TLR7 Expression Is Associated with M2 Macrophage Subset in Calcific Aortic Valve Stenosis

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Collection and Macroscopic Dissection of Human Tricuspid Aortic Valves
2.3. Valve mRNA Expression Microarrays
2.4. Immunofluorescence
2.5. Ex Vivo Human Aortic Valve Stimulation with TLR7 Agonist
2.6. Analysis of Cytokine Secretion
2.7. In Vitro Cell Differentiation and Stimulation
2.8. Statistical Analysis
3. Results
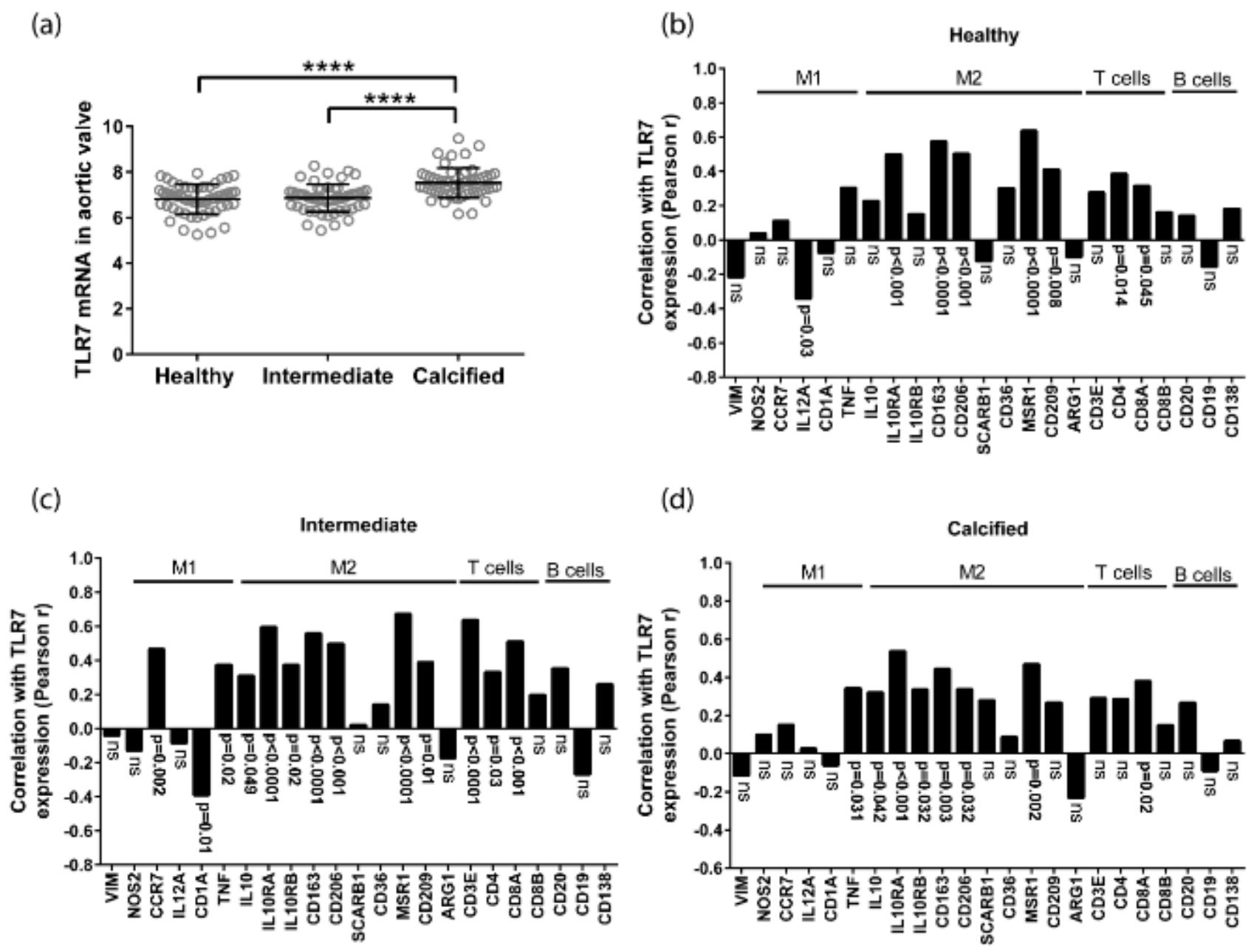
3.1. TLR7 mRNA Is Expressed in Human Aortic Valves and Correlates with M2 Macrophage Markers
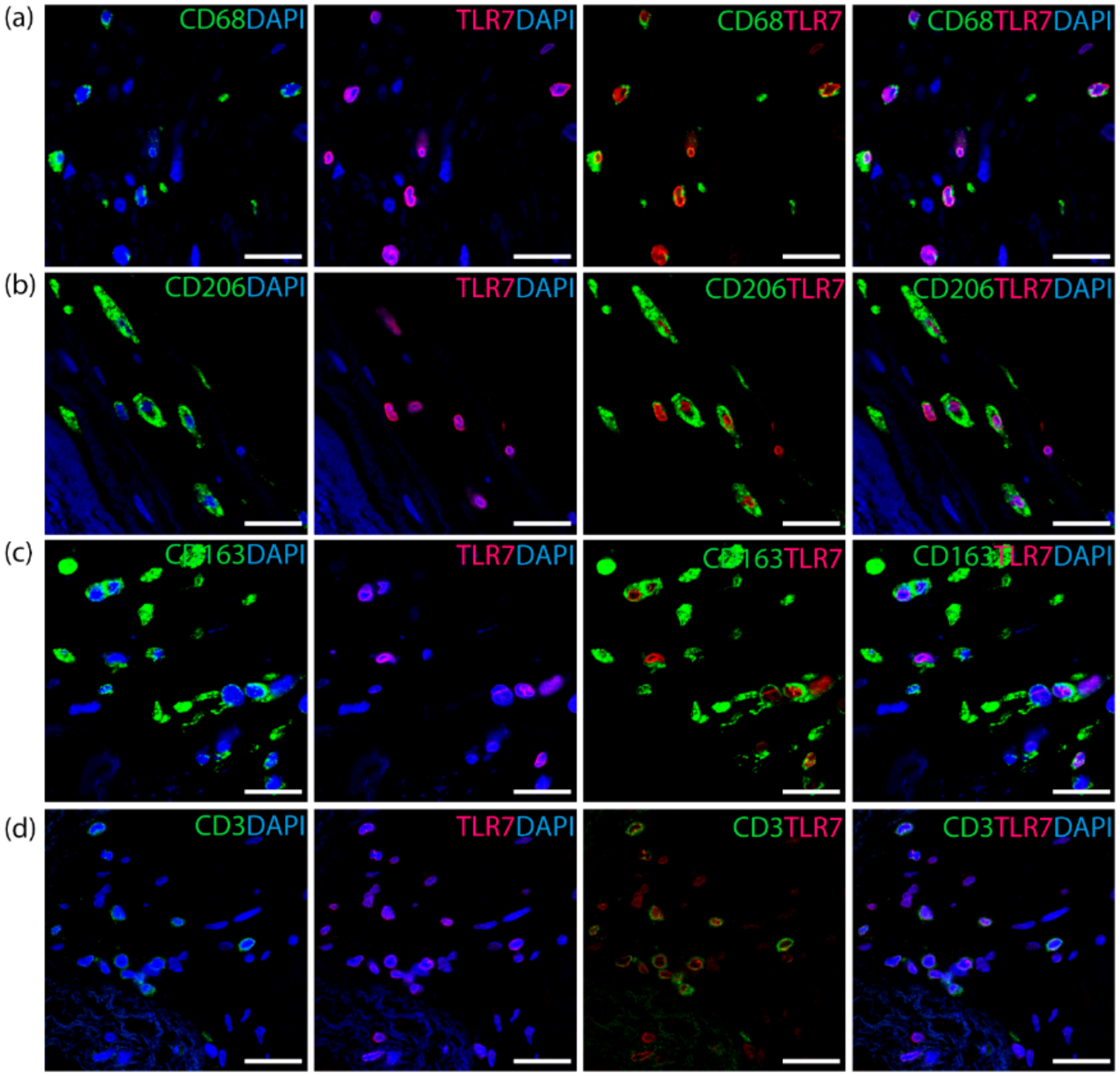
3.2. TLR7 Co-Localizes with T Cells and the M2 Macrophage Subset in Aortic Valves
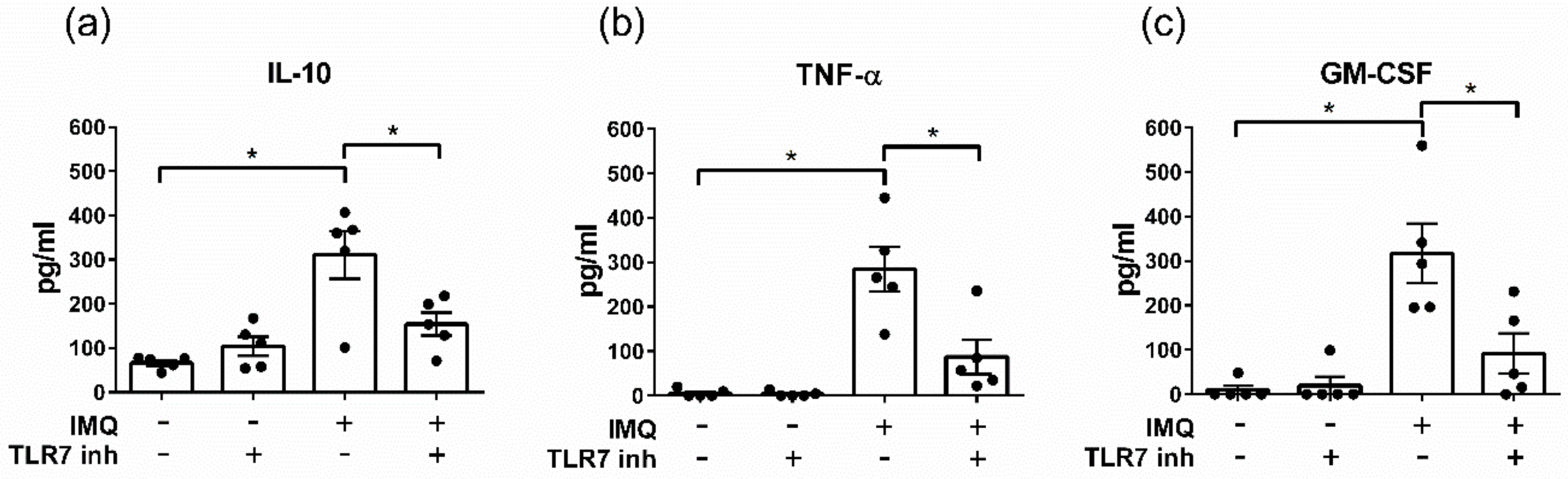
3.3. The TLR7 Agonist Imiquimod Alters Ex Vivo Cytokine Secretion from Human Aortic Valves
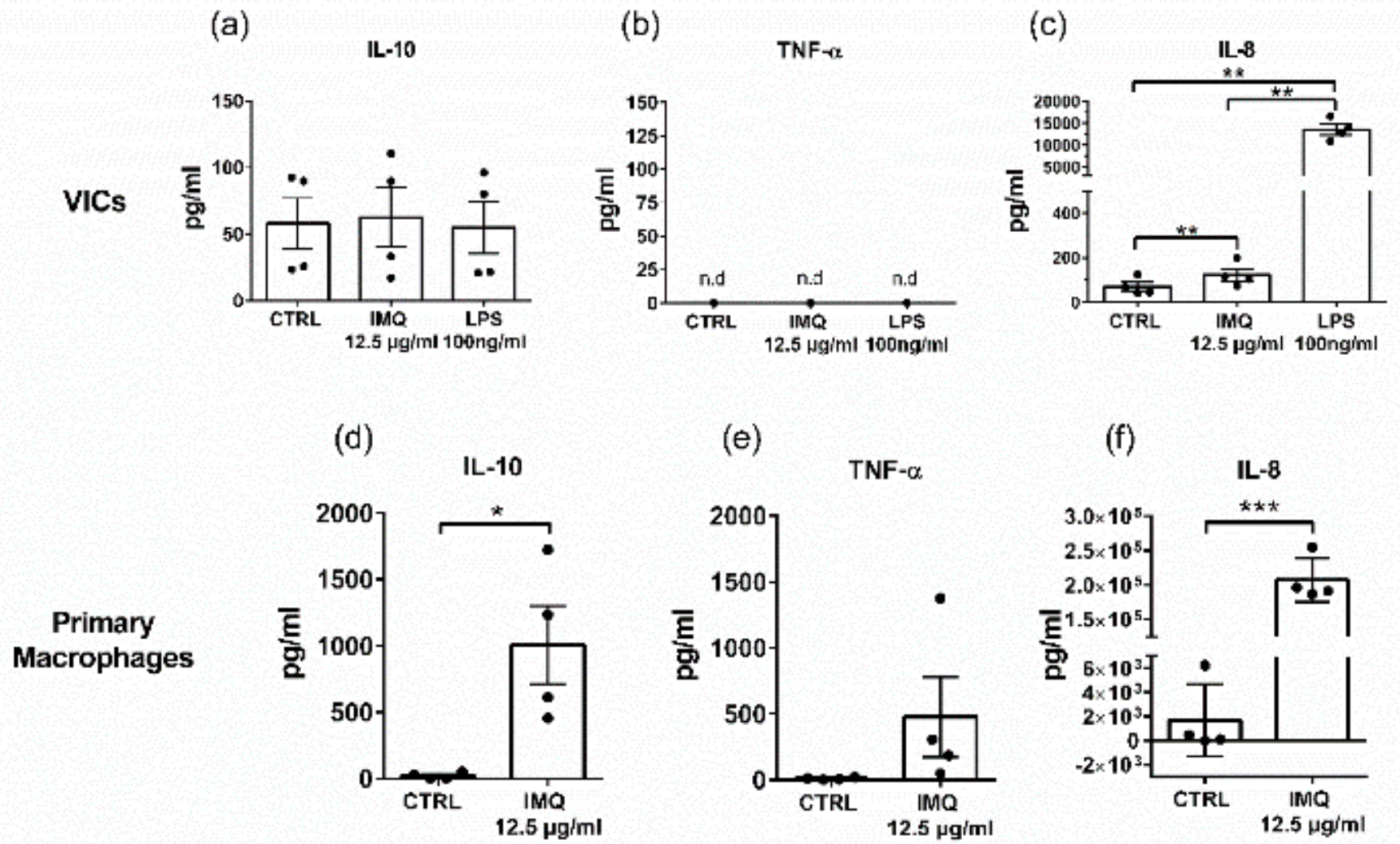
3.4. Secretion of IL-10 and TNF-a from Macrophages
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Go, A.S.; Mozaffarian, D.; Roger, V.L.; Benjamin, E.J.; Berry, J.D.; Borden, W.B.; Bravata, D.M.; Dai, S.; Ford, E.S.; Fox, C.S.; et al. Executive Summary: Heart Disease and Stroke Statistics—2013 Update. Circulation 2013, 127, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Lindman, B.R.; Clavel, M.-A.; Mathieu, P.; Iung, B.; Lancellotti, P.; Otto, C.M.; Pibarot, P. Calcific aortic stenosis. Nat. Rev. Dis. Primers 2016, 2, 16006. [Google Scholar] [CrossRef] [PubMed]
- Vavilis, G.; Bäck, M.; Occhino, G.; Trevisan, M.; Bellocco, R.; Evans, M.; Lindholm, B.; Szummer, K.; Carrero, J.J. Kidney Dysfunction and the Risk of Developing Aortic Stenosis. J. Am. Coll. Cardiol. 2019, 73, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.C.; Back, M.; Rees, J.M.B.; Mason, A.M.; Burgess, S. Body mass index and body composition in relation to 14 cardiovascular conditions in UK Biobank: A Mendelian randomization study. Eur. Heart J. 2019. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.C.; Wolk, A.; Back, M. Alcohol consumption, cigarette smoking and incidence of aortic valve stenosis. J. Intern. Med. 2017, 282, 332–339. [Google Scholar] [CrossRef]
- Larsson, S.C.; Wolk, A.; Hakansson, N.; Back, M. Overall and abdominal obesity and incident aortic valve stenosis: Two prospective cohort studies. Eur. Heart J. 2017, 38, 2192–2197. [Google Scholar] [CrossRef]
- Yutzey, K.E.; Demer, L.L.; Body, S.C.; Huggins, G.S.; Towler, D.A.; Giachelli, C.M.; Hofmann-Bowman, M.A.; Mortlock, D.P.; Rogers, M.B.; Sadeghi, M.M.; et al. Calcific Aortic Valve Disease. Arter. Thromb. Vasc. Biol. 2014, 34, 2387–2393. [Google Scholar] [CrossRef]
- Freeman, R.V.; Otto, C.M. Spectrum of Calcific Aortic Valve Disease. Circulation 2005, 111, 3316–3326. [Google Scholar] [CrossRef]
- Nishimura, R.A.; Otto, C.M.; Bonow, R.O.; Carabello, B.A.; Erwin, J.P.; Guyton, R.A.; O’Gara, P.T.; Ruiz, C.E.; Skubas, N.J.; Sorajja, P.; et al. 2014 AHA/ACC Guideline for the Management of Patients With Valvular Heart Disease: A Report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines. J. Am. Coll. Cardiol. 2014, 63, 57–185. [Google Scholar] [CrossRef]
- Members, A.T.F.; Vahanian, A.; Alfieri, O.; Andreotti, F.; Antunes, M.J.; Barón-Esquivias, G.; Baumgartner, H.; Borger, M.A.; Carrel, T.P.; De Bonis, M.; et al. Guidelines on the management of valvular heart disease (version 2012): The Joint Task Force on the Management of Valvular Heart Disease of the European Society of Cardiology (ESC) and the European Association for Cardio-Thoracic Surgery (EACTS). Eur. Heart J. 2012, 33, 2451–2496. [Google Scholar] [CrossRef]
- Raddatz, M.A.; Madhur, M.S.; Merryman, W.D. Adaptive immune cells in calcific aortic valve disease. Am. J. Physiol. Heart Circ. Physiol. 2019, 317, H141–H155. [Google Scholar] [CrossRef] [PubMed]
- Coté, N.; Mahmut, A.; Bosse, Y.; Couture, C.; Pagé, S.; Trahan, S.; Boulanger, M.-C.; Fournier, D.; Pibarot, P.; Mathieu, P. Inflammation Is Associated with the Remodeling of Calcific Aortic Valve Disease. Inflammation 2013, 36, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Artiach, G.; Carracedo, M.; Plunde, O.; Wheelock, C.E.; Thul, S.; Sjövall, P.; Franco-Cereceda, A.; Laguna-Fernandez, A.; Arnardottir, H.; Bäck, M. Omega-3 Polyunsaturated Fatty Acids Decrease Aortic Valve Disease through the Resolvin E1 and ChemR23 Axis. Circulation 2020. [Google Scholar] [CrossRef]
- Moore, K.J.; Sheedy, F.J.; Fisher, E.A. Macrophages in atherosclerosis: A dynamic balance. Nat. Rev. Immunol. 2013, 13, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.E.; Georgiou, E.; Monaco, C. The Expression and Functions of Toll-Like Receptors in Atherosclerosis. Mediat. Inflamm. 2010, 2010, 18. [Google Scholar] [CrossRef] [PubMed]
- Cohen, I.; Zanin-Zhorov, A. Signaling via TLR2 and TLR4 Directly Down-Regulates T Cell Effector Functions: The Regulatory Face of Danger Signals. Front. Immunol. 2013, 4. [Google Scholar] [CrossRef]
- Meng, X.; Ao, L.; Song, Y.; Babu, A.; Yang, X.; Wang, M.; Weyant, M.J.; Dinarello, C.A.; Joseph, C.; Cleveland, J. Expression of functional Toll-like receptors 2 and 4 in human aortic valve interstitial cells: Potential roles in aortic valve inflammation and stenosis. Am. J. Physiol. Cell Physiol. 2008, 294, C29–C35. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Karadimou, G.; Folkersen, L.; Berg, M.; Perisic, L.; Discacciati, A.; Roy, J.; Hansson, G.K.; Persson, J.; Paulsson-Berne, G. Low TLR7 gene expression in atherosclerotic plaques is associated with major adverse cardio- and cerebrovascular events. Cardiovasc. Res. 2016, 113, 30–39. [Google Scholar] [CrossRef]
- Salagianni, M.; Galani, I.E.; Lundberg, A.M.; Davos, C.H.; Varela, A.; Gavriil, A.; Lyytikäinen, L.-P.; Lehtimäki, T.; Sigala, F.; Folkersen, L.; et al. Toll-Like Receptor 7 Protects From Atherosclerosis by Constraining “Inflammatory” Macrophage Activation. Circulation 2012, 126, 952–962. [Google Scholar] [CrossRef]
- Karadimou, G.; Gisterå, A.; Gallina, A.L.; Caravaca, A.S.; Centa, M.; Salagianni, M.; Andreakos, E.; Hansson, G.K.; Malin, S.; Olofsson, P.S.; et al. Treatment with a Toll-like Receptor 7 ligand evokes protective immunity against atherosclerosis in hypercholesterolaemic mice. J. Int. Med. 2020. [Google Scholar] [CrossRef]
- Laguna-Fernandez, A.; Carracedo, M.; Jeanson, G.; Nagy, E.; Eriksson, P.; Caligiuri, G.; Franco-Cereceda, A.; Bäck, M. Iron alters valvular interstitial cell function and is associated with calcification in aortic stenosis. Eur. Heart J. 2016, 37, 3532–3535. [Google Scholar] [CrossRef] [PubMed]
- Nagy, E.; Andersson, D.C.; Caidahl, K.; Eriksson, M.J.; Eriksson, P.; Franco-Cereceda, A.; Hansson, G.K.; Bäck, M. Upregulation of the 5-Lipoxygenase Pathway in Human Aortic Valves Correlates With Severity of Stenosis and Leads to Leukotriene-Induced Effects on Valvular Myofibroblasts. Circulation 2011, 123, 1316–1325. [Google Scholar] [CrossRef] [PubMed]
- Mercier, N.; Pawelzik, S.-C.; Pirault, J.; Carracedo, M.; Persson, O.; Wollensack, B.; Franco-Cereceda, A.; Bäck, M. Semicarbazide-Sensitive Amine Oxidase Increases in Calcific Aortic Valve Stenosis and Contributes to Valvular Interstitial Cell Calcification. Oxidative Med. Cell. Longev. 2020, 2020. [Google Scholar] [CrossRef] [PubMed]
- Rutkovskiy, A.; Malashicheva, A.; Sullivan, G.; Bogdanova, M.; Kostareva, A.; Stensløkken, K.-O.; Fiane, A.; Vaage, J. Valve Interstitial Cells: The Key to Understanding the Pathophysiology of Heart Valve Calcification. J. Am. Heart Assoc. 2017, 6, e006339. [Google Scholar] [CrossRef]
- Zeng, Q.; Jin, C.; Ao, L.; Cleveland, J.C.; Song, R.; Xu, D.; Fullerton, D.A.; Meng, X. Cross-Talk Between the Toll-Like Receptor 4 and Notch1 Pathways Augments the Inflammatory Response in the Interstitial Cells of Stenotic Human Aortic Valves. Circulation 2012, 126, S222–S230. [Google Scholar] [CrossRef] [PubMed]
- García-Rodríguez, C.; Parra-Izquierdo, I.; Castaños-Mollor, I.; López, J.; San Román, J.A.; Sánchez Crespo, M. Toll-Like Receptors, Inflammation, and Calcific Aortic Valve Disease. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- López, J.; Fernández-Pisonero, I.; Dueñas, A.I.; Maeso, P.; Román, J.A.S.; Crespo, M.S.; García-Rodríguez, C. Viral and bacterial patterns induce TLR-mediated sustained inflammation and calcification in aortic valve interstitial cells. Int. J. Cardiol. 2012, 158, 18–25. [Google Scholar] [CrossRef]
- Kapelouzou, A.; Kontogiannis, C.; Tsilimigras, D.I.; Georgiopoulos, G.; Kaklamanis, L.; Tsourelis, L.; Cokkinos, D.V. Differential expression patterns of Toll Like Receptors and Interleukin-37 between calcific aortic and mitral valve cusps in humans. Cytokine 2019, 116, 150–160. [Google Scholar] [CrossRef]
- Gordon, S.; Plüddemann, A. Tissue macrophages: Heterogeneity and functions. BMC Biol. 2017, 15, 53. [Google Scholar] [CrossRef]
- Yap, J.; Cabrera-Fuentes, H.A.; Irei, J.; Hausenloy, D.J.; Boisvert, W.A. Role of Macrophages in Cardioprotection. Int. J. Mol. Sci 2019, 20, 2474. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, E.J.; Parker, A.E.; O’Neill, L.A.J. Targeting Toll-like receptors: Emerging therapeutics? Nat. Rev. Drug Discov. 2010, 9, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-F.; Wang, Y.; Zheng, D.-D.; Xu, H.-X.; Wang, T.; Pan, M.; Shi, J.-H.; Zhu, J.-H. M1 macrophages promote aortic valve calcification mediated by microRNA-214/TWIST1 pathway in valvular interstitial cells. Am J. Transl. Res. 2016, 8, 5773–5783. [Google Scholar] [PubMed]
- Shiraishi, M.; Shintani, Y.; Shintani, Y.; Ishida, H.; Saba, R.; Yamaguchi, A.; Adachi, H.; Yashiro, K.; Suzuki, K. Alternatively activated macrophages determine repair of the infarcted adult murine heart. J. Clin. Investig. 2016, 126, 2151–2166. [Google Scholar] [CrossRef]
- Larson, S.R.; Atif, S.M.; Gibbings, S.L.; Thomas, S.M.; Prabagar, M.G.; Danhorn, T.; Leach, S.M.; Henson, P.M.; Jakubzick, C.V. Ly6C(+) monocyte efferocytosis and cross-presentation of cell-associated antigens. Cell Death Differ. 2016, 23, 997–1003. [Google Scholar] [CrossRef]
- Celhar, T.; Pereira-Lopes, S.; Thornhill, S.I.; Lee, H.Y.; Dhillon, M.K.; Poidinger, M.; Connolly, J.E.; Lim, L.H.K.; Biswas, S.K.; Fairhurst, A.-M. TLR7 and TLR9 ligands regulate antigen presentation by macrophages. Int. Immunol. 2015, 28, 223–232. [Google Scholar] [CrossRef]
- Nagy, E.; Lei, Y.; Martínez-Martínez, E.; Body, S.C.; Schlotter, F.; Creager, M.; Assmann, A.; Khabbaz, K.; Libby, P.; Hansson, G.K.; et al. Interferon-γ Released by Activated CD8+ T Lymphocytes Impairs the Calcium Resorption Potential of Osteoclasts in Calcified Human Aortic Valves. Am. J. Pathol. 2017, 187, 1413–1425. [Google Scholar] [CrossRef]
- Clement, M.; Guedj, K.; Andreata, F.; Morvan, M.; Bey, L.; Khallou-Laschet, J.; Gaston, A.-T.; Delbosc, S.; Alsac, J.-M.; Bruneval, P.; et al. Control of the T Follicular Helper–Germinal Center B-Cell Axis by CD8+ Regulatory T Cells Limits Atherosclerosis and Tertiary Lymphoid Organ Development. Circulation 2015, 131, 560–570. [Google Scholar] [CrossRef]
- Shimoni, S.; Bar, I.; Meledin, V.; Gandelman, G.; George, J. Circulating regulatory T cells in patients with aortic valve stenosis: Association with disease progression and aortic valve intervention. Int. J. Cardiol. 2016, 218, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Van Duijn, J.; Kritikou, E.; Benne, N.; van der Heijden, T.; van Puijvelde, G.H.; Kröner, M.J.; Schaftenaar, F.H.; Foks, A.C.; Wezel, A.; Smeets, H.; et al. CD8+ T-cells contribute to lesion stabilization in advanced atherosclerosis by limiting macrophage content and CD4+ T-cell responses. Cardiovasc. Res. 2018, 115, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Boisvert, W.A. Interleukin-10 protects against atherosclerosis by modulating multiple atherogenic macrophage function. Thromb. Haemost. 2015, 113, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Mallat, Z.; Heymes, C.; Ohan, J.; Faggin, E.; Lesèche, G.; Tedgui, A. Expression of Interleukin-10 in Advanced Human Atherosclerotic Plaques. Arter. Thromb. Vasc. Biol. 1999, 19, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Kessler, B.; Rinchai, D.; Kewcharoenwong, C.; Nithichanon, A.; Biggart, R.; Hawrylowicz, C.M.; Bancroft, G.J.; Lertmemongkolchai, G. Interleukin 10 inhibits pro-inflammatory cytokine responses and killing of Burkholderia pseudomallei. Sci. Rep. 2017, 7, 42791. [Google Scholar] [CrossRef] [PubMed]
- Fadok, V.A.; Bratton, D.L.; Guthrie, L.; Henson, P.M. Differential Effects of Apoptotic Versus Lysed Cells on Macrophage Production of Cytokines: Role of Proteases. J. Immunol. 2001, 166, 6847. [Google Scholar] [CrossRef]
- Yu, Z.; Seya, K.; Daitoku, K.; Motomura, S.; Fukuda, I.; Furukawa, K.-I. Tumor Necrosis Factor-α Accelerates the Calcification of Human Aortic Valve Interstitial Cells Obtained from Patients with Calcific Aortic Valve Stenosis via the BMP2-Dlx5 Pathway. J. Pharmacol. Exp. Ther. 2011, 337, 16–23. [Google Scholar] [CrossRef]
- Durán, W.N. The double-edge sword of TNF-alpha in ischemia-reperfusion injury. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H2221–H2222. [Google Scholar] [CrossRef]
- Papathanasiou, S.; Rickelt, S.; Soriano, M.E.; Schips, T.G.; Maier, H.J.; Davos, C.H.; Varela, A.; Kaklamanis, L.; Mann, D.L.; Capetanaki, Y. Tumor necrosis factor-α confers cardioprotection through ectopic expression of keratins K8 and K18. Nat. Med. 2015, 21, 1076–1084. [Google Scholar] [CrossRef]
- Parra-Izquierdo, I.; Castaños-Mollor, I.; López, J.; Gómez, C.; Román, J.A.S.; Crespo, M.S.; García-Rodríguez, C. Calcification Induced by Type I Interferon in Human Aortic Valve Interstitial Cells Is Larger in Males and Blunted by a Janus Kinase Inhibitor. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 2148–2159. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Calcified vs Healthy | Calcified vs Intermediate | Intermediate vs Healthy | ||
|---|---|---|---|---|
| TLR7 | Fold change | 1.77 | 1.80 | 1.05 |
| CI | 1.57–1.97 | 1.53–2.07 | 0.93–1.17 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karadimou, G.; Plunde, O.; Pawelzik, S.-C.; Carracedo, M.; Eriksson, P.; Franco-Cereceda, A.; Paulsson-Berne, G.; Bäck, M. TLR7 Expression Is Associated with M2 Macrophage Subset in Calcific Aortic Valve Stenosis. Cells 2020, 9, 1710. https://doi.org/10.3390/cells9071710
Karadimou G, Plunde O, Pawelzik S-C, Carracedo M, Eriksson P, Franco-Cereceda A, Paulsson-Berne G, Bäck M. TLR7 Expression Is Associated with M2 Macrophage Subset in Calcific Aortic Valve Stenosis. Cells. 2020; 9(7):1710. https://doi.org/10.3390/cells9071710
Chicago/Turabian StyleKaradimou, Glykeria, Oscar Plunde, Sven-Christian Pawelzik, Miguel Carracedo, Per Eriksson, Anders Franco-Cereceda, Gabrielle Paulsson-Berne, and Magnus Bäck. 2020. "TLR7 Expression Is Associated with M2 Macrophage Subset in Calcific Aortic Valve Stenosis" Cells 9, no. 7: 1710. https://doi.org/10.3390/cells9071710
APA StyleKaradimou, G., Plunde, O., Pawelzik, S.-C., Carracedo, M., Eriksson, P., Franco-Cereceda, A., Paulsson-Berne, G., & Bäck, M. (2020). TLR7 Expression Is Associated with M2 Macrophage Subset in Calcific Aortic Valve Stenosis. Cells, 9(7), 1710. https://doi.org/10.3390/cells9071710