Inflammation Alters the Secretome and Immunomodulatory Properties of Human Skin-Derived Precursor Cells

 ,
,  , , , ,
, , , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Cultivation of Human SKP
2.2. Pro-Inflammatory Stimulation
2.3. Acquisition and Activation of Immune Cells for In Vitro Assays
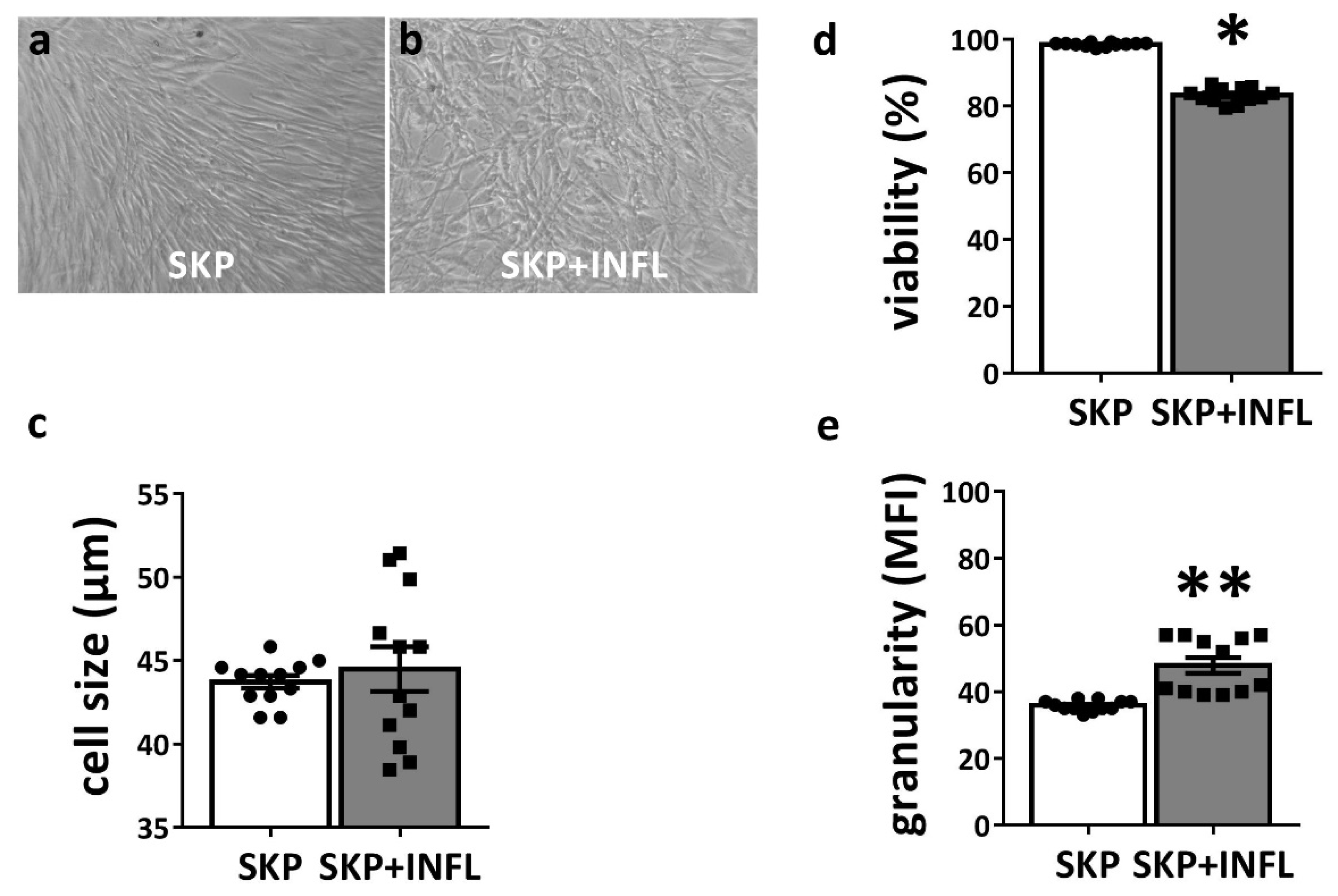
2.4. Morphology
2.5. Viability Assay
2.6. Cell Size and Granularity
2.7. Microarray Data Analysis
2.8. Immunogenicity Assay
2.9. Secretion Assay
2.10. Cotransplantation of PBMC and SKP in SCID Mice
2.11. Statistical Analysis
3. Results
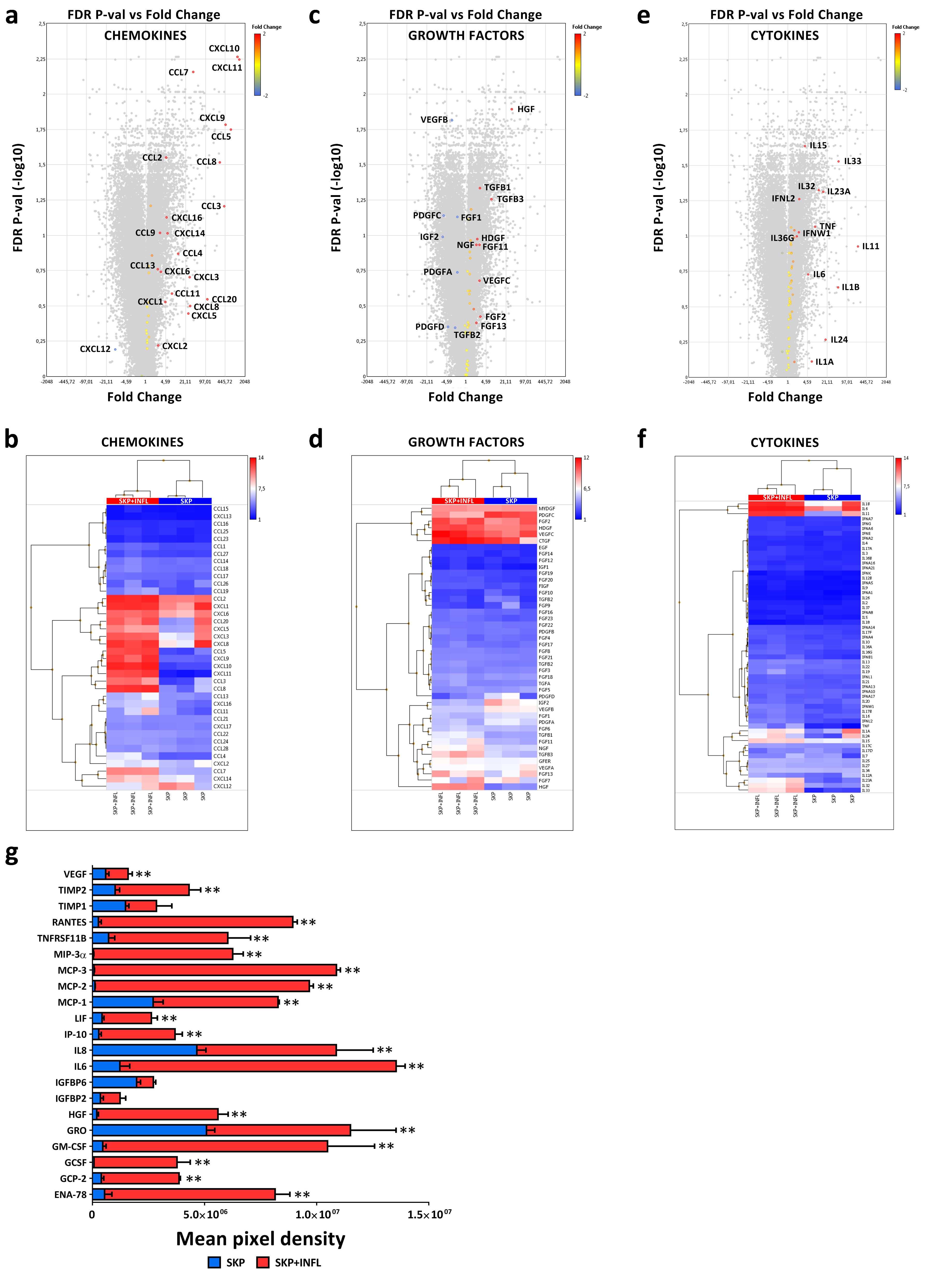
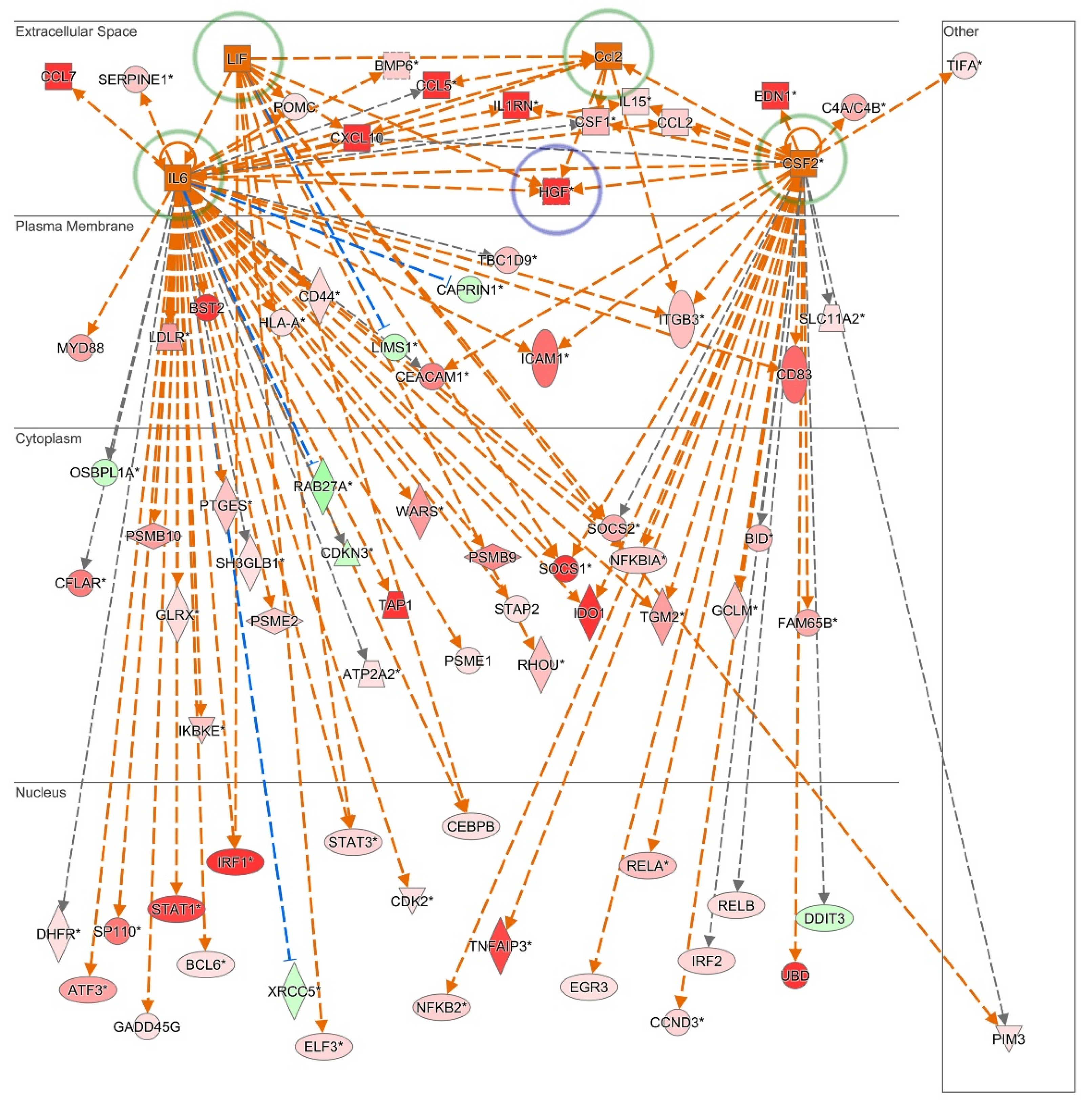
3.1. Inflammation Alters the Cytokine and Growth Factor Secretion Profile of SKP
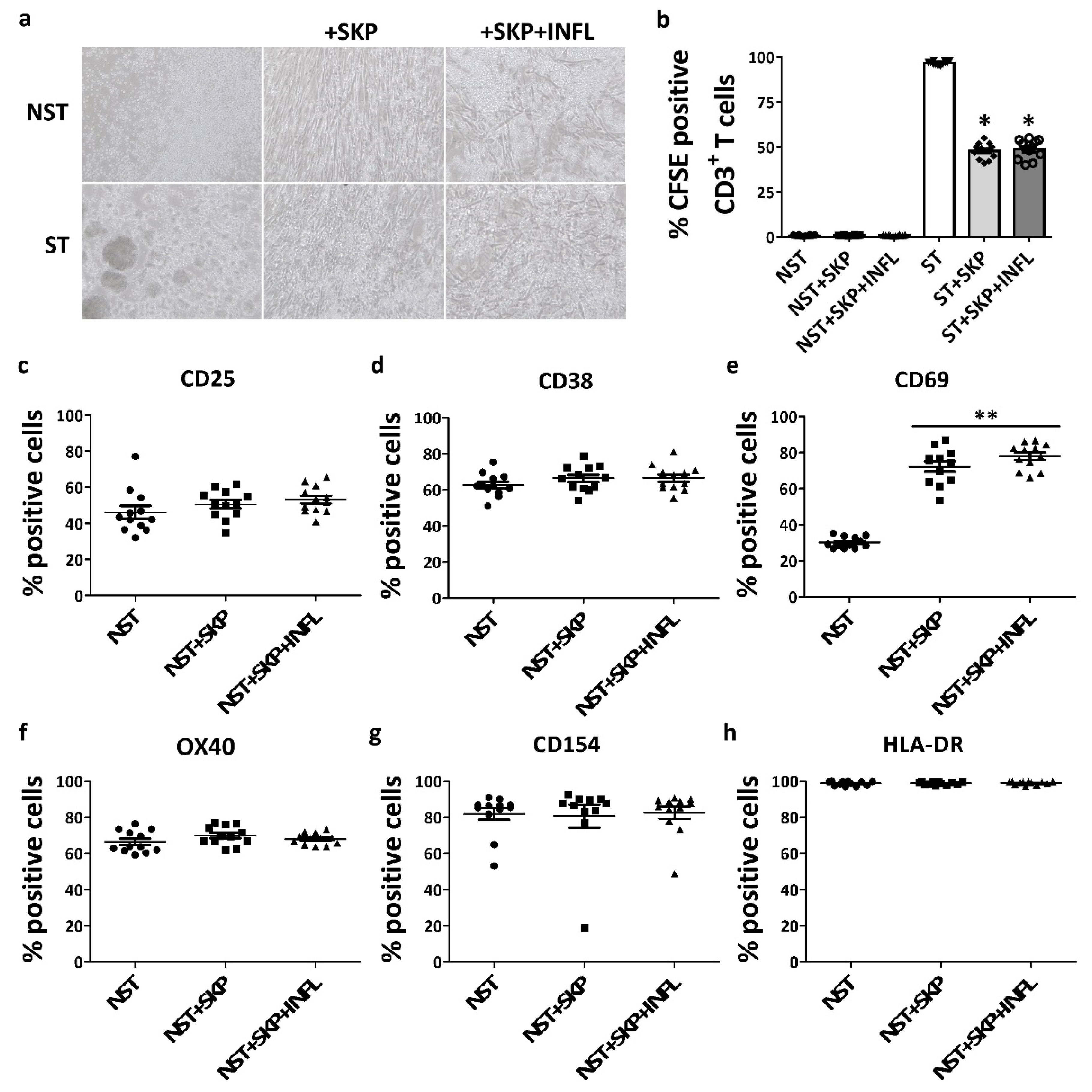
3.2. SKP Remain Immunosuppressive Upon In Vitro Pro-inflammatory Stimulation
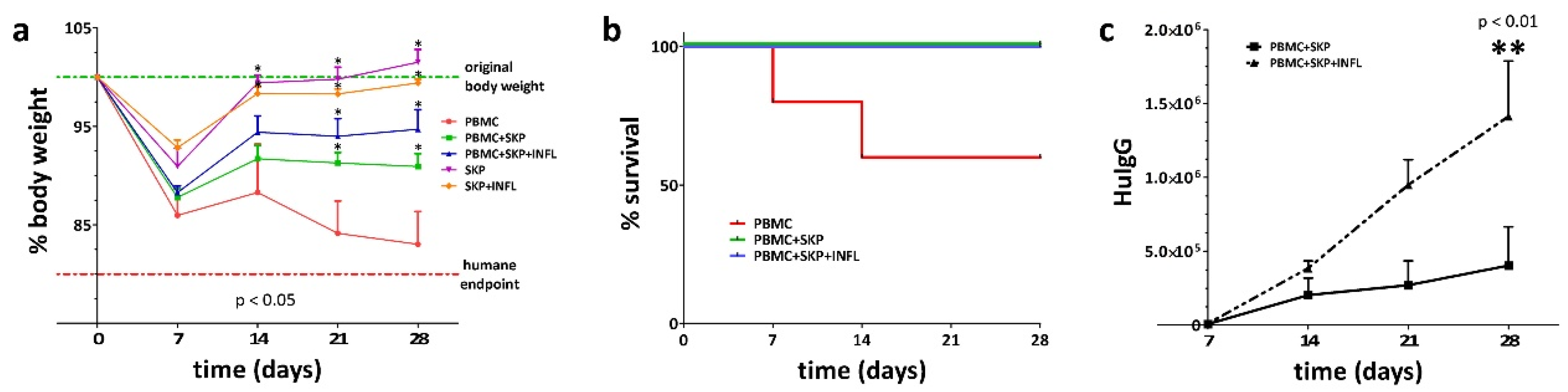
3.3. Inflammation Alters the Immunosuppressive Properties of SKP In Vivo
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hunt, D.P.J.; Jahoda, C.; Chandran, S. Multipotent skin-derived precursors: From biology to clinical translation. Curr. Opin. Biotechnol. 2009, 20, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Toma, J.G.; McKenzie, I.A.; Bagli, D.; Miller, F.D. Isolation and Characterization of Multipotent Skin-Derived Precursors from Human Skin. Stem Cells 2005, 23, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Gago, N.; Pérez-López, V.; Sanz-Jaka, J.P.; Cormenzana, P.; Eizaguirre, I.; Bernad, A.; Izeta, A. Age-dependent depletion of human skin-derived progenitor cells. Stem Cells 2009, 27, 1164–1172. [Google Scholar] [CrossRef] [PubMed]
- Buranasinsup, S.; Sila-Asna, M.; Bunyaratvej, N.; Bunyaratvej, A. In vitro osteogenesis from human skin-derived precursor cells. Dev. Growth Differ. 2006, 48, 263–269. [Google Scholar] [CrossRef] [PubMed]
- De Kock, J.; Rodrigues, R.M.; Buyl, K.; Vanhaecke, T.; Rogiers, V. Human skin-derived precursor cells: Isolation, expansion, and hepatic differentiation. In Methods in Molecular Biology; Springer: New York, NY, USA, 2014; Volume 1250, pp. 113–122. ISBN 9781493920747. [Google Scholar]
- Shih, D.T.; Lee, D.-C.; Chen, S.-C.; Tsai, R.-Y.; Huang, C.-T.; Tsai, C.-C.; Shen, E.-Y.; Chiu, W.-T. Isolation and Characterization of Neurogenic Mesenchymal Stem Cells in Human Scalp Tissue. Stem Cells 2005, 23, 1012–1020. [Google Scholar] [CrossRef] [PubMed]
- Biernaskie, J.; Sparling, J.S.; Liu, J.; Shannon, C.P.; Plemel, J.R.; Xie, Y.; Miller, F.D.; Tetzlaff, W. Skin-Derived Precursors Generate Myelinating Schwann Cells That Promote Remyelination and Functional Recovery after Contusion Spinal Cord Injury. J. Neurosci. 2007, 27, 9545–9559. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, J.-F.; Biernaskie, J.A.; Chen, Y.; Bagli, D.; Alman, B.; Kaplan, D.R.; Miller, F.D. Skin-Derived Precursors Differentiate Into Skeletogenic Cell Types and Contribute to Bone Repair. Stem Cells Dev. 2009, 18, 893–906. [Google Scholar] [CrossRef]
- Rodrigues, R.M.; De Kock, J.; Rogiers, V.; Vanhaecke, T. Human skin-derived stem cells as a novel cell source for in vitro hepatotoxicity screening of pharmaceuticals. Toxicol. Lett. 2015, 238, S186. [Google Scholar] [CrossRef][Green Version]
- De Kock, J.; Najar, M.; Bolleyn, J.; Al Battah, F.; Rodrigues, R.M.; Buyl, K.; Raicevic, G.; Govaere, O.; Branson, S.; Meganathan, K.; et al. Mesoderm-derived stem cells: The link between the transcriptome and their differentiation potential. Stem Cells Dev. 2012, 21, 3309–3323. [Google Scholar] [CrossRef]
- Etxaniz, U.; Vicente, A.P.S.; Gago-López, N.; Garciá-Dominguez, M.; Iribar, H.; Aduriz, A.; Pérez-López, V.; Burgoa, I.; Irizar, H.; Muñoz-Culla, M.; et al. Neural-competent cells of adult human dermis belong to the Schwann lineage. Stem Cell Rep. 2014, 3, 774–788. [Google Scholar] [CrossRef]
- Jinno, H.; Morozova, O.; Jones, K.L.; Biernaskie, J.A.; Paris, M.; Hosokawa, R.; Rudnicki, M.A.; Chai, Y.; Rossi, F.; Marra, M.A.; et al. Convergent Genesis of an Adult Neural Crest-Like Dermal Stem Cell from Distinct Developmental Origins. Stem Cells 2010, 28, 2027–2040. [Google Scholar] [CrossRef] [PubMed]
- Iribar, H.; Pérez-López, V.; Etxaniz, U.; Gutiérrez-Rivera, A.; Izeta, A. Schwann Cells in the Ventral Dermis Do Not Derive from Myf5-Expressing Precursors. Stem Cell Rep. 2017, 9, 1477–1487. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, I.A.; Biernaskie, J.; Toma, J.G.; Midha, R.; Miller, F.D. Skin-Derived Precursors Generate Myelinating Schwann Cells for the Injured and Dysmyelinated Nervous System. J. Neurosci. 2006, 26, 6651–6660. [Google Scholar] [CrossRef] [PubMed]
- Sparling, J.S.; Bretzner, F.; Biernaskie, J.; Assinck, P.; Jiang, Y.; Arisato, H.; Plunet, W.T.; Borisoff, J.; Liu, J.; Miller, F.D.; et al. Schwann Cells Generated from Neonatal Skin-Derived Precursors or Neonatal Peripheral Nerve Improve Functional Recovery after Acute Transplantation into the Partially Injured Cervical Spinal Cord of the Rat. J. Neurosci. 2015, 35, 6714–6730. [Google Scholar] [CrossRef]
- Fernandes, K.J.L.; Kobayashi, N.R.; Gallagher, C.J.; Barnabé-Heider, F.; Aumont, A.; Kaplan, D.R.; Miller, F.D. Analysis of the neurogenic potential of multipotent skin-derived precursors. Exp. Neurol. 2006, 201, 32–48. [Google Scholar] [CrossRef]
- Higashida, T.; Jitsuki, S.; Kubo, A.; Mitsushima, D.; Kamiya, Y.; Kanno, H. Skin-derived precursors differentiating into dopaminergic neuronal cells in the brains of Parkinson disease model rats. J. Neurosurg. 2010, 113, 648–655. [Google Scholar] [CrossRef]
- Wagner, J.P.; Sullins, V.F.; Dunn, J.C.Y. Skin-derived precursors generate enteric-type neurons in aganglionic jejunum. J. Pediatr. Surg. 2014, 49, 1809–1814. [Google Scholar] [CrossRef][Green Version]
- Steinbach, S.K.; El-Mounayri, O.; DaCosta, R.S.; Frontini, M.J.; Nong, Z.; Maeda, A.; Pickering, J.G.; Miller, F.D.; Husain, M. Directed Differentiation of Skin-Derived Precursors Into Functional Vascular Smooth Muscle Cells. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2938–2948. [Google Scholar] [CrossRef][Green Version]
- Rodrigues, R.M.; Branson, S.; De Boe, V.; Sachinidis, A.; Rogiers, V.; De Kock, J.; Vanhaecke, T. In vitro assessment of drug-induced liver steatosis based on human dermal stem cell-derived hepatic cells. Arch. Toxicol. 2015, 90, 677–689. [Google Scholar] [CrossRef]
- De Kock, J.; Vanhaecke, T.; Biernaskie, J.; Rogiers, V.; Snykers, S. Characterization and hepatic differentiation of skin-derived precursors from adult foreskin by sequential exposure to hepatogenic cytokines and growth factors reflecting liver development. Toxicol. In Vitro 2009, 23, 1522–1527. [Google Scholar] [CrossRef]
- Mehrabi, M.; Mansouri, K.; Hosseinkhani, S.; Yarani, R.; Yari, K.; Bakhtiari, M.; Mostafaie, A. Differentiation of human skin-derived precursor cells into functional islet-like insulin-producing cell clusters. In Vitro Cell. Dev. Biol. Anim. 2015, 51, 595–603. [Google Scholar] [CrossRef] [PubMed]
- De Kock, J.; Meuleman, P.; Raicevic, G.; Rodrigues, R.M.; Branson, S.; Meganathan, K.; De Boe, V.; Sachinidis, A.; Leroux-Roels, G.; Vanhaecke, T.; et al. Human skin-derived precursor cells are poorly immunogenic and modulate the allogeneic immune response. Stem Cells 2014, 32, 2215–2228. [Google Scholar] [CrossRef]
- Krause, M.; Dworski, S.; Feinberg, K.; Jones, K.; Johnston, A.W.; Paul, S.; Paris, M.; Peles, E.; Bagli, D.; Forrest, C.; et al. Direct Genesis of Functional Rodent and Human Schwann Cells from Skin Mesenchymal Precursors. Stem Cell Rep. 2014, 3, 85–100. [Google Scholar] [CrossRef]
- Agabalyan, N.A.; Rosin, N.L.; Rahmani, W.; Biernaskie, J. Hair follicle dermal stem cells and skin-derived precursor cells: Exciting tools for endogenous and exogenous therapies. Exp. Dermatol. 2017, 26, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, L.; Busuttil, V.; Botto, J.M. Multipotentiality of skin-derived precursors: Application to the regeneration of skin and other tissues. Int. J. Cosmet. Sci. 2020, 42, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, X.; Liu, J.; Cai, T.; Guo, L.; Wang, S.; Wang, J.; Cao, Y.; Ge, J.; Jiang, Y.; et al. Hair Follicle and Sebaceous Gland De Novo Regeneration With Cultured Epidermal Stem Cells and Skin-Derived Precursors. Stem Cells Transl. Med. 2016, 5, 1695–1706. [Google Scholar] [CrossRef]
- Bayati, V.; Abbaspour, M.R.; Neisi, N.; Hashemitabar, M. Skin-derived precursors possess the ability of differentiation into the epidermal progeny and accelerate burn wound healing. Cell Biol. Int. 2017, 41, 187–196. [Google Scholar] [CrossRef]
- Faden, A.I.; Wu, J.; Stoica, B.A.; Loane, D.J. Progressive inflammation-mediated neurodegeneration after traumatic brain or spinal cord injury. Br. J. Pharmacol. 2015, 173, 681–691. [Google Scholar] [CrossRef]
- Szabo, G.; Petrasek, J. Inflammasome activation and function in liver disease. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 387–400. [Google Scholar] [CrossRef]
- Goldring, S.R. Inflammatory signaling induced bone loss. Bone 2015, 80, 143–149. [Google Scholar] [CrossRef]
- Yang, Y.H.K.; Ogando, C.R.; Wang See, C.; Chang, T.Y.; Barabino, G.A. Changes in phenotype and differentiation potential of human mesenchymal stem cells aging in vitro. Stem Cell Res. Ther. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Raicevic, G.; Rouas, R.; Najar, M.; Stordeur, P.; Boufker, H.I.; Bron, D.; Martiat, P.; Goldman, M.; Nevessignsky, M.T.; Lagneaux, L. Inflammation modifies the pattern and the function of Toll-like receptors expressed by human mesenchymal stromal cells. Hum. Immunol. 2010, 71, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Irizarry, R.A.; Bolstad, B.M.; Collin, F.; Cope, L.M.; Hobbs, B.; Speed, T.P. Summaries of Affymetrix GeneChip probe level data. Nucleic Acids Res. 2003, 31, e15. [Google Scholar] [CrossRef] [PubMed]
- Depraetere, S.; Verhoye, L.; Leclercq, G.; Leroux-Roels, G. Human B Cell Growth and Differentiation in the Spleen of Immunodeficient Mice. J. Immunol. 2001, 166, 2929–2936. [Google Scholar] [CrossRef]
- Tournoy, K.G.; Depraetere, S.; Meuleman, P.; Leroux-Roels, G.; Pauwels, R.A. Murine IL-2 receptor beta chain blockade improves human leukocyte engraftment in SCID mice. Eur. J. Immunol. 1998, 28, 3221–3230. [Google Scholar] [CrossRef]
- Sharpe, M.E.; Morton, D.; Rossi, A. Nonclinical safety strategies for stem cell therapies. Toxicol. Appl. Pharmacol. 2012, 262, 223–231. [Google Scholar] [CrossRef]
- Le Blanc, K.; Davies, L.C. Mesenchymal stromal cells and the innate immune response. Immunol. Lett. 2015, 168, 140–146. [Google Scholar] [CrossRef]
- Najar, M.; Bouhtit, F.; Melki, R.; Afif, H.; Hamal, A.; Fahmi, H.; Merimi, M.; Lagneaux, L. Mesenchymal Stromal Cell-Based Therapy: New Perspectives and Challenges. J. Clin. Med. 2019, 8, 626. [Google Scholar] [CrossRef]
- Buyl, K.; Merimi, M.; Rodrigues, R.M.; Agha, D.M.; Melki, R.; Vanhaecke, T.; Bron, D.; Lewalle, P.; Meuleman, N.; Fahmi, H.; et al. The Impact of Cell-Expansion and Inflammation on The Immune-Biology of Human Adipose Tissue-Derived Mesenchymal Stromal Cells. J. Clin. Med. 2020, 9, 696. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Cao, W.; Shi, Y. Plasticity of mesenchymal stem cells in immunomodulation: Pathological and therapeutic implications. Nat. Immunol. 2014, 15, 1009–1016. [Google Scholar] [CrossRef]
- Sancho, D.; Gómez, M.; Viedma, F.; Esplugues, E.; Gordón-Alonso, M.; García-López, M.A.; De La Fuente, H.; Martínez-A, C.; Lauzurica, P.; Sánchez-Madrid, F. CD69 downregulates autoimmune reactivity through active transforming growth factor-β production in collagen-induced arthritis. J. Clin. Investig. 2003, 112, 872–882. [Google Scholar] [CrossRef]
- Cortés, J.R.; Sánchez-Díaz, R.; Bovolenta, E.R.; Barreiro, O.; Lasarte, S.; Matesanz-Marín, A.; Toribio, M.L.; Sánchez-Madrid, F.; Martín, P. Maintenance of immune tolerance by Foxp3+ regulatory T cells requires CD69 expression. J. Autoimmun. 2014, 55, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Carrero, R.; Cerrada, I.; Lledó, E.; Dopazo, J.; García-García, F.; Rubio, M.P.; Trigueros, C.; Dorronsoro, A.; Ruiz-Sauri, A.; Montero, J.A.; et al. IL1β Induces Mesenchymal Stem Cells Migration and Leucocyte Chemotaxis Through NF-κB. Stem Cell Rev. Rep. 2012, 8, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Zhang, L.; Zhao, X.; Xu, G.; Zhang, Y.; Roberts, A.I.; Zhao, R.C.; Shi, Y. Mesenchymal Stem Cell-Mediated Immunosuppression Occurs via Concerted Action of Chemokines and Nitric Oxide. Cell Stem Cell 2008, 2, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Buchsel, P.C.; Forgey, A.; Grape, F.B.; Hamann, S.S. Granulocyte Macrophage Colony-Stimulating Factor: Current Practice and Novel Approaches. Clin. J. Oncol. Nurs. 2002, 6, 198–205. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Benkhoucha, M.; Santiago-Raber, M.L.; Schneiter, G.; Chofflon, M.; Funakoshi, H.; Nakamura, T.; Lalive, P.H. Hepatocyte growth factor inhibits CNS autoimmunity by inducing tolerogenic dendritic cells and CD25+Foxp3+ regulatory T cells. Proc. Natl. Acad. Sci. USA 2010, 107, 6424–6429. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Sakai, K.; Nakamura, T.; Matsumoto, K. Hepatocyte growth factor twenty years on: Much more than a growth factor. J. Gastroenterol. Hepatol. 2011, 26, 188–202. [Google Scholar] [CrossRef]
- Okunishi, K.; Dohi, M.; Fujio, K.; Nakagome, K.; Tabata, Y.; Okasora, T.; Seki, M.; Shibuya, M.; Imamura, M.; Harada, H.; et al. Hepatocyte Growth Factor Significantly Suppresses Collagen-Induced Arthritis in Mice. J. Immunol. 2007, 179, 5504–5513. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Expression | Protein Secretion | |||||||
|---|---|---|---|---|---|---|---|---|
| Gene | Probeset | Fold Change | FDR p-Value | Protein | SKP | SKP+INFL | Fold Change | p-Value |
| CCL2 (*) | 216598_s_at | 4.7 | 2.81 × 10−2 | MCP-1 |  |  | 3.1 | 2.00 × 10−4 |
| CCL5 | 1405_i_at | 676.8 | 1.78 × 10−2 | RANTES |  |  | 34.2 | 1.00 × 10−4 |
| CCL7 | 208075_s_at | 37.6 | 7.00 × 10−3 | MCP-3 |  |  | 159.6 | 1.00 × 10−4 |
| CCL8 | 214038_at | 289.3 | 3.04 × 10−2 | MCP-2 |  |  | 85.2 | 1.00 × 10−4 |
| CCL20 | 205476_at | 111.2 | 2.85 × 10−1 | MIP-3α |  |  | 147.5 | 2.00 × 10−4 |
| CXCL1 | 204470_at | 4.4 | 2.97 × 10−1 | GRO |  |  | 2.3 | 3.63 × 10−2 |
| CXCL5 | 215101_s_at | 25.8 | 3.57 × 10−1 | ENA-78 |  |  | 15.2 | 5.00 × 10−4 |
| CXCL6 | 206336_at | 3.2 | 1.82 × 10−1 | GCP-2 |  |  | 9.8 | 1.00 × 10−4 |
| CXCL10 | 204533_at | 1135.8 | 5.50 × 10−3 | IP-10 |  |  | 13.2 | 6.00 × 10−4 |
| CSF2 (*) | 210229_s_at | 39.1 | 3.98 × 10−1 | GM-CSF |  |  | 22.9 | 8.80 × 10−3 |
| CSF3 | 207442_at | 38.0 | 1.00 × 10−1 | GCSF |  |  | 54.7 | 3.40 × 10−3 |
| HGF | 210997_at | 32.9 | 1.28 × 10−2 | HGF |  |  | 29.3 | 3.00 × 10−4 |
| IGFBP2 | 202718_at | 5.2 | 1.04 × 10−1 | IGFBP2 |  |  | 3.5 | 3.59 × 10−2 |
| IGFBP6 | 203851_at | 1.0 | 5.27 × 10−1 | IGFBP6 |  |  | 1.4 | 2.27 × 10−2 |
| IL6 (*) | 205207_at | 4.9 | 1.87 × 10−1 | IL6 |  |  | 11.1 | 1.00 × 10−4 |
| CXCL8 | 211506_s_at | 29.7 | 3.17 × 10−1 | IL8 |  |  | 2.3 | 2.17 × 10−2 |
| LIF (*) | 205266_at | 17.7 | 2.05 × 10−1 | LIF |  |  | 6.2 | 1.50 × 10−3 |
| TIMP1 | 201666_at | 2.1 | 6.81 × 10−2 | TIMP-1 |  |  | 1.9 | 1.24 × 10−1 |
| TIMP2 | 224560_at | −1.4 | 2.37 × 10−1 | TIMP-2 |  |  | 4.3 | 4.00 × 10−3 |
| TNFRSF11B | 204933_s_at | −5.2 | 9.46 × 10−1 | OPG |  |  | 8.4 | 7.20 × 10−3 |
| VEGFA | 210512_s_at | −1.5 | 2.73 × 10−1 | VEGF |  |  | 2.7 | 1.43 × 10−2 |
| VEGFB | 203683_s_at | −3.0 | 1.53 × 10−2 | |||||
| VEGFC | 209946_at | 2.8 | 6.88 × 10−2 | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Kock, J.; Rodrigues, R.M.; Branson, S.; Verhoye, L.; Colemonts-Vroninks, H.; Rombaut, M.; Boeckmans, J.; Neuckermans, J.; Lequeue, S.; Buyl, K.; et al. Inflammation Alters the Secretome and Immunomodulatory Properties of Human Skin-Derived Precursor Cells. Cells 2020, 9, 914. https://doi.org/10.3390/cells9040914
De Kock J, Rodrigues RM, Branson S, Verhoye L, Colemonts-Vroninks H, Rombaut M, Boeckmans J, Neuckermans J, Lequeue S, Buyl K, et al. Inflammation Alters the Secretome and Immunomodulatory Properties of Human Skin-Derived Precursor Cells. Cells. 2020; 9(4):914. https://doi.org/10.3390/cells9040914
Chicago/Turabian StyleDe Kock, Joery, Robim Marcelino Rodrigues, Steven Branson, Lieven Verhoye, Haaike Colemonts-Vroninks, Matthias Rombaut, Joost Boeckmans, Jessie Neuckermans, Sien Lequeue, Karolien Buyl, and et al. 2020. "Inflammation Alters the Secretome and Immunomodulatory Properties of Human Skin-Derived Precursor Cells" Cells 9, no. 4: 914. https://doi.org/10.3390/cells9040914
APA StyleDe Kock, J., Rodrigues, R. M., Branson, S., Verhoye, L., Colemonts-Vroninks, H., Rombaut, M., Boeckmans, J., Neuckermans, J., Lequeue, S., Buyl, K., Merimi, M., Agha, D. M., De Boe, V., Lagneaux, L., Meuleman, P., Vanhaecke, T., & Najar, M. (2020). Inflammation Alters the Secretome and Immunomodulatory Properties of Human Skin-Derived Precursor Cells. Cells, 9(4), 914. https://doi.org/10.3390/cells9040914