Association of NLRP1 Coding Polymorphism with Lung Function and Serum IL-1β Concentration in Patients Diagnosed with Chronic Obstructive Pulmonary Disease (COPD)

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Gene/SNP Selection and Genotyping
2.3. Gene Expression Analyses
2.4. Serum IL-1β Concentration
2.5. Statistical Analysis
3. Results
3.1. Association of the NLRP Coding Variants with COPD Risk and Disease Severity
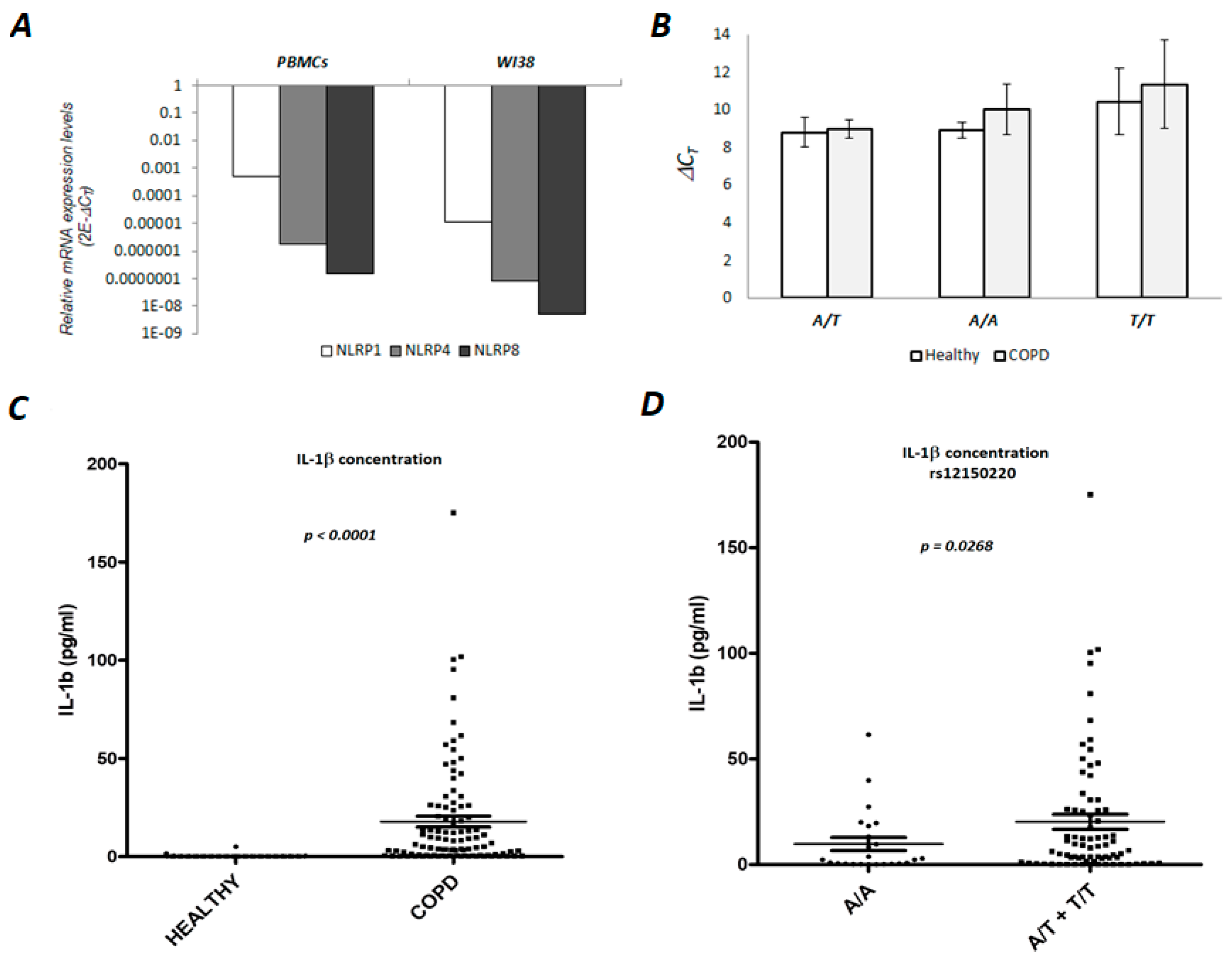
3.2. NLRP1 is Expressed in Human PBMCs and Lung Fibroblasts
3.3. Serum Concentration of the IL-1β in the COPD Patients is Associated with NLRP1 rs12150220 Genotype
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Akinbami, L.J.; Liu, X. American Lung Association. Trends in COPD (Chronic Bronchitis and Emphysema): Morbidity and Mortality. Am. Lung Assoc. Epidemiol. Stat. Unit Res. Health Educ. Div 2013, 1–8. [Google Scholar]
- Vogelmeier, C.F.; Criner, G.J.; Martinez, F.J.; Anzueto, A.; Barnes, P.J.; Bourbeau, J.; Celli, B.R.; Chen, R.; Decramer, M.; Fabbri, L.M.; et al. Global Strategy for the Diagnosis, Management, and Prevention of Chronic Obstructive Lung Disease 2017 Report. GOLD Executive Summary. Am. J. Respir. Crit. Care Med. Am. Thorac. Soc. 2017, 195, 557–582. [Google Scholar] [CrossRef] [PubMed]
- Angelis, N.; Porpodis, K.; Zarogoulidis, P.; Spyratos, D.; Kioumis, I.; Papaiwannou, A.; Pitsiou, G.; Tsakiridis, K.; Mpakas, A.; Arikas, S.; et al. Airway inflammation in chronic obstructive pulmonary disease. J. Thorac. Dis. 2014, 6, S167–S172. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Hammad, H. The role of dendritic and epithelial cells as master regulators of allergic airway inflammation. Lancet 2010, 376, 835–843. [Google Scholar] [CrossRef]
- Brusselle, G.G.; Joos, G.F.; Bracke, K.R. New insights into the immunology of chronic obstructive pulmonary disease. Lancet 2011, 378, 1015–1026. [Google Scholar] [CrossRef]
- Sin, D.D.; Anthonisen, N.R.; Soriano, J.B.; Agusti, A.G. Mortality in COPD: Role of comorbidities. Eur. Respir. J. 2006, 28, 1245–1257. [Google Scholar] [CrossRef]
- Chatila, W.M.; Thomashow, B.M.; Minai, O.A.; Criner, G.J.; Make, B.J. Comorbidities in Chronic Obstructive Pulmonary Disease. Proc. Am. Thorac. Soc. 2008, 5, 549–555. [Google Scholar] [CrossRef]
- Divo, M.; Cote, C.; De Torres, J.P.; Casanova, C.; Marín, J.M.; Pinto-Plata, V.; Zulueta, J.J.; Cabrera, C.; Zagaceta, J.; Hunninghake, G.; et al. Comorbidities and Risk of Mortality in Patients with Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2012, 186, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Brusselle, G.G.; Provoost, S.; Bracke, K.R.; Kuchmiy, A.; Lamkanfi, M. Inflammasomes in Respiratory Disease. Chest 2014, 145, 1121–1133. [Google Scholar] [CrossRef]
- De Vasconcelos, N.M.; Van Opdenbosch, N.; Lamkanfi, M. Inflammasomes as polyvalent cell death platforms. Cell Mol. Life Sci. 2016, 73, 2335–2347. [Google Scholar] [CrossRef]
- Bauernfeind, F.; Hornung, V. Of inflammasomes and pathogens—sensing of microbes by the inflammasome. EMBO Mol. Med. 2013, 5, 814–826. [Google Scholar] [CrossRef] [PubMed]
- Van de Veerdonk, F.L.; Netea, M.G.; Dinarello, C.; Joosten, L.B. Inflammasome activation and IL-1β and IL-18 processing during infection. Trends Immunol. 2011, 32, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Chen, X.; Liu, J.; Zhou, D.B.; Kuang, X.; Xiao, J.; Yu, Q.; Lu, X.; Li, W.; Xie, B.; et al. Serum IL-1β and IL-17 levels in patients with COPD: Associations with clinical parameters. Int. J. Chronic Obstr. Pulm. Dis. 2017, 12, 1247–1254. [Google Scholar] [CrossRef]
- Pauwels, N.S.; Bracke, K.R.; Dupont, L.L.; Van Pottelberge, G.R.; Provoost, S.; Berghe, T.V.; Vandenabeele, P.; Lambrecht, B.N.; Joos, G.F.; Brusselle, G.G. Role of IL-1 and the Nlrp3/caspase-1/IL-1 axis in cigarette smoke-induced pulmonary inflammation and COPD. Eur. Respir. J. 2011, 38, 1019–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, R.Y.; Pinkerton, J.W.; Gibson, P.G.; Cooper, M.A.; Horvat, J.C.; Hansbro, P.M. Inflammasomes in COPD and neutrophilic asthma. Thorax 2015, 70. [Google Scholar] [CrossRef] [PubMed]
- Huhn, S.; Miguel, I.; Filho, S.; Sanmuganantham, T.; Pichulik, T.; Catalano, C.; Pardini, B.; Naccarati, A.; Polakova-Vymetálkova, V.; Jiraskova, K.; et al. Coding variants in NOD-like receptors: An association study on risk and survival of colorectal cancer. PLoS ONE 2018, 5, e0199350. [Google Scholar] [CrossRef]
- Campo, C.; Kohler, A.; Figlioli, G.; Elisei, R.; Romei, C.; Cipollini, M. Inherited variants in genes somatically mutated in thyroid cancer. PLoS ONE 2017, 12, e0174995. [Google Scholar] [CrossRef]
- Singh, D.P.; Bagam, P.; Sahoo, M.K.; Batra, S. Immune-related gene polymorphisms in pulmonary diseases. Toxicology 2017, 383, 24–39. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Li, Y.; Zhu, L.; Liu, D.; Songyang, Z.; Wang, H.Y.; Wang, R.-F. NLRP4 negatively regulates type I interferon signaling by targeting the kinase TBK1 for degradation via the ubiquitin ligase DTX4. Nat. Immunol. 2012, 13, 387–395. [Google Scholar] [CrossRef]
- Jounai, N.; Kobiyama, K.; Shiina, M.; Ogata, K.; Ishii, K.J.; Takeshita, F. NLRP4 Negatively Regulates Autophagic Processes through an Association with Beclin1. J. Immunol. 2011, 186, 1646–1655. [Google Scholar] [CrossRef]
- Tian, X.; Pascal, G.; Monget, P. Evolution and functional divergence of NLRP genes in mammalian reproductive systems. BMC Evol. Boil. 2009, 9, 202. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Verma, S.; Kumar, S.; Ahmad, M.; Nischal, A.; Singh, S.; Dixit, R. Correlation of severity of chronic obstructive pulmonary disease with potential biomarkers. Immunol. Lett. 2018, 196, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Putcha, N.; Puhan, M.A.; Hansel, N.N.; Drummond, M.B.; Boyd, C.M. Impact of co-morbidities on self-rated health in self-reported COPD: An analysis of NHANES 2001-2008. COPD: J. Chronic Obstr. Pulm. Dis. 2013, 10, 324–332. [Google Scholar] [CrossRef]
- Smith, M.C.; Wrobel, J.P. Epidemiology and clinical impact of major comorbidities in patients with COPD. Int. J. Chronic Obstr. Pulm. Dis. 2014, 9, 871–888. [Google Scholar] [CrossRef] [PubMed]
- García-Rio, F.; Soriano, J.B.; Miravitlles, M.; Munoz, L.; Duran-Tauleria, E.; Sánchez, G.; Sobradillo, V.; Ancochea, J. Impact of Obesity on the Clinical Profile of a Population-Based Sample with Chronic Obstructive Pulmonary Disease. PLoS ONE 2014, 9, e105220. [Google Scholar] [CrossRef] [PubMed]
- Chaput, C.; Sander, L.E.; Suttorp, N.; Opitz, B. NOD-Like Receptors in Lung Diseases. Front. Immunol. 2013, 4, 1–12. [Google Scholar] [CrossRef]
- Li, Y.; Wei, X.; Ji, S.; Gui, S.; Zhang, S. In vivo effects of the NLRP1/NLRP3 inflammasome pathway on latent respiratory virus infection. Int. J. Mol. Med. 2018, 41, 3620–3628. [Google Scholar] [CrossRef]
- Crovella, S.; Moura, R.R.; Cappellani, S.; Celsi, F.; Trevisan, E.; Schneider, M.; Brollo, A.; Nicastro, E.M.; Vita, F.; Finotto, L.; et al. A genetic variant of NLRP1 gene is associated with asbestos body burden in patients with malignant pleural mesothelioma. J. Toxicol. Environ. Health Part A 2017, 81, 98–105. [Google Scholar] [CrossRef]
- Leal, V.N.C.; Genov, I.R.; Mallozi, M.C.; Solé, D.; Pontillo, A. Polymorphisms in inflammasome genes and risk of asthma in Brazilian children. Mol. Immunol. 2018, 93, 64–67. [Google Scholar] [CrossRef]
- Pontillo, A.; Girardelli, M.; Kamada, A.J.; Pancotto, J.A.; Donadi, E.A.; Crovella, S.; Sandrin-Garcia, P. Polimorphisms in Inflammasome Genes Are Involved in the Predisposition to Systemic Lupus Erythematosus. Autoimmunity 2012, 45, 271–278. [Google Scholar] [CrossRef]
- Levandowski, C.B.; Mailloux, C.M.; Ferrara, T.M.; Gowan, K.; Ben, S.; Jin, Y.; McFann, K.K.; Holland, P.J.; Fain, P.R.; Dinarello, C.A.; et al. NLRP1 haplotypes associated with vitiligo and autoimmunity increase interleukin-1β processing via the NLRP1 inflammasome. Proc. Natl. Acad. Sci. USA 2013, 110, 2952–2956. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Liu, W.; Kaur, M.; Luo, Y.; Domenico, J.; Samson, J.M.; Shellman, Y.G.; Norris, D.A.; Dinarello, C.A.; Spritz, R.A.; et al. NLRP1 promotes tumor growth by enhancing inflammasome activation and suppressing apoptosis in metastatic melanoma. Oncogene 2017, 36, 3820–3830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovarova, M.; Hesker, P.R.; Jania, L.; Nguyen, M.; Snouwaert, J.N.; Xiang, Z.; Lommatzsch, S.E.; Huang, M.T.; Ting, J.P.-Y.; Koller, B.H. NLRP1-dependent pyroptosis leads to acute lung injury and morbidity in mice. J. Immunol. 2012, 189, 2006–2016. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Characteristic | Chronic Obstructive Pulmonary Disease (COPD) Patients | Healthy Controls |
|---|---|---|
| Sex | ||
| Male | 487 (69.2) | 1116 (87.3) |
| Female | 217 (30.8) | 163 (12.7) |
| Age, year, median (SD, minimum and maximum) | 61 (8.25; 33–85) | 38 (12; 16–66) |
| Smoking History | ||
| Non-smoker | 56 (8.1) | 954 (74.4) |
| Former | 424 (61.5) | 3 (0.2) |
| Active | 209 (30.3) | 325 (25.3) |
| Missing data | 15 (2.1) | 1 (0.1) |
| Characteristic | COPD Patients |
|---|---|
| Smoking, pack-year, median (range) | 40 (0.8–160) |
| GOLD classification | |
| A | 172 (26.1) |
| B | 171 (25.9) |
| C | 172 (26.1) |
| D | 145 (21.9) |
| FEV1/FVC, %, mean (SD) | 56.66 (21.6) |
| FEV1, %, mean (SD) | 55.13 (22.0) |
| Cough | 597/704 (84.8) |
| mMRC 1 | |
| 0 | 59 (8.4) |
| 1 | 168 (23.9) |
| 2 | 238 (33.9) |
| 3 | 166 (23.6) |
| 4 | 72 (10.2) |
| Annual exacerbation rate | |
| 0 | 234 (33.2) |
| 2 | 346 (49.1) |
| >2 | 124 (17.6) |
| Hospital admission | 276 (39.2) |
| Number of comorbidities | |
| 0 | 117 (16.6) |
| 1 | 203 (28.8) |
| 2 | 201 (28.6) |
| ≥3 | 183 (26) |
| Comorbidities | |
| Pulmonary hypertension | 49 (7%) |
| Arterial hypertension | 353 (50.1%) |
| Lung cancer | 72 (10.2%) |
| Cardiovascular diseases | 196 (27.8%) |
| Diabetes mellitus | 100 (14.2%) |
| Extrapulmonary neoplasms | 63 (8.9%) |
| Chronic gastritis /GERD | 71 (10.1%) |
| Chronic kidney disease | 29 (4.1%) |
| Osteoporosis | 35 (5%) |
| Psychiatric disorders | 37 (5.3%) |
| GENE | SNP | MAF | Amino Acid Change | Domain | SIFT | PolyPhen |
|---|---|---|---|---|---|---|
| NLRP1 | rs12150220_A_T | 0.231 (T) | L155H | Deleterious 1 | possibly damaging 1 | |
| NLRP1 | rs35596958_T_C | 0.046 (C) | M1119V | FIIND | tolerated | benign |
| NLRP2 | rs1043673_C_A | 0.416 (A) | A1052E | tolerated | benign | |
| NLRP3 | rs35829419_C_A | 0.022 (A) | Q705K | tolerated | benign | |
| NLRP4 | rs441827_C_T | 0.383 (T) | A144T | tolerated | benign | |
| NLRP4 | rs12462372_G_A | 0.059 (A) | R708H | LRR | tolerated | benign/probably damaging 2 |
| NLRP4 | rs17857373_G_C | 0.013 (C) | P390Q | NACHT | tolerated | benign |
| NLRP4 | rs302453_A_T | 0.219 (T) | Q925L | LRR | tolerated | benign |
| NLRP5 | rs10409555_G_A | 0.258 (A) | V1181I | LRR | tolerated | benign |
| NLRP5 | rs16986899_T_C | 0.265 (C) | M912T | LRR | tolerated | benign |
| NLRP5 | rs471979_G_C | 0.125 (C) | M459I | NACHT | tolerated | benign |
| NLRP6 | rs56159585_T_A | 0.299 (A) | Y361F | NACHT | tolerated | benign |
| NLRP6 | rs77447196_C_G | 0.217 (G) | P244A | NACHT | tolerated | benign |
| NLRP8 | rs306457_C_G | 0.275 (G) | Stop lost | tolerated | benign | |
| NLRP8 | rs306481_G_A | 0.494 (A) | K937R | LRR | tolerated | benign |
| NLRP11 | rs12461110_G_A | 0.243 (A) | P438L | NACHT | tolerated | benign/possibly damaging/probably damaging 3 |
| NLRP11 | rs299163_A_C | 0.047 (C) | A188S | NACHT | tolerated | benign |
| NLRP12 | rs34436714_A_C | 0.267 (C) | G39V | NACHT | deleterious 1 | benign |
| NLRP12 | rs34971363_G_C | 0.051 (C) | F402L | NACHT | deleterious 1 | probably damaging 2 |
| NLRP13 | rs303997_C_T | 0.335 (T) | Q247R | NACHT | deleterious 1 | benign |
| Adjusted for Age/Gender/Smoking | |||||
|---|---|---|---|---|---|
| SNP | Genotype | COPD Cases | Controls | OR (95%CI 2) 1 | p |
| rs12150220 | A/A | 113 (29.20%) | 307 (24.64%) | 1.00 | |
| A/T | 190 (49.10%) | 642 (51.52%) | 0.54 (0.31–0.95) | 0.03 * | |
| T/T | 84 (21.71%) | 297 (23.84%) | 0.59 (0.28–1.26) | 0.17 | |
| A/T+T/T | 274 (70.80%) | 939 (75.36%) | 0.55 (0.32–0.95) | 0.03 * | |
| rs35596958 | T/T | 335 (86.79%) | 1086 (87.16%) | 1.00 | |
| C/T | 49 (12.69%) | 160 (12.84%) | 0.86 (0.42–1.78) | 0.69 | |
| C/C | 2 (0.52%) | 0 (0.00%) | - | - | |
| rs1043673 | C/C | 152 (39.79%) | 471 (37.83%) | 1.00 | |
| A/C | 177 (46.34%) | 589 (47.31%) | 1.13 (0.65–1.95) | 0.67 | |
| A/A | 53 (13.87%) | 185 (14.86%) | 0.92 (0.42–2.01) | 0.83 | |
| rs35829419 | C/C | 346 (89.41%) | 1141 (91.35%) | 1.00 | |
| A/C | 39 (10.08%) | 107 (8.57%) | 1.07 (0.44–2.58) | 0.88 | |
| A/A | 2 (0.52%) | 1 (0.08%) | - | 0.80 | |
| rs441827 | C/C | 155 (40.16%) | 464 (37.09%) | 1.00 | |
| T/C | 168 (43.52%) | 598 (47.80%) | 0.82 (0.47–1.44) | 0.49 | |
| T/T | 63 (16.32%) | 189 (15.11%) | 1.02 (0.50–2.10) | 0.95 | |
| rs12462372 | G/G | 354 (91.00%) | 1092 (88.21%) | 1.00 | |
| A/G | 34 (8.74%) | 143 (11.55%) | 0.36 (0.15–0.90) | 0.03 * | |
| A/A | 1 (0.26%) | 3 (0.24%) | 0.29 (0.00–949.77) | 0.77 | |
| A/G+A/A | 35 (9.00%) | 146 (11.79%) | 0.36 (0.15–0.90) | 0.03 * | |
| rs17857373 | G/G | 359 (92.76%) | 1162 (92.89%) | 1.00 | |
| C/G | 26 (6.72%) | 88 (7.03%) | 0.96 (0.34–2.69) | 0.94 | |
| C/C | 2 (0.52%) | 1 (0.08%) | - | - | |
| rs302453 | A/A | 215 (55.41%) | 681 (54.83%) | 1.00 | |
| A/T | 159 (40.98%) | 471 (37.92%) | 1.59 (0.94–2.71) | 0.09 | |
| T/T | 14 (3.61%) | 90 (7.25%) | 0.31 (0.09–0.99) | 0.05 | |
| rs10409555 | G/G | 212 (54.78%) | 664 (53.12%) | 1.00 | |
| A/G | 142 (36.69%) | 495 (39.60%) | 0.59 (0.34–1.03) | 0.06 | |
| A/A | 33 (8.53%) | 91 (7.28%) | 1.75 (0.73–4.20) | 0.21 | |
| rs16986899 | T/T | 268 (69.79%) | 857 (68.89%) | 1.00 | |
| C/T | 104 (27.08%) | 348 (27.97%) | 1.17 (0.66–2.07) | 0.60 | |
| C/C | 12 (3.13%) | 39 (3.14%) | 0.77 (0.19–3.06) | 0.71 | |
| rs471979 | G/G | 293 (79.62%) | 962 (80.37%) | 1.00 | |
| C/G | 68 (18.48%) | 219 (18.30%) | 0.72 (0.36–1.43) | 0.35 | |
| C/C | 7 (1.90%) | 16 (1.34%) | 4.14 (0.93–18.49) | 0.06 | |
| rs56159585 | T/T | 320 (82.69%) | 969 (79.23%) | 1.00 | |
| A/T | 61 (15.76%) | 238 (19.46%) | 0.54 (0.27–1.07) | 0.08 | |
| A/A | 6 (1.55%) | 16 (1.31%) | 0.11 (0.00–4.43) | 0.24 | |
| rs77447196 | C/C | 265 (68.48%) | 823 (66.42%) | 1.00 | |
| C/G | 117 (30.23%) | 363 (29.30%) | 0.72 (0.40–1.29) | 0.27 | |
| G/G | 5 (1.29%) | 53 (4.28%) | 0.38 (0.07–2.12) | 0.27 | |
| rs306457 | C/C | 227 (58.81%) | 757 (60.71%) | 1.00 | |
| C/G | 134 (34.72%) | 413 (33.12%) | 1.08 (0.63–1.85) | 0.77 | |
| G/G | 25 (6.48%) | 77 (6.17%) | 0.39 (0.12–1.30) | 0.12 | |
| rs306481 | G/G | 142 (36.50%) | 405 (32.58%) | 1.00 | |
| A/G | 190 (48.84%) | 604 (48.59%) | 0.95 (0.55–1.65) | 0.86 | |
| A/A | 57 (14.65%) | 234 (18.83%) | 0.60 (0.27–1.33) | 0.21 | |
| rs12461110 | G/G | 158 (41.15%) | 501 (40.14%) | 1.00 | |
| A/G | 160 (41.67%) | 568 (45.51%) | 0.76 (0.44–1.33) | 0.34 | |
| A/A | 66 (17.19%) | 179 (14.34%) | 1.26 (0.61–2.61) | 0.54 | |
| rs299163 | A/A | 349 (89.95%) | 1066 (85.76%) | 1.00 | |
| A/C | 38 (9.79%) | 172 (13.84%) | 0.93 (0.46–1.90) | 0.85 | |
| C/C | 1 (0.26%) | 5 (0.40%) | 0.02 (0.00–11710.48) | 0.57 | |
| rs34436714 | C/C | 251 (65.88%) | 771 (61.98%) | 1.00 | |
| A/C | 117 (30.71%) | 407 (32.72%) | 0.62 (0.35–1.08) | 0.09 | |
| A/A | 13 (3.41%) | 66 (5.31%) | 0.40 (0.11–1.43) | 0.16 | |
| rs34971363 | G/G | 325 (83.98%) | 1049 (83.79%) | 1.00 | |
| C/G | 60 (15.50%) | 191 (15.26%) | 1.26 (0.62–2.58) | 0.52 | |
| C/C | 2 (0.52%) | 12 (0.96%) | 0.41 (0.01–31.83) | 0.69 | |
| rs303997 | C/C | 124 (32.12%) | 418 (33.63%) | 1.00 | |
| C/T | 201 (52.07%) | 630 (50.68%) | 1.13 (0.65–1.98) | 0.67 | |
| T/T | 61 (15.80%) | 195 (15.69%) | 1.09 (0.51–2.30) | 0.83 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozretić, P.; da Silva Filho, M.I.; Catalano, C.; Sokolović, I.; Vukić-Dugac, A.; Šutić, M.; Kurtović, M.; Bubanović, G.; Popović-Grle, S.; Skrinjarić-Cincar, S.; et al. Association of NLRP1 Coding Polymorphism with Lung Function and Serum IL-1β Concentration in Patients Diagnosed with Chronic Obstructive Pulmonary Disease (COPD). Genes 2019, 10, 783. https://doi.org/10.3390/genes10100783
Ozretić P, da Silva Filho MI, Catalano C, Sokolović I, Vukić-Dugac A, Šutić M, Kurtović M, Bubanović G, Popović-Grle S, Skrinjarić-Cincar S, et al. Association of NLRP1 Coding Polymorphism with Lung Function and Serum IL-1β Concentration in Patients Diagnosed with Chronic Obstructive Pulmonary Disease (COPD). Genes. 2019; 10(10):783. https://doi.org/10.3390/genes10100783
Chicago/Turabian StyleOzretić, Petar, Miguel Inacio da Silva Filho, Calogerina Catalano, Irena Sokolović, Andrea Vukić-Dugac, Maja Šutić, Matea Kurtović, Gordana Bubanović, Sanja Popović-Grle, Sanda Skrinjarić-Cincar, and et al. 2019. "Association of NLRP1 Coding Polymorphism with Lung Function and Serum IL-1β Concentration in Patients Diagnosed with Chronic Obstructive Pulmonary Disease (COPD)" Genes 10, no. 10: 783. https://doi.org/10.3390/genes10100783
APA StyleOzretić, P., da Silva Filho, M. I., Catalano, C., Sokolović, I., Vukić-Dugac, A., Šutić, M., Kurtović, M., Bubanović, G., Popović-Grle, S., Skrinjarić-Cincar, S., Vugrek, O., Jukić, I., Rumora, L., Bosnar, M., Samaržija, M., Bals, R., Jakopović, M., Försti, A., & Knežević, J. (2019). Association of NLRP1 Coding Polymorphism with Lung Function and Serum IL-1β Concentration in Patients Diagnosed with Chronic Obstructive Pulmonary Disease (COPD). Genes, 10(10), 783. https://doi.org/10.3390/genes10100783