A Proximity Mapping Journey into the Biology of the Mammalian Centrosome/Cilium Complex


Abstract
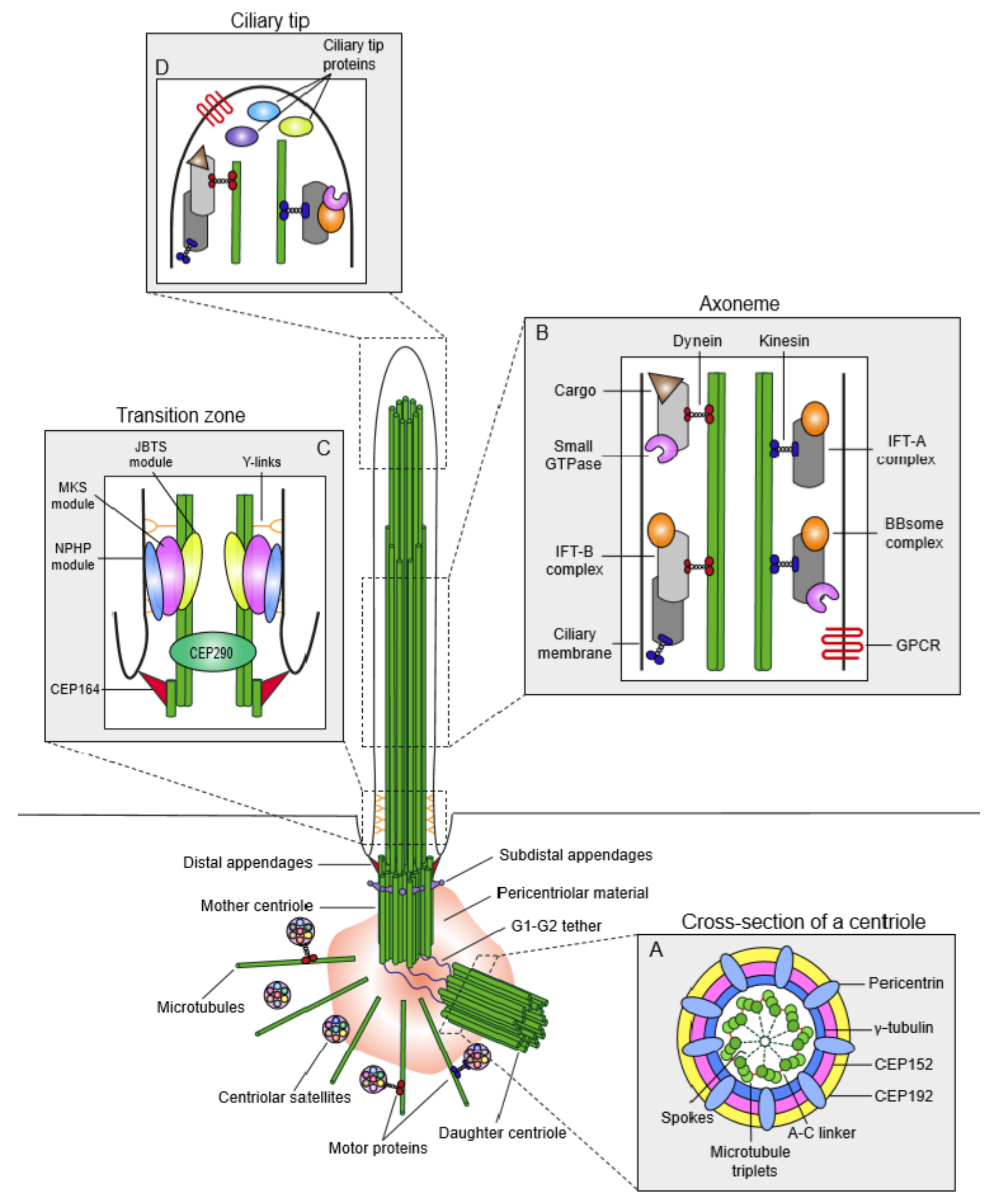
1. Structure of the Mammalian Centrosome/Cilium Complex
1.1. Centrosomes
1.2. Primary Cilium
1.3. Centriolar Satellites
2. Challenges in Mapping the Interactions at the Centrosome/Cilium Complex
2.1. Structural and Numerical Challenges
2.2. Temporal Challenges
3. Protein–Protein Interactions and Methods for Their Identification
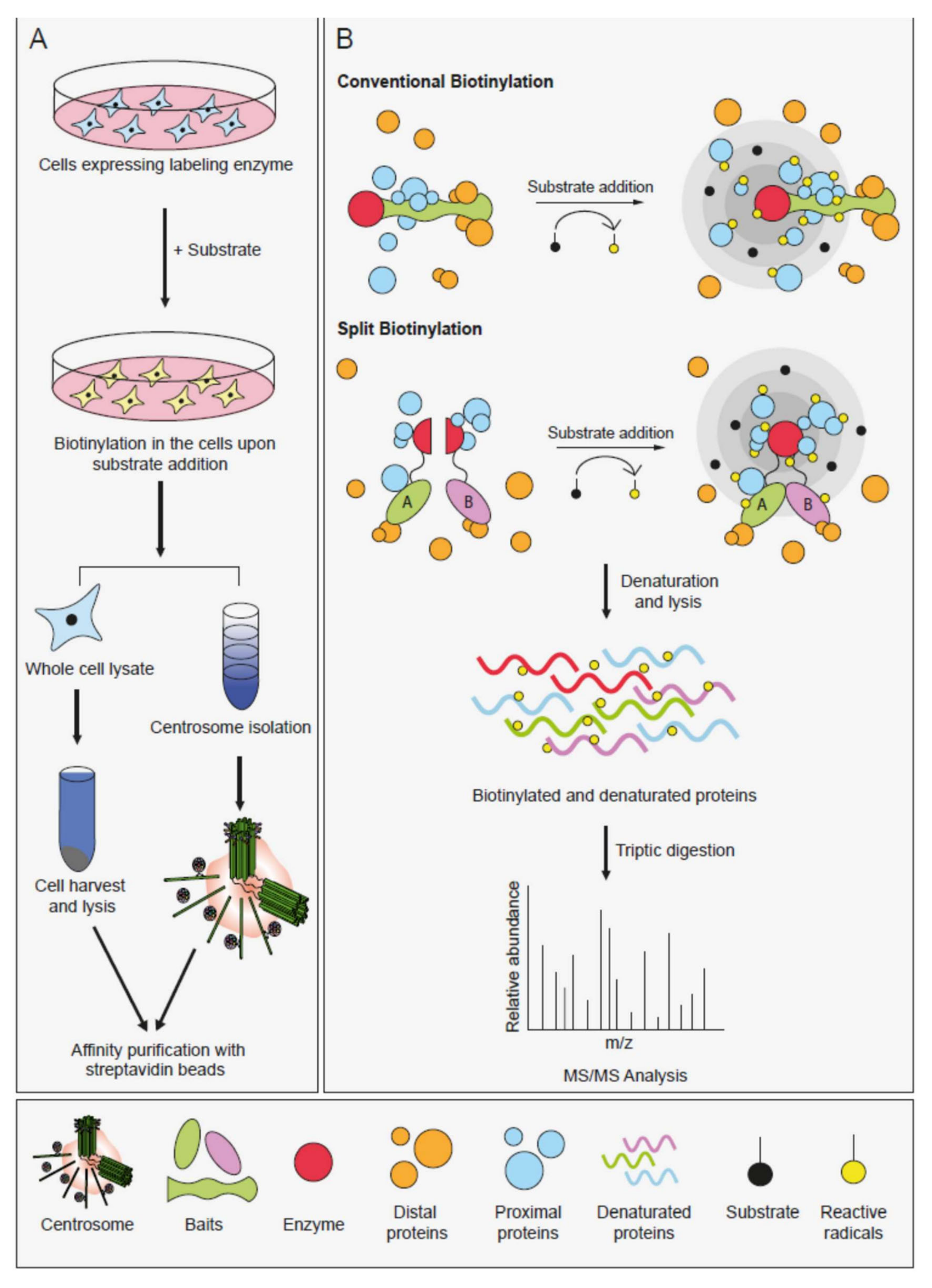
4. Proximity-Based Labeling Methods
5. Application of Proximity-Based Labeling Methods to the Centrosome/Cilium Complex
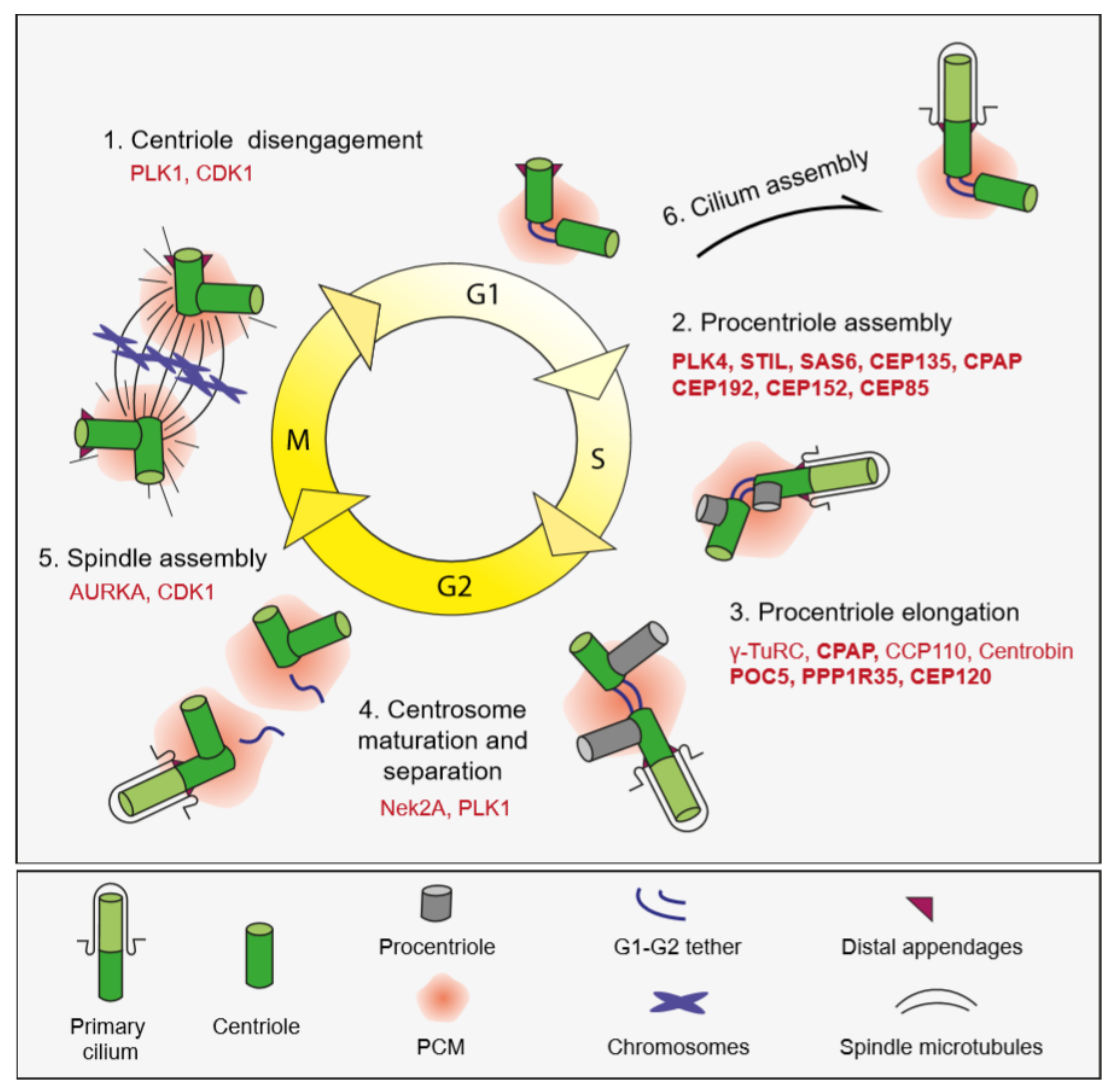
5.1. Centrosome Biogenesis
5.2. Primary Cilium Biogenesis and Proteome
5.3. Proximity Landscape of Centrosomes and Centriolar Satellites
6. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PCM | pericentriolar material |
| MTOC | microtubule organizing center |
| DAP | distal appendage protein |
| sDAP | subdistal appendage protein |
| U-ExM | Ultrastructure Expansion Microscopy |
| STORM | Stochastic Optical Reconstruction Microscopy |
| NPHP | nephronophthisis |
| MKS | Meckel-Gruber Syndrome |
| JBTS | Joubert syndrome |
| GAP | GTPase-activating protein |
| GEF | Guanine nucleotide exchange factor |
| AURKA | Aurora kinase A |
| Nek2A | NIMA related kinase 2A |
| GPCR | G protein-coupled receptor |
| TTLL | Tubulin tyrosine ligase |
| BioID | Biotin Identification |
| APEX | Ascorbate Peroxidase |
| HTR6 | 5-hydroxytryptamine receptor 6 |
| IFT | Intraflagellar Transport |
| BBS | Bardet-Biedl Syndrome |
References
- Breslow, D.K.; Holland, A.J. Mechanism and Regulation of Centriole and Cilium Biogenesis. Annu. Rev. Biochem. 2019, 88, 691–724. [Google Scholar] [CrossRef]
- Odabasi, E.; Batman, U.; Firat-Karalar, E.N. Unraveling the mysteries of centriolar satellites: Time to rewrite the textbooks about the centrosome/cilium complex. Mol. Biol. Cell 2020, 31, 866–872. [Google Scholar] [CrossRef]
- Prosser, S.L.; Pelletier, L. Centriolar satellite biogenesis and function in vertebrate cells. J. Cell Sci. 2020, 133. [Google Scholar] [CrossRef]
- Gomes, E.; Shorter, J. The molecular language of membraneless organelles. J. Biol. Chem. 2019, 294, 7115–7127. [Google Scholar] [CrossRef]
- Arquint, C.; Gabryjonczyk, A.M.; Nigg, E.A. Centrosomes as signalling centres. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef]
- Woodruff, J.B.; Wueseke, O.; Hyman, A.A. Pericentriolar material structure and dynamics. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef]
- Carvalho-Santos, Z.; Azimzadeh, J.; Pereira-Leal, J.B.; Bettencourt-Dias, M. Evolution: Tracing the origins of centrioles, cilia, and flagella. J. Cell. Biol. 2011, 194, 165–175. [Google Scholar] [CrossRef]
- Hodges, M.E.; Scheumann, N.; Wickstead, B.; Langdale, J.A.; Gull, K. Reconstructing the evolutionary history of the centriole from protein components. J. Cell Sci. 2010, 123, 1407–1413. [Google Scholar] [CrossRef]
- Alieva, I.; Staub, C.; Uzbekova, S.V.; Uzbekov, R. A Question of Flagella Origin for Spermatids: Mother or Daughter Centriole? Nova Sci. Publishers: New York, NY, USA, 2018; pp. 109–126. [Google Scholar]
- Azimzadeh, J.; Marshall, W.F. Building the centriole. Curr. Biol. 2010, 20, R816–R825. [Google Scholar] [CrossRef]
- Loncarek, J.; Bettencourt-Dias, M. Building the right centriole for each cell type. J. Cell Biol. 2018, 217, 823–835. [Google Scholar] [CrossRef]
- Wang, J.T.; Stearns, T. The ABCs of Centriole Architecture: The Form and Function of Triplet Microtubules. Cold Spring Harb. Symp. Quant. Biol. 2017, 82, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Winey, M.; O’Toole, E. Centriole structure. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef] [PubMed]
- Hamel, V.; Steib, E.; Hamelin, R.; Armand, F.; Borgers, S.; Fluckiger, I.; Busso, C.; Olieric, N.; Sorzano, C.O.S.; Steinmetz, M.O.; et al. Identification of Chlamydomonas Central Core Centriolar Proteins Reveals a Role for Human WDR90 in Ciliogenesis. Curr. Biol. 2017, 27, 2486–2498. [Google Scholar] [CrossRef]
- Le Guennec, M.; Klena, N.; Gambarotto, D.; Laporte, M.H.; Tassin, A.M.; van den Hoek, H.; Erdmann, P.S.; Schaffer, M.; Kovacik, L.; Borgers, S.; et al. A helical inner scaffold provides a structural basis for centriole cohesion. Sci. Adv. 2020, 6, 4137. [Google Scholar] [CrossRef]
- Agircan, F.G.; Schiebel, E.; Mardin, B.R. Separate to operate: Control of centrosome positioning and separation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef]
- Giet, R.; Uzbekov, R.; Cubizolles, F.; Le Guellec, K.; Prigent, C. The Xenopus laevis aurora-related protein kinase pEg2 associates with and phosphorylates the kinesin-related protein XlEg5. J. Biol. Chem. 1999, 274, 15005–15013. [Google Scholar] [CrossRef]
- Nigg, E.A.; Stearns, T. The centrosome cycle: Centriole biogenesis, duplication and inherent asymmetries. Nat. Cell Biol. 2011, 13, 1154–1160. [Google Scholar] [CrossRef]
- Remo, A.; Li, X.; Schiebel, E.; Pancione, M. The Centrosome Linker and Its Role in Cancer and Genetic Disorders. Trends Mol. Med. 2020, 26, 380–393. [Google Scholar] [CrossRef]
- Mardin, B.R.; Schiebel, E. Breaking the ties that bind: New advances in centrosome biology. J. Cell Biol. 2012, 197, 11–18. [Google Scholar] [CrossRef]
- Gupta, A.; Kitagawa, D. Ultrastructural diversity between centrioles of eukaryotes. J. Biochem. 2018, 164, 1–8. [Google Scholar] [CrossRef]
- Uzbekov, R.; Alieva, I. Who are you, subdistal appendages of centriole? Open Biol. 2018, 8. [Google Scholar] [CrossRef]
- Bowler, M.; Kong, D.; Sun, S.; Nanjundappa, R.; Evans, L.; Farmer, V.; Holland, A.; Mahjoub, M.R.; Sui, H.; Loncarek, J. High-resolution characterization of centriole distal appendage morphology and dynamics by correlative STORM and electron microscopy. Nat. Commun. 2019, 10, 993. [Google Scholar] [CrossRef]
- Chong, W.M.; Wang, W.J.; Lo, C.H.; Chiu, T.Y.; Chang, T.J.; Liu, Y.P.; Tanos, B.; Mazo, G.; Tsou, M.B.; Jane, W.N.; et al. Super-resolution microscopy reveals coupling between mammalian centriole subdistal appendages and distal appendages. eLife 2020, 9. [Google Scholar] [CrossRef]
- Yang, T.T.; Chong, W.M.; Wang, W.J.; Mazo, G.; Tanos, B.; Chen, Z.; Tran, T.M.N.; Chen, Y.D.; Weng, R.R.; Huang, C.E.; et al. Super-resolution architecture of mammalian centriole distal appendages reveals distinct blade and matrix functional components. Nat. Commun. 2018, 9, 2023. [Google Scholar] [CrossRef]
- Li, S.; Fernandez, J.J.; Marshall, W.F.; Agard, D.A. Three-dimensional structure of basal body triplet revealed by electron cryo-tomography. EMBO J. 2012, 31, 552–562. [Google Scholar] [CrossRef]
- Steib, E.; Gambarotto, D.; Laporte, M.; O’lieric, N.; Zheng, C.; Borgers, S.; Olieric, V.; Guennec, M.L.; Koll, F.; Tassin, A.M.; et al. WDR90 is a centriolar microtubule wall protein important for centriole architecture integrity. bioRxiv 2020. [Google Scholar] [CrossRef]
- Chang, J.; Cizmecioglu, O.; Hoffmann, I.; Rhee, K. PLK2 phosphorylation is critical for CPAP function in procentriole formation during the centrosome cycle. EMBO J. 2010, 29, 2395–2406. [Google Scholar] [CrossRef]
- Huang, N.; Xia, Y.; Zhang, D.; Wang, S.; Bao, Y.; He, R.; Teng, J.; Chen, J. Hierarchical assembly of centriole subdistal appendages via centrosome binding proteins CCDC120 and CCDC68. Nat. Commun. 2017, 8, 15057. [Google Scholar] [CrossRef]
- Mazo, G.; Soplop, N.; Wang, W.J.; Uryu, K.; Tsou, M.F. Spatial Control of Primary Ciliogenesis by Subdistal Appendages Alters Sensation-Associated Properties of Cilia. Dev. Cell 2016, 39, 424–437. [Google Scholar] [CrossRef]
- Tanos, B.E.; Yang, H.J.; Soni, R.; Wang, W.J.; Macaluso, F.P.; Asara, J.M.; Tsou, M.F. Centriole distal appendages promote membrane docking, leading to cilia initiation. Genes Dev. 2013, 27, 163–168. [Google Scholar] [CrossRef]
- Paz, J.; Luders, J. Microtubule-Organizing Centers: Towards a Minimal Parts List. Trends Cell Biol. 2018, 28, 176–187. [Google Scholar] [CrossRef]
- Alieva, I.B.; Uzbekov, R.E. Where are the limits of the centrosome? Bioarchitecture 2016, 6, 47–52. [Google Scholar] [CrossRef]
- Lawo, S.; Hasegan, M.; Gupta, G.D.; Pelletier, L. Subdiffraction imaging of centrosomes reveals higher-order organizational features of pericentriolar material. Nat. Cell Biol. 2012, 14, 1148–1158. [Google Scholar] [CrossRef]
- Luders, J. The amorphous pericentriolar cloud takes shape. Nat. Cell Biol. 2012, 14, 1126–1128. [Google Scholar] [CrossRef]
- Mennella, V.; Keszthelyi, B.; McDonald, K.L.; Chhun, B.; Kan, F.; Rogers, G.C.; Huang, B.; Agard, D.A. Subdiffraction-resolution fluorescence microscopy reveals a domain of the centrosome critical for pericentriolar material organization. Nat. Cell Biol. 2012, 14, 1159–1168. [Google Scholar] [CrossRef]
- Sonnen, K.F.; Schermelleh, L.; Leonhardt, H.; Nigg, E.A. 3D-structured illumination microscopy provides novel insight into architecture of human centrosomes. Biol. Open 2012, 1, 965–976. [Google Scholar] [CrossRef]
- Varadarajan, R.; Rusan, N.M. Bridging centrioles and PCM in proper space and time. Essays Biochem. 2018, 62, 793–801. [Google Scholar] [CrossRef]
- Golsteyn, R.M.; Mundt, K.E.; Fry, A.M.; Nigg, E.A. Cell cycle regulation of the activity and subcellular localization of Plk1, a human protein kinase implicated in mitotic spindle function. J. Cell Biol. 1995, 129, 1617–1628. [Google Scholar] [CrossRef]
- Roghi, C.; Giet, R.; Uzbekov, R.; Morin, N.; Chartrain, I.; Le Guellec, R.; Couturier, A.; Doree, M.; Philippe, M.; Prigent, C. The Xenopus protein kinase pEg2 associates with the centrosome in a cell cycle-dependent manner, binds to the spindle microtubules and is involved in bipolar mitotic spindle assembly. J. Cell Sci. 1998, 111, 557–572. [Google Scholar]
- Wang, G.; Jiang, Q.; Zhang, C. The role of mitotic kinases in coupling the centrosome cycle with the assembly of the mitotic spindle. J. Cell Sci. 2014, 127, 4111–4122. [Google Scholar] [CrossRef]
- Fry, A.M.; Sampson, J.; Shak, C.; Shackleton, S. Recent advances in pericentriolar material organization: Ordered layers and scaffolding gels. F1000 Res. 2017, 6, 1622. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Scholey, J.M. Assembly, Functions and Evolution of Archaella, Flagella and Cilia. Curr. Biol. 2018, 28, R278–R292. [Google Scholar] [CrossRef]
- Mitchell, D.R. The evolution of eukaryotic cilia and flagella as motile and sensory organelles. Adv. Exp. Med. Biol. 2007, 607, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Mitchison, H.M.; Valente, E.M. Motile and non-motile cilia in human pathology: From function to phenotypes. J. Pathol. 2017, 241, 294–309. [Google Scholar] [CrossRef]
- Nachury, M.V. How do cilia organize signalling cascades? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef]
- Nachury, M.V.; Mick, D.U. Establishing and regulating the composition of cilia for signal transduction. Nat. Rev. Mol. Cell Biol. 2019. [Google Scholar] [CrossRef]
- Reiter, J.F.; Leroux, M.R. Genes and molecular pathways underpinning ciliopathies. Nat. Rev. Mol. Cell Biol. 2017, 18, 533–547. [Google Scholar] [CrossRef]
- Mirvis, M.; Stearns, T.; James Nelson, W. Cilium structure, assembly, and disassembly regulated by the cytoskeleton. Biochem. J. 2018, 475, 2329–2353. [Google Scholar] [CrossRef]
- Pedersen, L.B.; Schroder, J.M.; Satir, P.; Christensen, S.T. The ciliary cytoskeleton. Compr. Physiol. 2012, 2, 779–803. [Google Scholar] [CrossRef]
- Kiesel, P.; Gonzalo, A.V.; Tsoy, T.; Maraspini, R.; Honigmann, A.; Gigino, G. The molecular structure of primary cilia revealed by cryo-electron tomography. bioRxiv 2020. [Google Scholar] [CrossRef]
- Sun, S.; Fisher, R.L.; Bowser, S.S.; Pentecost, B.T.; Sui, H. Three-dimensional architecture of epithelial primary cilia. Proc. Natl. Acad. Sci. USA 2019, 116, 9370–9379. [Google Scholar] [CrossRef]
- Soares, H.; Carmona, B.; Nolasco, S.; Viseu Melo, L.; Goncalves, J. Cilia Distal Domain: Diversity in Evolutionarily Conserved Structures. Cells 2019, 8, 160. [Google Scholar] [CrossRef]
- Nachury, M.V. The molecular machines that traffic signaling receptors into and out of cilia. Curr. Opin. Cell Biol. 2018, 51, 124–131. [Google Scholar] [CrossRef]
- Nakayama, K.; Katoh, Y. Ciliary protein trafficking mediated by IFT and BBSome complexes with the aid of kinesin-2 and dynein-2 motors. J. Biochem. 2018, 163, 155–164. [Google Scholar] [CrossRef]
- Lechtreck, K.F. IFT-Cargo Interactions and Protein Transport in Cilia. Trends Biochem. Sci. 2015, 40, 765–778. [Google Scholar] [CrossRef]
- Ye, F.; Nager, A.R.; Nachury, M.V. BBSome trains remove activated GPCRs from cilia by enabling passage through the transition zone. J. Cell Biol. 2018. [Google Scholar] [CrossRef]
- Garcia-Gonzalo, F.R.; Corbit, K.C.; Sirerol-Piquer, M.S.; Ramaswami, G.; Otto, E.A.; Noriega, T.R.; Seol, A.D.; Robinson, J.F.; Bennett, C.L.; Josifova, D.J.; et al. A transition zone complex regulates mammalian ciliogenesis and ciliary membrane composition. Nat. Genet. 2011, 43, 776–784. [Google Scholar] [CrossRef]
- Garcia-Gonzalo, F.R.; Reiter, J.F. Open Sesame: How Transition Fibers and the Transition Zone Control Ciliary Composition. Cold Spring Harb. Perspect. Biol. 2017, 9. [Google Scholar] [CrossRef]
- Goncalves, J.; Pelletier, L. The Ciliary Transition Zone: Finding the Pieces and Assembling the Gate. Mol. Cells 2017, 40, 243–253. [Google Scholar] [CrossRef]
- Sanchez, I.; Dynlacht, B.D. Cilium assembly and disassembly. Nat. Cell Biol. 2016, 18, 711–717. [Google Scholar] [CrossRef]
- Sorokin, S. Centrioles and the formation of rudimentary cilia by fibroblasts and smooth muscle cells. J. Cell Biol. 1962, 15, 363–377. [Google Scholar] [CrossRef]
- Sorokin, S.P. Reconstructions of centriole formation and ciliogenesis in mammalian lungs. J. Cell Sci. 1968, 3, 207–230. [Google Scholar]
- Molla-Herman, A.; Ghossoub, R.; Blisnick, T.; Meunier, A.; Serres, C.; Silbermann, F.; Emmerson, C.; Romeo, K.; Bourdoncle, P.; Schmitt, A.; et al. The ciliary pocket: An endocytic membrane domain at the base of primary and motile cilia. J. Cell Sci. 2010, 123, 1785–1795. [Google Scholar] [CrossRef]
- Wu, C.T.; Chen, H.Y.; Tang, T.K. Myosin-Va is required for preciliary vesicle transportation to the mother centriole during ciliogenesis. Nat. Cell Biol. 2018, 20, 175–185. [Google Scholar] [CrossRef]
- Ghossoub, R.; Molla-Herman, A.; Bastin, P.; Benmerah, A. The ciliary pocket: A once-forgotten membrane domain at the base of cilia. Biol. Cell 2011, 103, 131–144. [Google Scholar] [CrossRef]
- Pugacheva, E.N.; Jablonski, S.A.; Hartman, T.R.; Henske, E.P.; Golemis, E.A. HEF1-dependent Aurora A activation induces disassembly of the primary cilium. Cell 2007, 129, 1351–1363. [Google Scholar] [CrossRef]
- Ford, M.J.; Yeyati, P.L.; Mali, G.R.; Keighren, M.A.; Waddell, S.H.; Mjoseng, H.K.; Douglas, A.T.; Hall, E.A.; Sakaue-Sawano, A.; Miyawaki, A.; et al. A Cell/Cilia Cycle Biosensor for Single-Cell Kinetics Reveals Persistence of Cilia after G1/S Transition Is a General Property in Cells and Mice. Dev. Cell 2018, 47, 509–523. [Google Scholar] [CrossRef]
- Liang, Y.; Meng, D.; Zhu, B.; Pan, J. Mechanism of ciliary disassembly. Cell Mol. Life Sci. 2016, 73, 1787–1802. [Google Scholar] [CrossRef]
- Reilly, M.L.; Benmerah, A. Ciliary kinesins beyond IFT: Cilium length, disassembly, cargo transport and signalling. Biol. Cell 2019, 111, 79–94. [Google Scholar] [CrossRef]
- Wang, L.; Dynlacht, B.D. The regulation of cilium assembly and disassembly in development and disease. Development 2018, 145. [Google Scholar] [CrossRef]
- Mirvis, M.; Siemers, K.A.; Nelson, W.J.; Stearns, T.P. Primary cilium loss in mammalian cells occurs predominantly by whole-cilium shedding. PLoS Biol. 2019, 17, e3000381. [Google Scholar] [CrossRef] [PubMed]
- Avasthi, P.; Marshall, W.F. Ciliary regulation: Disassembly takes the spotlight. Curr. Biol. 2013, 23, R1001–R1003. [Google Scholar] [CrossRef] [PubMed]
- Jang, C.Y.; Coppinger, J.A.; Seki, A.; Yates, J.R., 3rd; Fang, G. Plk1 and Aurora A regulate the depolymerase activity and the cellular localization of Kif2a. J. Cell Sci. 2009, 122, 1334–1341. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, K.; Choi, J.H.; Ringstad, N.; Dynlacht, B.D. Nek2 activation of Kif24 ensures cilium disassembly during the cell cycle. Nat. Commun. 2015, 6, 8087. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Johmura, Y.; Yu, L.R.; Park, J.E.; Gao, Y.; Bang, J.K.; Zhou, M.; Veenstra, T.D.; Yeon Kim, B.; Lee, K.S. Identification of a novel Wnt5a-CK1varepsilon-Dvl2-Plk1-mediated primary cilia disassembly pathway. EMBO J. 2012, 31, 3104–3117. [Google Scholar] [CrossRef]
- Plotnikova, O.V.; Nikonova, A.S.; Loskutov, Y.V.; Kozyulina, P.Y.; Pugacheva, E.N.; Golemis, E.A. Calmodulin activation of Aurora-A kinase (AURKA) is required during ciliary disassembly and in mitosis. Mol. Biol. Cell 2012, 23, 2658–2670. [Google Scholar] [CrossRef]
- Ran, J.; Yang, Y.; Li, D.; Liu, M.; Zhou, J. Deacetylation of alpha-tubulin and cortactin is required for HDAC6 to trigger ciliary disassembly. Sci. Rep. 2015, 5, 12917. [Google Scholar] [CrossRef]
- Wang, G.; Chen, Q.; Zhang, X.; Zhang, B.; Zhuo, X.; Liu, J.; Jiang, Q.; Zhang, C. PCM1 recruits Plk1 to the pericentriolar matrix to promote primary cilia disassembly before mitotic entry. J. Cell Sci. 2013, 126, 1355–1365. [Google Scholar] [CrossRef]
- Wang, W.; Wu, T.; Kirschner, M.W. The master cell cycle regulator APC-Cdc20 regulates ciliary length and disassembly of the primary cilium. eLife 2014, 3, e03083. [Google Scholar] [CrossRef]
- Conkar, D.; Bayraktar, H.; Firat-Karalar, E.N. Centrosomal and ciliary targeting of CCDC66 requires cooperative action of centriolar satellites, microtubules and molecular motors. Sci. Rep. 2019, 9, 14250. [Google Scholar] [CrossRef]
- Kubo, A.; Sasaki, H.; Yuba-Kubo, A.; Tsukita, S.; Shiina, N. Centriolar satellites: Molecular characterization, ATP-dependent movement toward centrioles and possible involvement in ciliogenesis. J. Cell Biol. 1999, 147, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Gheiratmand, L.; Coyaud, E.; Gupta, G.D.; Laurent, E.M.; Hasegan, M.; Prosser, S.L.; Goncalves, J.; Raught, B.; Pelletier, L. Spatial and proteomic profiling reveals centrosome-independent features of centriolar satellites. EMBO J. 2019. [Google Scholar] [CrossRef] [PubMed]
- Quarantotti, V.; Chen, J.X.; Tischer, J.; Gonzalez Tejedo, C.; Papachristou, E.K.; D’Santos, C.S.; Kilmartin, J.V.; Miller, M.L.; Gergely, F. Centriolar satellites are acentriolar assemblies of centrosomal proteins. EMBO J. 2019. [Google Scholar] [CrossRef] [PubMed]
- Aydin, O.A.; Taflan, S.O.; Gurkaslar, C.; Firat-Karalar, E.N. Acute inhibition of centriolar satellite function and positioning reveals their functions at the primary cilium. bioRxiv 2020. [Google Scholar] [CrossRef]
- Odabasi, E.; Gul, S.; Kavakli, I.H.; Firat-Karalar, E.N. Centriolar satellites are required for efficient ciliogenesis and ciliary content regulation. EMBO Rep. 2019. [Google Scholar] [CrossRef]
- Wang, L.; Lee, K.; Malonis, R.; Sanchez, I.; Dynlacht, B.D. Tethering of an E3 ligase by PCM1 regulates the abundance of centrosomal KIAA0586/Talpid3 and promotes ciliogenesis. eLife 2016, 5. [Google Scholar] [CrossRef]
- Kubo, A.; Tsukita, S. Non-membranous granular organelle consisting of PCM-1: Subcellular distribution and cell-cycle-dependent assembly/disassembly. J. Cell Sci. 2003, 116, 919–928. [Google Scholar] [CrossRef]
- Rai, A.K.; Chen, J.X.; Selbach, M.; Pelkmans, L. Kinase-controlled phase transition of membraneless organelles in mitosis. Nature 2018, 559, 211–216. [Google Scholar] [CrossRef]
- Fais, D.A.; Nadezhdina, E.S.; Chentsov, Y.S. The centriolar rim. The structure that maintains the configuration of centrioles and basal bodies in the absence of their microtubules. Exp. Cell Res. 1986, 164, 27–34. [Google Scholar] [CrossRef]
- Klotz, C.; Dabauvalle, M.C.; Paintrand, M.; Weber, T.; Bornens, M.; Karsenti, E. Parthenogenesis in Xenopus eggs requires centrosomal integrity. J. Cell Biol. 1990, 110, 405–415. [Google Scholar] [CrossRef]
- Schnackenberg, B.J.; Palazzo, R.E. Identification and function of the centrosome centromatrix. Biol. Cell 1999, 91, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Chakraborti, S.; Natarajan, K.; Curiel, J.; Janke, C.; Liu, J. The emerging role of the tubulin code: From the tubulin molecule to neuronal function and disease. Cytoskeleton 2016, 73, 521–550. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, H.G.; Abia, D.; Janowski, R.; Mortuza, G.; Bertero, M.G.; Boutin, M.; Guarin, N.; Mendez-Giraldez, R.; Nunez, A.; Pedrero, J.G.; et al. Structure and non-structure of centrosomal proteins. PLoS ONE 2013, 8, e62633. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, M.; Hyman, A.A.; Beyer, A. Coiled-coil proteins facilitated the functional expansion of the centrosome. PLoS Comput. Biol. 2014, 10, e1003657. [Google Scholar] [CrossRef] [PubMed]
- Bayless, B.A.; Navarro, F.M.; Winey, M. Motile Cilia: Innovation and Insight From Ciliate Model Organisms. Front. Cell Dev. Biol. 2019, 7, 265. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, K.; Bustamante-Marin, X.; Yin, W.; Goshe, M.B.; Ostrowski, L.E. Quantitative Proteomic Analysis of Human Airway Cilia Identifies Previously Uncharacterized Proteins of High Abundance. J. Proteome Res. 2017, 16, 1579–1592. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, L.E.; Blackburn, K.; Radde, K.M.; Moyer, M.B.; Schlatzer, D.M.; Moseley, A.; Boucher, R.C. A proteomic analysis of human cilia: Identification of novel components. Mol. Cell Proteom. 2002, 1, 451–465. [Google Scholar] [CrossRef]
- Sigg, M.A.; Menchen, T.; Lee, C.; Johnson, J.; Jungnickel, M.K.; Choksi, S.P.; Garcia, G., 3rd; Busengdal, H.; Dougherty, G.W.; Pennekamp, P.; et al. Evolutionary Proteomics Uncovers Ancient Associations of Cilia with Signaling Pathways. Dev. Cell 2017, 43, 744–762. [Google Scholar] [CrossRef]
- Zhao, L.; Hou, Y.; Picariello, T.; Craige, B.; Witman, G.B. Proteome of the central apparatus of a ciliary axoneme. J. Cell Biol. 2019, 218, 2051–2070. [Google Scholar] [CrossRef]
- Nigg, E.A.; Holland, A.J. Once and only once: Mechanisms of centriole duplication and their deregulation in disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 297–312. [Google Scholar] [CrossRef]
- Berggard, T.; Linse, S.; James, P. Methods for the detection and analysis of protein-protein interactions. Proteomics 2007, 7, 2833–2842. [Google Scholar] [CrossRef] [PubMed]
- Keskin, O.; Tuncbag, N.; Gursoy, A. Predicting Protein-Protein Interactions from the Molecular to the Proteome Level. Chem. Rev. 2016, 116, 4884–4909. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Wang, J.; Peng, W.; Wu, F.X.; Pan, Y. Protein-protein interactions: Detection, reliability assessment and applications. Brief. Bioinform. 2017, 18, 798–819. [Google Scholar] [CrossRef] [PubMed]
- Podobnik, M.; Krasevec, N.; Bedina Zavec, A.; Naneh, O.; Flasker, A.; Caserman, S.; Hodnik, V.; Anderluh, G. How to Study Protein-protein Interactions. Acta Chim. Slov. 2016, 63, 424–439. [Google Scholar] [CrossRef] [PubMed]
- Stynen, B.; Tournu, H.; Tavernier, J.; Van Dijck, P. Diversity in genetic in vivo methods for protein-protein interaction studies: From the yeast two-hybrid system to the mammalian split-luciferase system. Microbiol. Mol. Biol. Rev. 2012, 76, 331–382. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Thompson, J.; Yates, J.R., 3rd; Marshall, W.F. Proteomic analysis of mammalian primary cilia. Curr. Biol. 2012, 22, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, L.; Vanselow, K.; Skogs, M.; Toyoda, Y.; Lundberg, E.; Poser, I.; Falkenby, L.G.; Bennetzen, M.; Westendorf, J.; Nigg, E.A.; et al. Novel asymmetrically localizing components of human centrosomes identified by complementary proteomics methods. EMBO J. 2011, 30, 1520–1535. [Google Scholar] [CrossRef]
- Gentzel, M.; Pardo, M.; Subramaniam, S.; Stewart, A.F.; Choudhary, J.S. Proteomic navigation using proximity-labeling. Methods 2019, 164, 67–72. [Google Scholar] [CrossRef]
- Gingras, A.C.; Abe, K.T.; Raught, B. Getting to know the neighborhood: Using proximity-dependent biotinylation to characterize protein complexes and map organelles. Curr. Opin. Chem. Biol. 2019, 48, 44–54. [Google Scholar] [CrossRef]
- Trinkle-Mulcahy, L. Recent advances in proximity-based labeling methods for interactome mapping. F1000 Res. 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Roux, K.J.; Kim, D.I.; Raida, M.; Burke, B. A promiscuous biotin ligase fusion protein identifies proximal and interacting proteins in mammalian cells. J. Cell Biol. 2012, 196, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.I.; Birendra, K.C.; Zhu, W.; Motamedchaboki, K.; Doye, V.; Roux, K.J. Probing nuclear pore complex architecture with proximity-dependent biotinylation. Proc. Natl. Acad. Sci. USA 2014, 111, E2453–E2461. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.I.; Jensen, S.C.; Noble, K.A.; Kc, B.; Roux, K.H.; Motamedchaboki, K.; Roux, K.J. An improved smaller biotin ligase for BioID proximity labeling. Mol. Biol. Cell 2016, 27, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Branon, T.C.; Bosch, J.A.; Sanchez, A.D.; Udeshi, N.D.; Svinkina, T.; Carr, S.A.; Feldman, J.L.; Perrimon, N.; Ting, A.Y. Efficient proximity labeling in living cells and organisms with TurboID. Nat. Biotechnol. 2018, 36, 880–887. [Google Scholar] [CrossRef]
- Nguyen, T.M.T.; Kim, J.; Doan, T.T.; Lee, M.W.; Lee, M. APEX Proximity Labeling as a Versatile Tool for Biological Research. Biochemistry 2020, 59, 260–269. [Google Scholar] [CrossRef]
- Cho, K.F.; Tess, C.; Branon, S.R.; Svinkina, T.; Namrata, D.U.; Thoudam, T.; Kwak, C.; Rhee, H.W.; Lee, I.; Carr, S.A.; et al. Split-TurboID enables contact-dependent proximity labeling in cells. bioRxiv 2020. [Google Scholar] [CrossRef]
- De Munter, S.; Gornemann, J.; Derua, R.; Lesage, B.; Qian, J.; Heroes, E.; Waelkens, E.; Van Eynde, A.; Beullens, M.; Bollen, M. Split-BioID: A proximity biotinylation assay for dimerization-dependent protein interactions. FEBS Lett. 2017, 591, 415–424. [Google Scholar] [CrossRef]
- Schopp, I.M.; Amaya Ramirez, C.C.; Debeljak, J.; Kreibich, E.; Skribbe, M.; Wild, K.; Bethune, J. Split-BioID a conditional proteomics approach to monitor the composition of spatiotemporally defined protein complexes. Nat. Commun. 2017, 8, 15690. [Google Scholar] [CrossRef]
- Xue, M.; Hou, J.; Wang, L.; Cheng, D.; Lu, J.; Zheng, L.; Xu, T. Optimizing the fragment complementation of APEX2 for detection of specific protein-protein interactions in live cells. Sci. Rep. 2017, 7, 12039. [Google Scholar] [CrossRef]
- Lambert, J.P.; Tucholska, M.; Go, C.; Knight, J.D.; Gingras, A.C. Proximity biotinylation and affinity purification are complementary approaches for the interactome mapping of chromatin-associated protein complexes. J. Proteom. 2015, 118, 81–94. [Google Scholar] [CrossRef]
- He, K.; Ma, X.; Xu, T.; Li, Y.; Hodge, A.; Zhang, Q.; Torline, J.; Huang, Y.; Zhao, J.; Ling, K.; et al. Axoneme polyglutamylation regulated by Joubert syndrome protein ARL13B controls ciliary targeting of signaling molecules. Nat. Commun. 2018, 9, 3310. [Google Scholar] [CrossRef] [PubMed]
- Mick, D.U.; Rodrigues, R.B.; Leib, R.D.; Adams, C.M.; Chien, A.S.; Gygi, S.P.; Nachury, M.V. Proteomics of Primary Cilia by Proximity Labeling. Dev. Cell 2015, 35, 497–512. [Google Scholar] [CrossRef] [PubMed]
- Kohli, P.; Hohne, M.; Jungst, C.; Bertsch, S.; Ebert, L.K.; Schauss, A.C.; Benzing, T.; Rinschen, M.M.; Schermer, B. The ciliary membrane-associated proteome reveals actin-binding proteins as key components of cilia. EMBO Rep. 2017, 18, 1521–1535. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.D.; Coyaud, E.; Goncalves, J.; Mojarad, B.A.; Liu, Y.; Wu, Q.; Gheiratmand, L.; Comartin, D.; Tkach, J.M.; Cheung, S.W.; et al. A Dynamic Protein Interaction Landscape of the Human Centrosome-Cilium Interface. Cell 2015, 163, 1484–1499. [Google Scholar] [CrossRef] [PubMed]
- Firat-Karalar, E.N.; Rauniyar, N.; Yates, J.R., 3rd; Stearns, T. Proximity interactions among centrosome components identify regulators of centriole duplication. Curr. Biol. 2014, 24, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Gurkaslar, H.K.; Culfa, E.; Arslanhan, M.D.; Lince-Faria, M.; Firat-Karalar, E.N. CCDC57 Cooperates with Microtubules and Microcephaly Protein CEP63 and Regulates Centriole Duplication and Mitotic Progression. Cell Rep. 2020, 31, 107630. [Google Scholar] [CrossRef]
- Barenz, F.; Kschonsak, Y.T.; Meyer, A.; Jafarpour, A.; Lorenz, H.; Hoffmann, I. Ccdc61 controls centrosomal localization of Cep170 and is required for spindle assembly and symmetry. Mol. Biol. Cell 2018. [Google Scholar] [CrossRef]
- Conkar, D.; Culfa, E.; Odabasi, E.; Rauniyar, N.; Yates, J.R., 3rd; Firat-Karalar, E.N. Centriolar satellite protein CCDC66 interacts with CEP290 and functions in cilium formation and trafficking. J. Cell Sci. 2017. [Google Scholar] [CrossRef]
- Uddin, B.; Partscht, P.; Chen, N.P.; Neuner, A.; Weiss, M.; Hardt, R.; Jafarpour, A.; Hessling, B.; Ruppert, T.; Lorenz, H.; et al. The human phosphatase CDC14A modulates primary cilium length by regulating centrosomal actin nucleation. EMBO Rep. 2019, 20. [Google Scholar] [CrossRef]
- Firat-Karalar, E.N.; Stearns, T. Probing mammalian centrosome structure using BioID proximity-dependent biotinylation. Methods Cell Biol. 2015, 129, 153–170. [Google Scholar] [CrossRef]
- Joseph, N.; Al-Jassar, C.; Johnson, C.M.; Andreeva, A.; Barnabas, D.D.; Freund, S.M.V.; Gergely, F.; van Breugel, M. Disease-Associated Mutations in CEP120 Destabilize the Protein and Impair Ciliogenesis. Cell Rep. 2018, 23, 2805–2818. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gupta, G.D.; Barnabas, D.D.; Agircan, F.G.; Mehmood, S.; Wu, D.; Coyaud, E.; Johnson, C.M.; McLaughlin, S.H.; Andreeva, A.; et al. Direct binding of CEP85 to STIL ensures robust PLK4 activation and efficient centriole assembly. Nat. Commun. 2018, 9, 1731. [Google Scholar] [CrossRef] [PubMed]
- Bozal-Basterra, L.; Gonzalez-Santamarta, M.; Bermejo-Arteagabeitia, A.; Da Fonseca, C.; Pampliega, O.; Andrade, R.; Martin-Martin, N.; Branon, T.C.; Ting, A.Y.; Carracedo, A.; et al. LUZP1, a novel regulator of primary cilia and the actin cytoskeleton, is altered in Townes-Brocks Syndrome. bioRxiv 2019. [Google Scholar] [CrossRef]
- Goncalves, J.; Coyaud, E.; Laurent, E.M.N.; Raught, B.; Pelletier, L. LUZP1 and the tumour suppressor EPLIN are negative regulators of primary cilia formation. bioRxiv 2019. [Google Scholar] [CrossRef]
- Dho, S.E.; Silva-Gagliardi, N.; Morgese, F.; Coyaud, E.; Lamoureux, E.; Berry, D.M.; Raught, B.; McGlade, C.J. Proximity interactions of the ubiquitin ligase Mind bomb 1 reveal a role in regulation of epithelial polarity complex proteins. Sci. Rep. 2019, 9, 12471. [Google Scholar] [CrossRef] [PubMed]
- Kazazian, K.; Go, C.; Wu, H.; Brashavitskaya, O.; Xu, R.; Dennis, J.W.; Gingras, A.C.; Swallow, C.J. Plk4 Promotes Cancer Invasion and Metastasis through Arp2/3 Complex Regulation of the Actin Cytoskeleton. Cancer Res. 2017, 77, 434–447. [Google Scholar] [CrossRef]
- Sydor, A.M.; Coyaud, E.; Rovelli, C.; Laurent, E.; Liu, H.; Raught, B.; Mennella, V. PPP1R35 is a novel centrosomal protein that regulates centriole length in concert with the microcephaly protein RTTN. eLife 2018, 7. [Google Scholar] [CrossRef]
- Bozal-Basterra, L.; Martin-Ruiz, I.; Pirone, L.; Liang, Y.; Sigurethsson, J.O.; Gonzalez-Santamarta, M.; Giordano, I.; Gabicagogeascoa, E.; de Luca, A.; Rodriguez, J.A.; et al. Truncated SALL1 Impedes Primary Cilia Function in Townes-Brocks Syndrome. Am. J. Hum. Genet. 2018, 102, 249–265. [Google Scholar] [CrossRef]
- Agircan, F.G.; Hata, S.; Nussbaum-Krammer, C.; Atorino, E.; Schiebel, E. Proximity mapping of human separase by the BioID approach. Biochem. Biophys. Res. Commun. 2016, 478, 656–662. [Google Scholar] [CrossRef]
- Firat-Karalar, E.N. Proximity mapping of the microtubule plus-end tracking protein SLAIN2 using the BioID approach. Turk. J. Biol. 2020, 44, 61–72. [Google Scholar] [CrossRef]
- Godinho, S.A.; Pellman, D. Causes and consequences of centrosome abnormalities in cancer. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef] [PubMed]
- Arquint, C.; Nigg, E.A. The PLK4-STIL-SAS-6 module at the core of centriole duplication. Biochem. Soc. Trans. 2016, 44, 1253–1263. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.S.; Park, J.E.; Shukla, A.; Choi, S.; Murugan, R.N.; Lee, J.H.; Ahn, M.; Rhee, K.; Bang, J.K.; Kim, B.Y.; et al. Hierarchical recruitment of Plk4 and regulation of centriole biogenesis by two centrosomal scaffolds, Cep192 and Cep152. Proc. Natl. Acad. Sci. USA 2013, 110, E4849–E4857. [Google Scholar] [CrossRef] [PubMed]
- Sonnen, K.F.; Gabryjonczyk, A.M.; Anselm, E.; Stierhof, Y.D.; Nigg, E.A. Human Cep192 and Cep152 cooperate in Plk4 recruitment and centriole duplication. J. Cell Sci. 2013, 126, 3223–3233. [Google Scholar] [CrossRef] [PubMed]
- Guichard, P.; Hamel, V.; Gonczy, P. The Rise of the Cartwheel: Seeding the Centriole Organelle. Bioessays 2018, 40, e1700241. [Google Scholar] [CrossRef]
- Arquint, C.; Gabryjonczyk, A.M.; Imseng, S.; Bohm, R.; Sauer, E.; Hiller, S.; Nigg, E.A.; Maier, T. STIL binding to Polo-box 3 of PLK4 regulates centriole duplication. eLife 2015, 4. [Google Scholar] [CrossRef]
- Vulprecht, J.; David, A.; Tibelius, A.; Castiel, A.; Konotop, G.; Liu, F.; Bestvater, F.; Raab, M.S.; Zentgraf, H.; Izraeli, S.; et al. STIL is required for centriole duplication in human cells. J. Cell Sci. 2012, 125, 1353–1362. [Google Scholar] [CrossRef]
- Kodani, A.; Yu, T.W.; Johnson, J.R.; Jayaraman, D.; Johnson, T.L.; Al-Gazali, L.; Sztriha, L.; Partlow, J.N.; Kim, H.; Krup, A.L.; et al. Centriolar satellites assemble centrosomal microcephaly proteins to recruit CDK2 and promote centriole duplication. eLife 2015, 4. [Google Scholar] [CrossRef]
- Comartin, D.; Gupta, G.D.; Fussner, E.; Coyaud, E.; Hasegan, M.; Archinti, M.; Cheung, S.W.; Pinchev, D.; Lawo, S.; Raught, B.; et al. CEP120 and SPICE1 cooperate with CPAP in centriole elongation. Curr. Biol. 2013, 23, 1360–1366. [Google Scholar] [CrossRef]
- Al-Jassar, C.; Andreeva, A.; Barnabas, D.D.; McLaughlin, S.H.; Johnson, C.M.; Yu, M.; van Breugel, M. The Ciliopathy-Associated Cep104 Protein Interacts with Tubulin and Nek1 Kinase. Structure 2017, 25, 146–156. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Tag | Wild Type Enzymes | Source | Molecular Weight | Substrate | Half life of Radicals | Biotinylation Time | |
|---|---|---|---|---|---|---|---|
| Conventional Biotinylation | BioID | Biotin Ligase | E. coli | 35 kDa | Biotin | Minutes | 18–24 h |
| BioID2 | Biotin Ligase | A. Aoelicus | 26 kDa | Biotin | Minutes | 18–24 h | |
| TurboID | Biotin Ligase | E. coli | 35 kDa | Biotin-phenol | Minutes | 10 min | |
| miniTurboID | Biotin Ligase | E. coli | 28 kDa | Biotin | Minutes | 10 min | |
| APEX | Ascorbate peroxidase | Soybean | 27 kDa | Biotin-phenol | <1 ms | 1 min | |
| APEX2 | Ascorbate peroxidase | Soybean | 27 kDa | Biotin-phenol | <1 ms | 1 min | |
| Split Biotinylation | Split-BioID | Biotin Ligase | E. coli | 35 kDa | Biotin | Minutes | 18–24 h |
| Split-TurboID | Biotin Ligase | E. coli | 35 kDa | Biotin | Minutes | 10 minutes | |
| Split-APEX | Ascorbate peroxidase | Soybean | 27 kDa | Biotin-phenol | <1 ms | 1 min |
| Tag | Protein Name | Function | Localization | Cell Line | Reference | ||||
|---|---|---|---|---|---|---|---|---|---|
| Centrosome | CS | Cilia | Spindle | Nucleus | |||||
| APEX | ARL13B | GTP binding/Ciliogenesis | - | - | + | - | - | RCTE | He et al., 2018 [122] |
| NPHP3(1–203) | Ciliary membrane receptor | - | - | + | - | - | IMCD3 | Mick et al., 2015 [123] | |
| APEX2 | HTR6 | Ciliary membrane receptor | - | - | + | - | - | IMCD3 NIH3T3 | Kohli et al., 2017 [124] |
| BirA | AHI1 | Ciliogenesis and vesicle trafficking | + | - | + | - | - | HEK293 | Gupta et al., 2015 [125] |
| B9D1 | Transition zone component/Hedgehog signalling | - | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| B9D2 | Transition zone component/Ciliogenesis | - | - | + | - | + | HEK293 | Gupta et al., 2015 [125] | |
| BBS4 | Ciliogenesis and ciliary sorting | + | + | + | - | - | HEK293 | Gheiratmand et al., 2019 [83] | |
| C11orf49 | Unknown | n/a | + | n/a | n/a | n/a | HEK293 | Gheiratmand et al., 2019 [83] | |
| CC2D2A | Transition zone component/Hedgehog signalling | - | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| CCDC11 | Ciliary Beating/Ciliogenesis | - | + | + | - | - | HEK293 | Gheiratmand et al., 2019 [83] | |
| CCDC112 | Unknown | + | HEK293 | Gheiratmand et al., 2019 [83] | |||||
| CCDC13 | Primary cilium formation/Localization of BBS4 to cilia | - | + | + | - | - | HEK293 | Gheiratmand et al., 2019 [83] | |
| CCDC14 | Centriole duplication | - | + | - | - | - | HEK293 | Fırat-Karalar et al., 2014 [126] Gheiratmand et al., 2019 [83] | |
| CCDC18 | Unknown | n/a | + | n/a | n/a | n/a | HEK293 | Gheiratmand et al., 2019 [83] | |
| CCDC57 | Centriole duplication | + | + | - | + | - | HEK293 | Gurkaslar et al., 2020 [127] | |
| CCDC61 | Spindle assembly | + | - | - | - | - | HEK293 | Barenz et al., 2018 [128] | |
| CCDC66 | Ciliogenesis/Retina morphogenesis and homeostasis | + | + | + | - | - | HEK293 | Conkar et al., 2017 [129] Gheiratmand et al., 2019 [83] | |
| CCP110 | Centriole length regulation/Negative regulator of ciliogenesis | + | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| CDC14A | Cilium length regulation | + | - | - | + | - | RPE1 | Uddin et al., 2019 [130] | |
| CENPJ/CPAP | Centriole duplication/Centriole elongation | + | - | - | - | - | HEK293 | Fırat-Karalar et al., 2014 [126] Gupta et al., 2015 [125] | |
| CEP104 | Ciliogenesis and cilium length regulation | + | - | + | + | - | HEK293 | Gupta et al., 2015 [131] | |
| CEP120 | Centriole Duplication and Ciliogenesis | + | - | - | - | - | HEK293 | Gupta et al., 2015 [131] | |
| CEP120 (C2B) | Centriole Duplication and Ciliogenesis | + | - | - | - | - | HEK293 | Joseph et al., 2018 [132] | |
| CEP128 | Subdistal appendages/TGFbeta signaling | + | - | - | + | - | HEK293 | Gupta et al., 2015 [131] | |
| CEP131 | Ciliogenesis/Spindle pole integrity | + | + | + | - | - | HEK293 | Gheiratmand et al., 2019 [83] | |
| CEP135 | Centriole duplication/elongation | + | - | - | - | - | HEK293 | Gupta et al., 2015 [125] | |
| CEP152 | Centriole duplication | + | - | - | - | - | HEK293 | Fırat-Karalar et al., 2014 [126] Gupta et al., 2015 [125] Liu et al., 2018 [133] | |
| CEP162 | Transition zone assembly | + | - | + | + | + | HEK293 | Gupta et al., 2015 [125] | |
| CEP164 | Primary cilium docking | + | - | - | - | + | HEK293 | Gupta et al., 2015 [125] | |
| CEP170 | Subdistal appendage assembly | + | - | - | + | - | HEK293 | Gupta et al., 2015 [125] | |
| CEP19 | Recruitment of ciliary vesicles | + | - | + | + | - | HEK293 | Gupta et al., 2015 [125] | |
| CEP192 | Centriole duplication/centrosome maturation/spindle pole assembly | + | + | - | - | - | HEK293 | Fırat-Karalar et al., 2014 [126] Liu et al., 2018 [133] | |
| CEP290 | Transition zone component/early ciliogenesis/ciliary targeting of cargos | + | + | + | - | + | HEK293 | Gupta et al., 2015 [125] | |
| CEP44 | Centriole to centrosome conversion | + | - | - | + | - | HEK293 | Gupta et al., 2015 [125] | |
| CEP63 | Centriole duplication/Spindle assembly | + | + | - | - | - | HEK293 | Fırat-Karalar et al., 2014 [126] Gupta et al., 2015 [125] Liu et al., 2018 [133] Gheiratmand et al., 2019 [83] | |
| CEP72 | Centriole duplication/spindle pole assembly | + | + | - | - | - | HEK293 | Conkar et al., 2017 [129] Gheiratmand et al., 2019 [83] | |
| CEP83 | Distal appendages/Ciliogenesis | + | - | - | - | - | HEK293 | Gupta et al., 2015 [125] | |
| CEP85 | Negative regulator of centrosome integrity | + | - | - | + | + | HEK293 | Liu et al., 2018 [133] | |
| CEP89 | Ciliogenesis and regulation | + | - | - | + | - | HEK293 | Gupta et al., 2015 [125] | |
| CEP97 | Negative regulator of ciliogenesis | + | - | - | - | - | HEK293 | Gupta et al., 2015 [125] | |
| CETN2 | Centriole duplication | + | - | - | + | - | HEK293 | Gupta et al., 2015 [125] | |
| BirA | CNTRL | Cell cycle and cytokinesis | + | - | - | - | - | HEK293 | Gupta et al., 2015 [125] |
| CNTROB | Centriole Duplication | + | - | - | - | - | HEK293 | Gupta et al., 2015 [125] | |
| DCTN1 | Dynein-mediated transport | + | - | - | + | + | HEK293 | Gupta et al., 2015 [125] | |
| DYNLT1 | Retrograde transport | - | - | + | + | - | HEK293 | Gupta et al., 2015 [125] | |
| EVC2 | Hedgehog signalling | - | - | + | - | + | HEK293 | Gupta et al., 2015 [125] | |
| FBF1 | Transition fiber/Cilium content regulation | + | - | - | + | - | HEK293 | Gupta et al., 2015 [125] | |
| FGFR1OP | Cell cycle/Ciliogenesis | + | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| FOPNL | Cilium biogenesis/PLK1 recruitment to centrosome | + | + | + | - | - | HEK293 | Gheiratmand et al., 2019 [83] | |
| KIAA0753 | Centriole Duplication | + | + | - | - | - | HEK293 | Gupta et al., 2015 [125] | |
| LCA5 | Intraflagellar protein transport | + | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| LUZP1 | Actin-Stabilizing protein/Ciliogenesis/Hedgehog signaling | - | - | - | - | + | HEK293 | Bozal-Basterra et al., 2019 [134] Goncalves et al., 2019 [135] | |
| MIB1 | Ubiquitination/Ciliogenesis | - | + | - | - | - | HEK293 | Gheiratmand et al., 2019 [83] Dho et al., 2019 [136] | |
| MKS1 | Transition zone regulation | + | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| NEK8 | Protein targeting to cilium | + | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| NIN | Microtubule (-) end binding protein | + | - | - | - | - | HEK293 | Gupta et al., 2015 [125] | |
| NINL | Microtubule organization | + | - | - | - | - | HEK293 | Gupta et al., 2015 [125] | |
| NPHP1 | Spermatogenesis/Epithelial cell polarity | - | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| NPHP4 | Ciliary trafficking | + | - | + | - | + | HEK293 | Gupta et al., 2015 [125] | |
| ODF2 | Microtubule organization | + | - | + | + | - | HEK293 | Gupta et al., 2015 [125] | |
| OFD1 | Ciliogenesis/Centriole length control | + | + | + | - | + | HEK293 | Gupta et al., 2015 [125] Gheiratmand et al., 2019 [83] | |
| PCM1 | Molecular marker for centriolar satellites/Ciliogenesis/Hedgehog signaling | + | + | + | - | - | HEK293 | Gupta et al., 2015 [125] Gheiratmand et al., 2019 [83] Aydin et al., 2020 [85] | |
| PIBF1 | Ciliogenesis/Spindle pole integrity | + | + | - | - | + | HEK293 | Gheiratmand et al., 2019 [83] | |
| PLK4 | Centriole duplication/Cell migration | + | - | - | - | + | HEK293 | Fırat-Karalar et al., 2014 [126] Kazazian et al.,2016 [137] | |
| POC1A | Centriole stability | + | - | + | + | - | HEK293 | Gupta et al., 2015 [125] | |
| POC1B | Centriole stability | + | - | + | + | - | HEK293 | Gupta et al., 2015 [125] | |
| POC5 | Centriole stability | + | - | - | - | - | HEK293 | Gupta et al., 2015 [125] | |
| PPP1R35 | Centriole elongation | + | - | - | - | - | HEK293 | Sydor et al., 2018 [138] | |
| RPGR | Ciliogenesis/Photoreceptor integrity | + | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| RPGRIP1 | Photoreceptor morphogenesis/survival | - | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| RPGRIP1L | Transition zone regulation | + | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| SALL1 | Transcriptional repressor in organogenesis | - | - | - | - | + | HEK293 | Bozal-Basterra et al., 2018 [139] | |
| SASS6 | Cartwheel formation during procentriole assembly | + | - | - | - | - | HEK293 | Gupta et al., 2015 [125] Liu et al., 2018 [133] | |
| SCLT1 | Distal appendage assembly | + | - | - | - | - | HEK293 | Gupta et al., 2015 [125] | |
| SEPARASE | Centriole disengagement/Sister chromatid separation | + | - | - | - | + | U2OS | Agircan et al., 2016 [140] | |
| SLAIN2 | Microtubule (+) end binding | HEK293 | Fırat-Karalar, 2020 [141] | ||||||
| SPICE1 | Chromosome segregation/Centriole elongation | + | - | - | + | - | HEK293 | Gupta et al., 2015 [125] | |
| SSX2IP | Centrosome maturation | - | + | + | - | + | HEK293 | Gupta et al., 2015 [125] Gheiratmand et al., 2019 [83] | |
| STIL | Centriole duplication | + | - | - | - | - | HEK293 | Gupta et al., 2015 [125] Liu et al., 2018 [133] | |
| TBC1D31 | Unknown | + | + | - | - | - | HEK293 | Gheiratmand et al., 2019 [83] | |
| TCTN1 | Transition zone regulation | - | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| TCTN2 | Transition zone regulation | - | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| TCTN3 | Transition zone regulation | - | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| TEX9 | Ciliogenesis | + | HEK293 | Gheiratmand et al., 2019 [83] | |||||
| TMEM17 | Transition zone regulation | - | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| TMEM216 | Transition zone regulation | - | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| TMEM67 | Transition zone regulation | - | - | + | - | - | HEK293 | Gupta et al., 2015 [125] | |
| WRAP73 | Spindle pole anchoring | + | + | - | - | - | HEK293 | Gheiratmand et al., 2019 [83] | |
| Biological Process | New Findings | |
|---|---|---|
| Centrosome Biogenesis | Centriole Duplication: Initiation | Identification of new duplication factors including CEP85 and STIL |
| The regulatory role of centriolar satellites during initiation of centriole duplication | ||
| Interaction of CEP152 and CDK5RAP2 in PCM recruitment | ||
| Requiriement for MIB1 during centriole duplication | ||
| Centriole Duplication: Elongation | Identification of CEP120-SPICE1-CPAP functional complex | |
| Identification of PPP1R35 as a new centriole elongation factor | ||
| Centriole Disengament | Identification of Separase proximity interactors | |
| Actin Cytoskeleton | Phosphorylation of Arp2 by PLK4 for cancer cell movement | |
| Mitosis | Spindle Assembly | Interaction of CCDC61 and CEP170 in spindle formation |
| Primary Cilium Biogenesis | Cilium Assembly | CCDC66 and CEP290 interaction for cilium assembly |
| Involvement of CEP120 in cilium assembly | ||
| Role of actin stabilizing protein LUZP1 in cilium assembly | ||
| Cilium Length | Requirement of CEP104 and NEK1 interaction for cilium length regulation | |
| Cilia length regulation by CDC14A, which mediates centrosomal actin nucletion | ||
| Ciliary Functions | Rab11 and ARL13B interaction in axonemal glutamylation | |
| Requirement for TTLL5 and TTLL6 glutamylases | ||
| Identification of AC6/cAMP/PKA signaling module | ||
| Compartment Proteome | Centrosome | Identification of <1700 unique proteins in a netwrok of <7000 interactors |
| Centriolar Satellites | Highly overlapped interactome in centrosome and satellites | |
| Identification of 2113 high confidance interactors among 660 unique proteins | ||
| Cilium | Identification of over 200 proteins | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arslanhan, M.D.; Gulensoy, D.; Firat-Karalar, E.N. A Proximity Mapping Journey into the Biology of the Mammalian Centrosome/Cilium Complex. Cells 2020, 9, 1390. https://doi.org/10.3390/cells9061390
Arslanhan MD, Gulensoy D, Firat-Karalar EN. A Proximity Mapping Journey into the Biology of the Mammalian Centrosome/Cilium Complex. Cells. 2020; 9(6):1390. https://doi.org/10.3390/cells9061390
Chicago/Turabian StyleArslanhan, Melis Dilara, Dila Gulensoy, and Elif Nur Firat-Karalar. 2020. "A Proximity Mapping Journey into the Biology of the Mammalian Centrosome/Cilium Complex" Cells 9, no. 6: 1390. https://doi.org/10.3390/cells9061390
APA StyleArslanhan, M. D., Gulensoy, D., & Firat-Karalar, E. N. (2020). A Proximity Mapping Journey into the Biology of the Mammalian Centrosome/Cilium Complex. Cells, 9(6), 1390. https://doi.org/10.3390/cells9061390