Transcription Factor CEBPB Inhibits the Expression of the Human HTR1A by Binding to 5′ Regulatory Region In Vitro

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioinformatics Analysis and Prediction of Transcription Factors
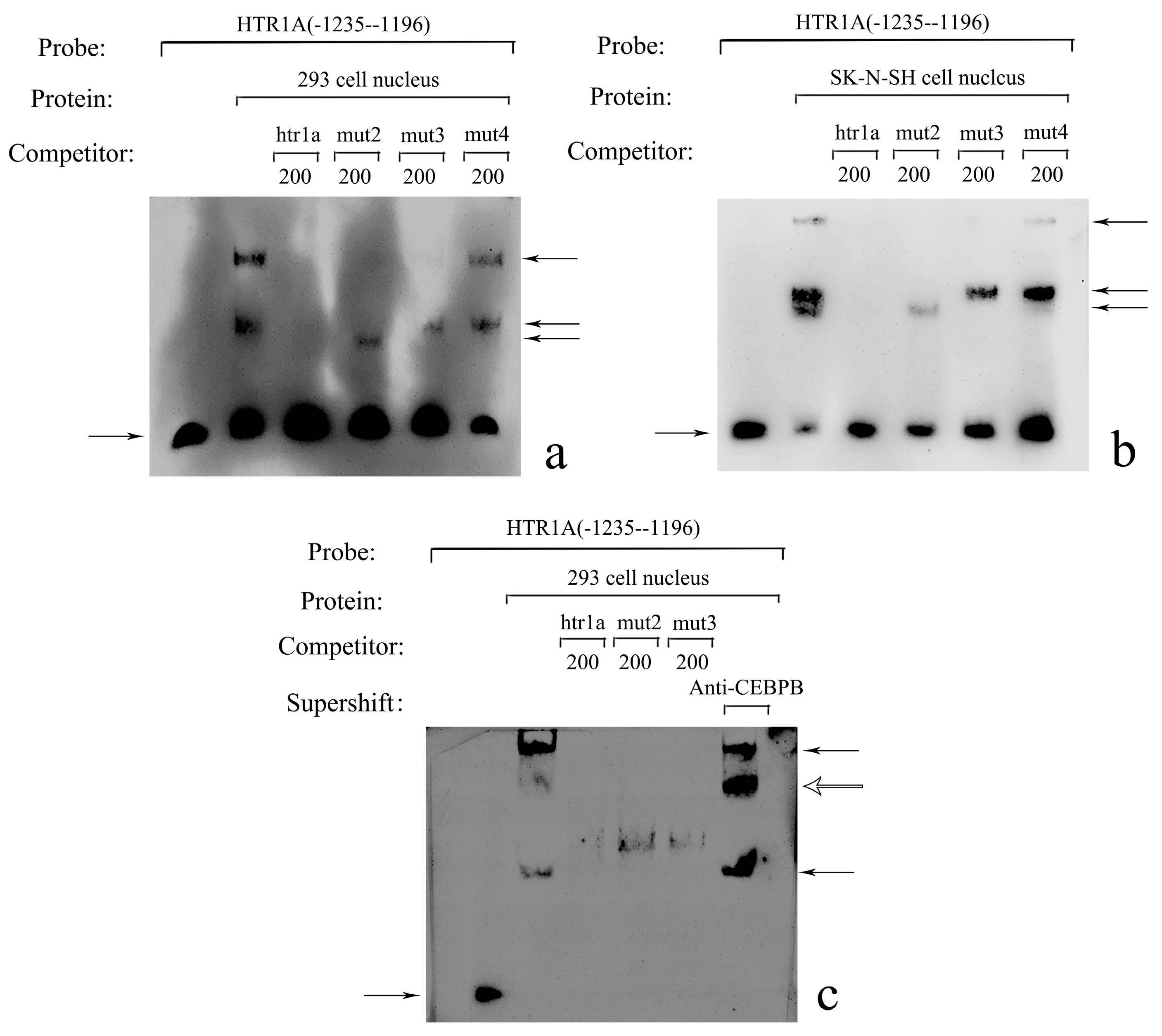
2.2. Electrophoretic Mobility Shift Assay (EMSA)
2.3. Construction of pGL3 Recombinant Vector
2.4. Cell Culture
2.5. Transient Transfection of pGL3-HTR1A-1235, pGL3-HTR1A-1227, pGL3-HTR1A-1208 and pGL3-HTR1A-1196 Recombinant Vectors
2.6. Transient Transfection of pGL3-HTR1A Recombinant Vectors with CEBPB Over-Expressing Vector
2.7. Transfection of CEBPB Over-Expressing Vector
2.8. Dual Luciferase Reporter Assay
2.9. Quantitative Real-Time PCR
2.10. Western Blot
2.11. Statistical Analysis
3. Results
3.1. Prediction and Screening of Transcription Factors
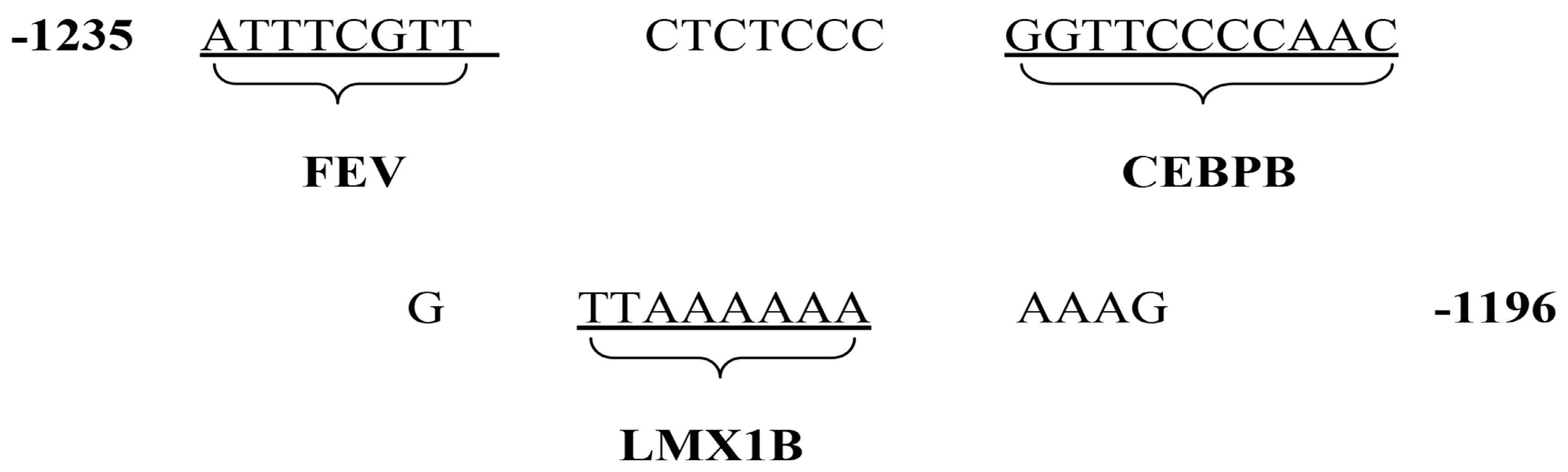
3.2. Sequence Location Identified by CEBPB
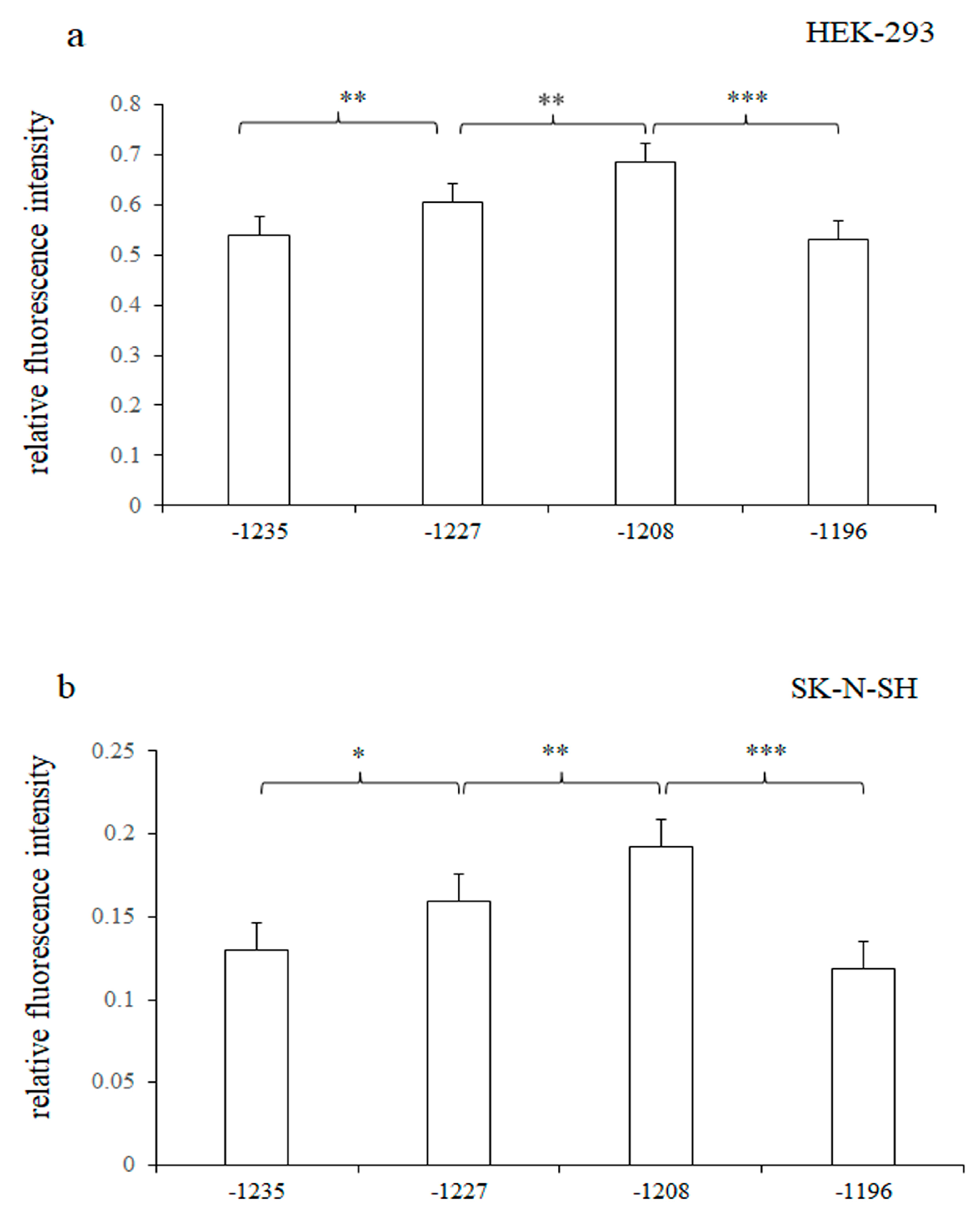
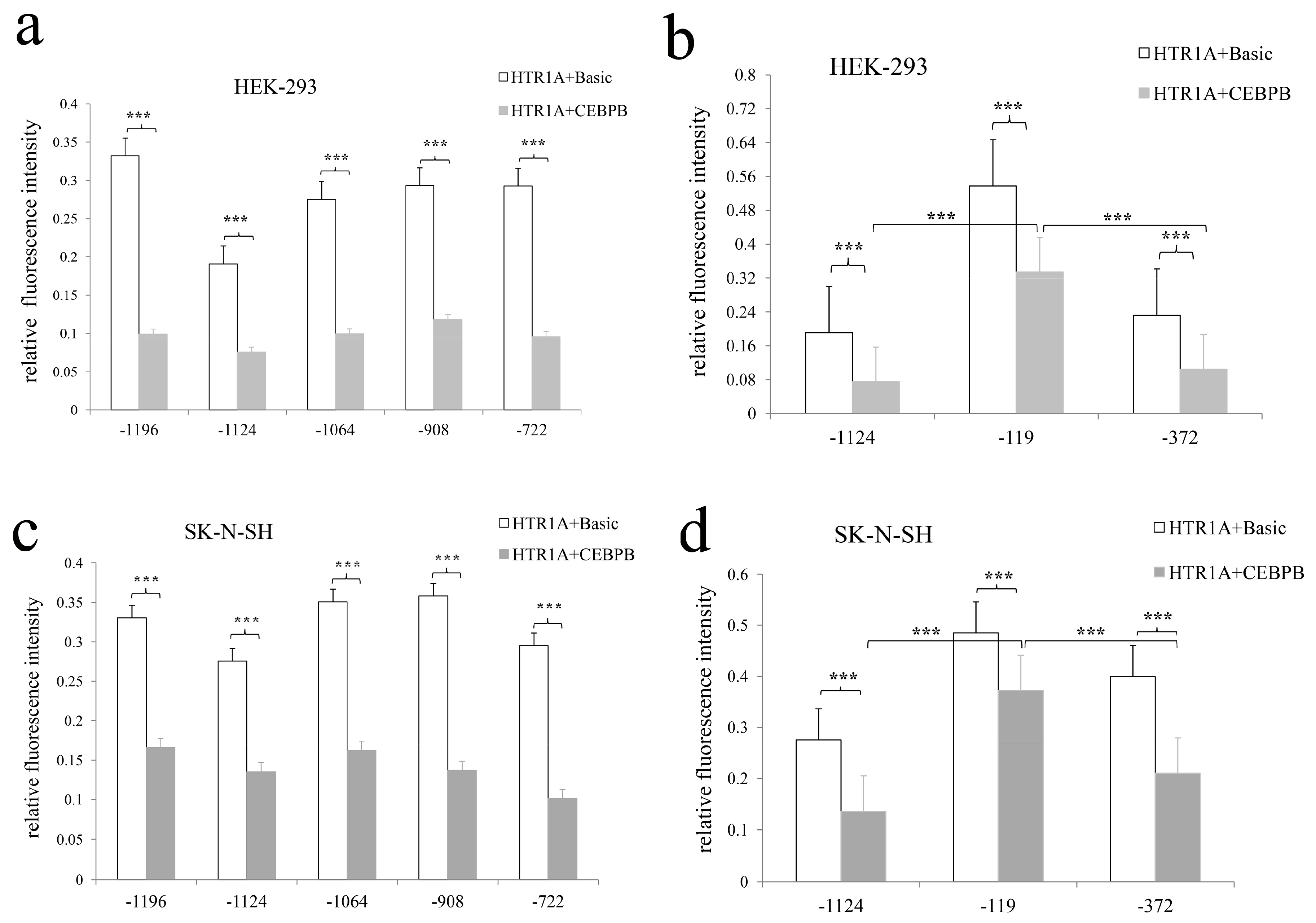
3.3. Significant Differences in Relative Fluorescence Intensity of pGL3-HTR1A-1235, pGL3-HTR1A-1227, pGL3-HTR1A-1208 and pGL3-HTR1A-1196 Recombinant Vectors
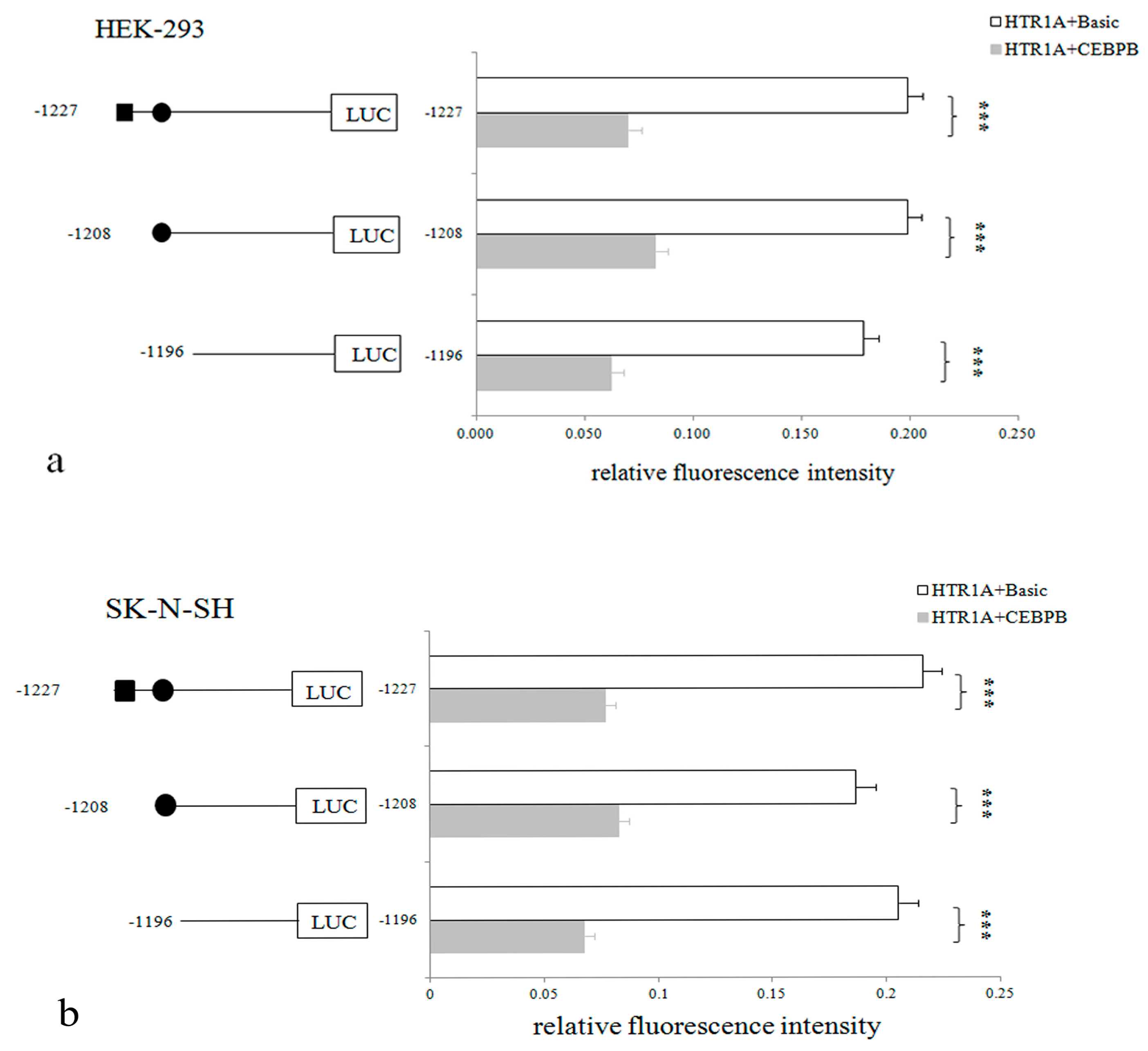
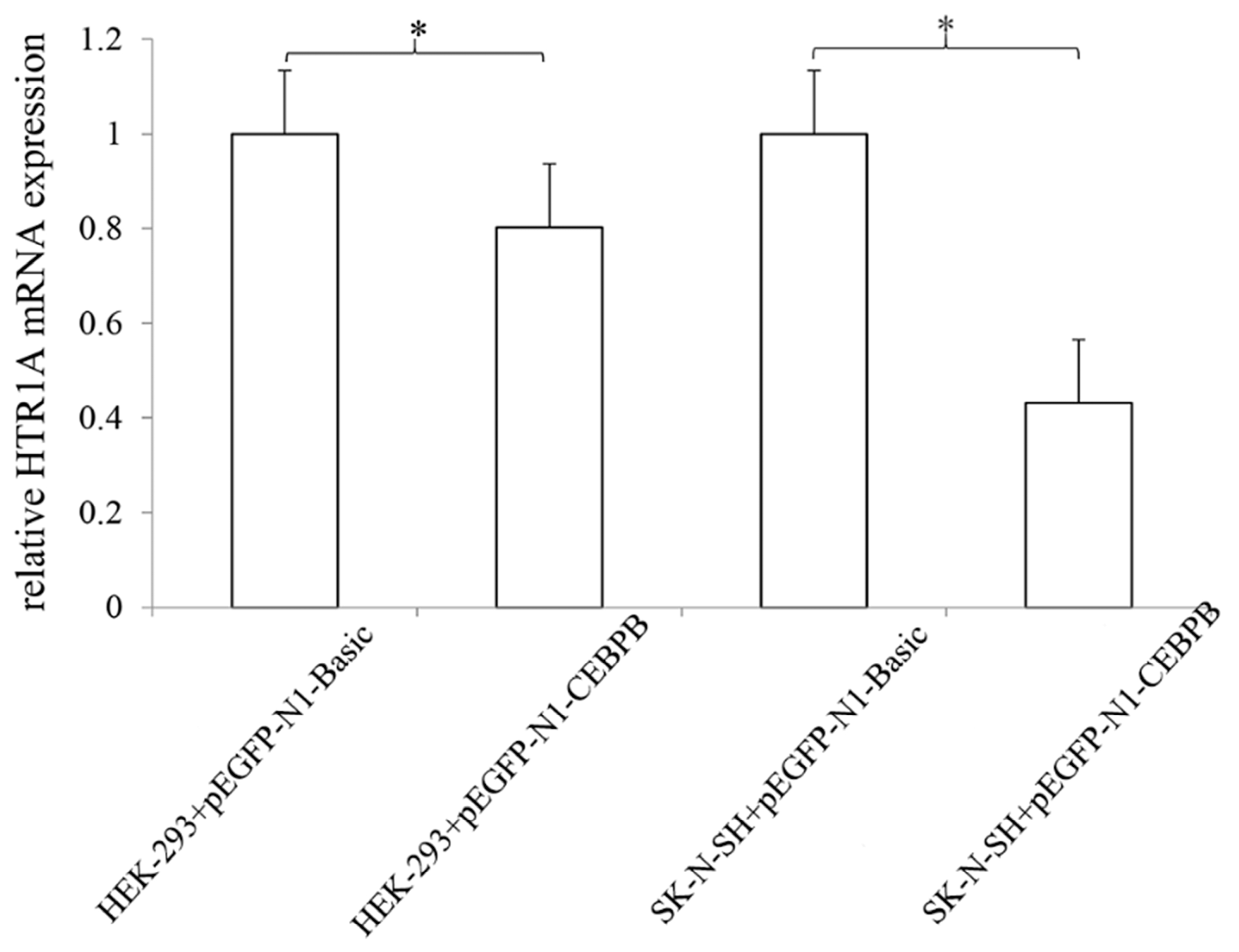
3.4. CEBPB Significantly Inhibited HTR1A Expression
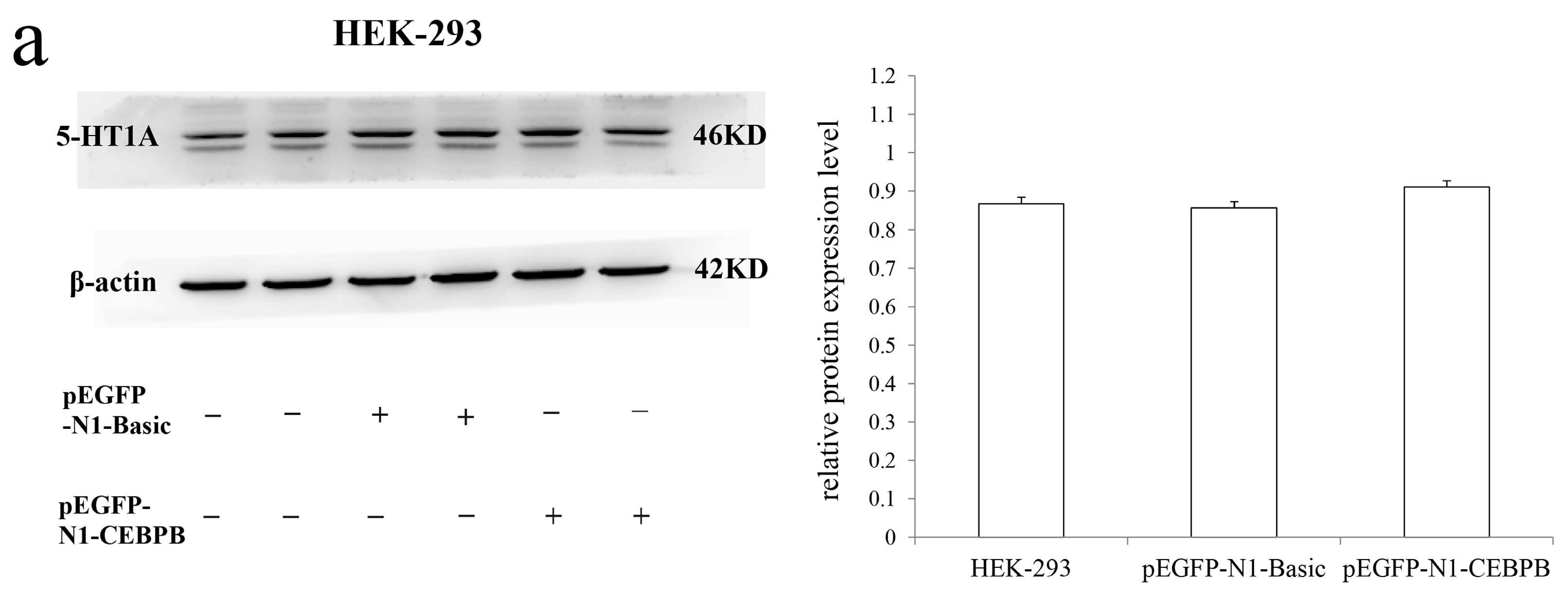
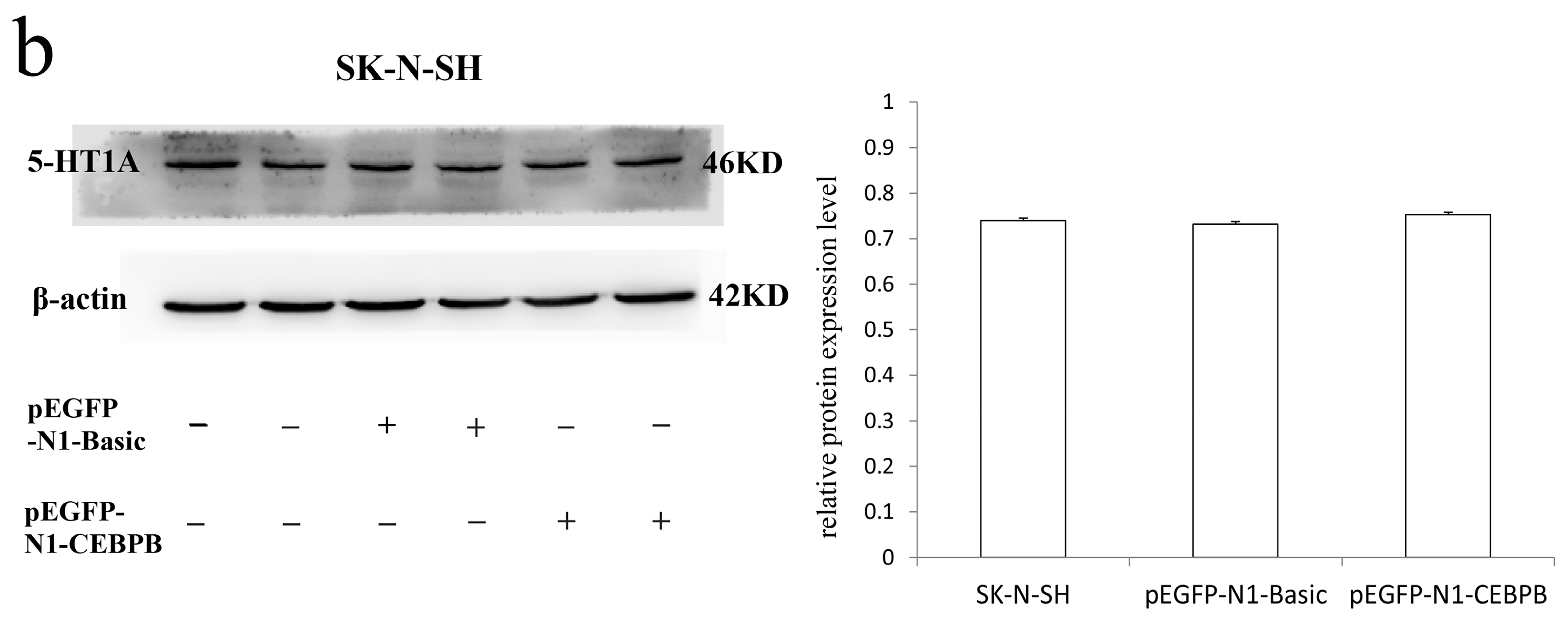
3.5. The Effect of CEBPB on the Endogenous 5-HT1A Receptor Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cervenka, I.; Agudelo, L.Z.; Ruas, J.L. Kynurenines: Tryptophan′s metabolites in exercise, inflammation, and mental health. Science 2017, 357, 369. [Google Scholar] [CrossRef]
- Brezo, J.; Bureau, A.; Merette, C.; Jomphe, V.; Barker, E.D.; Vitaro, F.; Hebert, M.; Carbonneau, R.; Tremblay, R.E.; Turecki, G. Differences and similarities in the serotonergic diathesis for suicide attempts and mood disorders: A 22-year longitudinal gene-environment study. Mol. Psychiatry 2010, 15, 831–843. [Google Scholar] [CrossRef] [PubMed]
- Sadkowski, M.; Dennis, B.; Clayden, R.C.; Elsheikh, W.; Rangarajan, S.; Dejesus, J.; Samaan, Z. The role of the serotonergic system in suicidal behavior. Neuropsychiatr. Dis. Treat. 2013, 9, 1699–1716. [Google Scholar] [PubMed] [Green Version]
- Provini, C.; Everett, J.R.; Pfeffer, C.R. Adults mourning suicide: Self-reported concerns about bereavement, needs for assistance, and help-seeking behavior. Death Stud. 2000, 24, 1–19. [Google Scholar] [PubMed]
- Geyer, M.A.; Vollenweider, F.X. Serotonin research: Contributions to understanding psychoses. Trends Pharmacol. Sci. 2008, 29, 445–453. [Google Scholar] [CrossRef]
- Kishi, T.; Okochi, T.; Tsunoka, T.; Okumura, T.; Kitajima, T.; Kawashima, K.; Yamanouchi, Y.; Kinoshita, Y.; Naitoh, H.; Inada, T.; et al. Serotonin 1A receptor gene, schizophrenia and bipolar disorder: An association study and meta-analysis. Psychiatry Res. 2011, 185, 20–26. [Google Scholar] [CrossRef]
- David, S.P.; Murthy, N.V.; Rabiner, E.A.; Munafo, M.R.; Johnstone, E.C.; Jacob, R.; Walton, R.T.; Grasby, P.M. A functional genetic variation of the serotonin (5-HT) transporter affects 5-HT1A receptor binding in humans. J. Neurosci. 2005, 25, 2586–2590. [Google Scholar] [CrossRef]
- Lemonde, S.; Rogaeva, A.; Albert, P.R. Cell type-dependent recruitment of trichostatin A-sensitive repression of the human 5-HT1A receptor gene. J. Neurochem. 2004, 88, 857–868. [Google Scholar] [CrossRef]
- Lin, H.; Lei, Y.; Zhang, B.; Dai, Z.; Lu, X. Common variants of HTR1A and SLC6A4 confer the increasing risk of Schizophrenia susceptibility: A population-based association and epistasis analysis. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2015, 168, 749–755. [Google Scholar] [CrossRef]
- Ressler, K.J.; Nemeroff, C.B. Role of serotonergic and noradrenergic systems in the pathophysiology of depression and anxiety disorders. Depress. Anxiety 2000, 12, 2–19. [Google Scholar] [CrossRef]
- Milak, M.S.; Pantazatos, S.; Rashid, R.; Zanderigo, F.; DeLorenzo, C.; Hesselgrave, N.; Ogden, R.T.; Oquendo, M.A.; Mulhern, S.T.; Miller, J.M.; et al. Higher 5-HT1A autoreceptor binding as an endophenotype for major depressive disorder identified in high risk offspring—A pilot study. Psychiatry Res. Neuroimaging 2018, 276, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, J.; Sullivan, G.M.; Yang, J.; Ogden, R.T.; Miller, J.M.; Oquendo, M.A.; Mann, J.J.; Parsey, R.V.; DeLorenzo, C. Quantification of the Serotonin 1A Receptor Using PET: Identification of a Potential Biomarker of Major Depression in Males. Neuropsychopharmacology 2015, 40, 1692–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhagwagar, Z.; Cowen, P.J. ‘It’s not over when it’s over’: Persistent neurobiological abnormalities in recovered depressed patients. Psychol. Med. 2008, 38, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Albert, P.R.; Fiori, L.M. Transcriptional dys-regulation in anxiety and major depression: 5-HT1A gene promoter architecture as a therapeutic opportunity. Curr. Pharm. Des. 2014, 20, 3738–3750. [Google Scholar] [CrossRef] [PubMed]
- Cronholm, B.; Asberg, M.; Montgomery, S.; Schalling, D. Suicidal behaviour syndrome with low CSF 5-HIAA. Br. Med. J. 1977, 1, 776. [Google Scholar] [CrossRef] [PubMed]
- Asberg, M. Neurotransmitters and suicidal behavior. The evidence from cerebrospinal fluid studies. Ann. N. Y. Acad. Sci. 1997, 836, 158–181. [Google Scholar] [CrossRef]
- Lesch, K.P.; Araragi, N.; Waider, J.; van den Hove, D.; Gutknecht, L. Targeting brain serotonin synthesis: Insights into neurodevelopmental disorders with long-term outcomes related to negative emotionality, aggression and antisocial behaviour. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 2426–2443. [Google Scholar] [CrossRef]
- Ansorge, M.; Tanneberger, C.; Davies, B.; Theuring, F.; Kusserow, H. Analysis of the murine 5-HT receptor gene promoter in vitro and in vivo. Eur. J. Neurosci. 2004, 20, 363–374. [Google Scholar] [CrossRef]
- Miquel, M.C.; Doucet, E.; Riad, M.; Adrien, J.; Verge, D.; Hamon, M. Effect of the selective lesion of serotoninergic neurons on the regional distribution of 5-HT1A receptor mRNA in the rat brain. Brain Res. Mol. 1992, 14, 357–362. [Google Scholar] [CrossRef]
- Richardson-Jones, J.W.; Craige, C.P.; Guiard, B.P.; Stephen, A.; Metzger, K.L.; Kung, H.F.; Gardier, A.M.; Dranovsky, A.; David, D.J.; Beck, S.G.; et al. 5-HT1A autoreceptor levels determine vulnerability to stress and response to antidepressants. Neuron 2010, 65, 40–52. [Google Scholar] [CrossRef]
- Albert, P.R.; Le Francois, B.; Millar, A.M. Transcriptional dysregulation of 5-HT1A autoreceptors in mental illness. Mol. Brain 2011, 4, 21. [Google Scholar] [CrossRef] [PubMed]
- Stockmeier, C.A.; Shapiro, L.A.; Dilley, G.E.; Kolli, T.N.; Friedman, L.; Rajkowska, G. Increase in serotonin-1A autoreceptors in the midbrain of suicide victims with major depression-postmortem evidence for decreased serotonin activity. J. Neurosci. 1998, 18, 7394–7401. [Google Scholar] [CrossRef] [PubMed]
- Tork, I. Anatomy of the serotonergic system. Ann. N. Y. Acad. Sci. 1990, 600, 9–34. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, D.; Clarke, D.E.; Fozard, J.R.; Hartig, P.R.; Martin, G.R.; Mylecharane, E.J.; Saxena, P.R.; Humphrey, P.P. International Union of Pharmacology classification of receptors for 5-hydroxytryptamine (Serotonin). Pharmacol. Rev. 1994, 46, 157–203. [Google Scholar] [PubMed]
- Arango, V.; Underwood, M.D.; Boldrini, M.; Tamir, H.; Kassir, S.A.; Hsiung, S.; Chen, J.J.; Mann, J.J. Serotonin 1A receptors, serotonin transporter binding and serotonin transporter mRNA expression in the brainstem of depressed suicide victims. Neuropsychopharmacology 2001, 25, 892–903. [Google Scholar] [CrossRef]
- Drevets, W.C.; Frank, E.; Price, J.C.; Kupfer, D.J.; Greer, P.J.; Mathis, C. Serotonin type-1A receptor imaging in depression. Nucl. Med. Biol. 2000, 27, 499–507. [Google Scholar] [CrossRef]
- Parks, C.L.; Shenk, T. The serotonin 1a receptor gene contains a TATA-less promoter that responds to MAZ and Sp1. J. Biol. Chem. 1996, 271, 4417–4430. [Google Scholar] [CrossRef]
- Ou, X.M.; Storring, J.M.; Kushwaha, N.; Albert, P.R. Heterodimerization of mineralocorticoid and glucocorticoid receptors at a novel negative response element of the 5-HT1A receptor gene. J. Biol. Chem. 2001, 276, 14299–14307. [Google Scholar] [CrossRef]
- Meijer, O.C.; de Kloet, E.R. Corticosterone and serotonergic neurotransmission in the hippocampus: Functional implications of central corticosteroid receptor diversity. Crit. Rev. Neurobiol. 1998, 12, 1–20. [Google Scholar] [CrossRef]
- Ou, X.M.; Jafar-Nejad, H.; Storring, J.M.; Meng, J.H.; Lemonde, S.; Albert, P.R. Novel dual repressor elements for neuronal cell-specific transcription of the rat 5-HT1A receptor gene. J. Biol. Chem. 2000, 275, 8161–8168. [Google Scholar] [CrossRef]
- Souslova, T.; Miredin, K.; Millar, A.M.; Albert, P.R. Recruitment by the repressor Freud-1 of histone deacetylase-brg1 chromatin remodeling complexes to strengthen HTR1A gene repression. Mol. Neurobiol. 2017, 54, 8263–8277. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xu, F.L.; Ding, M.; Zhang, J.J.; Yao, J.; Wang, B.J. Characterization and functional analyses of the human HTR1A gene: 5′ regulatory region modulates gene expression in vitro. BMC Genet. 2018, 19, 115. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, K.A.; Steinmann, M.; Magistretti, P.J.; Halfon, O.; Cardinaux, R.J. C/EBPβ couples dopamine signalling to substance P precursor gene expression in striatal neurones. J. Neurochem. 2006, 98, 1390–1399. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Chen, W.; Liu, H.; Yang, H.; Liu, T. Transcription factor CEBPB inhibits the proliferation of osteosarcoma by regulating downstream target gene CLEC5A. J. Clin. Lab. Anal. 2019, 22985. [Google Scholar] [CrossRef] [PubMed]
- Ramji, D.P.; Foka, P. CCAAT/enhancer-binding proteins: Structure, function and regulation. Biochem. J. 2002, 365, 561–575. [Google Scholar] [CrossRef]
- Huber, R.; Panterodt, T.; Welz, B.; Christmann, M.; Friesenhagen, J.; Westphal, A.; Pietsch, D.; Brand, K. C/EBPβ-LAP*/LAP expression is mediated by RSK/eIF4B-dependent signalling and boosted by increased protein stability in models of monocytic differentiation. PLoS ONE 2015, 10, e0144338. [Google Scholar] [CrossRef]
- Adzic, M.; Djordjevic, J.; Mitic, M.; Brkic, Z.; Lukic, I.; Radojcic, M. The contribution of hypothalamic neuroendocrine, neuroplastic and neuroinflammatory processes to lipopolysaccharide-induced depressive-like behaviour in female and male rats: Involvement of glucocorticoid receptor and C/EBP-beta. Behav. Brain Res. 2015, 291, 130–139. [Google Scholar] [CrossRef]
- Patel, D.N.; King, C.A.; Bailey, S.R.; Holt, J.W.; Venkatachalam, K.; Agrawal, A.; Valente, A.J.; Chandrasekar, B. Interleukin-17 stimulates C-reactive protein expression in hepatocytes and smooth muscle cells via p38 MAPK and ERK1/2-dependent NF-kappaB and C/EBPβ activation. J. Biol. Chem. 2007, 282, 27229–27238. [Google Scholar] [CrossRef]
- Simpson-Abelson, M.R.; Childs, E.E.; Ferreira, M.C.; Bishu, S.; Conti, H.R.; Gaffen, S.L. C/EBPβ promotes immunity to oral candidiasis through regulation of β-defensins. PLoS ONE 2015, 10, e0136538. [Google Scholar] [CrossRef]
- Zhou, X.; Mao, Y.; Zhu, J.; Meng, F.; Chen, Q.; Tao, L.; Li, R.; Fu, F.; Liu, C.; Hu, Y.; et al. TGF-β1 promotes colorectal cancer immune escape by elevating B7-H3 and B7-H4 via the miR-155/miR-143 axis. Oncotarget 2016, 7, 67196–67211. [Google Scholar]
- Shim, S.H.; Hwangbo, Y.; Kwon, Y.J.; Jeong, H.Y.; Lee, B.H.; Hwang, J.A.; Kim, Y.K. A case-control association study of serotonin 1A receptor gene and tryptophan hydroxylase 2 gene in attention deficit hyperactivity disorder. Prog. Neuropsychopharmacol. Biol. Psychiatry 2010, 34, 974–979. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Laurence, A.; Kanno, Y.; Pacher-Zavisin, M.; Zhu, B.M.; Tato, C.; Yoshimura, A.; Hennighausen, L.; O’Shea, J.J. Selective regulatory function of Socs3 in the formation of IL-17-secreting T cells. Proc. Natl. Acad. Sci. USA 2006, 103, 8137–8142. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequences |
|---|---|
| HTR1A(−1235–−1196) | 5′-ATTTCGTTCTCTCCCGGTTCCCCAACGTTAAAAAAAAAG-3′(WT) |
| mut2 (CEBPB) | 5′-ATTTCGTTCTCTCCCAACCTTTTGGAGTTAAAAAAAAAG-3′(MT) |
| mut3 (FEV) | 5′-CGGGATCCCTCTCCCGGTTCCCCAACGTTAAAAAAAAAG-3′(MT) |
| mut4(LMX1B) | 5′-ATTTCGTTCTCTCCCGGTTCCCCAACGCCGGGCCGAAAG-3′(MT) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.-P.; Wu, X.; Meng, J.-H.; Ding, M.; Xu, F.-L.; Zhang, J.-J.; Yao, J.; Wang, B.-J. Transcription Factor CEBPB Inhibits the Expression of the Human HTR1A by Binding to 5′ Regulatory Region In Vitro. Genes 2019, 10, 802. https://doi.org/10.3390/genes10100802
Liu Y-P, Wu X, Meng J-H, Ding M, Xu F-L, Zhang J-J, Yao J, Wang B-J. Transcription Factor CEBPB Inhibits the Expression of the Human HTR1A by Binding to 5′ Regulatory Region In Vitro. Genes. 2019; 10(10):802. https://doi.org/10.3390/genes10100802
Chicago/Turabian StyleLiu, Yong-Ping, Xue Wu, Jing-Hua Meng, Mei Ding, Feng-Ling Xu, Jing-Jing Zhang, Jun Yao, and Bao-Jie Wang. 2019. "Transcription Factor CEBPB Inhibits the Expression of the Human HTR1A by Binding to 5′ Regulatory Region In Vitro" Genes 10, no. 10: 802. https://doi.org/10.3390/genes10100802