Effects of Pyridoxine Deficiency on Hippocampal Function and Its Possible Association with V-Type Proton ATPase Subunit B2 and Heat Shock Cognate Protein 70


 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Novel Object Recognition Test
2.3. High-Performance Liquid Chromatography Analysis
2.4. Immunohistochemistry
2.5. Proteomic Analysis
2.5.1. Protein Preparation for 2DE
2.5.2. Analysis of 2DE Gels
2.5.3. Trypsin Digestion
2.5.4. Protein Identification Using MALDI-TOF MS
2.5.5. Data Searches for Protein Identification
2.6. Validation by Western Blot Analyses
2.7. Data Analysis
2.8. Statistical Analysis
3. Results
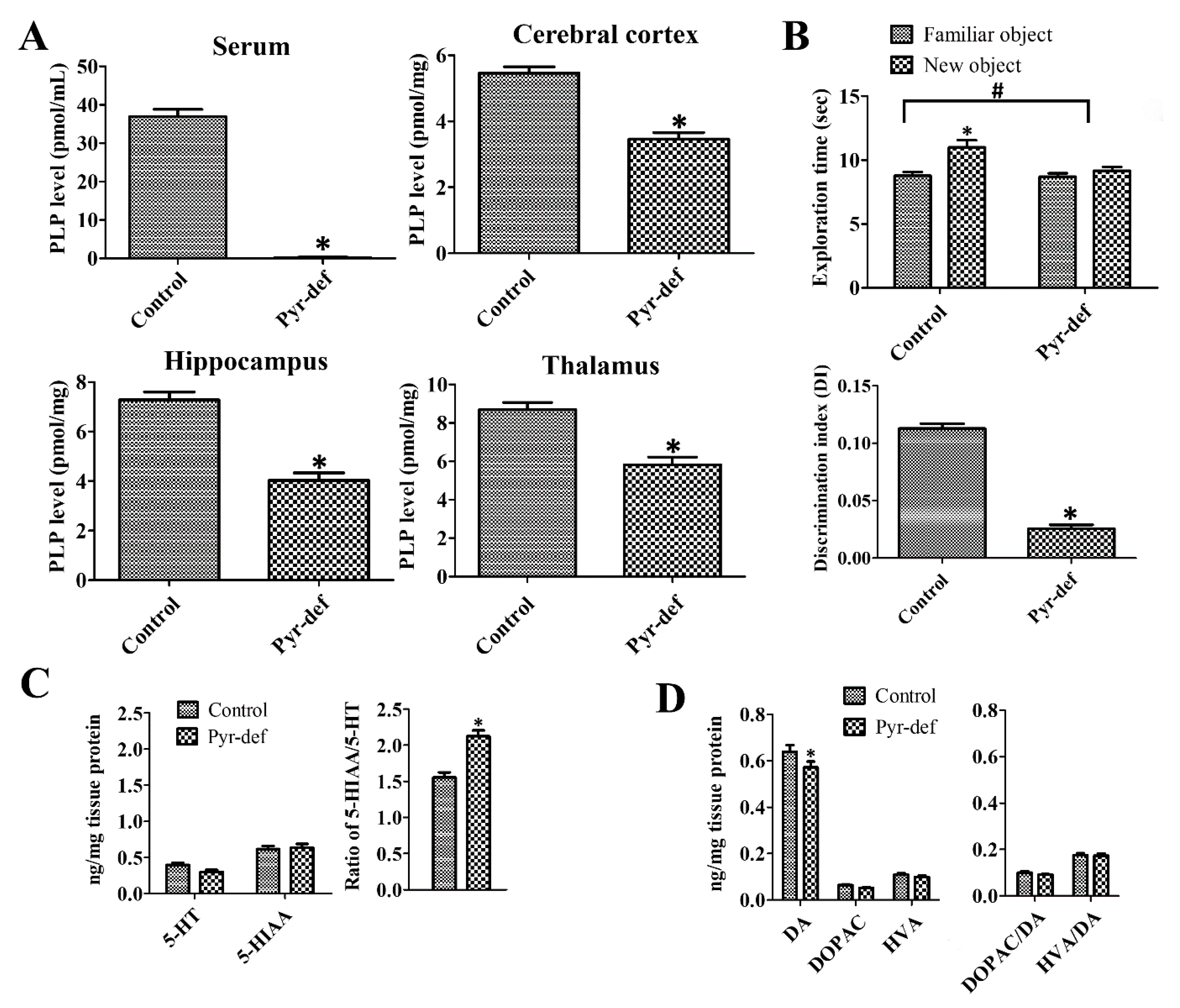
3.1. Effects of Pyridoxine Deficiency on PLP Levels in Serum and Brain Tissues
3.2. Effects of Pyridoxine Deficiency on Novel Object Recognition
3.3. Effects of Pyridoxine Deficiency on Monoamine Levels in the Hippocampus
3.4. Effects of Pyridoxine Deficiency on Ki67 and DCX Immunoreactivity
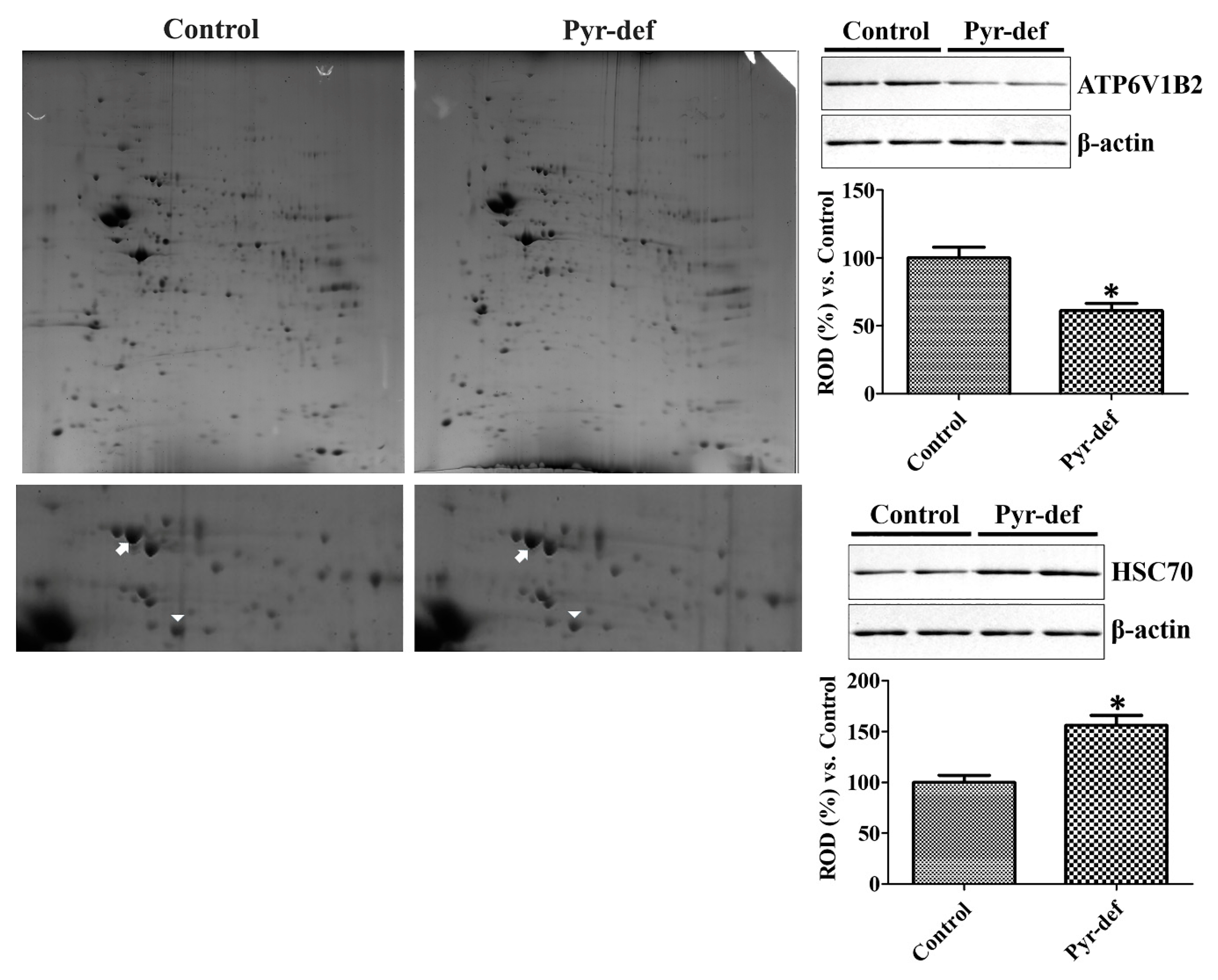
3.5. Protein Identification by 2D-DIGE Followed by MALDI-TOF MS in the Hippocampus
3.6. Validation of Identified Proteins by Western Blot
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sibbe, M.; Kulik, A. GABAergic regulation of adult hippocampal neurogenesis. Mol. Neurobiol. 2017, 54, 5497–5510. [Google Scholar] [CrossRef]
- Hodge, R.D.; Kowalczyk, T.D.; Wolf, S.A.; Encinas, J.M.; Rippey, C.; Enikolopov, G.; Kempermann, G.; Hevner, R.F. Intermediate progenitors in adult hippocampal neurogenesis: Tbr2 expression and coordinate regulation of neuronal output. J. Neurosci. 2008, 28, 3707–3717. [Google Scholar] [CrossRef]
- Yau, S.Y.; Li, A.; So, K.F. Involvement of adult hippocampal neurogenesis in learning and forgetting. Neural Plast. 2015, 2015, 717958. [Google Scholar] [CrossRef] [PubMed]
- Freund, T.F.; Buzsáki, G. Interneurons of the hippocampus. Hippocampus 1996, 6, 347–470. [Google Scholar] [CrossRef]
- Jessberger, S.; Clark, R.E.; Broadbent, N.J.; Clemenson, G.D., Jr.; Consiglio, A.; Lie, D.C.; Squire, L.R.; Gage, F.H. Dentate gyrus-specific knockdown of adult neurogenesis impairs spatial and object recognition memory in adult rats. Learn. Mem. 2009, 16, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Franco, M. Pyridoxine 5′-phosphate synthase: De novo synthesis of vitamin B6 and beyond. Biochim. Biophys. Acta 2003, 1647, 92–97. [Google Scholar] [CrossRef]
- Lheureux, P.; Penaloza, A.; Gris, M. Pyridoxine in clinical toxicology: A review. Eur. J. Emerg. Med. 2005, 12, 78–85. [Google Scholar] [CrossRef]
- Mooney, S.; Leuendorf, J.E.; Hendrickson, C.; Hellmann, H. Vitamin B6: A long known compound of surprising complexity. Molecules 2009, 14, 329–351. [Google Scholar] [CrossRef]
- Baldewicz, T.; Goodkin, K.; Feaster, D.J.; Blaney, N.T.; Kumar, M.; Kumar, A.; Shor-Posner, G.; Baum, M. Plasma pyridoxine deficiency is related to increased psychological distress in recently bereaved homosexual men. Psychosom. Med. 1998, 60, 297–308. [Google Scholar] [CrossRef]
- Jung, H.Y.; Kim, D.W.; Nam, S.M.; Kim, J.W.; Chung, J.Y.; Won, M.H.; Seong, J.K.; Yoon, Y.S.; Yoo, D.Y.; Hwang, I.K. Pyridoxine improves hippocampal cognitive function via increases of serotonin turnover and tyrosine hydroxylase, and its association with CB1 cannabinoid receptor-interacting protein and the CB1 cannabinoid receptor pathway. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 3142–3153. [Google Scholar] [CrossRef]
- McCarty, M.F. High-dose pyridoxine as an ‘anti-stress’ strategy. Med. Hypotheses 2000, 54, 803–807. [Google Scholar] [CrossRef] [PubMed]
- Yoo, D.Y.; Kim, W.; Kim, D.W.; Yoo, K.Y.; Chung, J.Y.; Youn, H.Y.; Yoon, Y.S.; Choi, S.Y.; Won, M.H.; Hwang, I.K. Pyridoxine enhances cell proliferation and neuroblast differentiation by upregulating the GABAergic system in the mouse dentate gyrus. Neurochem. Res. 2011, 36, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Dakshinamurti, K.; Dakshinamurti, S.; Czubryt, M.P. Vitamin B6: Effects of deficiency, and metabolic and therapeutic functions. In Handbook of Famine, Starvation, and Nutrient Deprivation; Preedy, V., Patel, V., Eds.; Springer: Cham, Switzerland, 2017; pp. 1–23. [Google Scholar]
- Stephens, M.C.; Havlicek, V.; Dakshinamurti, K. Pyridoxine deficiency and development of the central nervous system in the rat. J. Neurochem. 1971, 18, 2407–2416. [Google Scholar] [CrossRef] [PubMed]
- Stephens, M.C.; Dakshinamurti, K. Brain lipids in pyridoxine-deficient young rats. Neurobiology 1975, 5, 262–269. [Google Scholar]
- Williamson, B.; Coniglio, J.G. The effects of pyridoxine deficiency and of caloric restriction on lipids in the developing rat brain. J. Neurochem. 1971, 18, 267–276. [Google Scholar] [CrossRef]
- Abdou, H.M.; Wahby, M.M. Neuroprotection of grape seed extract and pyridoxine against triton-induced neurotoxicity. Oxid. Med. Cell. Longev. 2016, 2016, 8679506. [Google Scholar] [CrossRef]
- Dakshinamurti, S.; Dakshinamurti, K. Antihypertensive and neuroprotective actions of pyridoxine and its derivatives. Can. J. Physiol. Pharmacol. 2015, 93, 1083–1090. [Google Scholar] [CrossRef]
- Hwang, I.K.; Yoo, K.Y.; Kim, D.H.; Lee, B.H.; Kwon, Y.G.; Won, M.H. Time course of changes in pyridoxal 5′-phosphate (vitamin B6 active form) and its neuroprotection in experimental ischemic damage. Exp. Neurol. 2007, 206, 114–125. [Google Scholar] [CrossRef]
- Kuypers, N.J.; Hoane, M.R. Pyridoxine administration improves behavioral and anatomical outcome after unilateral contusion injury in the rat. J. Neurotrauma 2010, 27, 1275–1282. [Google Scholar] [CrossRef]
- Wei, Y.; Lu, M.; Mei, M.; Wang, H.; Han, Z.; Chen, M.; Yao, H.; Song, N.; Ding, X.; Ding, J.; et al. Pyridoxine induces glutathione synthesis via PKM2-mediated Nrf2 transactivation and confers neuroprotection. Nat. Commun. 2020, 11, 941. [Google Scholar] [CrossRef]
- Jeanclos, E.; Albersen, M.; Ramos, R.J.J.; Raab, A.; Wilhelm, C.; Hommers, L.; Lesch, K.P.; Verhoeven-Duif, N.M.; Gohla, A. Improved cognition, mild anxiety-like behavior and decreased motor performance in pyridoxal phosphatase-deficient mice. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Bayoumi, R.A.; Smith, W.R. Some effects of dietary vitamin B6 deficiency on γ-aminobutyric acid metabolism in developing rat brain. J. Neurochem. 1972, 19, 1883–1897. [Google Scholar] [CrossRef] [PubMed]
- Krishna, A.P.; Ramakrishna, T. Effect of pyridoxine deficiency on the structural and functional development of hippocampus. Indian J. Physiol. Pharmacol. 2004, 48, 304–310. [Google Scholar] [PubMed]
- Brown, M.J.; Beier, K. Vitamin B6 deficiency (pyridoxine). In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Masisi, K.; Suidasari, S.; Zhang, P.; Okazaki, Y.; Yanaka, N.; Kato, N. Comparative study on the responses of concentrations of B6-vitamers in several tissues of mice to the dietary level of pyridoxine. J. Nutr. Sci. Vitaminol. (Tokyo) 2012, 58, 446–451. [Google Scholar] [CrossRef][Green Version]
- Rowland, N.E.; Dunn, A.J. Effect of dexfenfluramine on metabolic and neurochemical measures in restraint-stressed ob/ob mice. Physiol. Behav. 1995, 58, 749–754. [Google Scholar] [CrossRef]
- Franklin, K.B.J.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates; Academic Press: San Diego, CA, USA, 1997. [Google Scholar]
- Yoo, D.Y.; Cho, S.B.; Jung, H.Y.; Kim, W.; Choi, G.M.; Won, M.H.; Kim, D.W.; Hwang, I.K.; Choi, S.Y.; Moon, S.M. Tat-protein disulfide-isomerase A3: A possible candidate for preventing ischemic damage in the spinal cord. Cell Death Dis. 2017, 8, e3075. [Google Scholar] [CrossRef]
- Bowling, F.G. Pyridoxine supply in human development. Semin. Cell Dev. Biol. 2011, 22, 611–618. [Google Scholar] [CrossRef]
- Georgieff, M.K.; Brunette, K.E.; Tran, P.V. Early life nutrition and neural plasticity. Dev. Psychopathol. 2015, 27, 411–423. [Google Scholar] [CrossRef]
- Guilarte, T.R.; Miceli, R.C.; Moran, T.H. Developmental effects of vitamin B-6 restriction on the locomotor behavior of rats. Brain Res. Bull. 1991, 26, 857–861. [Google Scholar] [CrossRef]
- Dakshinamurti, K.; LeBlancq, W.D.; Herchl, R.; Havlicek, V. Nonparallel changes in brain monoamines of pyridoxine-deficient growing rats. Exp. Brain Res. 1976, 26, 355–366. [Google Scholar] [CrossRef]
- Guilarte, T.R.; Wagner, H.N., Jr.; Frost, J.J. Effects of perinatal vitamin B6 deficiency on dopaminergic neurochemistry. J. Neurochem. 1987, 48, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Guilarte, T.R. Effect of vitamin B-6 nutrition on the levels of dopamine, dopamine metabolites, dopa decarboxylase activity, tyrosine, and GABA in the developing rat corpus striatum. Neurochem. Res. 1989, 14, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.R.; Mabasa, L.; Crane, C.; Park, C.S.; Venâncio, V.P.; Bianchi, M.L.; Antunes, L.M. Maternal vitamin B6 deficient or supplemented diets on expression of genes related to GABAergic, serotonergic, or glutamatergic pathways in hippocampus of rat dams and their offspring. Mol. Nutr. Food Res. 2016, 60, 1615–1624. [Google Scholar] [CrossRef] [PubMed]
- Glier, M.B.; Green, T.J.; Devlin, A.M. Methyl nutrients, DNA methylation, and cardiovascular disease. Mol. Nutr. Food Res. 2014, 58, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Gilarte, T.R. The role of vitamin B6 in central nervous system development: Neurochemistry and behavior. In Vitamin B6 Metabolism in Pregnancy, Lactation, and Infancy; CRC Press: Boca Raton, FL, USA, 1995; pp. 77–92. [Google Scholar]
- Qian, B.; Shen, S.; Zhang, J.; Jing, P. Effects of vitamin B6 deficiency on the composition and functional potential of T cell populations. J. Immunol. Res. 2017, 2017, 2197975. [Google Scholar] [CrossRef]
- Cohen, S.J.; Stackman, R.W., Jr. Assessing rodent hippocampal involvement in the novel object recognition task. A review. Behav. Brain Res. 2015, 285, 105–117. [Google Scholar] [CrossRef]
- Root, E.J.; Longenecker, J.B. Brain cell alterations suggesting premature aging induced by dietary deficiency of vitamin B6 and/or copper. Am. J. Clin. Nutr. 1983, 37, 540–552. [Google Scholar] [CrossRef]
- Zhuo, J.M.; Praticò, D. Acceleration of brain amyloidosis in an Alzheimer’s disease mouse model by a folate, vitamin B6 and B12-deficient diet. Exp. Gerontol. 2010, 45, 195–201. [Google Scholar] [CrossRef]
- Li, P.; Zhu, M.L.; Pan, G.P.; Lu, J.X.; Zhao, F.R.; Jian, X.; Liu, L.Y.; Wan, G.R.; Chen, Y.; Ping, S.; et al. Vitamin B6 prevents isocarbophos-induced vascular dementia in rats through N-methyl-D-aspartate receptor signaling. Clin. Exp. Hypertens. 2018, 40, 192–201. [Google Scholar] [CrossRef]
- Fernandez, S.P.; Muzerelle, A.; Scotto-Lomassese, S.; Barik, J.; Gruart, A.; Delgado-García, J.M.; Gaspar, P. Constitutive and acquired serotonin deficiency alters memory and hippocampal synaptic plasticity. Neuropsychopharmacology 2017, 42, 512–523. [Google Scholar] [CrossRef]
- Teixeira, C.M.; Rosen, Z.B.; Suri, D.; Sun, Q.; Hersh, M.; Sargin, D.; Dincheva, I.; Morgan, A.A.; Spivack, S.; Krok, A.C.; et al. Hippocampal 5-HT input regulates memory formation and Schaffer collateral excitation. Neuron 2018, 98, 992–1004.e4. [Google Scholar] [CrossRef] [PubMed]
- Kempadoo, K.A.; Mosharov, E.V.; Choi, S.J.; Sulzer, D.; Kandel, E.R. Dopamine release from the locus coeruleus to the dorsal hippocampus promotes spatial learning and memory. Proc. Natl. Acad. Sci. USA 2016, 113, 14835–14840. [Google Scholar] [CrossRef] [PubMed]
- Zurkovsky, L.; Bychkov, E.; Tsakem, E.L.; Siedlecki, C.; Blakely, R.D.; Gurevich, E.V. Cognitive effects of dopamine depletion in the context of diminished acetylcholine signaling capacity in mice. Dis. Model Mech. 2013, 6, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Sahay, A.; Scobie, K.N.; Hill, A.S.; O’Carroll, C.M.; Kheirbek, M.A.; Burghardt, N.S.; Fenton, A.A.; Dranovsky, A.; Hen, R. Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation. Nature 2011, 472, 466–470. [Google Scholar] [CrossRef]
- Kirksey, A.; Morré, D.M.; Wasynczuk, A.Z. Neuronal development in vitamin B6 deficiency. Ann. N. Y. Acad. Sci. 1990, 585, 202–218. [Google Scholar] [CrossRef]
- Wagner, C.A.; Finberg, K.E.; Breton, S.; Marshansky, V.; Brown, D.; Geibel, J.P. Renal vacuolar H+-ATPase. Physiol. Rev. 2004, 84, 1263–1314. [Google Scholar] [CrossRef]
- Woody, S.K.; Zhou, H.; Ibrahimi, S.; Dong, Y.; Zhao, L. Human ApoE ε2 promotes regulatory mechanisms of bioenergetic and synaptic function in female brain: A focus on V-type H+-ATPase. J. Alzheimers Dis. 2016, 53, 1015–1031. [Google Scholar] [CrossRef]
- Zhao, W.; Gao, X.; Qiu, S.; Gao, B.; Gao, S.; Zhang, X.; Kang, D.; Han, W.; Dai, P.; Yuan, Y. A subunit of V-ATPases, ATP6V1B2, underlies the pathology of intellectual disability. EBioMedicine 2019, 45, 408–421. [Google Scholar] [CrossRef]
- Gonda, X.; Eszlari, N.; Anderson, I.M.; Deakin, J.F.; Bagdy, G.; Juhasz, G. Association of ATP6V1B2 rs1106634 with lifetime risk of depression and hippocampal neurocognitive deficits: Possible novel mechanisms in the etiopathology of depression. Transl. Psychiatry 2016, 6, e945. [Google Scholar] [CrossRef]
- Piedrahita, D.; Castro-Alvarez, J.F.; Boudreau, R.L.; Villegas-Lanau, A.; Kosik, K.S.; Gallego-Gomez, J.C.; Cardona-Gómez, G.P. β-Secretase 1’s targeting reduces hyperphosphorilated Tau, implying autophagy actors in 3xTg-AD mice. Front. Cell. Neurosci. 2016, 9, 498. [Google Scholar] [CrossRef]
- Yang, X.; Tohda, C. Heat shock cognate 70 inhibitor, VER-155008, reduces memory deficits and axonal degeneration in a mouse model of Alzheimer’s disease. Front. Pharmacol. 2018, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Abisambra, J.; Jinwal, U.K.; Miyata, Y.; Rogers, J.; Blair, L.; Li, X.; Seguin, S.P.; Wang, L.; Jin, Y.; Bacon, J.; et al. Allosteric heat shock protein 70 inhibitors rapidly rescue synaptic plasticity deficits by reducing aberrant tau. Biol. Psychiatry 2013, 74, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, S.N.; Martin, M.D.; Akoury, E.; Assimon, V.A.; Borysov, S.; Nordhues, B.A.; Sabbagh, J.J.; Cockman, M.; Gestwicki, J.E.; Zweckstetter, M.; et al. The active Hsc70/tau complex can be exploited to enhance tau turnover without damaging microtubule dynamics. Hum. Mol. Genet. 2015, 24, 3971–3981. [Google Scholar] [CrossRef] [PubMed]
- Jinwal, U.K.; Akoury, E.; Abisambra, J.F.; O’Leary, J.C., 3rd; Thompson, A.D.; Blair, L.J.; Jin, Y.; Bacon, J.; Nordhues, B.A.; Cockman, M.; et al. Imbalance of Hsp70 family variants fosters tau accumulation. FASEB J. 2013, 27, 1450–1459. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Proteins (gI Accession Number) | Number of Peptides Matched in the Identified Protein | Protein Sequence Coverage (%) | pI, Mr (kDa) | MOWSE | Fold Decrease |
|---|---|---|---|---|---|
| APG-1 (705391) | 1 | 1 | 5.53, 95.3 | 39 | |
| Zinc finger protein 831 (150010581) | 1 | 0 | 8.18 176.1 | 40 | 2.3 |
| Murine valosin-containing protein (55217) | 3 | 3 | 5.14 89.9 | 105 | 2.5 |
| Heat-shock protein hsp84 (194027) | 2 | 3 | 4.95 83.5 | 89 | 2.0 |
| V-type proton ATPase subunit B2 (17105370) | 38 | 25 | 5.57 56.9 | 638 | 2.3 |
| Vacuolar adenosine triphosphatase subunit B (1184661) | 1 | 2 | 5.57 56.9 | 45 | 3.1 |
| Acyltransferase (1129118) | 1 | 2 | 8.88 53.5 | 48 | 2.8 |
| Anti-DNA immunoglobulin light chain IgG (1870366) | 1 | 8 | 9.11 11.1 | 39 | 2.1 |
| Poly(rC)-binding protein 1 (6754994) | 1 | 2 | 6.66 38.0 | 40 | 2.2 |
| Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta-1 (6680045) | 12 | 12 | 5.60 38.2 | 243 | 8.8 |
| Glyceraldehyde-3-phosphate dehydrogenase (2494630) | 1 | 1 | 8.14 48.4 | 34 | 3.0 |
| Charged multivesicular body protein 4b (2807749) | 2 | 11 | 4.76 24.9 | 85 | 2.1 |
| Voltage-dependent anion-selective channel protein 2 (6755965) | 3 | 7 | 7.44 32.3 | 102 | 3.4 |
| Serine-threonine kinase receptor-associated protein (4063383) | 1 | 3 | 4.99 38.8 | 36 | 2.8 |
| Osmotic stress protein 94 (1098541) | 5 | 5 | 5.50 95.2 | 215 | 3.3 |
| Proteins (gI Accession Number) | Number of Peptides Matched in the Identified Protein | Protein Sequence Coverage (%) | pI, Mr (kDa) | MOWSE | Fold Increase |
|---|---|---|---|---|---|
| NADH dehydrogenase (13879366) | 2 | 2 | 5.51 80.7 | 68 | 2.0 |
| Heat shock cognate protein 70 (309319) | 49 | 29 | 5.37 71.0 | 1066 | 2.4 |
| Heat shock-related 70 kDa protein 2 (31560686) | 20 | 21 | 5.51 69.9 | 620 | 3.5 |
| Dihydrolipoamide dehydrogenase (2078522) | 1 | 2 | 7.97 54.7 | 41 | 2.3 |
| ATP synthase subunit alpha, mitochondrial precursor (6680748) | 10 | 9 | 9.22 59.8 | 308 | 2.7 |
| Aldehyde dehydrogenase, mitochondrial isoform 1 precursor (6753036) | 2 | 3 | 7.53 57.0 | 96 | 2.0 |
| Type II keratin subunit protein (4159806) | 1 | 1 | 8.97 65.7 | 40 | 2.8 |
| Spermatogenesis-associated protein 7 homolog isoform 1 | 1 | 1 | 6.23 66.2 | 34 | 3.0 |
| ATP-specific succinyl-CoA synthetase beta subunit | 16 | 13 | 5.65 46.6 | 366 | 3.6 |
| Protein phosphatase type 2A catalytic subunit alpha isoform (3342500) | 1 | 2 | 5.30 36.2 | 48 | 2.4 |
| Histidine triad nucleotide-binding protein 1 (33468857) | 1 | 11 | 6.36 13.9 | 41 | 2.1 |
| Epidermal keratin subunit I, partial (387397) | 17 | 8 | 5.01 58.0 | 304 | 4.0 |
| 2-Oxoglutarate dehydrogenase-like, mitochondrial (568987556) | 2 | 83.2 | 96 | 2.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, H.Y.; Kim, W.; Hahn, K.R.; Kwon, H.J.; Nam, S.M.; Chung, J.Y.; Yoon, Y.S.; Kim, D.W.; Yoo, D.Y.; Hwang, I.K. Effects of Pyridoxine Deficiency on Hippocampal Function and Its Possible Association with V-Type Proton ATPase Subunit B2 and Heat Shock Cognate Protein 70. Cells 2020, 9, 1067. https://doi.org/10.3390/cells9051067
Jung HY, Kim W, Hahn KR, Kwon HJ, Nam SM, Chung JY, Yoon YS, Kim DW, Yoo DY, Hwang IK. Effects of Pyridoxine Deficiency on Hippocampal Function and Its Possible Association with V-Type Proton ATPase Subunit B2 and Heat Shock Cognate Protein 70. Cells. 2020; 9(5):1067. https://doi.org/10.3390/cells9051067
Chicago/Turabian StyleJung, Hyo Young, Woosuk Kim, Kyu Ri Hahn, Hyun Jung Kwon, Sung Min Nam, Jin Young Chung, Yeo Sung Yoon, Dae Won Kim, Dae Young Yoo, and In Koo Hwang. 2020. "Effects of Pyridoxine Deficiency on Hippocampal Function and Its Possible Association with V-Type Proton ATPase Subunit B2 and Heat Shock Cognate Protein 70" Cells 9, no. 5: 1067. https://doi.org/10.3390/cells9051067
APA StyleJung, H. Y., Kim, W., Hahn, K. R., Kwon, H. J., Nam, S. M., Chung, J. Y., Yoon, Y. S., Kim, D. W., Yoo, D. Y., & Hwang, I. K. (2020). Effects of Pyridoxine Deficiency on Hippocampal Function and Its Possible Association with V-Type Proton ATPase Subunit B2 and Heat Shock Cognate Protein 70. Cells, 9(5), 1067. https://doi.org/10.3390/cells9051067