |
| 9 | 24 | 19.2 (2.6) | 0.32 (0.07) |
| 10 | 30 | 20.4 (3.8) | 0.32 (0.10) |
| 11 | 30 | 20.9 (3.9) | 0.31(0.11) |
| 12 | 28 | 22.0 (4.7) | 0.32 (0.09) |
| 13 | 34 | 22.1 (3.9) | 0.33 (0.09) |
| 14 | 37 | 23.8 (4.8) | 0.37 (0.13) |
| 15 | 23 | 23.0 (4.6) | 0.33 (0.08) |
| 16 | 24 | 25.7 (6.1) | 0.33 (0.12) |
| Boys | |||
| 3 | 28 | 16.1 (2.2) | 0.24 (0.16) |
| 4 | 25 | 16.6 (2.9) | 0.22 (0.13) |
| 5 | 28 | 16.5 (2.6) | 0.18 (0.13) |
| 6 | 28 | 16.3 (2.0) | 0.24 (0.11) |
| 7 | 22 | 17.8 (2.3) | 0.27 (0.12) |
| 8 | 40 | 18.3 (3.1) | 0.26 (0.10) |
| 9 | 46 | 20.4 (4.2) | 0.31(0.10) |
| 10 | 29 | 22.6 (5.3) | 0.34 (0.10) |
| 11 | 32 | 21.6 (4.5) | 0.30 (0.11) |
| 12 | 41 | 21.2 (4.9) | 0.28 (0.10) |
| 13 | 28 | 22.3 (4.1) | 0.27 (0.10) |
| 14 | 36 | 21.9 (4.9) | 0.24 (0.12) |
| 15 | 20 | 23.6 (4.1) | 0.26 (0.10) |
| 16 | 20 | 25.5 (4.6) | 0.27 (0.09) |
| Gene | Description | rs | En Id | Chr | As | MA | MAF | P/G | β | Punadj | PBonf | R2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BMI associated | ||||||||||||
| CERS3 | Ceramide synthase 3 | rs72757283 | ENSG00000154227 | 15 | A/C | C | 0.02 | intron | 3.4 | 2.18 × 10−8 | 0.005 | 0.95 |
| CERS3 | rs75661572 | ENSG00000154227 | 15 | T/C | C | 0.16 | intron | 1.3 | 8.97 × 10−8* | 0.020 | ||
| CYP2E1 | Cytochrome P450 family 2 subfamily E member 1 | rs72866768 | ENSG00000130649 | 10 | A/G | G | 0.02 | intron | 5.1 | 3.39 × 10−8 | 0.014 | |
| PCDH15 | Protocadherin related 15 | rs2680328 | ENSG00000150275 | 10 | G/A | G | 0.16 | GUTV | 1.4 | 5.86 × 10−8* | 0.020 | 0.97 |
| PCDH15 | rs2799617 | ENSG00000150275 | 10 | G/A | G | 0.16 | GUTV | 1.7 | 5.86 × 10−8* | 0.024 | ||
| %BF associated | ||||||||||||
| ANKS1B | Ankyrin repeat and sterile alpha motif domain containing 1B | rs116928965 | ENSG00000185046 | 12 | G/A | A | 0.130 | 3’ UTR | 0.2 | 3.27 × 10−8 | 0.012 | 0.60 |
| ARNTL2 | Aryl hydrocarbon receptor nuclear translocator like 2 | rs111392859 | ENSG00000029153 | 12 | G/A | A | 0.012 | intron | 0.2 | 3.61 × 10−8 | 0.014 | |
| ENAM | Enamelin | rs115766831 | ENSG00000132464 | 4 | T/A | A | 0.014 | 3’ UTR | 0.1 | 5.25 × 10−8* | 0.031 | |
| KCNS3 | Potassium voltage-gated channel modifier subfamily S member 3 | rs67939090 | ENSG00000170745 | 2 | A/T | T | 0.012 | intron | 0.2 | 7.31 × 10−11 | 4.89 × 10−5 | 0.90 |
| KCNS3 | rs2198300 | ENSG00000170745 | 2 | A/G | G | 0.012 | intron | 0.2 | 2.49 × 10−11 | 1.66 × 10−5 | ||
| KCNS3 | rs111366249 | ENSG00000170745 | 2 | G/A | A | 0.012 | intron | 0.2 | 3.67 × 10−12 | 2.45 × 10−6 | ||
| KCNS3 | rs34364120 | ENSG00000170745 | 2 | C/A | A | 0.012 | intron | 0.2 | 3.67 × 10−12 | 2.45 × 10−6 | ||
| KCNS3 | rs13002427 | ENSG00000170745 | 2 | A/T | T | 0.012 | intron | 0.2 | 1.26 × 10−11 | 8.40 × 10−6 | ||
| LMNB1 | Lamin B1 | rs140680370 | ENSG00000113368 | 5 | G/A | A | 0.012 | intron | 0.2 | 1.40 × 10−8 | 0.007 | 0.57 |
| MARCH3 | Membrane associated ring-CH-type finger 3 | rs77016412 | ENSG00000173926 | 5 | G/A | A | 0.011 | intron | 0.2 | 9.17 × 10−8* | 0.046 | |
| PHF20L1 | Phd finger protein 20 like 1 | rs78211770 | ENSG00000129292 | 8 | A/G | G | 0.050 | intron | 0.2 | 8.48 × 10−9 | 0.003 | 1 |
| ZC3H3 | Zinc finger CCCH-type containing 3 | rs35110652 | ENSG00000014164 | 8 | C/T | C | 0.015 | GDTV | 0.1 | 5.29 × 0−8* | 0.023 | |
| SLC6A1 | Solute carrier family 6 member 1 | rs58053962 | ENSG00000157103 | 3 | G/C | C | 0.015 | intron | 0.2 | 6.64 × 10−8* | 0.038 | |
| SRGAP3 | Slit-robo Rho GTPase activating protein 3 | rs544274585 | ENSG00000196220 | 3 | C/A | A | 0.015 | intron | 0.2 | 2.31 × 10−8 | 0.0133 | |
| TRPC7 | Transient receptor potential cation channel subfamily C member 7 | rs3756699 | ENSG00000069018 | 5 | A/G | G | 0.012 | intron | 0.1 | 3.53 × 10−8 | 0.0178 | 0.99 |
| TRPC7 | rs55996753 | ENSG00000069018 | 5 | G/A | A | 0.050 | intron | 0.1 | 3.53 × 10−8 | 0.0178 | ||
| TRPC7 | rs34999969 | ENSG00000279240 | 5 | C/A | A | 0.050 | near | 0.1 | 7.18 × 10−8* | 0.036 |
| Biological Process | P |
|---|---|
| BMI | |
| Monoterpenoid metabolic process (GO:0016098) | 9.0 × 10−4 |
| Equilibrioception (GO:0050957) | 9.0 × 10−4 |
| Terpenoid metabolic process (GO:0006721) | 1.0 × 10−3 |
| Benzene-containing compound metabolic process (GO:0042537) | 1.2 × 10−3 |
| Neuromuscular process controlling balance (GO:0050885) | 2.1 × 10−3 |
| Epoxygenase P450 pathway (GO:0019373) | 3.1 × 10−3 |
| Exogenous drug catabolic process (GO:0042738) | 3.4 × 10−3 |
| Drug catabolic process (GO:0042737) | 3.7 × 10−3 |
| Ceramide biosynthetic process (GO:0046513) | 4.9 × 10−3 |
| Ceramide metabolic process (GO:0006672) | 6.4 × 10−3 |
| %BF | |
| Regulation of mRNA export from nucleus (GO:0010793) | 3.3 × 10−3 |
| Regulation of ribonucleoprotein complex localization (GO:2000197) | 3.3 × 10−3 |
| Manganese ion transport (GO:0006828) | 4.9 × 10−3 |
| Positive regulation of circadian rhythm (GO:0042753) | 5.5 × 10−3 |
| Poly(A) + mRNA export from nucleus (GO:0016973) | 7.1 × 10−3 |
| Regulation of RNA export from nucleus (GO:0046831) | 8.2 × 10−3 |
| mRNA polyadenylation (GO:0006378) | 0.02 |
| Transition metal ion transport (GO:0000041) | 0.02 |
| RNA polyadenylation (GO:0043631) | 0.02 |
| Regulation of protein export from nucleus (GO:0046825) | 0.02 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa-Urrutia, P.; Colistro, V.; Jiménez-Osorio, A.S.; Cárdenas-Hernández, H.; Solares-Tlapechco, J.; Ramirez-Alcántara, M.; Granados, J.; Ascencio-Montiel, I.d.J.; Rodríguez-Arellano, M.E. Genome-Wide Association Study of Body Mass Index and Body Fat in Mexican-Mestizo Children. Genes 2019, 10, 945. https://doi.org/10.3390/genes10110945
Costa-Urrutia P, Colistro V, Jiménez-Osorio AS, Cárdenas-Hernández H, Solares-Tlapechco J, Ramirez-Alcántara M, Granados J, Ascencio-Montiel IdJ, Rodríguez-Arellano ME. Genome-Wide Association Study of Body Mass Index and Body Fat in Mexican-Mestizo Children. Genes. 2019; 10(11):945. https://doi.org/10.3390/genes10110945
Chicago/Turabian StyleCosta-Urrutia, Paula, Valentina Colistro, Angélica Saraí Jiménez-Osorio, Helios Cárdenas-Hernández, Jacqueline Solares-Tlapechco, Miryam Ramirez-Alcántara, Julio Granados, Iván de Jesús Ascencio-Montiel, and Martha Eunice Rodríguez-Arellano. 2019. "Genome-Wide Association Study of Body Mass Index and Body Fat in Mexican-Mestizo Children" Genes 10, no. 11: 945. https://doi.org/10.3390/genes10110945
APA StyleCosta-Urrutia, P., Colistro, V., Jiménez-Osorio, A. S., Cárdenas-Hernández, H., Solares-Tlapechco, J., Ramirez-Alcántara, M., Granados, J., Ascencio-Montiel, I. d. J., & Rodríguez-Arellano, M. E. (2019). Genome-Wide Association Study of Body Mass Index and Body Fat in Mexican-Mestizo Children. Genes, 10(11), 945. https://doi.org/10.3390/genes10110945