Tomato Natural Resistance Genes in Controlling the Root-Knot Nematode

 ,
,  ,
,  ,
,
Abstract
1. Introduction
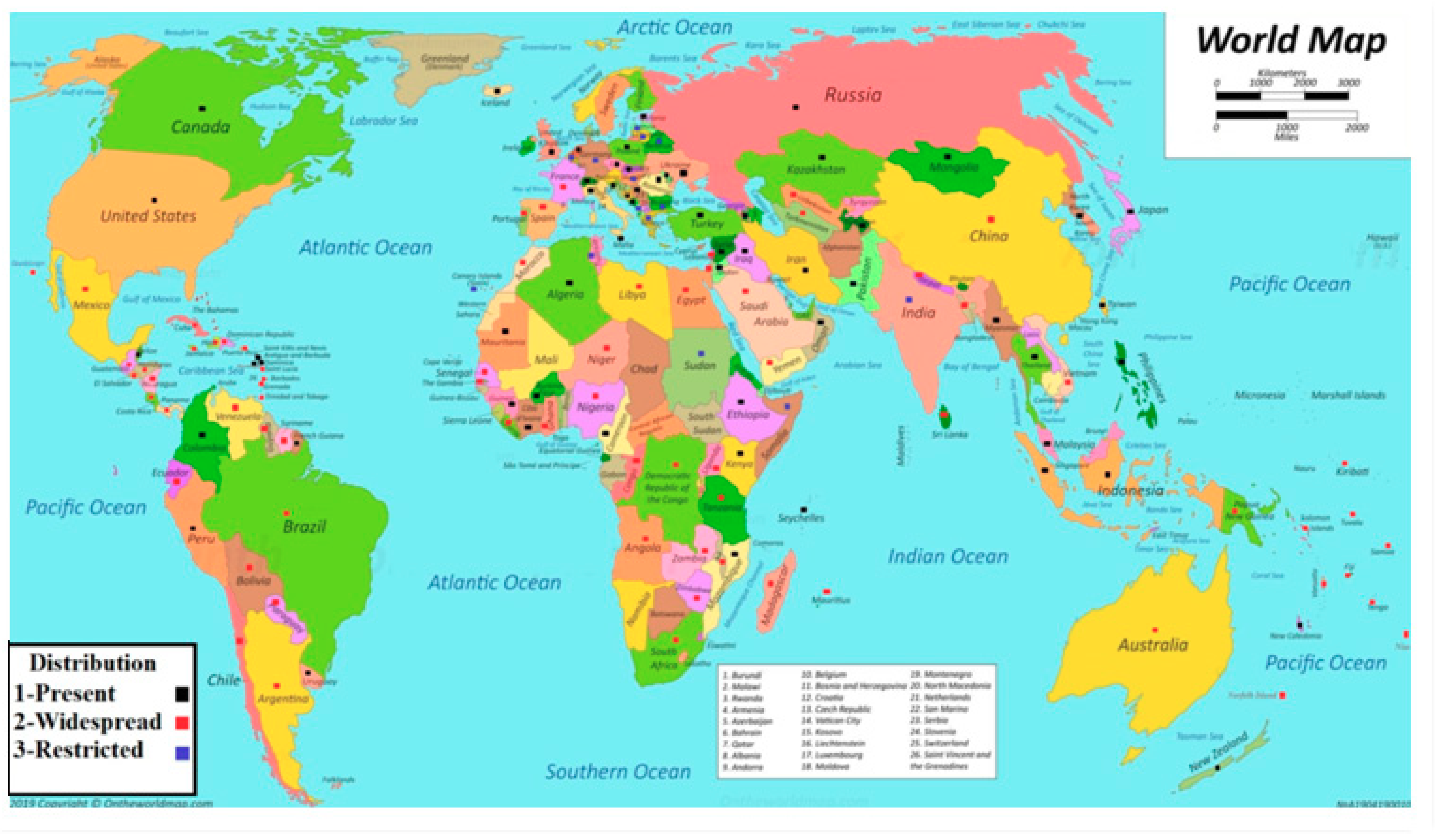
2. Distribution of Meloidogyne Species around the World
3. Naturally Resistant Resources
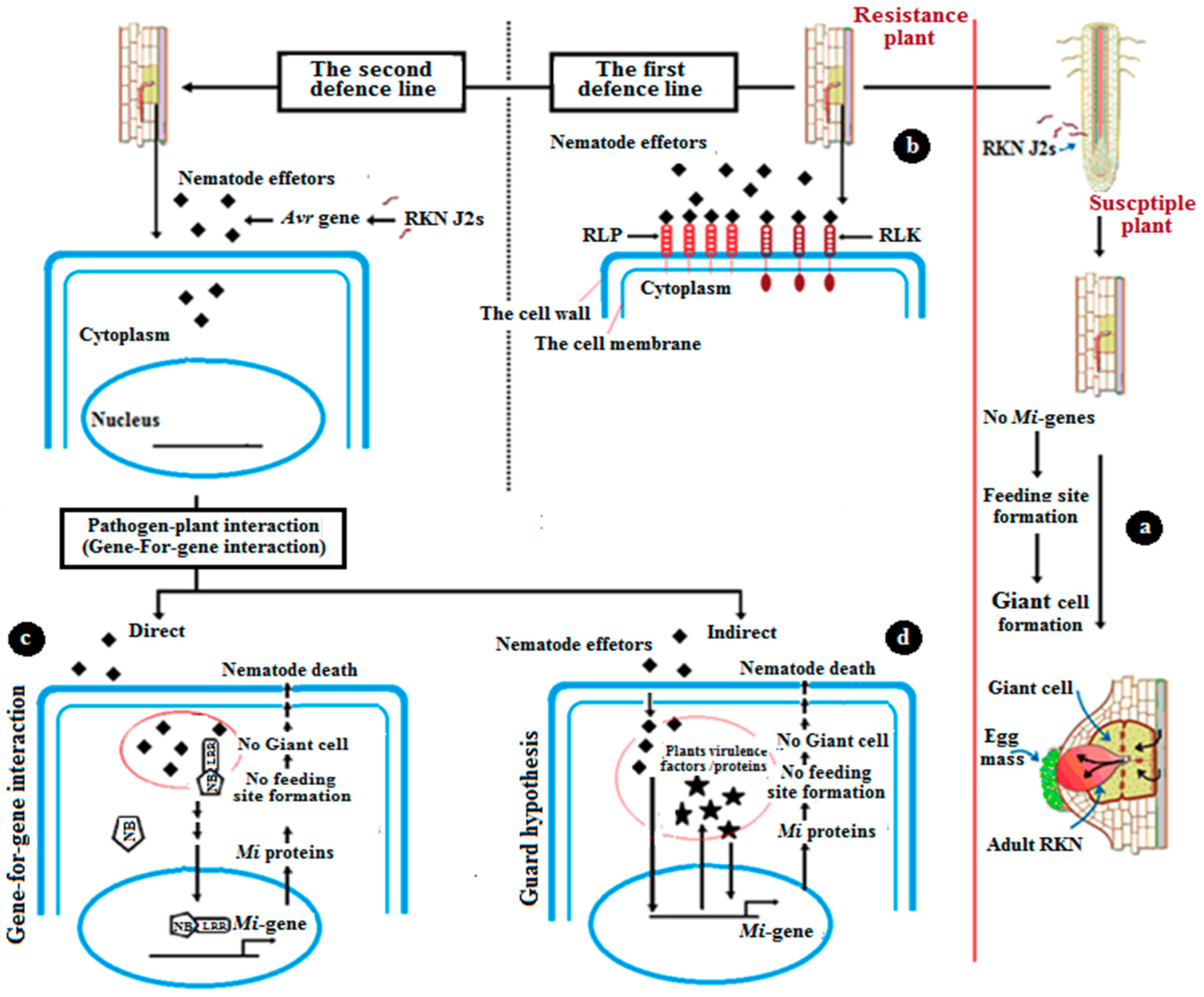
4. The Mechanism of Natural Resistance
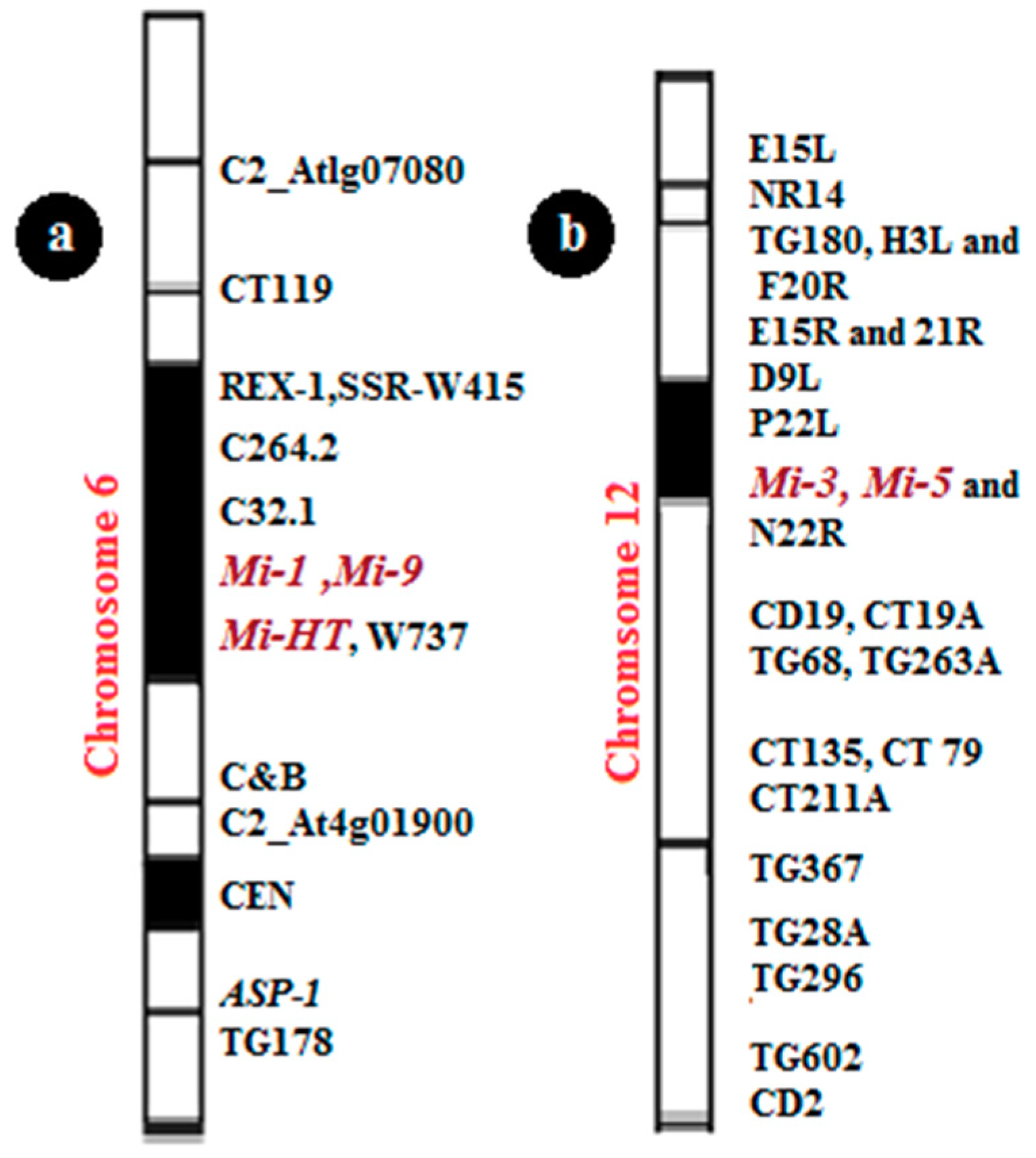
5. The Genetics of Natural Resistance
5.1. Meloidogyne Incognita (Mi) Genes
5.1.1. Mi (Mi-1)
5.1.2. Mi-2
5.1.3. Mi-3
5.1.4. Mi-4
5.1.5. Mi-5
5.1.6. Mi-6
5.1.7. Mi-7
5.1.8. Mi-8
5.1.9. Mi-9
5.1.10. Mi-HT
5.2. Elements Mediating R-genes
6. Problems of Natural Resistance
6.1. Not Effective against All Types of Nematode
6.2. Varieties of Resistance Breakdown
6.3. Genetics of Virulence in Nematodes
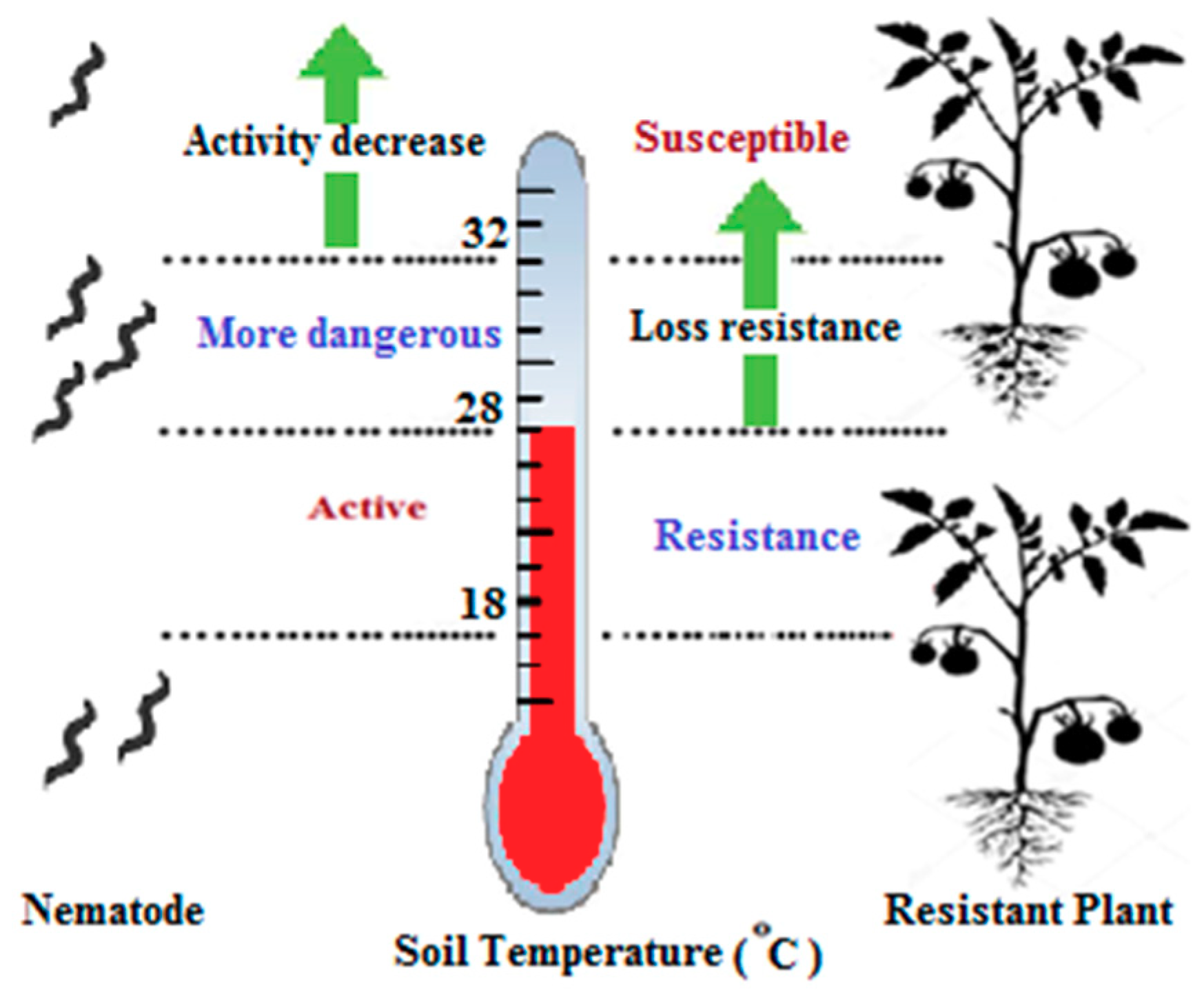
6.4. The Temperature Factor
7. Different Approaches to Strengthening Natural Resistance
7.1. Marker-Assisted Selectionin Breeding Programs
7.2. Genetic Engineering in Controlling RKN
7.2.1. Transfer Resistance Genes
7.2.2. Resistance Effectors
7.2.3. Gene Silencing
Author Contributions
Funding
Conflicts of Interest
References
- Giovannucci, E. Tomatoes, tomato-based products, lycopene, and cancer: A review of the epidemiologic literature. J. Natl. Cancer 1999, 91, 317–331. [Google Scholar] [CrossRef]
- Neal, J.C. The Root-Knot Disease of the Peach, Orange, and Other Plants in Florida due to the Work of Anguillula; U.S. Dept. of Agriculture, Division of Entomology: Washington, DC, USA, 1889.
- Seebold, K.W. Root-Knot Nematode in Commercial & Residential Crops; Plant Pathology Fact Sheet; Plant Pathology Extension; College of Agriculture, University of Kentucky: Lexington, KY, USA, 2014; PPFS-GEN-10. [Google Scholar]
- Trudgill, D.L.; Blok, V.C. Apomictic, polyphagous root knot nematode: Exceptionally successful and damaging biotrophic root pathogens. Annu. Rev. Phytopathol. 2001, 39, 53–77. [Google Scholar] [CrossRef] [PubMed]
- Bailey, D.M. The seedling test method for root-knot nematode resistance. Proc. Am. Soc. Hortic. Sci. 1941, 38, 573–575. [Google Scholar]
- Smith, P.G. Embryo culture of a tomato species hybrid. Proc. Am. Soc. Hortic. Sci. 1944, 44, 413–416. [Google Scholar]
- Ho, J.-Y.; Weide, R.; Ma, H.M.; Wordragen, M.F.; Lambert, K.N.; Koornneef, M.; Zabel, P.; Williamson, V.M. The root-knot nematode resistance gene (Mi) in tomato: Construction of a molecular linkage map and identification of dominant cDNA markers in resistant genotypes. Plant J. 1992, 2, 971–982. [Google Scholar] [PubMed]
- Cap, G.B.; Roberts, P.; Thomason, I.J. Inheritance of heat-stable resistance to Meloidogyne incognita in Lycopersicon peruvianum and its relationship to the Mi gene. Theor. Appl. Genet. 1993, 85, 777–783. [Google Scholar] [CrossRef]
- Martinez de Ilarduya, O.; Moore, A.E.; Kaloshian, I. The tomato Rme1 locus is required for Mi-1-mediated resistance to root-knot nematodes and the potato aphid. Plant J. 2001, 27, 417–425. [Google Scholar] [CrossRef]
- Veremis, J.C.; Roberts, P.A. Relationship between Meloidogyne incognita resistance gene in Lycopersicon peruvianum differentiated by heat sensitivity and nematode virulence. Theor. Appl. Genet. 1996, 93, 950–959. [Google Scholar] [CrossRef]
- Wu, W.-W.; Shen, H.-L.; Yang, W.-C. Sources for Heat-Stable Resistance to Southern Root-Knot Nematode (Meloidogyne incognita) in Solanum lycopersicum. Agric. Sci. China 2009, 8, 697–702. [Google Scholar] [CrossRef]
- Yaghoobi, J.; Yates, J.L.; Williamson, V.M. Fine mapping of the nematode resistance gene Mi-3 in Solanum peruvianum and construction of an S. lycopersicum DNA contig spanning the locus. Mol. Genet. Genom. 2005, 274, 60–69. [Google Scholar] [CrossRef]
- Jablonska, B.; Ammiraju, J.S.; Bhattarai, K.K.; Mantelin, S.; Martinez de Ilarduya, O.; Roberts, P.A.; Kaloshian, I. The Mi-9 gene from Solanum arcanum conferring heat-stable resistance to root-knot nematodes is a homolog of Mi-1. Plant Physiol. 2007, 143, 1044–1054. [Google Scholar] [CrossRef] [PubMed]
- Ammiraju, J.S.S.; Veremis, J.C.; Huang, X.; Roberts, P.A.; Kaloshian, I. The heat-stable, root-knot nematode-resistance gene Mi-9 from Lycopersicon peruvianum is localized on the short arm of chromosome 6. Theor. Appl. Genet. 2003, 106, 478–484. [Google Scholar] [CrossRef] [PubMed]
- CABI/EPPO. Meloidogyne Incognita; Distribution Maps of Plant Diseases, No. 854; CAB International: Wallingford, UK, 2002. [Google Scholar]
- Olsen, M.W. Root-Knot Nematode; University of Arizona, Arizona Cooperative Extension: Tucson, AZ, USA, 2000; pp. 1–3. [Google Scholar]
- Abad, P.; Favery, B.; Rosso, M.N.; Castagnone-Sereno, P. Root-knot nematode parasitism and host response: Molecular basis of a sophisticated interaction. Mol. Plant Pathol. 2003, 4, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Ralmi, N.H.A.A.; Khandaker, M.M.; Nashriyah, M. Occurrence and control of root knot nematode in crops: A review. Aust. J. Crop Sci. 2016, 10, 1649–1654. [Google Scholar] [CrossRef]
- Jones, J.T.; Haegemen, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Palomares-Rius, J.E.; Wesemael, W.M.L.; Perry, R.N. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef]
- Yaghoobi, J.; Kaloshian, I.; Wen, Y.; Williamson, V.M. Mapping a new nematode resistance locus in Lycopersicon peruvianum. Theor. Appl. Genet. 1995, 91, 457–464. [Google Scholar] [CrossRef]
- Seah, S.; Yaghoobi, J.; Rossi, M.; Gleason, C.A.; Williamson, V.M. The nematode-resistance gene, Mi-1, is associated with an inverted chromosomal segment in susceptible compared to resistant tomato. Theor. Appl. Genet. 2004, 108, 1635–1642. [Google Scholar] [CrossRef]
- Desaeger, J.A.; Csinos, A.S. Root-knot nematode management in double-cropped plasticulture vegetables. J. Nematol. 2006, 38, 59–67. [Google Scholar]
- Rani, C.I.; Muthuvel, I.; Veeraragavathatham, D. Evaluation of 14 tomato genotypes for yield and root knot nematode resistance parameters. Pest Technol. 2009, 3, 76–80. [Google Scholar]
- Devran, Z.; Söğüt, M.A. The occurrence of virulent root-knot nematode populations on tomatoes bearing the Mi gene in protected vegetable-growing areas of Turkey. Phytoparasitica 2010, 38, 245–251. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, W.; Zhang, W.; Han, Q.; Feng, M.; Shen, H. Mapping of a heat-stable gene for resistance to southern root-knot nematode in Solanum lycopersicum. Plant Mol. Biol. Rep. 2013, 31, 352–362. [Google Scholar] [CrossRef]
- Milligan, S.B.; Bodeau, J.; Yaghoobi, J.; Kaloshian, I.; Zabel, P.; Williamson, V.M. The root knot nematode resistance gene Mi from tomato is a member of the leucine zipper, nucleotide binding, leucine-rich repeat family of plant genes. Plant Cell 1998, 10, 1307–1319. [Google Scholar] [CrossRef]
- Jaiteh, F.; Kwoseh, C.; Akromah, R. Evaluation of tomato genotypes for resistance to root-knot nematodes. Afr. Crop Sci. J. 2012, 20, 41–49. [Google Scholar]
- Okorley, B.A.; Agyeman, C.; Amissah, N.; Nyaku, S.T. Screening Selected Solanum Plants as Potential Rootstocks for the Management of Root-Knot Nematodes (Meloidogyne incognita). Int. J. Agron. 2018. [Google Scholar] [CrossRef]
- Rashed, M.H.; Al-Marmum, M.H.; Uddin, M.N. How Durable is root knot nematode resistance in tomato? Plant Breed. Biotechnol. 2017, 5, 143–162. [Google Scholar] [CrossRef]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef]
- Caplan, J.L.; Zhu, X.; Mamillapalli, P.; Marathe, R.; Anandalakshmi, R.; Dinesh-Kumar, S.P. Induced ER chaperones regulate a receptor-like kinase to mediate antiviral innate immune response in plants. Cell Host Microbe 2009, 6, 457–469. [Google Scholar] [CrossRef]
- Flor, H.H. Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Bakker, E.G.; Toomajian, C.; Kreitman, M.; Bergelson, J. A genome-wide survey of R gene polymorphisms in Arabidopsis. Plant Cell 2006, 18, 1803–1818. [Google Scholar] [CrossRef]
- Heath, M.C. Hypersensitive response-related death. Plant Mol. Biol. 2000, 44, 321–334. [Google Scholar] [CrossRef]
- Dangl, J.L.; Jones, J.D. Plant pathogens and integrated defence responses to infection. Nature 2001, 411, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Dropkin, V.H. Cellular responses of plants to nematode infections. Ann. Rev. Phytopathol. 1969, 7, 101–122. [Google Scholar] [CrossRef]
- Roberts, P.; May, D. Meloidogyne incognita resistance characteristics in tomato genotypes developed for processing. J. Nematol. 1986, 18, 353–358. [Google Scholar]
- Dropkin, V. The necrotic reaction of tomatoes and other hosts resistant to Meloidogyne: Reversal by temperature. Phytopathology 1969, 59, 1632–1637. [Google Scholar]
- Williamson, V.M.; Hwang, C.F.; Truesdell, G.; Bhakta, A.V.; Fort, K.P. The nematode resistance gene, Mi. In Biology of Plant Microbe Interactions; De Wit, P.J.G., Bisseling, T., Stiekema, W.J., Eds.; International Society for Molecular Plant Microbe Interactions: St. Paul, MN, USA, 2000; Volume 2, pp. 88–92. [Google Scholar]
- Seah, S.; Williamson, V.M.; Garcia, B.E.; Mejia, L.; Salus, M.S.; Martin, C.T.; Maxwell, D.P. Evaluation of a co-dominant SCAR marker for detection of the Mi-1 locus for resistance to root-knot nematode in tomato germplasm. Rep. Tomato Genet. Coop. 2007, 57, 37–40. [Google Scholar]
- Messeguer, R.; Ganal, M.; Devicente, M.C.; Young, N.D.; Bolkan, H.; Tanksley, S.D. High-resolution RFLP map around the root-knot nematode resistance gene (Mi) in tomato. Theor. Appl. Genet. 1991, 82, 529–536. [Google Scholar] [CrossRef]
- Van Wordragen, M.F.; Weide, R.; Liharska, T.; Van Der Steen, A.; Koornneef, M.; Zabel, P. Genetic and molecular organization of the short arm and pericentromeric region of tomato chromosome 6. Euphytica 1994, 79, 169–174. [Google Scholar] [CrossRef]
- Kaloshian, I.; Yaghoobi, J.; Liharska, T.; Hontelez, J.; Hanson, D.; Hogan, P.; Jesse, T.; Wijbrandi, J.; Simons, G.; Vos, P.; et al. Genetic and physical localization of the root-knot nematode resistance locus Mi in tomato. Mol. Gen. Genet. 1998, 257, 376–385. [Google Scholar] [CrossRef]
- Rossi, M.; Goggin, F.L.; Milligan, S.B.; Kaloshian, I.; Ullman, D.E.; Williamson, V.M. The nematode resistance gene Mi of tomato confers resistance against the potato aphid. Proc. Natl. Acad. Sci. USA 1998, 95, 9750–9754. [Google Scholar] [CrossRef]
- Goggin, F.L.; Williamson, V.M.; Ullman, D.E. Variability in the response of Macrosiphum euphorbiae and Myzus persicae (Hemiptera: Aphididae) to the tomato resistance gene Mi. Environ. Entomol. 2001, 30, 101–106. [Google Scholar] [CrossRef]
- Ammati, M. Resistance to Meloidogyne in Lycopersicon, Its Stability at High Soil Temperature and Use of Tissue Culture Techniques to Transfer the Resistance to Tomato. Ph.D. Thesis, University of California, Riverside, Oakland, CA, USA, 1985. [Google Scholar]
- Ammati, M.; Thomason, I.J.; McKinney, H.E. Retention of resistance to Meloidogyne incognita in Lycopersicon genotypes at high soil temperature. J. Nematol. 1986, 18, 491–495. [Google Scholar] [PubMed]
- Roberts, P.; Dalmasso, A.; Cap, G.B.; Castagnone-Sereno, P. Resistance in Lycopersicon peruvianum to isolates of Mi gene compatible Meloidogyne populations. J. Nematol. 1990, 22, 585. [Google Scholar] [PubMed]
- Huang, X.; McGiffen, M.; Kaloshian, I. Reproduction of Mi-virulent Meloidogyne incognita isolates on Lycopersicon spp. J. Nematol. 2004, 36, 69–75. [Google Scholar] [PubMed]
- Doganlar, S.; Frary, A.; Tanksley, S.D. Production of interspecific F-1 hybrids, BC1, BC2 and BC3 populations between Lycopersicon esculentum and two accessions of Lycopersicon peruvianum carrying new root-knot nematode resistance genes. Euphytica 1997, 95, 203–207. [Google Scholar] [CrossRef]
- Moretti, A.; Bongiovanni, M.; Castagnone-Sereno, P.; Caranta, C. Introgression of resistance against Mi1-virulent Meloidogyne spp. from Lycopersicon peruvianum into L. esculentum. Tomato Genet. Coop. Rep. 2002, 52, 21–23. [Google Scholar]
- Roupe van der Voort, J.; Wolters, P.; Folkertsma, R.; van Zandvoort, P.; Vinke, H.; Kanyuka, K.; Bendahmane, A.; Jacobsen, E.; janssen, R.; Bakker, J. Mapping of the cyst nematode resistance locus Gpa2 in potato using a strategy based on co-migrating AFLP markers. Theor. Appl. Genet. 1997, 95, 874–880. [Google Scholar] [CrossRef]
- Stamova, B.S.; Chetelat, R.T. Inheritance and genetic mapping of cucumber mosaic virus resistance introgressed from Lycopersicon chilense into tomato. Theor. Appl. Genet. 2000, 101, 527–537. [Google Scholar] [CrossRef]
- Djian-Caporalino, C.; Pijarowski, L.; Fazari, A.; Samson, M.; Gaveau, L.; O’Byrne, C.; Lefebvre, V.; Caranta, C.; Palloix, A.; Abad, P. High-resolution genetic mapping of pepper (Capsicum annuum L.) resistance loci Me3 and Me4 conferring heat-stable resistance root-knot nematodes (Meloidogyne spp.). Theor. Appl. Genet. 2001, 103, 592–600. [Google Scholar] [CrossRef]
- Grube, R.C.; Radwanski, E.R.; Jahn, M. Comparative genetics of disease resistance within Solanaceae. Genetics 2000, 155, 873–887. [Google Scholar]
- Veremis, J.C. Genetic Characterization of Novel Resistance to Root-Knot Nematodes (Meloidogyne spp.) in Wild Tomato (Lycopersicon peruvianum). Ph.D. Thesis, University of California, Riverside, Oakland, CA, USA, 1995. [Google Scholar]
- Barbary, A.; Djian-Caporalino, C.; Palloix, A.; Castagnone-Sereno, P. Host genetic resistance to root-knot nematodes, Meloidogyne spp., in Solanaceae: From genes to the field. Pest Manag. Sci. 2015, 71, 1591–1598. [Google Scholar] [CrossRef]
- Hwang, C.F.; Bhakta, A.V.; Truesdell, G.M.; Pudlo, W.M.; Williamson, V.M. Evidence for a role of the N terminus and leucine-rich repeat region of the Mi gene product in regulation of localized cell death. Plant Cell 2000, 12, 1319–1329. [Google Scholar] [CrossRef] [PubMed]
- Veremis, J.C.; van Heusden, A.W.; Roberts, P.A. Mapping a novel heat-stable resistance to Meloidogyne in Lycopersicon peruvianum. Theor. Appl. Genet. 1999, 98, 274–280. [Google Scholar] [CrossRef]
- Carter, W.W. Influence of soil temperature on Meloidogyne incognita resistant and susceptible cotton, Gossypium hirsutum. J. Nematol. 1982, 14, 343. [Google Scholar] [PubMed]
- Veremis, J.C.; Roberts, P.A. Identification of resistance to Meloidogyne javanica in the Lycopersicon peruvianum complex. Theor. Appl. Genet. 1996, 93, 894–901. [Google Scholar] [CrossRef]
- Schulze-Lefert, P. Plant immunity: The origami of receptor activation. Curr. Biol. 2004, 14, R22–R24. [Google Scholar] [CrossRef]
- Schaff, J.E.; Nielsen, D.M.; Smith, C.P.; Scholl, E.H.; Bird, D.M. Comprehensive transcriptome profiling in tomato reveals a role for glycosyltransferase in Mi-mediated nematode resistance. Plant Physiol. 2007, 144, 1079–1092. [Google Scholar] [CrossRef]
- Bhattarai, K.K.; Li, Q.; Liu, Y.; Dinesh-Kumar, S.P.; Kaloshian, I. The Mi-1-mediated pest resistance requires Hsp90 and Sgt1. Plant Physiol. 2007, 144, 312–323. [Google Scholar] [CrossRef]
- Azevedo, C.; Sadanandom, A.; Kitagawa, K.; Freialdenhoven, A.; Shirasu, K.; Schulze-Lefert, P. The RAR1 interactor SGT1, an essential component of R gene-triggered disease resistance. Science 2002, 295, 2073–2076. [Google Scholar] [CrossRef]
- Takahashi, A.; Casais, C.; Ichimura, K.; Shirasu, K. HSP90 interacts with RAR1 and SGT1 and is essential for RPS2-mediated disease resistance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 11777–11782. [Google Scholar] [CrossRef]
- Abdelsamad, N.; Regmi, H.; Desaeger, J.; DiGennaro, P. Nicotinamide adenine dinucleotide induced resistance against root-knot nematode Meloidogyne hapla is based on increased tomato basal defense. J. Nematol. 2019, 51, 1–10. [Google Scholar] [CrossRef]
- Chinnapandi, B.; Bucki, P.; Fitoussi, N.; Kolomiets, M.; Borrego, E.; Braun Miyara, S. Tomato SlWRKY3 acts as a positive regulator for resistance against the root-knot nematode Meloidogyne javanica by activating lipids and hormone-mediated defense-signaling pathways. Plant Signal. Behav. 2019, 14, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Branch, C.; Hwang, C.F.; Navarre, D.A.; Williamson, V.M. Salicylic acid is part of the Mi-1-mediated defense response to root-knot nematode in tomato. Mol. Plant–Microbe Interact. 2004, 17, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Kiewnick, S.; Dessimoz, M.; Franck, L. Effects of the Mi-1 and the N root-knot nematode-resistance gene on infection and reproduction of Meloidogyne enterolobii on tomato and pepper cultivars. J. Nematol. 2009, 41, 134–139. [Google Scholar] [PubMed]
- Brito, J.A.; Stanley, J.D.; Kaur, R.; Cetintas, R.; Di Vito, M.; Thies, J.A.; Dickson, D.W. Effects of the Mi-1, N and Tabasco genes on infection and reproduction of Meloidogyne mayaguensis on tomato and pepper genotypes. J. Nematol. 2007, 39, 327–332. [Google Scholar]
- Williamson, V.M. Plant nematode resistance genes. Curr. Opin. Plant Biol. 1999, 2, 327–331. [Google Scholar] [CrossRef]
- Thies, J.A.; Dickson, D.W.; Fery, R.L. Stability of resistance to root-knot nematodes in ‘Charlston Belle’ an ‘Carolina Wonder’ bell pepper in a sub-tropical environment. HortScience 2008, 43, 188–190. [Google Scholar] [CrossRef]
- Cetintas, R.; Kaur, R.; Brito, J.A.; Mendes, M.L.; Nyczepir, A.P.; Dickson, D.W. Pathogenicity and reproductive potential of Meloidogyne mayaguensis and M. floridensis compared to three common Meloidogyne spp. Nematropica 2007, 37, 21–31. [Google Scholar]
- Brito, J.A.; Stanley, J.D.; Mendes, M.L.; Cetintas, R.; Dickson, D.W. Host status of selected cultivated plants to Meloidogyne mayaguensis in Florida. Nematropica 2007, 37, 65–71. [Google Scholar]
- Kaloshian, I.; Williamson, V.M.; Miyao, G.; Lawn, D. Resistance-breaking nematodes identified in California tomatoes. Calif. Agric. 1996, 50, 18–30. [Google Scholar] [CrossRef]
- Philis, J.; Vakis, N. Resistance of tomato varieties to the root-knot nematode Meloidogyne javanica in Cyprus. Nematol. Mediterr. 1977, 5, 39–44. [Google Scholar]
- Tzortzakakis, E.G.; Gowen, S.R. Occurrence of resistance breaking pathotypes of Meloidogyne javanica on tomatoes in Crete, Greece. Fundam. Appl. Nematol. 2005, 19, 283–288. [Google Scholar]
- Castagnone-Sereno, P.; Wajnberg, E.; Bongiovanni, M.; Leroy, F.; Dalmasso, A. Genetic variation in Meloidogyne incognita virulence against the tomato Mi resistance gene: Evidence from isofemale line selection studies. Theor. Appl. Genet. 1994, 88, 749–753. [Google Scholar] [CrossRef] [PubMed]
- Jarquin-Barberena, H.; Dalmasso, A.; de Guiran, G.; Cardin, M. Acquired virulence in the plant parasitic nematode Meloidogyne incognita. I. Biological analysis of the phenomenon. Rev. Nematol. 1991, 14, 261–275. [Google Scholar]
- Eddaoudi, M.; Ammati, M.; Rammah, H. Identification of resistance breaking populations of Meloidogyne on tomatoes in Morocco and their effect on new sources of resistance. Fundam. Appl. Nematol. 1997, 20, 285–289. [Google Scholar]
- Molinari, S.; Miacola, C. Interactions between resistant tomato cvs Meloidogyne spp. in vitro. Nematol. Mediterr. 1997, 25, 63–71. [Google Scholar]
- Ornat, C.; Verdejo-Lucas, S.; Sorribas, F.J. A population of Meloidogyne javanica in Spain virulent to the Mi resistance gene in tomato. Plant Dis. 2001, 85, 271–276. [Google Scholar] [CrossRef]
- Roberts, P.A.; Thomason, I.J. Variability in reproduction of isolates of Meloidogyne incognita and Meloidogyne javanica on resistant tomato genotypes. Plant Dis. 1986, 70, 547–551. [Google Scholar] [CrossRef]
- Ogallo, J.L.; McClure, M.A. Systemic acquired resistance and susceptibility to root-knot nematodes in tomato. Phytopathology 1996, 86498–86501. [Google Scholar] [CrossRef]
- Verdejo-Lucas, S.; Cortada, L.; Sorribas, F.J.; Ornat, C. Selection of virulent isolates of Meloidogyne javanica by repeated cultivation of Mi resistance gene tomato rootstocks under field conditions. Plant Pathol. 2009, 58, 990–998. [Google Scholar] [CrossRef]
- Triantaphyllou, A.C. Genetics of Nematode Parasitism on Plants; Veech, J.A., Dickson, D.W., Eds.; Society of Nematologists, Inc.: Hyattsville, MD, USA, 1987; pp. 354–363. [Google Scholar]
- Ferris, H.; Van Gundy, S.D. Meloidogyne ecology and host interrelationships. In Root-Knot Nematodes (Meloidogyne Species); Systematics, Biology and Control; Lamberti, F., Taylor, C.E., Eds.; Acad. Press: London, UK, 1979; pp. 205–230. [Google Scholar]
- Wang, Y.; Bao, Z.; Zhu, Y.; Hua, J. Analysis of temperature modulation of plant defence against biotrophic microbes. Mol. Plant-Microbe Interact. 2009, 22, 498–506. [Google Scholar] [CrossRef]
- Williamson, V. Root-knot nematode resistance genes in tomato and their potential for future use. Annu. Rev. Phytopathol. 1998, 36, 277–293. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Baki, A.; Hroon, S.A.; Chitwood, D.J. Temperature effects on resistance to Meloidogyne spp. in excised tomato roots. HortScience 1996, 31, 147–149. [Google Scholar] [CrossRef]
- Foolad, M.R. Genome mapping and molecular breeding of tomato. Int. J. Plant Genom. 2007, 10, 1–52. [Google Scholar] [CrossRef] [PubMed]
- Williamson, V.M.; Ho, J.Y.; Wu, F.F.; Miller, N.; Kaloshian, I. A PCR based marker tightly linked to the nematode resistance gene, Mi, in tomato. Theor. Appl. Genet. 1994, 87, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Devran, Z.; Söğüt, M.A. Response of heat-stable tomato genotypes to Mi-1 virulent root-knot nematode populations. Turk. J. Entomol. 2014, 38, 229–238. [Google Scholar] [CrossRef][Green Version]
- El Mehrach, K.; Mejía, L.; Gharsallah-Couchane, S.; Salus, M.S.; Martin, C.T.; Hatimi, A.; Vidavski, F.; Williamson, V.; Maxwell, D.P. PCR-based methods for tagging the Mi-1 locus for resistance to root-knot nematode in begomovirus-resistant tomato germplasm. Acta Hortic. 2005, 695, 263–270. [Google Scholar] [CrossRef]
- Devran, Z.; Söğüt, M.A.; Gözel, U.; Tör, M.; Elekcioglu, I.H. Analysis of genetic variation between populations of Meloidogyne spp. from Turkey. Russ. J. Nematol. 2008, 16, 143–149. [Google Scholar]
- Tai, T.H.; Dahlbeck, D.; Clark, E.T.; Gajiwala, P.; Pasion, R.; Whalen, M.C.; Stall, R.E.; Staskawicz, B.J. Expression of the Bs2 pepper gene confers resistance to bacterial spot disease in tomato. Proc. Natl. Acad. Sci. USA 1999, 96, 14153–14158. [Google Scholar] [CrossRef]
- Van der Vossen, E.A.G.; van der Voort, J.; Kanyuka, K.; Bendahmane, A.; Sandbrink, H.; Baulcombe, D.C.; Bakker, J.; Stiekema, W.J.; Klein-Lankhorst, R.M. Homologues of a single resistance-gene cluster in potato confer resistance to distinct pathogens: A virus and a nematode. Plant J. 2000, 23, 567–576. [Google Scholar] [CrossRef]
- Khallouk, S.; Voisin, R.; Ghelder, C.V.; Engler, G.; Amiri, S.; Esmenjaud, D. Histological Mechanisms of the Resistance Conferred by the Ma Gene Against Meloidogyne incognita in Prunus spp. Am. Phytopathol. Soc. Nematol. 2011, 101, 945–951. [Google Scholar] [CrossRef][Green Version]
- Chan, Y.-L.; Yang, A.-H.; Chen, J.-T.; Yeh, K.-W.; Chan, M.-T. Heterologous expression of taro cystatin protects transgenic tomato against Meloidogyne incognita infection by means of interfering sex determination and suppressing gall formation. Plant Cell Rep. 2010, 29, 231–238. [Google Scholar] [CrossRef]
- Goggin, F.L.; Jia, L.; Shah, G.; Hebert, S.; Williamson, V.M.; Ullman, D.E. Heterologous Expression of the Mi-1.2 Gene from Tomato Confers Resistance Against Nematodes but Not Aphids in Eggplant. Mol. Plant-Microbe Interact. 2006, 19, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.-L.; He, Y.; Hsiao, T.-T.; Wang, C.-J.; Tian, Z.; Yeh, K.-W. Pyramiding taro cystatin and fungal chitinase genes driven by a synthetic promoter enhances resistance in tomato to root-knot nematode Meloidogyne incognita. Plant Sci. 2015, 231, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Q.; Wei, J.Z.; Tan, A.; Aroian, R.V. Resistance to root-knot nematode in tomato roots expressing a nematicidal Bacillus thuringiensis crystal protein. Plant Biotechnol. J. 2007, 5, 455–464. [Google Scholar] [CrossRef]
- Ali, M.A.; Azeem, F.; Abbas, A.; Joyia, F.A.; Li, H.; Dababat, A.A. Transgenic Strategies for Enhancement of Nematode Resistance in Plants. Front. Plant Sci. 2017, 8, 750. [Google Scholar] [CrossRef]
- Urwin, P.E.; Lilley, C.J.; Mcpherson, M.J.; Atkinson, H.J. Resistance to both cyst and root-knot nematodes conferred by transgenic Arabidopsis expressing a modified plant cystatin. Plant J. 1997, 12, 455–461. [Google Scholar] [CrossRef]
- Lilley, C.J.; Urwin, P.E.; Johnston, K.A.; Atkinson, H.J. Preferential expression of a plant cystatin at nematode feeding sites confers resistance to Meloidogyne incognita and Globodera pallida. Plant Biotechnol. J. 2003, 2, 3–12. [Google Scholar] [CrossRef]
- Neveu, C.; Philippe, P.A.; Sereno, C. Molecular cloning and characterization of an intestinal cathepsin L protease from the root-knot nematode Meloidogyne incognita. Physiol. Mol. Plant Pathol. 2003, 63, 159–165. [Google Scholar] [CrossRef]
- Da Rocha Fragoso, R.; Batista, J.A.N.; Neto, O.P.O.; Grossi de sa, M.F. Isolation and characterization of a cDNA encoding a serine proteinase from the root-knot nematodes Meloidogyne incognita. Exp. Parasitol. 2005, 110, 123–133. [Google Scholar] [CrossRef][Green Version]
- Hepher, A.; Atkinson, H.J. Nematode Control with Proteinase Inhibitors. U.S. Patent 5,494,813, 5 March 1992. [Google Scholar]
- Urwin, P.E.; Atkinson, H.J.; Waller, D.A.; Mcpherson, M.J. Engineered Oryzacystatin-I expressed in transgenic hairy roots confers resistance to Globodera pallida. Plant J. 1995, 8, 121–131. [Google Scholar] [CrossRef]
- Urwin, P.E.; Levesley, A.; Mcpherson, M.J.; Atkinson, H.J. Transgenic resistance to the nematode Rotylenchulus reniformis conferred by Arabidopsis thaliana plants expressing proteinase inhibitors. Mol. Breed. 2000, 6, 257–264. [Google Scholar] [CrossRef]
- Urwin, P.E.; Green, J.; Atkinson, H.J. Expression of a plant cystatin confers partial resistance to Globodera; full resistance is achieved by pyramiding a cystatin with natural resistance. Mol. Breed. 2003, 12, 263–269. [Google Scholar] [CrossRef]
- Vain, P.; Worland, B.; Clarke, M.C.; Richard, G.; Beavis, M.; Liu, H.; Kohli, A.; Leech, M.; Snape, J.; Christou, P.; et al. Expression of an engineered cysteine proteinase inhibitor (Oryzacystatin-I1D86) for nematode resistance in transgenic rice plants. Theor. Appl. Genet. 1998, 96, 266–271. [Google Scholar] [CrossRef]
- Atkinson, H.J.; Grimwood, S.; Johnston, K.; Green, J. Proto type demonstration of transgenic resistance to the nematode Radopholus similis conferred on banana by a cystatin. Transgenic Res. 2004, 13, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Fanellia, E.; Di Vito, M.; Jonesc, J.T.; de Giorgi, C. Analysis of chitin synthase function in a plant parasitic nematode, Meloidogyne artiellia, using RNAi. Gene 2005, 349, 87–95. [Google Scholar] [CrossRef]
- Yadav, B.C.; Veluthambi, K.; Subramaniam, K. Host-generated double stranded RNA induces RNAi in plant-parasitic nematodes and protects the host from infection. Mol. Biochem. Parasitol. 2006, 148, 219–222. [Google Scholar] [CrossRef]
- Huang, G.; Allen, R.; Davis, E.L.; Baum, T.J.; Hussey, R.S. Engineering broad root-knot resistance in transgenic plants by RNAi silencing of a conserved and essential root-knot nematode parasitism gene. Proc. Natl. Acad. Sci. USA 2006, 103, 14302–14306. [Google Scholar] [CrossRef]
- Shingles, J.; Lilley, C.J.; Atkinson, H.J.; Urwin, P.E. Meloidogyne incognita: Molecular and biochemical characterisation of a cathepsin L cysteine proteinase and the effect on parasitism following RNAi. Exp. Parasitol. 2007, 115, 114–120. [Google Scholar] [CrossRef]
- Choudhary, D.; Koulagi, R.; Rohatagi, D.; Kumar, A.; Jain, P.K.; Sirohi, A. Engineering Resistance Against Root-Knot Nematode, Meloidogyne Incognita, by Host Delivered RNAi in Abstracts of International Conference on Plant Biotechnology for food Security: New Frontiers; National Agricultural Science Centre: New Delhi, India, 2012; pp. 21–24. [Google Scholar]
- Niu, J.H.; Jian, H.; Xu, J.; Chen, C.; Guo, Q. RNAi silencing of the Meloidogyne incognita Rpn7 gene reduces nematode parasitic success. Eur. J. Plant Pathol. 2012, 134, 131–144. [Google Scholar] [CrossRef]
- Dutta, T.K.; Papolu, P.K.; Banakar, P.; Choudhary, D.; Sirohi, A.; Rao, U. Tomato transgenic plants expressing hairpin construct of a nematode protease gene conferred enhanced resistance to root-knot nematodes. Front. Microbiol. 2015, 6, 260, PMID:25883594. [Google Scholar] [CrossRef]
- National Institute of Food and Agriculture. Variability, Adaptation and Management of Nematodes Impacting Crop Production and Trade; California University: Oakland, CA, USA, 2019. Available online: https://portal.nifa.usda.gov/web/crisprojectpages/1018832-variability-adaptation-and-management-of-nematodes-impacting-crop-production-and-trade.html (accessed on 7 November 2019).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype/Lines/Cultivars | Mi Genes | Notes | Reference |
|---|---|---|---|
| (L. peruvianum) PI128657 | Mi-1 | High-level resistance and the main source of resistance. | [6] |
| (L. esculentum) VFNT | Mi-1 | Resistance | [20,21] |
| (L. esculentum) Mobile | Mi-1 | Resistance | [21] |
| (L. esculentum) Ontario | Mi-1 | Resistance | [21] |
| Solanum lycopersicum cv. Amelia | Mi-1 | Resistance | [22] |
| (L. esculentum) CLN2026C | Mi-2 | Resistance | [23] |
| (L. esculentum) CLN2026E | Mi-2 | Resistance | [23] |
| (L. esculentum) CLN1464A | Mi-2 | Resistance | [23] |
| (L. peruvianum) 2R2-clone PI270435 | Mi-2 | Heat-stable resistance | [8,24] |
| (L. esculentum) VWP2 | Mi-3 | Heat-stable resistance | [12] |
| (L. peruvianum) 1MH-clone PI126443 | Mi-3 | Heat-stable resistance | [24] |
| (L.peruvianum) Maranon LA1708 | Mi-4 | Heat-stable resistance | [10]. |
| (L. peruvianum) 1MH-clone PI126443 | Mi-5 | Heat-stable resistance | [10] |
| (L.peruvianum) 3MH-clone PI270435 | Mi-6 | Heat-stable resistance | [10] |
| (L.peruvianum) 3MH-clonePI270435 | Mi-7 | Resistance to RKN, including strains virulent on Mi | [10] |
| (L. peruvianum) 2R2-clone PI270435 | Mi-8 | Resistance to RKN, including strains virulent on Mi | [10] |
| (S.Arcanum)\LA2157 | Mi-9 | Heat-stable resistance | [13,14] |
| ZN48 | Mi-HT | Heat-stable resistance | [25] |
| ZN17 | Mi-HT | Heat-stable resistance | [25] |
| LA0385 | Heat-stable resistance | [25] | |
| CastlerockII | Resistance | [26] | |
| Sun6082 | Resistance | [26] | |
| (S. lycopersicum) Tomato Mongal T-11 | High resistance | [27] | |
| (S. lycopersicum L) Samrudhi F1 | Resistance | [28] | |
| (L.esculentum) LE812 | Resistance | [23] |
| No. | MarkerName | Marker | Gene | Sequence | Reference |
|---|---|---|---|---|---|
| 1 | C&B | CAPS | Mi-9 | 5′-TACCCACGCCCCATCAATG-3′ 5′-TGCAAGAGGGTGAATATTGAGTGC-3′ | [14] |
| 2 | Mint-1 | SCAR | Mi-1.1, 1.2, 1.4 and 1.6 | 5′-TTCTCTAGCTAAACTTCAGCC-3′ 5′-TTTTCGTTTTTCCATGATTCTAC-3′ | [13] |
| 3 | TG180 | SCAR | Mi-3 | 5′-ATACTTCTTTRCAGGAACAGCTCA-3′ 5′-ACTTAGTGATCATAAAGTACCA-3′ | [12] |
| 4 | REX-1 | CAPS | Mi-1.2 | 5′-TCGGAGCCTTGGTCTGAATT-3′ 5′-GCCAGAGATGATTCGTGAGA-3′ | [14,93] |
| 5 | JB-1 | CAPS | Mi-1 | 5′-AACCATTATCCGGTTCACTC-3′ 5′-TTTCCATTCCTTGTTTCTCTG-3′ | [94] |
| 6 | PMi12 | SCAR | Mi-1 | 5′-CCTGCTCGTTTACCATTACTTTTCCAACC-3′ 5′-CTGCTCGTTTACCATTACTTTTCCAACC-3′ | [95] |
| 7 | Mi23 | SCAR | Mi-1.2 | 5′-TGGAAAAATGTTGAATTTCTTTTG-3′ 5′-GCATACTATATGGCTTTTTACCC-3′ | [40] |
| 8 | APS-1 | CAPS | Mi | 5′-GGATTTTCGTGTTCTTGGTG-3′ 5′-GCCCAGTCAGCAAGAAAACT-3′ | [14] |
| 9 | CT119 | CAPS | Mi | 5′-TCAGGTATCGAACCAAAACC-3′ 5′-TAAAAGGTTCATCCTAATAC-3′ | [14] |
| 10 | C1/2 | RAPD | Mi1.1 | 5′-CAGTGAAGTGGAAGTGATGA-3′ | [20,96] |
| 11 | C2S4 | RAPD | Mi1.2 | 5′-CTAAGAGGAATCTCATCACAGG-3′ | [20,96] |
| 12 | TG-263 | SCAR | Mi-3 | 5′-GCTGAGAAATAAAGCTCTTGAGG-3′ 5′-TACCCTTAATGCTTCGGCAGTGG-3′ | [12] |
| 13 | Cf-2 | CAPS | Mi-1.1, 1.2 and 1.3 | 5′-CTAGGCAGCGATTTCCATTT-3′ 5′-CGGAATAGGTAATGGCCTTC-3′ | [13] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Sappah, A.H.; M. M., I.; H. El-awady, H.; Yan, S.; Qi, S.; Liu, J.; Cheng, G.-t.; Liang, Y. Tomato Natural Resistance Genes in Controlling the Root-Knot Nematode. Genes 2019, 10, 925. https://doi.org/10.3390/genes10110925
El-Sappah AH, M. M. I, H. El-awady H, Yan S, Qi S, Liu J, Cheng G-t, Liang Y. Tomato Natural Resistance Genes in Controlling the Root-Knot Nematode. Genes. 2019; 10(11):925. https://doi.org/10.3390/genes10110925
Chicago/Turabian StyleEl-Sappah, Ahmed H., Islam M. M., Hamada H. El-awady, Shi Yan, Shiming Qi, Jingyi Liu, Guo-ting Cheng, and Yan Liang. 2019. "Tomato Natural Resistance Genes in Controlling the Root-Knot Nematode" Genes 10, no. 11: 925. https://doi.org/10.3390/genes10110925
APA StyleEl-Sappah, A. H., M. M., I., H. El-awady, H., Yan, S., Qi, S., Liu, J., Cheng, G.-t., & Liang, Y. (2019). Tomato Natural Resistance Genes in Controlling the Root-Knot Nematode. Genes, 10(11), 925. https://doi.org/10.3390/genes10110925