IGFBP-3/IGFBP-3 Receptor System as an Anti-Tumor and Anti-Metastatic Signaling in Cancer


Abstract
1. Introduction
2. IGFBP-3
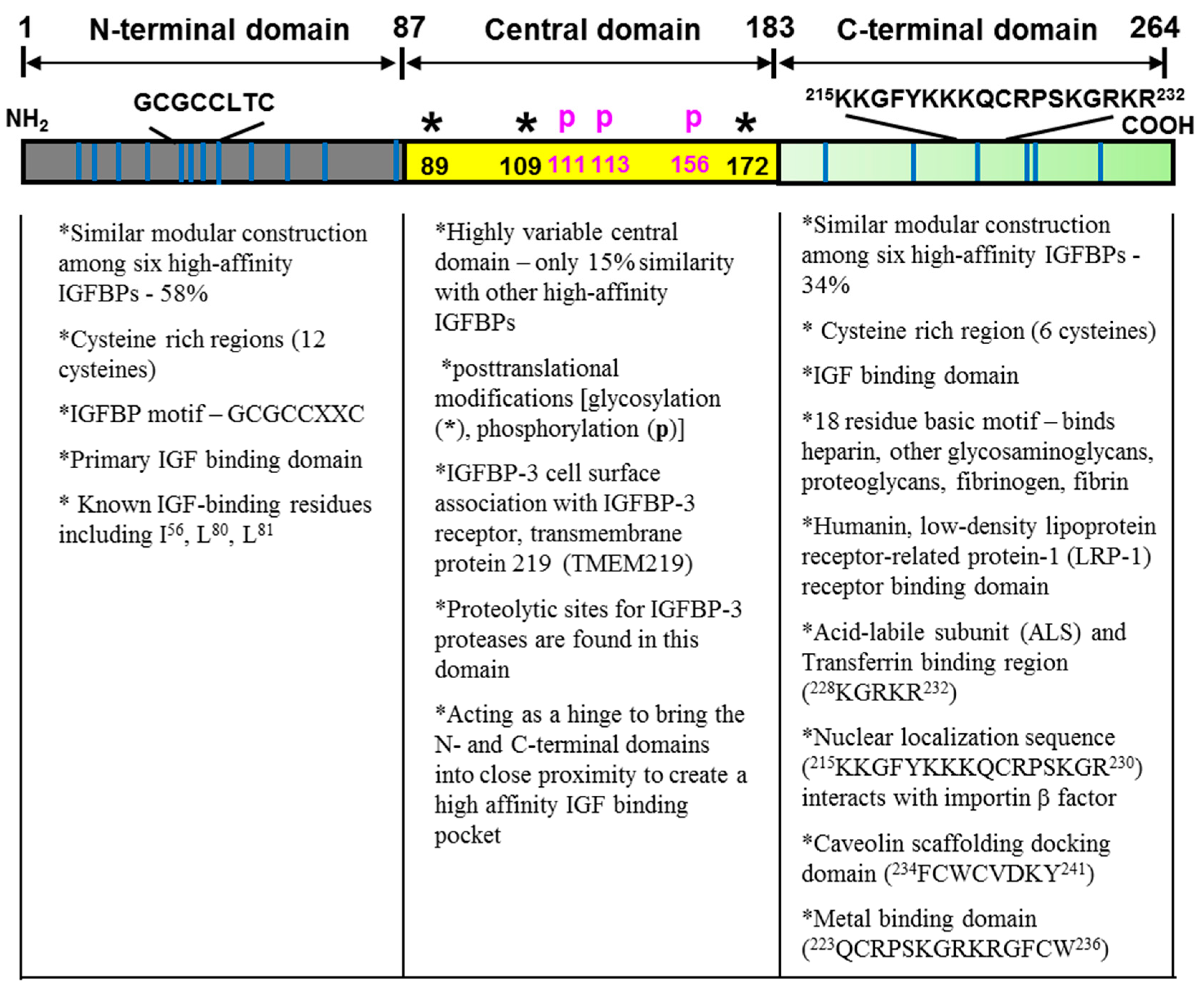
2.1. Structure-Function Analysis
2.1.1. The Conserved N-Terminal Domain
2.1.2. The Variable Central Domain
2.1.3. The Conserved C-Terminal Domain
2.2. IGF/IGF-IR Dependent Actions of IGFBP-3
2.3. IGF/IGF-IR Independent Actions of IGFBP-3
2.3.1. IGFBP-3 Binding Partners on the Cell Surface
2.3.2. IGFBP-3 Binding Partners within Cells
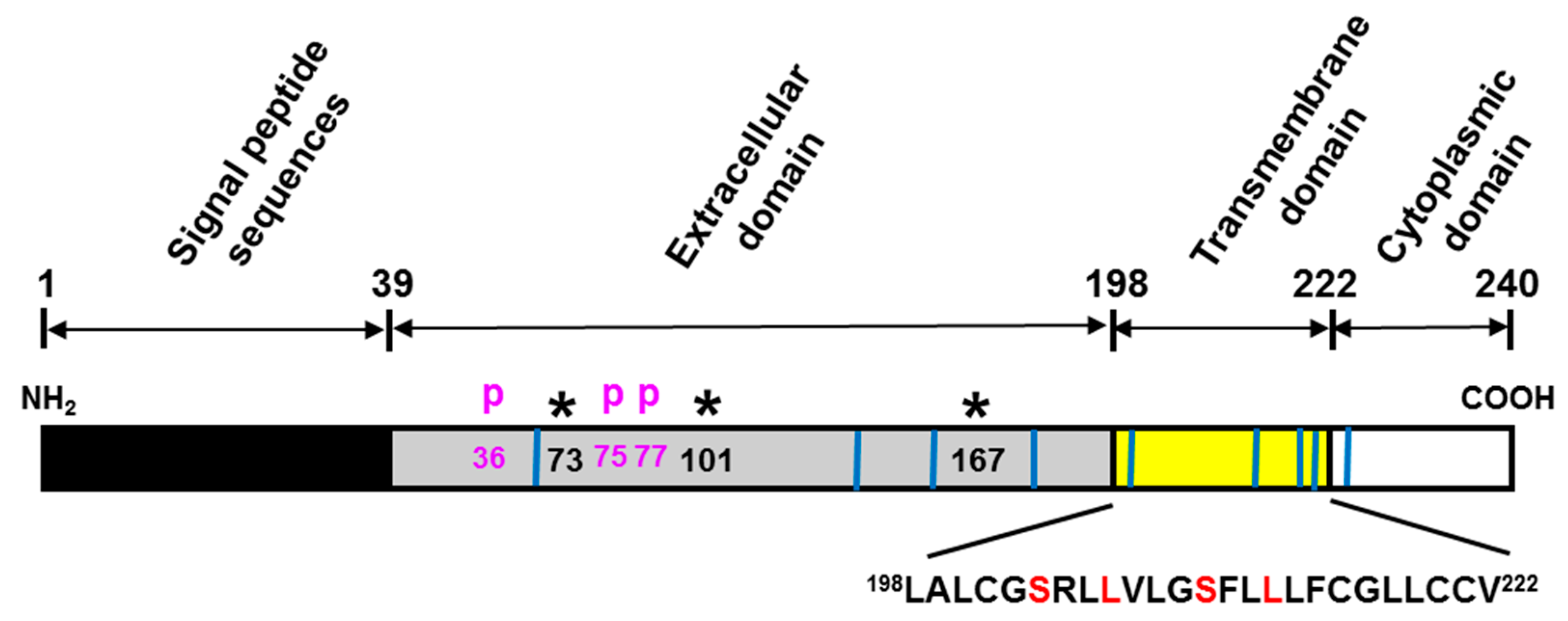
3. TMEM219 as an IGFBP-3 Specific Receptor
4. Clinical Insights of IGFBP-3/IGFBP-3R (TMEM219) System in Cancer
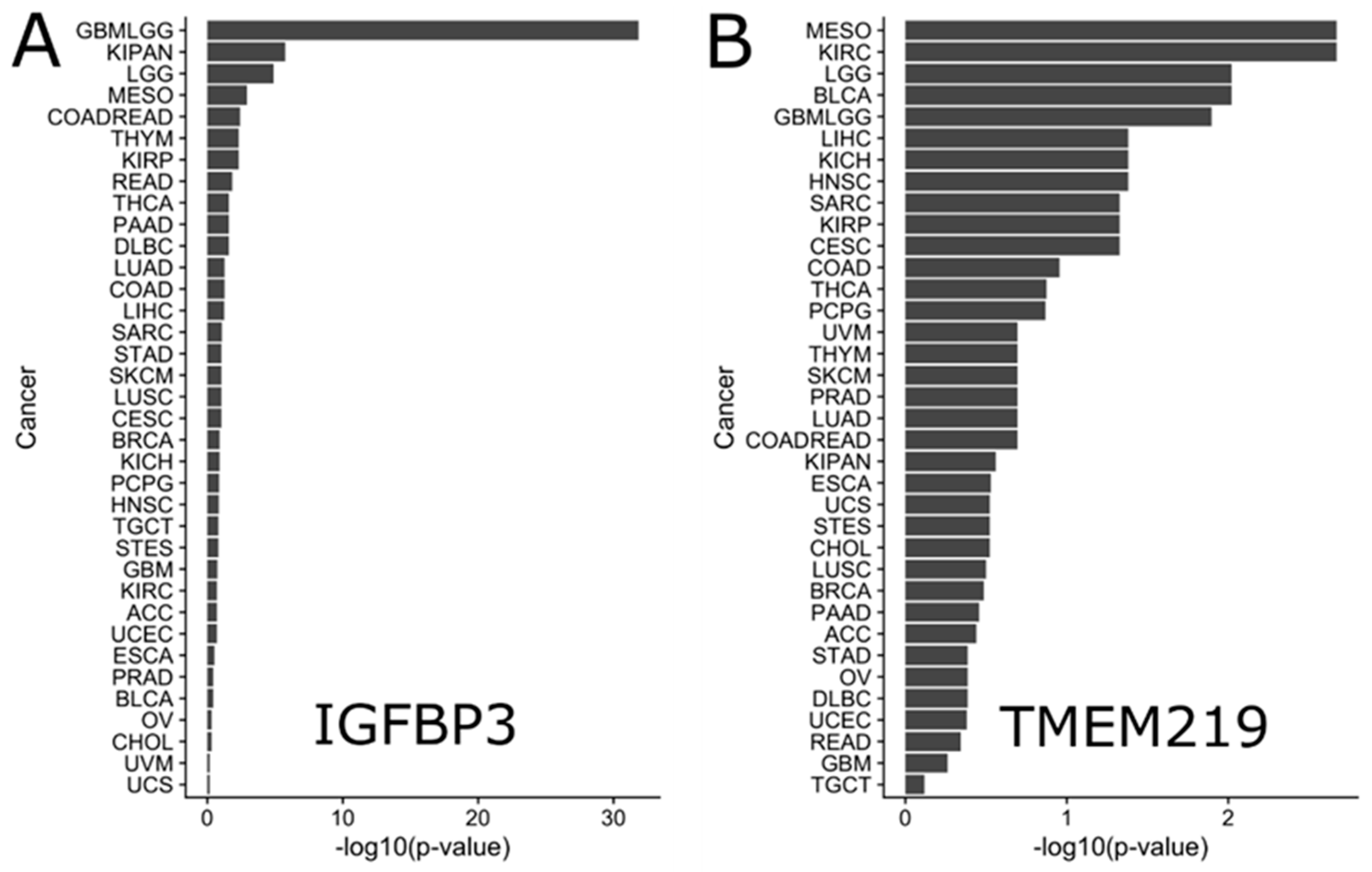
4.1. IGFBP-3 and TMEM219 Gene Expression in Tumor and Normal Samples
4.2. Pan-Cancer Survival Effect of IGFBP-3 and TMEM219
4.3. Survival Effect of IGFBP-3 and TMEM219 in Clinical Subcategories
4.4. Survival Effect of IGFBP-3 and TMEM219 in Breast Cancer
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wood, W.I.; Cachianes, G.; Henzel, W.J.; Winslow, G.A.; Spencer, S.A.; Hellmiss, R.; Martin, J.L.; Baxter, R.C. Cloning and expression of the growth hormone-dependent insulin-like growth factor-binding protein. Mol. Endocrinol. 1988, 2, 1176–1185. [Google Scholar] [CrossRef] [PubMed]
- Hwa, V.; Oh, Y.; Rosenfeld, R.G. The insulin-like growth factor-binding protein (IGFBP) superfamily. Endocr. Rev. 1999, 20, 761–787. [Google Scholar] [CrossRef] [PubMed]
- Jogie-Brahim, S.; Feldman, D.; Oh, Y. Unraveling insulin-like growth factor binding protein-3 actions in human disease. Endocr. Rev. 2009, 30, 417–437. [Google Scholar] [CrossRef] [PubMed]
- Yamada, P.M.; Lee, K.W. Perspectives in mammalian IGFBP-3 biology: Local vs. systemic action. Am. J. Physiol. Cell Physiol. 2009, 296, C954–C976. [Google Scholar] [CrossRef]
- Baxter, R.C. IGF binding proteins in cancer: Mechanistic and clinical insights. Nat. Rev. Cancer 2014, 14, 329–341. [Google Scholar] [CrossRef]
- Johnson, M.A.; Firth, S.M. IGFBP-3: A cell fate pivot in cancer and disease. Growth Horm. IGF Res. 2014, 24, 164–173. [Google Scholar] [CrossRef]
- Buckway, C.K.; Wilson, E.M.; Ahlsen, M.; Bang, P.; Oh, Y.; Rosenfeld, R.G. Mutation of three critical amino acids of the N-terminal domain of IGF-binding protein-3 essential for high affinity IGF binding. J. Clin. Endocrinol. Metab. 2001, 86, 4943–4950. [Google Scholar] [CrossRef]
- Firth, S.M.; Baxter, R.C. Characterisation of recombinant glycosylation variants of insulin-like growth factor binding protein-3. J. Endocrinol. 1999, 160, 379–387. [Google Scholar] [CrossRef]
- Hoeck, W.G.; Mukku, V.R. Identification of the major sites of phosphorylation in IGF binding protein-3. J. Cell Biochem. 1994, 56, 262–273. [Google Scholar] [CrossRef]
- Bang, P.; Fielder, P.J. Human pregnancy serum contains at least two distinct proteolytic activities with the ability to degrade insulin-like growth factor binding protein-3. Endocrinology 1997, 138, 3912–3917. [Google Scholar] [CrossRef]
- Firth, S.M.; Baxter, R.C. Cellular actions of the insulin-like growth factor binding proteins. Endocr. Rev. 2002, 23, 824–854. [Google Scholar] [CrossRef]
- Schedlich, L.J.; Nilsen, T.; John, A.P.; Jans, D.A.; Baxter, R.C. Phosphorylation of insulin-like growth factor binding protein-3 by deoxyribonucleic acid-dependent protein kinase reduces ligand binding and enhances nuclear accumulation. Endocrinology 2003, 144, 1984–1993. [Google Scholar] [CrossRef] [PubMed]
- Cobb, L.J.; Liu, B.; Lee, K.W.; Cohen, P. Phosphorylation by DNA-dependent protein kinase is critical for apoptosis induction by insulin-like growth factor binding protein-3. Cancer Res. 2006, 66, 10878–10884. [Google Scholar] [CrossRef] [PubMed]
- Fowlkes, J.L.; Thrailkill, K.M.; Serra, D.M.; Suzuki, K.; Nagase, H. Matrix metalloproteinases as insulin-like growth factor binding protein-degrading proteinases. Prog. Growth Factor Res. 1995, 6, 255–263. [Google Scholar] [CrossRef]
- Ingermann, A.R.; Yang, Y.F.; Han, J.; Mikami, A.; Garza, A.E.; Mohanraj, L.; Fan, L.; Idowu, M.; Ware, J.L.; Kim, H.S.; et al. Identification of a novel cell death receptor mediating IGFBP-3-induced anti-tumor effects in breast and prostate cancer. J. Biol. Chem. 2010, 285, 30233–30246. [Google Scholar] [CrossRef] [PubMed]
- Spencer, E.M.; Chan, K. A 3-dimensional model for the insulin-like growth factor binding proteins (IGFBPs); supporting evidence using the structural determinants of the IGF binding site on IGFBP-3. Prog. Growth Factor Res. 1995, 6, 209–214. [Google Scholar] [CrossRef]
- Ho, P.J.; Baxter, R.C. Characterization of truncated insulin-like growth factor-binding protein-2 in human milk. Endocrinology 1997, 138, 3811–3818. [Google Scholar] [CrossRef]
- Devi, G.R.; Yang, D.H.; Rosenfeld, R.G.; Oh, Y. Differential effects of insulin-like growth factor (IGF)-binding protein-3 and its proteolytic fragments on ligand binding, cell surface association, and IGF-I receptor signaling. Endocrinology 2000, 141, 4171–4179. [Google Scholar] [CrossRef]
- Galanis, M.; Firth, S.M.; Bond, J.; Nathanielsz, A.; Kortt, A.A.; Hudson, P.J.; Baxter, R.C. Ligand-binding characteristics of recombinant amino- and carboxyl-terminal fragments of human insulin-like growth factor-binding protein-3. J. Endocrinol. 2001, 169, 123–133. [Google Scholar] [CrossRef]
- Fowlkes, J.L.; Serra, D.M. Characterization of glycosaminoglycan-binding domains present in insulin-like growth factor-binding protein-3. J. Biol. Chem. 1996, 271, 14676–14679. [Google Scholar] [CrossRef]
- Smith, E.P.; Lu, L.; Chernausek, S.D.; Klein, D.J. Insulin-like growth factor-binding protein-3 (IGFBP-3) concentration in rat Sertoli cell-conditioned medium is regulated by a pathway involving association of IGFBP-3 with cell surface proteoglycans. Endocrinology 1994, 135, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Campbell, P.G.; Durham, S.K.; Hayes, J.D.; Suwanichkul, A.; Powell, D.R. Insulin-like growth factor-binding protein-3 binds fibrinogen and fibrin. J. Biol. Chem. 1999, 274, 30215–30221. [Google Scholar] [CrossRef] [PubMed]
- Baxter, R.C.; Martin, J.L.; Beniac, V.A. High molecular weight insulin-like growth factor binding protein complex. Purification and properties of the acid-labile subunit from human serum. J. Biol. Chem. 1989, 264, 11843–11848. [Google Scholar] [PubMed]
- Huq, A.; Singh, B.; Meeker, T.; Mascarenhas, D. The metal-binding domain of IGFBP-3 selectively delivers therapeutic molecules into cancer cells. Anticancer Drugs 2009, 20, 21–31. [Google Scholar] [CrossRef]
- Schedlich, L.J.; Le Page, S.L.; Firth, S.M.; Briggs, L.J.; Jans, D.A.; Baxter, R.C. Nuclear import of insulin-like growth factor-binding protein-3 and -5 is mediated by the importin beta subunit. J. Biol. Chem. 2000, 275, 23462–23470. [Google Scholar] [CrossRef]
- Lee, K.W.; Liu, B.; Ma, L.; Li, H.; Bang, P.; Koeffler, H.P.; Cohen, P. Cellular internalization of insulin-like growth factor binding protein-3: Distinct endocytic pathways facilitate re-uptake and nuclear localization. J. Biol. Chem. 2004, 279, 469–476. [Google Scholar] [CrossRef]
- Mohan, S.; Baylink, D.J.; Pettis, J.L. Insulin-like growth factor (IGF)-binding proteins in serum--do they have additional roles besides modulating the endocrine IGF actions? J. Clin. Endocrinol. Metab. 1996, 81, 3817–3820. [Google Scholar] [CrossRef][Green Version]
- Rajaram, S.; Baylink, D.J.; Mohan, S. Insulin-like growth factor-binding proteins in serum and other biological fluids: Regulation and functions. Endocr. Rev. 1997, 18, 801–831. [Google Scholar]
- Clemmons, D.R. Role of insulin-like growth factor binding proteins in controlling IGF actions. Mol. Cell Endocrinol. 1998, 140, 19–24. [Google Scholar] [CrossRef]
- Kelley, K.M.; Oh, Y.; Gargosky, S.E.; Gucev, Z.; Matsumoto, T.; Hwa, V.; Ng, L.; Simpson, D.M.; Rosenfeld, R.G. Insulin-like growth factor-binding proteins (IGFBPs) and their regulatory dynamics. Int. J. Biochem. Cell Biol. 1996, 28, 619–637. [Google Scholar] [CrossRef]
- Ferry, R.J., Jr.; Cerri, R.W.; Cohen, P. Insulin-like growth factor binding proteins: New proteins, new functions. Horm. Res. 1999, 51, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Collett-Solberg, P.F.; Cohen, P. Genetics, chemistry, and function of the IGF/IGFBP system. Endocrine 2000, 12, 121–136. [Google Scholar] [CrossRef]
- Oh, Y.; Muller, H.L.; Lamson, G.; Rosenfeld, R.G. Insulin-like growth factor (IGF)-independent action of IGF-binding protein-3 in Hs578T human breast cancer cells. Cell surface binding and growth inhibition. J. Biol. Chem. 1993, 268, 14964–14971. [Google Scholar] [PubMed]
- Rajah, R.; Valentinis, B.; Cohen, P. Insulin-like growth factor (IGF)-binding protein-3 induces apoptosis and mediates the effects of transforming growth factor-beta1 on programmed cell death through a p53-and IGF-independent mechanism. J. Biol. Chem. 1997, 272, 12181–12188. [Google Scholar] [CrossRef] [PubMed]
- Leal, S.M.; Huang, S.S.; Huang, J.S. Interactions of high affinity insulin-like growth factor-binding proteins with the type V transforming growth factor-beta receptor in mink lung epithelial cells. J. Biol. Chem. 1999, 274, 6711–6717. [Google Scholar] [CrossRef]
- Wu, H.B.; Kumar, A.; Tsai, W.C.; Mascarenhas, D.; Healey, J.; Rechler, M.M. Characterization of the inhibition of DNA synthesis in proliferating mink lung epithelial cells by insulin-like growth factor binding protein-3. J. Cell Biochem. 2000, 77, 288–297. [Google Scholar] [CrossRef]
- Ikonen, M.; Liu, B.; Hashimoto, Y.; Ma, L.; Lee, K.W.; Niikura, T.; Nishimoto, I.; Cohen, P. Interaction between the Alzheimer’s survival peptide humanin and insulin-like growth factor-binding protein 3 regulates cell survival and apoptosis. Proc. Natl. Acad. Sci. USA 2003, 100, 13042–13047. [Google Scholar] [CrossRef]
- Chen, X.; Ferry, R.J., Jr. Novel actions of IGFBP-3 on intracellular signaling pathways of insulin-secreting cells. Growth Horm. IGF Res. 2006, 16, 41–48. [Google Scholar] [CrossRef]
- Chang, K.H.; Chan-Ling, T.; McFarland, E.L.; Afzal, A.; Pan, H.; Baxter, L.C.; Shaw, L.C.; Caballero, S.; Sengupta, N.; Li Calzi, S.; et al. IGF binding protein-3 regulates hematopoietic stem cell and endothelial precursor cell function during vascular development. Proc. Natl. Acad. Sci. USA 2007, 104, 10595–10600. [Google Scholar] [CrossRef]
- Grimberg, A.; Coleman, C.M.; Burns, T.F.; Himelstein, B.P.; Koch, C.J.; Cohen, P.; El-Deiry, W.S. p53-Dependent and p53-independent induction of insulin-like growth factor binding protein-3 by deoxyribonucleic acid damage and hypoxia. J. Clin. Endocrinol. Metab. 2005, 90, 3568–3574. [Google Scholar] [CrossRef]
- Lofqvist, C.; Chen, J.; Connor, K.M.; Smith, A.C.; Aderman, C.M.; Liu, N.; Pintar, J.E.; Ludwig, T.; Hellstrom, A.; Smith, L.E. IGFBP3 suppresses retinopathy through suppression of oxygen-induced vessel loss and promotion of vascular regrowth. Proc. Natl. Acad. Sci. USA 2007, 104, 10589–10594. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.C.; Smartt, H.; H-Zadeh, A.; MMacfarlane, M.; Paraskeva, C.; Collard, T.J. Insulin-like growth factor binding protein 3 (IGFBP-3) potentiates TRAIL-induced apoptosis of human colorectal carcinoma cells through inhibition of NF-kappaB. Cell Death Differ. 2007, 14, 137–145. [Google Scholar] [CrossRef]
- Mehta, H.H.; Gao, Q.; Galet, C.; Paharkova, V.; Wan, J.; Said, J.; Sohn, J.J.; Lawson, G.; Cohen, P.; Cobb, L.J.; et al. IGFBP-3 is a metastasis suppression gene in prostate cancer. Cancer Res. 2011, 71, 5154–5163. [Google Scholar] [CrossRef] [PubMed]
- Mohanraj, L.; Kim, H.S.; Li, W.; Cai, Q.; Kim, K.E.; Shin, H.J.; Lee, Y.J.; Lee, W.J.; Kim, J.H.; Oh, Y. IGFBP-3 inhibits cytokine-induced insulin resistance and early manifestations of atherosclerosis. PLoS ONE 2013, 8, e55084. [Google Scholar] [CrossRef] [PubMed]
- Min, H.K.; Maruyama, H.; Jang, B.K.; Shimada, M.; Mirshahi, F.; Ren, S.; Oh, Y.; Puri, P.; Sanyal, A.J. Suppression of IGF binding protein-3 by palmitate promotes hepatic inflammatory responses. FASEB J. 2016, 30, 4071–4082. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Cai, Q.; Oh, Y. Therapeutic potential of alpha-1 antitrypsin in human disease. Ann. Pediatr. Endocrinol. Metab. 2018, 23, 131–135. [Google Scholar] [CrossRef]
- Arab, J.P.; Cabrera, D.; Sehrawat, T.S.; Jalan-Sakrikar, N.; Verma, V.K.; Simonetto, D.; Cao, S.; Yaqoob, U.; Leon, J.; Freire, M.; et al. Hepatic stellate cell activation promotes alcohol-induced steatohepatitis through Igfbp3 and SerpinA12. J. Hepatol. 2020. [Google Scholar] [CrossRef]
- Gucev, Z.S.; Oh, Y.; Kelley, K.M.; Rosenfeld, R.G. Insulin-like growth factor binding protein 3 mediates retinoic acid-and transforming growth factor beta2-induced growth inhibition in human breast cancer cells. Cancer Res. 1996, 56, 1545–1550. [Google Scholar]
- Hembree, J.R.; Agarwal, C.; Beard, R.L.; Chandraratna, R.A.; Eckert, R. Retinoid X receptor-specific retinoids inhibit the ability of retinoic acid receptor-specific retinoids to increase the level of insulin-like growth factor binding protein-3 in human ectocervical epithelial cells. Cancer Res. 1996, 56, 1794–1799. [Google Scholar]
- Rozen, F.; Zhang, J.; Pollak, M. Antiproliferative action of tumor necrosis factor-alpha on MCF-7 breastcancer cells is associated with increased insulin-like growth factor binding protein-3 accumulation. Int. J. Oncol. 1998, 13, 865–869. [Google Scholar] [CrossRef]
- Colston, K.W.; Perks, C.M.; Xie, S.P.; Holly, J.M. Growth inhibition of both MCF-7 and Hs578T human breast cancer cell lines by vitamin D analogues is associated with increased expression of insulin-like growth factor binding protein-3. J. Mol. Endocrinol. 1998, 20, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Huynh, H.; Yang, X.; Pollak, M. Estradiol and antiestrogens regulate a growth inhibitory insulin-like growth factor binding protein 3 autocrine loop in human breast cancer cells. J. Biol. Chem. 1996, 271, 1016–1021. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.L.; Baxter, R.C. Transforming growth factor-beta stimulates production of insulin-like growth factor-binding protein-3 by human skin fibroblasts. Endocrinology 1991, 128, 1425–1433. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Muller, H.L.; Ng, L.; Rosenfeld, R.G. Transforming growth factor-beta-induced cell growth inhibition in human breast cancer cells is mediated through insulin-like growth factor-binding protein-3 action. J. Biol. Chem. 1995, 270, 13589–13592. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Wang, S.; Ying, Y.; Zhou, R.; Mao, P. miR-196b/miR-1290 participate in the antitumor effect of resveratrol via regulation of IGFBP3 expression in acute lymphoblastic leukemia. Oncol. Rep. 2017, 37, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Jin, L.; Zhang, B.; Xue, H.; Li, Q.; Xu, Y. The potentiation of curcumin on insulin-like growth factor-1 action in MCF-7 human breast carcinoma cells. Life Sci. 2007, 80, 2161–2169. [Google Scholar] [CrossRef] [PubMed]
- Vijayababu, M.R.; Arunkumar, A.; Kanagaraj, P.; Arunakaran, J. Effects of quercetin on insulin-like growth factors (IGFs) and their binding protein-3 (IGFBP-3) secretion and induction of apoptosis in human prostate cancer cells. J. Carcinog. 2006, 5, 10. [Google Scholar] [CrossRef]
- Buckbinder, L.; Talbott, R.; Velasco-Miguel, S.; Takenaka, I.; Faha, B.; Seizinger, B.R.; Kley, N. Induction of the growth inhibitor IGF-binding protein 3 by p53. Nature 1995, 377, 646–649. [Google Scholar] [CrossRef]
- Bourdon, J.C.; Deguin-Chambon, V.; Lelong, J.C.; Dessen, P.; May, P.; Debuire, B.; May, E. Further characterisation of the p53 responsive element--identification of new candidate genes for trans-activation by p53. Oncogene 1997, 14, 85–94. [Google Scholar] [CrossRef]
- Ludwig, R.L.; Bates, S.; Vousden, K.H. Differential activation of target cellular promoters by p53 mutants with impaired apoptotic function. Mol. Cell Biol. 1996, 16, 4952–4960. [Google Scholar] [CrossRef]
- Barbieri, C.E.; Perez, C.A.; Johnson, K.N.; Ely, K.A.; Billheimer, D.; Pietenpol, J.A. IGFBP-3 is a direct target of transcriptional regulation by DeltaNp63alpha in squamous epithelium. Cancer Res. 2005, 65, 2314–2320. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.S.; Wang, L.; Liu, D.; Mao, L.; Hong, W.K.; Khuri, F.R.; Lee, H.Y. Correlation between insulin-like growth factor-binding protein-3 promoter methylation and prognosis of patients with stage I non-small cell lung cancer. Clin. Cancer Res. 2002, 8, 3669–3675. [Google Scholar] [PubMed]
- Wiley, A.; Katsaros, D.; Fracchioli, S.; Yu, H. Methylation of the insulin-like growth factor binding protein-3 gene and prognosis of epithelial ovarian cancer. Int. J. Gynecol. Cancer 2006, 16, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Ibanez de Caceres, I.; Cortes-Sempere, M.; Moratilla, C.; Machado-Pinilla, R.; Rodriguez-Fanjul, V.; Manguan-Garcia, C.; Cejas, P.; Lopez-Rios, F.; Paz-Ares, L.; de CastroCarpeno, J.; et al. IGFBP-3 hypermethylation-derived deficiency mediates cisplatin resistance in non-small-cell lung cancer. Oncogene 2010, 29, 1681–1690. [Google Scholar] [CrossRef] [PubMed]
- Harada, A.; Jogie-Brahim, S.; Oh, Y. Tobacco specific carcinogen 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone suppresses a newly identified anti-tumor IGFBP-3/IGFBP-3R system in lung cancer cells. Lung Cancer 2013, 80, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Perez-Carbonell, L.; Balaguer, F.; Toiyama, Y.; Egoavil, C.; Rojas, E.; Guarinos, C.; Andreu, M.; Llor, X.; Castells, A.; Jover, R.; et al. IGFBP3 methylation is a novel diagnostic and predictive biomarker in colorectal cancer. PLoS ONE 2014, 9, e104285. [Google Scholar] [CrossRef] [PubMed]
- Pernia, O.; Belda-Iniesta, C.; Pulido, V.; Cortes-Sempere, M.; Rodriguez, C.; Vera, O.; Soto, J.; Jimenez, J.; Taus, A.; Rojo, F.; et al. Methylation status of IGFBP-3 as a useful clinical tool for deciding on a concomitant radiotherapy. Epigenetics 2014, 9, 1446–1453. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Perks, C.M.; Holly, J.M. Epigenetic regulation of insulin-like growth factor binding protein-3 (IGFBP-3) in cancer. J. Cell Commun. Signal 2015, 9, 159–166. [Google Scholar] [CrossRef]
- Fu, T.; Pappou, E.P.; Guzzetta, A.A.; Calmon Mde, F.; Sun, L.; Herrera, A.; Li, F.; Wolfgang, C.L.; Baylin, S.B.; Iacobuzio-Donahue, C.A.; et al. IGFBP-3 Gene Methylation in Primary Tumor Predicts Recurrence of Stage II Colorectal Cancers. Ann. Surg. 2016, 263, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Muller, H.L.; Pham, H.; Rosenfeld, R.G. Demonstration of receptors for insulin-like growth factor binding protein-3 on Hs578T human breast cancer cells. J. Biol. Chem. 1993, 268, 26045–26048. [Google Scholar]
- Huang, S.S.; Ling, T.Y.; Tseng, W.F.; Huang, Y.H.; Tang, F.M.; Leal, S.M.; Huang, J.S. Cellular growth inhibition by IGFBP-3 and TGF-beta1 requires LRP-1. FASEB J. 2003, 17, 2068–2081. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Raz, A.; Murphy, L.J. Insulin-like growth factor binding protein-3 interacts with autocrine motility factor/phosphoglucose isomerase (AMF/PGI) and inhibits the AMF/PGI function. Cancer Res. 2004, 64, 2516–2522. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Weinzimer, S.A.; Gibson, T.B.; Collett-Solberg, P.F.; Khare, A.; Liu, B.; Cohen, P. Transferrin is an insulin-like growth factor-binding protein-3 binding protein. J. Clin. Endocrinol. Metab. 2001, 86, 1806–1813. [Google Scholar] [CrossRef] [PubMed]
- Leal, S.M.; Liu, Q.; Huang, S.S.; Huang, J.S. The type V transforming growth factor beta receptor is the putative insulin-like growth factor-binding protein 3 receptor. J. Biol. Chem. 1997, 272, 20572–20576. [Google Scholar] [CrossRef]
- Huang, S.S.; Leal, S.M.; Chen, C.L.; Liu, I.H.; Huang, J.S. Identification of insulin receptor substrate proteins as key molecules for the TbetaR-V/LRP-1-mediated growth inhibitory signaling cascade in epithelial and myeloid cells. FASEB J. 2004, 18, 1719–1721. [Google Scholar] [CrossRef]
- Lee, S.H.; Takahashi, M.; Honke, K.; Miyoshi, E.; Osumi, D.; Sakiyama, H.; Ekuni, A.; Wang, X.; Inoue, S.; Gu, J.; et al. Loss of core fucosylation of low-density lipoprotein receptor-related protein-1 impairs its function, leading to the upregulation of serum levels of insulin-like growth factor-binding protein 3 in Fut8-/-mice. J. Biochem. 2006, 139, 391–398. [Google Scholar] [CrossRef]
- Mangasser-Stephan, K.; Gressner, A.M. Molecular and functional aspects of latent transforming growth factor-beta binding protein: Just a masking protein? Cell Tissue Res. 1999, 297, 363–370. [Google Scholar] [CrossRef]
- Gui, Y.; Murphy, L.J. Interaction of insulin-like growth factor binding protein-3 with latent transforming growth factor-beta binding protein-1. Mol. Cell Biochem. 2003, 250, 189–195. [Google Scholar] [CrossRef]
- Rifkin, D.B. Latent transforming growth factor-beta (TGF-beta) binding proteins: Orchestrators of TGF-beta availability. J. Biol. Chem. 2005, 280, 7409–7412. [Google Scholar] [CrossRef]
- Nwosu, Z.C.; Ebert, M.P.; Dooley, S.; Meyer, C. Caveolin-1 in the regulation of cell metabolism: A cancer perspective. Mol. Cancer 2016, 15, 71. [Google Scholar] [CrossRef]
- Oufattole, M.; Lin, S.W.; Liu, B.; Mascarenhas, D.; Cohen, P.; Rodgers, B.D. Ribonucleic acid polymerase II binding subunit 3 (Rpb3), a potential nuclear target of insulin-like growth factor binding protein-3. Endocrinology 2006, 147, 2138–2146. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Yao, G.; Zou, M.; Chen, G.; Wang, M.; Liu, J.; Wang, J.; Xu, D. N-Acetylgalactosaminyltransferase 14, a novel insulin-like growth factor binding protein-3 binding partner. Biochem. Biophys. Res. Commun. 2007, 357, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Harada, A.; Oh, Y. IGFBP-3 sensitizes antiestrogen-resistant breast cancer cells through interaction with GRP78. Cancer Lett. 2012, 325, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Grkovic, S.; O’Reilly, V.C.; Han, S.; Hong, M.; Baxter, R.C.; Firth, S.M. IGFBP-3 binds GRP78, stimulates autophagy and promotes the survival of breast cancer cells exposed to adverse microenvironments. Oncogene 2013, 32, 2412–2420. [Google Scholar] [CrossRef]
- Liu, B.; Lee, H.Y.; Weinzimer, S.A.; Powell, D.R.; Clifford, J.L.; Kurie, J.M.; Cohen, P. Direct functional interactions between insulin-like growth factor-binding protein-3 and retinoid X receptor-alpha regulate transcriptional signaling and apoptosis. J. Biol. Chem. 2000, 275, 33607–33613. [Google Scholar] [CrossRef] [PubMed]
- Schedlich, L.J.; O’Han, M.K.; Leong, G.M.; Baxter, R.C. Insulin-like growth factor binding protein-3 prevents retinoid receptor heterodimerization: Implications for retinoic acid-sensitivity in human breast cancer cells. Biochem. Biophys. Res. Commun. 2004, 314, 83–88. [Google Scholar] [CrossRef]
- Lee, K.W.; Cobb, L.J.; Paharkova-Vatchkova, V.; Liu, B.; Milbrandt, J.; Cohen, P. Contribution of the orphan nuclear receptor Nur77 to the apoptotic action of IGFBP-3. Carcinogenesis 2007, 28, 1653–1658. [Google Scholar] [CrossRef]
- Moreno-Santos, I.; Castellano-Castillo, D.; Lara, M.F.; Fernandez-Garcia, J.C.; Tinahones, F.J.; Macias-Gonzalez, M. IGFBP-3 Interacts with the Vitamin D Receptor in Insulin Signaling Associated with Obesity in Visceral Adipose Tissue. Int. J. Mol. Sci. 2017, 18, 2349. [Google Scholar] [CrossRef]
- Chan, S.S.; Schedlich, L.J.; Twigg, S.M.; Baxter, R.C. Inhibition of adipocyte differentiation by insulin-like growth factor-binding protein-3. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E654–E663. [Google Scholar] [CrossRef]
- Njomen, E.; Evans, H.G.; Gedara, S.H.; Heyl, D.L. Humanin Peptide Binds to Insulin-Like Growth Factor-Binding Protein 3 (IGFBP3) and Regulates Its Interaction with Importin-beta. Protein. Pept. Lett. 2015, 22, 869–876. [Google Scholar] [CrossRef]
- Wang, H.; Tachibana, K.; Zhang, Y.; Iwasaki, H.; Kameyama, A.; Cheng, L.; Guo, J.; Hiruma, T.; Togayachi, A.; Kudo, T.; et al. Cloning and characterization of a novel UDP-GalNAc:polypeptide N-acetylgalactosaminyltransferase, pp-GalNAc-T14. Biochem. Biophys. Res. Commun. 2003, 300, 738–744. [Google Scholar] [CrossRef]
- Kwon, O.S.; Oh, E.; Park, J.R.; Lee, J.S.; Bae, G.Y.; Koo, J.H.; Kim, H.; Choi, Y.L.; Choi, Y.S.; Kim, J.; et al. GalNAc-T14 promotes metastasis through Wnt dependent HOXB9 expression in lung adenocarcinoma. Oncotarget 2015, 6, 41916–41928. [Google Scholar] [CrossRef]
- Yang, J.; Hu, Y.; Wu, J.; Kong, S. Effects of IGFBP-3 and GalNAc-T14 on proliferation and cell cycle of glioblastoma cells and its mechanism. J. Pharm. Pharmacol. 2020, 72, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ni, M.; Lee, B.; Barron, E.; Hinton, D.R.; Lee, A.S. The unfolded protein response regulator GRP78/BiP is required for endoplasmic reticulum integrity and stress-induced autophagy in mammalian cells. Cell Death Differ. 2008, 15, 1460–1471. [Google Scholar] [CrossRef] [PubMed]
- Bertolotti, A.; Zhang, Y.; Hendershot, L.M.; Harding, H.P.; Ron, D. Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response. Nat. Cell Biol. 2000, 2, 326–332. [Google Scholar] [CrossRef]
- Wang, M.; Wey, S.; Zhang, Y.; Ye, R.; Lee, A.S. Role of the unfolded protein response regulator GRP78/BiP in development, cancer, and neurological disorders. Antioxid. Redox. Signal 2009, 11, 2307–2316. [Google Scholar] [CrossRef] [PubMed]
- Jing, G.; Wang, J.J.; Zhang, S.X. ER stress and apoptosis: A new mechanism for retinal cell death. Exp. Diabetes Res. 2012, 2012, 589589. [Google Scholar] [CrossRef]
- Lee, K.W.; Ma, L.; Yan, X.; Liu, B.; Zhang, X.K.; Cohen, P. Rapid apoptosis induction by IGFBP-3 involves an insulin-like growth factor-independent nucleomitochondrial translocation of RXRalpha/Nur77. J. Biol. Chem. 2005, 280, 16942–16948. [Google Scholar] [CrossRef]
- Kastner, P.; Mark, M.; Chambon, P. Nonsteroid nuclear receptors: What are genetic studies telling us about their role in real life? Cell 1995, 83, 859–869. [Google Scholar] [CrossRef]
- Mangelsdorf, D.J.; Evans, R.M. The RXR heterodimers and orphan receptors. Cell 1995, 83, 841–850. [Google Scholar] [CrossRef]
- Germain, P.; Staels, B.; Dacquet, C.; Spedding, M.; Laudet, V. Overview of nomenclature of nuclear receptors. Pharmacol. Rev. 2006, 58, 685–704. [Google Scholar] [CrossRef] [PubMed]
- Hazel, T.G.; Nathans, D.; Lau, L.F. A gene inducible by serum growth factors encodes a member of the steroid and thyroid hormone receptor superfamily. Proc. Natl. Acad. Sci. USA 1988, 85, 8444–8448. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, N.; Pechhold, K.; Shahjee, H.; Zappala, G.; Elbi, C.; Raaka, B.; Wiench, M.; Hong, J.; Rechler, M.M. Nonsecreted insulin-like growth factor binding protein-3 (IGFBP-3) can induce apoptosis in human prostate cancer cells by IGF-independent mechanisms without being concentrated in the nucleus. J. Biol. Chem. 2006, 281, 24588–24601. [Google Scholar] [CrossRef] [PubMed]
- Zappala, G.; Elbi, C.; Edwards, J.; Gorenstein, J.; Rechler, M.M.; Bhattacharyya, N. Induction of apoptosis in human prostate cancer cells by insulin-like growth factor binding protein-3 does not require binding to retinoid X receptor-alpha. Endocrinology 2008, 149, 1802–1812. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, Y.; Fowlkes, J.L.; Wilson, E.M.; Rosenfeld, R.G.; Oh, Y. Characterization of insulin-like growth factor binding protein-3 (IGFBP-3) binding to human breast cancer cells: Kinetics of IGFBP-3 binding and identification of receptor binding domain on the IGFBP-3 molecule. Endocrinology 1999, 140, 1319–1328. [Google Scholar] [CrossRef][Green Version]
- Ruan, W.; Becker, V.; Klingmuller, U.; Langosch, D. The interface between self-assembling erythropoietin receptor transmembrane segments corresponds to a membrane-spanning leucine zipper. J. Biol. Chem. 2004, 279, 3273–3279. [Google Scholar] [CrossRef]
- Noordeen, N.A.; Carafoli, F.; Hohenester, E.; Horton, M.A.; Leitinger, B. A transmembrane leucine zipper is required for activation of the dimeric receptor tyrosine kinase DDR1. J. Biol. Chem. 2006, 281, 22744–22751. [Google Scholar] [CrossRef]
- Ding, H.F.; Lin, Y.L.; McGill, G.; Juo, P.; Zhu, H.; Blenis, J.; Yuan, J.; Fisher, D.E. Essential role for caspase-8 in transcription-independent apoptosis triggered by p53. J. Biol. Chem. 2000, 275, 38905–38911. [Google Scholar] [CrossRef]
- Zhao, H.; Ross, F.P.; Teitelbaum, S.L. Unoccupied alpha(v)beta3 integrin regulates osteoclast apoptosis by transmitting a positive death signal. Mol. Endocrinol. 2005, 19, 771–780. [Google Scholar] [CrossRef][Green Version]
- Baxter, R.C.; Martin, J.L. Structure of the Mr 140,000 growth hormone-dependent insulin-like growth factor binding protein complex: Determination by reconstitution and affinity-labeling. Proc. Natl. Acad. Sci. USA 1989, 86, 6898–6902. [Google Scholar] [CrossRef]
- Han, J.; Jogie-Brahim, S.; Harada, A.; Oh, Y. Insulin-like growth factor-binding protein-3 suppresses tumor growth via activation of caspase-dependent apoptosis and cross-talk with NF-kappaB signaling. Cancer Lett. 2011, 307, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y. Treatment of Diseases Related to IGFBP-3 and Its Receptor. International Application Number No. PCT/US2017/059244, International Publication Number WO/2018/085252, 11 May 2018. [Google Scholar]
- Zhang, X.D.; Nguyen, T.; Thomas, W.D.; Sanders, J.E.; Hersey, P. Mechanisms of resistance of normal cells to TRAIL induced apoptosis vary between different cell types. FEBS Lett. 2000, 482, 193–199. [Google Scholar] [CrossRef]
- Wagner, K.W.; Punnoose, E.A.; Januario, T.; Lawrence, D.A.; Pitti, R.M.; Lancaster, K.; Lee, D.; von Goetz, M.; Yee, S.F.; Totpal, K.; et al. Death-receptor O-glycosylation controls tumor-cell sensitivity to the proapoptotic ligand Apo2L/TRAIL. Nat. Med. 2007, 13, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Dubuisson, A.; Micheau, O. Antibodies and Derivatives Targeting DR4 and DR5 for Cancer Therapy. Antibodies 2017, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12. [Google Scholar] [CrossRef] [PubMed]
- R2: Genomics Analysis and Visualization Platform. Available online: http://r2.amc.nl (accessed on 12 May 2020).
- Mihaly, Z.; Kormos, M.; Lanczky, A.; Dank, M.; Budczies, J.; Szasz, M.A.; Gyorffy, B. A meta-analysis of gene expression-based biomarkers predicting outcome after tamoxifen treatment in breast cancer. Breast Cancer Res. Treat. 2013, 140, 219–232. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 289–300. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TCGA ID | Description | log2 Fold Change | p-Value | Mean log2(RSEM) Normal | Mean log2(RSEM) Cancer |
|---|---|---|---|---|---|
| Upregulated in Tumor | |||||
| KIRC | Kidney renal clear cell carcinoma | 3.44 | 3.11 10−53 | 11.86 | 15.30 |
| LUSC | Lung squamous cell carcinoma | 2.26 | 1.22 10−28 | 10.61 | 12.86 |
| LUAD | Lung adenocarcinoma | 1.94 | 9.28 10−21 | 10.31 | 12.26 |
| HNSC | Head and Neck squamous cell carcinoma | 1.32 | 2.29 10−7 | 10.82 | 12.14 |
| STAD | Stomach adenocarcinoma | 1.18 | 8.67 10−7 | 10.82 | 12.00 |
| THCA | Thyroid carcinoma | 1.11 | 4.85 10−16 | 9.46 | 10.57 |
| BLCA | Bladder urothelial carcinoma | 1.05 | 3.57 10−3 | 12.56 | 13.61 |
| COAD | Colon adenocarcinoma | 0.65 | 4.01 10−9 | 10.45 | 11.10 |
| KIRP | Kidney renal papillary cell carcinoma | 0.34 | 5.19 10−2 | 11.69 | 12.03 |
| Downregulated in Tumor | |||||
| LIHC | Liver hepatocellular carcinoma | −2.53 | 4.17 10−81 | 14.18 | 11.65 |
| KICH | Kidney Chromophobe | −1.07 | 6.80 10−5 | 12.32 | 11.25 |
| BRCA | Breast invasive carcinoma | −0.82 | 9.74 10−24 | 12.04 | 11.23 |
| PRAD | Prostate adenocarcinoma | −0.56 | 1.47 10−5 | 11.27 | 10.72 |
| UCEC | Uterine Corpus Endometrial Carcinoma | −0.43 | 1.53 10−1 | 13.29 | 12.86 |
| TCGA ID | Description | log2 Fold Change | p-Value | Mean log2(RSEM) Normal | Mean log2(RSEM) Cancer |
|---|---|---|---|---|---|
| Upregulated in Tumor | |||||
| KIRP | Kidney renal papillary cell carcinoma | 0.53 | 3.16 10−14 | 11.07 | 11.60 |
| THCA | Thyroid carcinoma | 0.40 | 2.77 10−8 | 10.94 | 11.34 |
| BRCA | Breast invasive carcinoma | 0.39 | 1.00 10−19 | 10.71 | 11.10 |
| KIRC | Kidney renal clear cell carcinoma | 0.29 | 4.30 10−11 | 10.88 | 11.18 |
| BLCA | Bladder urothelial carcinoma | 0.27 | 1.08 10−1 | 10.92 | 11.19 |
| UCEC | Uterine Corpus Endometrial Carcinoma | 0.22 | 4.47 10−3 | 11.10 | 11.32 |
| LIHC | Liver hepatocellular carcinoma | 0.09 | 2.11 10−1 | 11.30 | 11.39 |
| PRAD | Prostate adenocarcinoma | 0.08 | 2.52 10−1 | 11.18 | 11.26 |
| Downregulated in Tumor | |||||
| LUSC | Lung squamous cell carcinoma | −0.74 | 6.56 10−23 | 11.18 | 10.45 |
| STAD | Stomach adenocarcinoma | −0.42 | 5.93 10−4 | 10.86 | 10.44 |
| COAD | Colon adenocarcinoma | −0.36 | 1.47 10−7 | 11.25 | 10.89 |
| HNSC | Head and neck squamous cell carcinoma | −0.32 | 9.00 10−4 | 10.63 | 10.30 |
| LUAD | Lung adenocarcinoma | −0.12 | 3.97 10−2 | 11.18 | 11.06 |
| KICH | Kidney Chromophobe | −0.10 | 3.20 10−1 | 11.24 | 11.14 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, Q.; Dozmorov, M.; Oh, Y. IGFBP-3/IGFBP-3 Receptor System as an Anti-Tumor and Anti-Metastatic Signaling in Cancer. Cells 2020, 9, 1261. https://doi.org/10.3390/cells9051261
Cai Q, Dozmorov M, Oh Y. IGFBP-3/IGFBP-3 Receptor System as an Anti-Tumor and Anti-Metastatic Signaling in Cancer. Cells. 2020; 9(5):1261. https://doi.org/10.3390/cells9051261
Chicago/Turabian StyleCai, Qing, Mikhail Dozmorov, and Youngman Oh. 2020. "IGFBP-3/IGFBP-3 Receptor System as an Anti-Tumor and Anti-Metastatic Signaling in Cancer" Cells 9, no. 5: 1261. https://doi.org/10.3390/cells9051261
APA StyleCai, Q., Dozmorov, M., & Oh, Y. (2020). IGFBP-3/IGFBP-3 Receptor System as an Anti-Tumor and Anti-Metastatic Signaling in Cancer. Cells, 9(5), 1261. https://doi.org/10.3390/cells9051261