Focusing on Adenosine Receptors as a Potential Targeted Therapy in Human Diseases


Abstract
1. Introduction
2. Adenosine: Production, Transport, and Metabolism
3. Adenosine Receptors
3.1. A1 Adenosine Receptor (A1AR)
3.2. A2A Adenosine Receptor (A2AAR)
3.3. A2B Adenosine Receptor (A2BAR)
3.4. A3 Adenosine Receptor (A3AR)
4. Adenosine Receptors and Diseases
4.1. A1AR
4.1.1. A1AR in Inflammation
4.1.2. A1AR in the Respiratory Systems
4.1.3. A1AR in the Cardiovascular Systems
4.1.4. A1AR in CNS
4.1.5. A1AR in Metabolic and other Diseases
4.2. A2AAR
4.2.1. A2AAR in Inflammation
4.2.2. A2AAR in the Respiratory System
4.2.3. A2AAR in the Cardiovascular Systems
4.2.4. A2AAR in the CNS
4.3. A2BAR
4.3.1. A2BAR in Inflammation
4.3.2. A2BAR in the Respiratory System
4.3.3. A2BAR in Cardiovascular Systems
4.3.4. A2BAR in Metabolic Diseases
4.3.5. A2BAR in Cancer
4.4. A3AR
4.4.1. A3AR in Inflammation
4.4.2. A3AR in the Respiratory System
4.4.3. A3AR in Cardiovascular Systems
4.4.4. A3AR in CNS Systems
4.4.5. A3AR in the Digestive and Renal Systems
4.4.6. A3AR in Cancer
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Drury, A.N.; Szent-Györgyi, A. The physiological activity of adenine compounds with especial reference to their action upon the mammalian heart. J. Physiol. 1929, 68, 213–237. [Google Scholar] [CrossRef] [PubMed]
- Cobbin, L.B.; Einstein, R.; Maguire, M.H. Studies on the coronary dilator actions of some adenosine analogues. Br. J. Pharmacol. 1974, 50, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Degubareff, T.; Sleator, W.J. Effects of caffeine on mammalian atrial muscle, and its interaction with adenosine and calcium. J. Pharmacol. Exp. Ther. 1965, 148, 202–214. [Google Scholar] [PubMed]
- Sattin, A.; Rall, T.W. The effect of adenosine and adenine nucleotides on the cyclic adenosine 3′, 5′-phosphate content of guinea pig cerebral cortex slices. Mol. Pharmacol. 1970, 6, 13–23. [Google Scholar]
- Burnstock, G. Purinergic nerves and receptors. Prog. Biochem. Pharmacol. 1980, 16, 141–154. [Google Scholar]
- Burnstock, G.; Verkhratsky, A. Purinergic signaling. Wiley Interdiscip. Rev. Membr. Transp. Signal. 2012, 1, 116–125. [Google Scholar] [CrossRef]
- Burnstock, G. Autonomic neurotransmitters and trophic factors. J. Auton. Nerv. Syst. 1983, 7, 213–217. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Abbracchio, M.P.; Burnstock, G.; Daly, J.W.; Harden, T.K.; Jacobson, K.A.; Leff, P.; Williams, M. Nomenclature and classification of purinoceptors. Pharmacol. Rev. 1994, 46, 143–156. [Google Scholar]
- Krishtal, O.A.; Marchenko, S.M.; Pidoplichko, V.I. Receptor for ATP in the membrane of mammalian sensory neurones. Neurosci. Lett. 1983, 35, 41–45. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Abbracchio, M.P.; Burnstock, G.; Dubyak, G.R.; Harden, T.K.; Jacobson, K.A.; Schwabe, U.; Williams, M. Towards a revised nomenclature for P1 and P2 receptors. Trends Pharmacol. Sci. 1997, 18, 79–82. [Google Scholar] [CrossRef]
- Fredholm, B.B.; IJzerman, A.P.; Jacobson, K.A.; Klotz, K.N.; Linden, J. International Union of Pharmacology. XXV. Nomenclature and classification of adenosine receptors. Pharmacol. Rev. 2001, 53, 527–552. [Google Scholar] [PubMed]
- Olah, M.E.; Stiles, G.L. Adenosine receptor subtypes: Characterization and therapeutic regulation. Annu. Rev. Pharmacol. Toxicol. 1995, 35, 581–606. [Google Scholar] [CrossRef] [PubMed]
- Shryock, J.C.; Belardinelli, L. Adenosine and adenosine receptors in the cardiovascular system: Biochemistry, physiology, and pharmacology. Am. J. Cardiol. 1997, 79, 2–10. [Google Scholar] [CrossRef]
- Spicuzza, L.; Di Maria, G.; Polosa, R. Adenosine in the airways: Implications and applications. Eur. J. Pharmacol. 2006, 533, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Sala-Newby, G.B.; Skladanowski, A.C.; Newby, A.C. The mechanism of adenosine formation in cells. Cloning of cytosolic 5′-nucleotidase-I. J. Biol. Chem. 1999, 274, 17789–17793. [Google Scholar] [CrossRef]
- Sala-Newby, G.B.; Freeman, N.V.E.; Skladanowski, A.C.; Newby, A.C. Distinct Roles for Recombinant Cytosolic 5′-Nucleotidase-I and -II in AMP and IMP Catabolism in COS-7 and H9c2 Rat Myoblast Cell Lines. J. Biol. Chem. 2000, 275, 11666–11671. [Google Scholar] [CrossRef]
- Broch, O.J.; Ueland, P.M. Regional and subcellular distribution of S-adenosylhomocysteine hydrolase in the adult rat brain. J. Neurochem. 1980, 35, 484–488. [Google Scholar] [CrossRef]
- Kroll, K.; Decking, U.K.; Dreikorn, K.; Schrader, J. Rapid turnover of the AMP-adenosine metabolic cycle in the guinea pig heart. Circ. Res. 1993, 73, 846–856. [Google Scholar] [CrossRef]
- Layland, J.; Carrick, D.; Lee, M.; Oldroyd, K.; Berry, C. Adenosine: Physiology, pharmacology, and clinical applications. JACC Cardiovasc. Interv. 2014, 7, 581–591. [Google Scholar] [CrossRef]
- Saito, H.; Nishimura, M.; Shinano, H.; Makita, H.; Tsujino, I.; Shibuya, E.; Sato, F.; Miyamoto, K.; Kawakami, Y. Plasma concentration of adenosine during normoxia and moderate hypoxia in humans. Am. J. Respir. Crit. Care Med. 1999, 159, 1014–1018. [Google Scholar] [CrossRef]
- Latini, S.; Pedata, F. Adenosine in the central nervous system: Release mechanisms and extracellular concentrations. J. Neurochem. 2001, 79, 463–484. [Google Scholar] [CrossRef] [PubMed]
- Sheth, S.; Brito, R.; Mukherjea, D.; Rybak, L.P.; Ramkumar, V. Adenosine receptors: Expression, function and regulation. Int. J. Mol. Sci. 2014, 15, 2024–2052. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, M.R.; Vance, C.O.; Morschl, E.; Wilson, C.N. Adenosine receptors and inflammation. In Adenosine Receptors in Health and Disease; Wilson, C.N., Mustafa, S.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 215–269. [Google Scholar]
- Polosa, R.; Holgate, S.T. Adenosine receptors as promising therapeutic targets for drug development in chronic airway inflammation. Curr. Drug Targets 2006, 7, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Yegutkin, G.G. Nucleotide- and nucleoside-converting ectoenzymes: Important modulators of purinergic signalling cascade. Biochim. Biophys. Acta 2008, 1783, 673–694. [Google Scholar] [CrossRef] [PubMed]
- Eltzschig, H.K. Adenosine: An old drug newly discovered. Anesthesiology 2009, 111, 904–915. [Google Scholar] [CrossRef]
- Zimmermann, H. Biochemistry, localization and functional roles of ecto-nucleotidases in the nervous system. Prog. Neurobiol. 1996, 49, 589–618. [Google Scholar] [CrossRef]
- Marquardt, D.L.; Gruber, H.E.; Wasserman, S.I. Adenosine release from stimulated mast cells. Proc. Natl. Acad. Sci. USA 1984, 81, 6192–6196. [Google Scholar] [CrossRef]
- Mann, J.S.; Renwick, A.G.; Holgate, S.T. Release of adenosine and its metabolites from activated human leucocytes. Clin. Sci. (Lond.) 1986, 70, 461–468. [Google Scholar] [CrossRef]
- Madara, J.L.; Patapoff, T.W.; Gillece-Castro, B.; Colgan, S.P.; Parkos, C.A.; Delp, C.; Mrsny, R.J. 5′-adenosine monophosphate is the neutrophil-derived paracrine factor that elicits chloride secretion from T84 intestinal epithelial cell monolayers. J. Clin. Investig. 1993, 91, 2320–2325. [Google Scholar] [CrossRef]
- Resnick, M.B.; Colgan, S.P.; Patapoff, T.W.; Mrsny, R.J.; Awtrey, C.S.; Delp-Archer, C.; Weller, P.F.; Madara, J.L. Activated eosinophils evoke chloride secretion in model intestinal epithelia primarily via regulated release of 5′-AMP. J. Immunol. 1993, 151, 5716–5723. [Google Scholar]
- Newby, A.C. Adenosine and the concept of ‘retaliatory metabolites. Trends Biochem. Sci. 1984, 9, 42–44. [Google Scholar] [CrossRef]
- Cronstein, B.N.; Sitkovsky, M. Adenosine and adenosine receptors in the pathogenesis and treatment of rheumatic diseases. Nat. Rev. Rheumatol. 2017, 13, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, H. Extracellular purine metabolism. Drug Dev. Res. 1996, 39, 337–352. [Google Scholar] [CrossRef]
- Maliszewski, C.R.; Delespesse, G.J.; Schoenborn, M.A.; Armitage, R.J.; Fanslow, W.C.; Nakajima, T.; Baker, E.; Sutherland, G.R.; Poindexter, K.; Birks, C. The CD39 lymphoid cell activation antigen. Molecular cloning and structural characterization. J. Immunol. 1994, 153, 3574–3583. [Google Scholar] [PubMed]
- Kaczmarek, E.; Koziak, K.; Sevigny, J.; Siegel, J.B.; Anrather, J.; Beaudoin, A.R.; Bach, F.H.; Robson, S.C. Identification and characterization of CD39/vascular ATP diphosphohydrolase. J. Biol. Chem. 1996, 271, 33116–33122. [Google Scholar] [CrossRef]
- Zimmermann, H.; Braun, N.; Kegel, B.; Heine, P. New insights into molecular structure and function of ectonucleotidases in the nervous system. Neurochem. Int. 1998, 32, 421–425. [Google Scholar] [CrossRef]
- Kansas, G.S.; Wood, G.S.; Tedder, T.F. Expression, distribution, and biochemistry of human CD39. Role in activation-associated homotypic adhesion of lymphocytes. J. Immunol. 1991, 146, 2235–2244. [Google Scholar]
- Robson, S.C.; Ph, D.; Wu, Y.; Ph, D.; Sun, X.; Knosalla, C.; Dwyer, K.; Ph, D.; Enjyoji, K.; Ph, D. Ectonucleotidases of CD39 Family Modulate Vascular Inflammation and Thrombosis in Transplantation. Semin. Thromb. Hemost. 2005, 31, 217–233. [Google Scholar] [CrossRef]
- Bono, M.R.; Fernandez, D.; Flores-Santibanez, F.; Rosemblatt, M.; Sauma, D. CD73 and CD39 ectonucleotidases in T cell differentiation: Beyond immunosuppression. FEBS Lett. 2015, 589, 3454–3460. [Google Scholar] [CrossRef]
- Zimmermann, H. 5’-Nucleotidase: Molecular structure and functional aspects. Biochem. J. 1992, 285, 345–365. [Google Scholar] [CrossRef]
- Blackburn, M.R.; Kellems, R.E. Regulation and Function of Adenosine Deaminase in Mice. Prog. Nucleic Acid Res. Mol. Biol. 1996, 55, 195–226. [Google Scholar]
- Leiva, A.; Guzman-Gutierrez, E.; Contreras-Duarte, S.; Fuenzalida, B.; Cantin, C.; Carvajal, L.; Salsoso, R.; Gutierrez, J.; Pardo, F.; Sobrevia, L. Adenosine receptors: Modulators of lipid availability that are controlled by lipid levels. Mol. Aspects Med. 2017, 55, 26–44. [Google Scholar] [CrossRef] [PubMed]
- Van Calker, D.; Muller, M.; Hamprecht, B. Adenosine regulates via two different types of receptors, the accumulation of cyclic AMP in cultured brain cells. J. Neurochem. 1979, 33, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.K.Y.; Segala, E.; Robertson, N.; Deflorian, F.; Doré, A.S.; Errey, J.C.; Fiez-Vandal, C.; Marshall, F.H.; Cooke, R.M. Structures of Human A1and A2AAdenosine Receptors with Xanthines Reveal Determinants of Selectivity. Structure 2017, 25, 1275–1285.e4. [Google Scholar] [CrossRef] [PubMed]
- Piirainen, H.; Ashok, Y.; Nanekar, R.T.; Jaakola, V.P. Structural features of adenosine receptors: From crystal to function. Biochim. Biophys. Acta Biomembr. 2011, 1808, 1233–1244. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey Conn, P.; Christopoulos, A.; Lindsley, C.W. Allosteric modulators of GPCRs: A novel approach for the treatment of CNS disorders. Nat. Rev. Drug Discov. 2009, 8, 41–54. [Google Scholar] [CrossRef]
- Christopoulos, A. Allosteric binding sites on cell-surface receptors: Novel targets for drug discovery. Nat. Rev. Drug Discov. 2002, 1, 198–210. [Google Scholar] [CrossRef]
- Peleli, M.; Fredholm, B.B.; Sobrevia, L.; Carlström, M. Pharmacological targeting of adenosine receptor signaling. Mol. Aspects Med. 2017, 55, 4–8. [Google Scholar] [CrossRef]
- Fredholm, B.B.; IJzerman, A.P.; Jacobson, K.A.; Linden, J.; Müller, C.E. International union of basic and clinical pharmacology. LXXXI. Nomenclature and classification of adenosine receptors—An update. Pharmacol. Rev. 2011, 63, 1–34. [Google Scholar] [CrossRef]
- Stiles, G.L. Adenosine Receptors. J. Biol. Chem. 1992, 10, 6451–6454. [Google Scholar]
- Haskó, G.; Linden, J.; Cronstein, B.; Pacher, P. Adenosine receptors: Therapeutic aspects for inflammatory and immune diseases. Nat. Rev. Drug Discov. 2008, 7, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Burbiel, J.C.; Maaß, A.; Müller, C.E. Adenosine receptor agonists: From basic medicinal chemistry to clinical development. Expert Opin. Emerg. Drugs 2003, 8, 537–576. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B.; Arslan, G.; Halldner, L.; Kull, B.; Schulte, G.; Wasserman, W. Structure and function of adenosine receptors and their genes. Naunyn. Schmiedebergs. Arch. Pharmacol. 2000, 362, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B.; Chern, Y.; Franco, R.; Sitkovsky, M. Aspects of the general biology of adenosine A2A signaling. Prog. Neurobiol. 2007, 83, 263–276. [Google Scholar] [CrossRef]
- Varani, K.; Vincenzi, F.; Tosi, A.; Gessi, S.; Casetta, I.; Granieri, G.; Fazio, P.; Leung, E.; MacLennan, S.; Granieri, E.; et al. A2A adenosine receptor overexpression and functionality, as well as TNF-α levels, correlate with motor symptoms in Parkinson’s disease. FASEB J. 2010, 24, 587–598. [Google Scholar] [CrossRef]
- Ledent, C.; Vaugeoist, J.M.; Schiffmann, S.N.; Pedrazzini, T.; El Yacoubi, M.; Vanderhaeghen, J.J.; Costentin, J.; Heath, J.K.; Vassart, G.; Parmentier, M. Aggressiveness, hypoalgesia and high blood pressure in mice lacking the adenosine A2a receptor. Nature 1997, 388, 674–678. [Google Scholar] [CrossRef]
- Baraldi, P.G.; Tabrizi, M.A.; Gessi, S.; Borea, P.A. Adenosine receptor antagonists: Translating medicinal chemistry and pharmacology into clinical utility. Chem. Rev. 2008, 108, 238–263. [Google Scholar] [CrossRef]
- Gessi, S.; Merighi, S.; Fazzi, D.; Stefanelli, A.; Varani, K.; Borea, P.A. Adenosine receptor targeting in health and disease. Expert Opin. Investig. Drugs 2011, 20, 1591–1609. [Google Scholar] [CrossRef]
- Herve, D.; Levi-Strauss, M.; Marey-Semper, I.; Verney, C.; Tassin, J.P.; Glowinski, J.; Girault, J.A. G(olf) and Gs in rat basal ganglia: Possible involvement of G(olf) in the coupling of dopamine D1 receptor with adenylyl cyclase. J. Neurosci. 1993, 13, 2237–2248. [Google Scholar] [CrossRef]
- De Lera Ruiz, M.; Lim, Y.H.; Zheng, J. Adenosine A2A receptor as a drug discovery target. J. Med. Chem. 2014, 57, 3623–3650. [Google Scholar] [CrossRef]
- Cristalli, G.; Müller, C.E.; Volpini, R. Recent developments in Adenosine A2A receptor ligands. In Adenosine Receptors in Health and Disease; Wilson, C.N., Mustafa, S.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 193, pp. 59–98. [Google Scholar]
- Haskó, G.; Csóka, B.; Németh, Z.H.; Vizi, E.S.; Pacher, P. A2B adenosine receptors in immunity and inflammation. Trends Immunol. 2009, 30, 263–270. [Google Scholar] [CrossRef]
- Kalla, R.V.; Zablocki, J.; Tabrizi, M.A.; Baraldi, P.G. Recent developments in A2B adenosine receptor ligands. In Adenosine Receptors in Health and Disease; Wilson, C.N., Mustafa, S.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 193, pp. 99–122. [Google Scholar]
- Cohen, M.V.; Yang, X.; Downey, J.M. A 2b adenosine receptors can change their spots. Br. J. Pharmacol. 2010, 159, 1595–1597. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.Y.; Li, C.; Olah, M.E.; Johnson, R.A.; Stiles, G.L.; Civelli, O. Molecular cloning and characterization of an adenosine receptor: The A3 adenosine receptor. Proc. Natl. Acad. Sci. USA 1992, 89, 7432–7436. [Google Scholar] [CrossRef] [PubMed]
- Meyerhof, W.; Paust, H.J.; Schonrock, C.; Richter, D. Cloning of a cDNA encoding a novel putative G-protein-coupled receptor expressed in specific rat brain regions. DNA Cell Biol. 1991, 10, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Borea, P.A.; Varani, K.; Vincenzi, F.; Baraldi, P.G.; Tabrizi, M.A.; Merighi, S.; Gessi, S. The A3 adenosine receptor: History and perspectives. Pharmacol. Rev. 2015, 67, 74–102. [Google Scholar] [CrossRef]
- Jacobson, K.A.; Merighi, S.; Varani, K.; Borea, P.A.; Baraldi, S.; Aghazadeh Tabrizi, M.; Romagnoli, R.; Baraldi, P.G.; Ciancetta, A.; Tosh, D.K.; et al. A3 Adenosine Receptors as Modulators of Inflammation: From Medicinal Chemistry to Therapy. Med. Res. Rev. 2018, 38, 1031–1072. [Google Scholar] [CrossRef]
- Salvatore, C.A.; Jacobson, M.A.; Taylor, H.E.; Linden, J.; Johnson, R.G. Molecular cloning and characterization of the human A3 adenosine receptor. Proc. Natl. Acad. Sci. USA 1993, 90, 10365–10369. [Google Scholar] [CrossRef]
- Baraldi, P.G.; Preti, D.; Borea, P.A.; Varani, K. Medicinal Chemistry of A3 Adenosine Receptor Modulators: Pharmacological Activities and Therapeutic Implications. J. Med. Chem. 2012, 55, 5676–5703. [Google Scholar] [CrossRef]
- Jacobson, K.A.; Klutz, A.M.; Tosh, D.K.; Ivanov, A.A.; Preti, D.; Baraldi, P.G. Medicinal chemistry of the A 3 adenosine receptor: Agonists, antagonists, and receptor engineering. In Adenosine Receptors in Health and Disease; Wilson, C.N., Mustafa, S.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 193, pp. 123–159. [Google Scholar]
- Baraldi, P.G.; Cacciari, B.; Romagnoli, R.; Merighi, S.; Varani, K.; Borea, P.A.; Spalluto, G. A(3) adenosine receptor ligands: History and perspectives. Med. Res. Rev. 2000, 20, 103–128. [Google Scholar] [CrossRef]
- Cronstein, B.N.; Levin, R.I.; Philips, M.; Hirschhorn, R.; Abramson, S.B.; Weissmann, G. Neutrophil adherence to endothelium is enhanced via adenosine A1 receptors and inhibited via adenosine A2 receptors. J. Immunol. 1992, 148, 2201–2206. [Google Scholar]
- Zahler, S.; Becker, B.F. Adhesion of neutrophils to cultured human endothelial cells is enhanced by stimulation of adenosine A1-receptors. Drug Dev. Res. 1998, 45, 350–355. [Google Scholar] [CrossRef]
- Zahler, S.; Becker, B.F.; Raschke, P.; Gerlach, E. Stimulation of endothelial adenosine A1 receptors enhances adhesion of neutrophils in the intact guinea pig coronary system. Cardiovasc. Res. 1994, 28, 1366–1372. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Salmon, J.E.; Brogle, N.; Brownlie, C.; Edberg, J.C.; Kimberly, R.P.; Chen, B.X.; Erlanger, B.F. Human mononuclear phagocytes express adenosine A1 receptors. A novel mechanism for differential regulation of Fc gamma receptor function. J. Immunol. 1993, 151, 2775–2785. [Google Scholar] [PubMed]
- Clark, A.N.; Youkey, R.; Liu, X.; Jia, L.; Blatt, R.; Day, Y.-J.; Sullivan, G.W.; Linden, J.; Tucker, A.L. A1 adenosine receptor activation promotes angiogenesis and release of VEGF from monocytes. Circ. Res. 2007, 101, 1130–1138. [Google Scholar] [CrossRef]
- Kara, F.M.; Chitu, V.; Sloane, J.; Axelrod, M.; Fredholm, B.B.; Stanley, E.R.; Cronstein, B.N. Adenosine A1 receptors (A1Rs) play a critical role in osteoclast formation and function. FASEB J. 2010, 24, 2325–2333. [Google Scholar] [CrossRef]
- Takahashi, H.K.; Iwagaki, H.; Hamano, R.; Wake, H.; Kanke, T.; Liu, K.; Yoshino, T.; Tanaka, N.; Nishibori, M. Effects of adenosine on adhesion molecule expression and cytokine production in human PBMC depend on the receptor subtype activated. Br. J. Pharmacol. 2007, 150, 816–822. [Google Scholar] [CrossRef]
- Gallos, G.; Ruyle, T.D.; Emala, C.W.; Lee, H.T. A1 adenosine receptor knockout mice exhibit increased mortality, renal dysfunction, and hepatic injury in murine septic peritonitis. Am. J. Physiol. Ren. Physiol. 2005, 289, F369–F376. [Google Scholar] [CrossRef]
- Kim, J.; Kim, M.; Song, J.H.; Lee, H.T. Endogenous A1 adenosine receptors protect against hepatic ischemia reperfusion injury in mice. Liver Transplant. 2008, 14, 845–854. [Google Scholar] [CrossRef]
- Park, S.W.; Chen, S.W.C.; Kim, M.; Brown, K.M.; D’Agati, V.D.; Lee, H.T. Protection against acute kidney injury via A1 adenosine receptor-mediated Akt activation reduces liver injury after liver ischemia and reperfusion in mice. J. Pharmacol. Exp. Ther. 2010, 333, 736–747. [Google Scholar] [CrossRef]
- Hua, X.; Erikson, C.J.; Chason, K.D.; Rosebrock, C.N.; Deshpande, D.A.; Penn, R.B.; Tilley, S.L. Involvement of A1 adenosine receptors and neural pathways in adenosine-induced bronchoconstriction in mice. Am. J. Physiol. Cell. Mol. Physiol. 2007, 293, L25–L32. [Google Scholar] [CrossRef][Green Version]
- Wilson, C.N.; Nadeem, A.; Spina, D.; Brown, R.; Page, C.P.; Mustafa, S.J. Adenosine receptors and asthma. In Adenosine Receptors in Health and Disease; Wilson, C.N., Mustafa, S.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 193, pp. 329–362. [Google Scholar]
- Brown, R.A.; Spina, D.; Page, C.P. Adenosine receptors and asthma. Br. J. Pharmacol. 2008, 153, S446–S456. [Google Scholar] [CrossRef] [PubMed]
- Obiefuna, P.C.M.; Batra, V.K.; Nadeem, A.; Borron, P.; Wilson, C.N.; Jamal Mustafa, S. A novel A1 adenosine receptor antagonist, L-97-1 [3-[2-(4-aminophenyl)-ethyl]-8-benzyl-7-{2-ethyl-(2-hydroxy-ethyl)-amino]-ethyl} -1-propyl-3,7-dihydro-purine-2,6-dione], reduces allergic responses to house dust mite in an allergic rabbit model of asthma. J. Pharmacol. Exp. Ther. 2005, 315, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Ball, H.A.; Van Scott, M.K.; Robinson, C.B. Sense and antisense: Therapeutic potential of oligonucleotides and interference RNA in asthma and allergic disorders. Clin. Rev. Allergy Immunol. 2004, 27, 207–217. [Google Scholar] [CrossRef]
- Chunn, J.L.; Young, H.W.; Banerjee, S.K.; Colasurdo, G.N.; Blackburn, M.R. Adenosine-dependent airway inflammation and hyperresponsiveness in partially adenosine deaminase-deficient mice. J. Immunol. 2001, 167, 4676–4685. [Google Scholar] [CrossRef] [PubMed]
- Ngamsri, K.C.; Wagner, R.; Vollmer, I.; Stark, S.; Reutershan, J. Adenosine Receptor A1 Regulates Polymorphonuclear Cell Trafficking and Microvascular Permeability in Lipopolysaccharide-Induced Lung Injury. J. Immunol. 2010, 185, 4374–4384. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, L.G.; Sharma, A.K.; Lapar, D.J.; Kron, I.L.; Laubach, V.E. Adenosine A1 receptor activation attenuates lung ischemia-reperfusion injury. J. Thorac. Cardiovasc. Surg. 2013, 145, 1654–1659. [Google Scholar] [CrossRef] [PubMed]
- Aeffner, F.; Woods, P.S.; Davis, I.C. Activation of A1-Adenosine Receptors Promotes Leukocyte Recruitment to the Lung and Attenuates Acute Lung Injury in Mice Infected with Influenza A/WSN/33 (H1N1) Virus. J. Virol. 2014, 88, 10214–10227. [Google Scholar] [CrossRef]
- Romano, F.D.; Dobson, J.G. Adenosine modulates β-adrenergic signal transduction in guinea-pig heart ventricular membranes. J. Mol. Cell. Cardiol. 1990, 22, 1359–1370. [Google Scholar] [CrossRef]
- Fenton, R.A.; Dobson, J.G., Jr. Adenosine A1 and A2A receptor effects on G-protein cycling in β-adrenergic stimulated ventricular membranes. J. Cell. Physiol. 2007, 213, 785–792. [Google Scholar] [CrossRef]
- Fenton, R.A.; Shea, L.G.; Doddi, C.; Dobson, J.G., Jr. Myocardial adenosine A(1)-receptor-mediated adenoprotection involves phospholipase C, PKC-epsilon, and p38 MAPK, but not HSP27. Am. J. Physiol. Heart Circ. Physiol. 2010, 298, H1671–H1678. [Google Scholar] [CrossRef]
- Lorbar, M.; Chung, E.S.; Nabi, A.; Skalova, K.; Fenton, R.A.; Dobson, J.G.; Meyer, T.E. Receptors subtypes involved in adenosine-mediated modulation of norepinephrine release from cardiac nerve terminals. Can. J. Physiol. Pharmacol. 2004, 82, 1026–1031. [Google Scholar] [CrossRef]
- Burgdorf, C. Adenosine inhibits norepinephrine release in the postischemic rat heart: The mechanism of neuronal stunning. Cardiovasc. Res. 2001, 49, 713–720. [Google Scholar] [CrossRef]
- Tawfik, H.E.; Schnermann, J.; Oldenburg, P.J.; Mustafa, S.J. Role of A1 adenosine receptors in regulation of vascular tone. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H1411–H1416. [Google Scholar] [CrossRef] [PubMed]
- Nayeem, M.A.; Zeldin, D.C.; Boegehold, M.A.; Morisseau, C.; Marowsky, A.; Ponnoth, D.S.; Roush, K.P.; Falck, J.R. Modulation by salt intake of the vascular response mediated through adenosine A(2A) receptor: Role of CYP epoxygenase and soluble epoxide hydrolase. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R325–R333. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Halenda, S.P.; Sturek, M.; Wilden, P.A. Cell-signaling evidence for adenosine stimulation of coronary smooth muscle proliferation via the A1 adenosine receptor. Circ. Res. 2005, 97, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Maeda, T.; Koos, B.J. Adenosine A1 and A2a receptors modulate insulinemia, glycemia, and lactatemia in fetal sheep. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R693–R701. [Google Scholar] [CrossRef] [PubMed]
- Johansson, S.M.; Yang, J.-N.; Lindgren, E.; Fredholm, B.B. Eliminating the antilipolytic adenosine A1 receptor does not lead to compensatory changes in the antilipolytic actions of PGE2 and nicotinic acid. Acta Physiol. 2007, 190, 87–96. [Google Scholar] [CrossRef]
- Urmaliya, V.B.; Church, J.E.; Coupar, I.M.; Rose’Meyer, R.B.; Pouton, C.W.; White, P.J. Cardioprotection induced by adenosine A1 receptor agonists in a cardiac cell ischemia model involves cooperative activation of adenosine A2A and A2B receptors by endogenous adenosine. J. Cardiovasc. Pharmacol. 2009, 53, 424–433. [Google Scholar] [CrossRef]
- Urmaliya, V.B.; Pouton, C.W.; Ledent, C.; Short, J.L.; White, P.J. Cooperative cardioprotection through adenosine A1 and A 2A receptor agonism in ischemia-reperfused isolated mouse heart. J. Cardiovasc. Pharmacol. 2010, 56, 379–388. [Google Scholar] [CrossRef]
- Headrick, J.P.; Peart, J.N.; Reichelt, M.E.; Haseler, L.J. Adenosine and its receptors in the heart: Regulation, retaliation and adaptation. Biochim. Biophys. Acta Biomembr. 2011, 1808, 1413–1428. [Google Scholar] [CrossRef]
- Liao, Y.; Takashima, S.; Asano, Y.; Asakura, M.; Ogai, A.; Shintani, Y.; Minamino, T.; Asanuma, H.; Sanada, S.; Kim, J.; et al. Activation of Adenosine A 1 Receptor Attenuates Cardiac Hypertrophy and Prevents Heart Failure in Murine Left Ventricular Pressure-Overload Model. Circ. Res. 2003, 93, 759–766. [Google Scholar] [CrossRef]
- Greenberg, B.; Thomas, I.; Banish, D.; Goldman, S.; Havranek, E.; Massie, B.M.; Zhu, Y.; Ticho, B.; Abraham, W.T. Effects of Multiple Oral Doses of an A1 Adenosine Antagonist, BG9928, in Patients With Heart Failure. Results of a Placebo-Controlled, Dose-Escalation Study. J. Am. Coll. Cardiol. 2007, 50, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Voors, A.A.; Dittrich, H.C.; Massie, B.M.; Delucca, P.; Mansoor, G.A.; Metra, M.; Cotter, G.; Weatherley, B.D.; Ponikowski, P.; Teerlink, J.R.; et al. Effects of the adenosine A1 receptor antagonist rolofylline on renal function. J. Am. Coll. Cardiol. 2011, 57, 1899–1907. [Google Scholar] [CrossRef] [PubMed]
- Fozard, J.R.; Ellis, K.M.; Villela Dantas, M.F.; Tigani, B.; Mazzoni, L. Effects of CGS 21680, a selective adenosine A2A receptor agonist, on allergic airways inflammation in the rat. Eur. J. Pharmacol. 2002, 438, 183–188. [Google Scholar] [CrossRef]
- Trevethick, M.A.; Mantell, S.J.; Stuart, E.F.; Barnard, A.; Wright, K.N.; Yeadon, M. Treating lung inflammation with agonists of the adenosine A 2A receptor: Promises, problems and potential solutions. Br. J. Pharmacol. 2008, 155, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Luijk, B.; Van Den Berge, M.; Kerstjens, H.A.M.; Postma, D.S.; Cass, L.; Sabin, A.; Lammers, J.W.J. Effect of an inhaled adenosine A2A agonist on the allergen-induced late asthmatic response. Allergy 2008, 63, 75–80. [Google Scholar]
- Golzar, Y.; Doukky, R. Regadenoson use in patients with chronic obstructive pulmonary disease: The state of current knowledge. Int. J. COPD 2014, 9, 129–137. [Google Scholar]
- Haskó, G.; Xu, D.Z.; Lu, Q.; Németh, Z.H.; Jabush, J.; Berezina, T.L.; Zaets, S.B.; Csóka, B.; Deitch, E.A. Adenosine A2A receptor activation reduces lung injury in trauma/hemorrhagic shock. Crit. Care Med. 2006, 34, 1119–1125. [Google Scholar] [CrossRef]
- Konrad, F.M.; Neudeck, G.; Vollmer, I.; Ngamsri, K.C.; Thiel, M.; Reutershan, J. Protective effects of pentoxifylline in pulmonary inflammation are adenosine receptor A2A dependent. FASEB J. 2013, 27, 3524–3535. [Google Scholar] [CrossRef]
- Greer, S.; Page, C.W.; Joshi, T.; Yan, D.; Newton, R.; Giembycz, M.A. Concurrent agonism of adenosine a2b and glucocorticoid receptors in human airway epithelial cells cooperatively induces genes with anti-inflammatory potential: A novel approach to treat chronic obstructive pulmonary disease. J. Pharmacol. Exp. Ther. 2013, 346, 473–485. [Google Scholar] [CrossRef]
- Mustafa, S.J.; Nadeem, A.; Fan, M.; Zhong, H.; Belardinelli, L.; Zeng, D. Effect of a specific and selective A2B adenosine receptor antagonist on adenosine agonist AMP and allergen-induced airway responsiveness and cellular influx in a mouse model of asthma. J. Pharmacol. Exp. Ther. 2007, 320, 1246–1251. [Google Scholar] [CrossRef] [PubMed]
- Eckle, T.; Grenz, A.; Laucher, S.; Eltzschig, H.K. A2B adenosine receptor signaling attenuates acute lung injury by enhancing alveolar fluid clearance in mice. J. Clin. Investig. 2008, 118, 3301–3315. [Google Scholar] [CrossRef] [PubMed]
- Karmouty-Quintana, H.; Zhong, H.; Acero, L.; Weng, T.; Melicoff, E.; West, J.D.; Hemnes, A.; Grenz, A.; Eltzschig, H.K.; Blackwell, T.S.; et al. The A2B adenosine receptor modulates pulmonary hypertension associated with interstitial lung disease. FASEB J. 2012, 26, 2546–2557. [Google Scholar] [CrossRef]
- Chan, E.S.L.; Cronstein, B.N. Adenosine in fibrosis. Mod. Rheumatol. 2010, 20, 114–122. [Google Scholar] [CrossRef]
- Rivo, J.; Zeira, E.; Galun, E.; Matot, I. Activation of A 3 adenosine receptors attenuates lung injury after in vivo reperfusion. Anesthesiology 2004, 101, 1153–1159. [Google Scholar] [CrossRef] [PubMed]
- Tendera, M.; Gaszewska-Zurek, E.; Parma, Z.; Ponikowski, P.; Jankowska, E.; Kawecka-Jaszcz, K.; Czarnecka, D.; Krzemińska-Pakuła, M.; Bednarkiewicz, Z.; Sosnowski, M.; et al. The new oral adenosine A1 receptor agonist capadenoson in male patients with stable angina. Clin. Res. Cardiol. 2012, 101, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Baltos, J.A.; Vecchio, E.A.; Harris, M.A.; Qin, C.X.; Ritchie, R.H.; Christopoulos, A.; White, P.J.; May, L.T. Capadenoson, a clinically trialed partial adenosine A 1 receptor agonist, can stimulate adenosine A 2B receptor biased agonism. Biochem. Pharmacol. 2017, 135, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, K.A.; Tosh, D.K.; Jain, S.; Gao, Z.G. Historical and current adenosine receptor agonists in preclinical and clinical development. Front. Cell. Neurosci. 2019, 13, 124. [Google Scholar] [CrossRef]
- Voors, A.A.; Düngen, H.D.; Senni, M.; Nodari, S.; Agostoni, P.; Ponikowski, P.; Bax, J.J.; Butler, J.; Kim, R.J.; Dorhout, B.; et al. Safety and Tolerability of Neladenoson Bialanate, a Novel Oral Partial Adenosine A1 Receptor Agonist, in Patients With Chronic Heart Failure. J. Clin. Pharmacol. 2017, 57, 440–451. [Google Scholar] [CrossRef]
- Headrick, J.P.; Lasley, R.D. Adenosine receptors and reperfusion injury of the heart. In Adenosine Receptors in Health and Disease; Wilson, C.N., Mustafa, S.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 193, pp. 189–214. [Google Scholar]
- McIntosh, V.J.; Lasley, R.D. Adenosine receptor-mediated cardioprotection: Are all 4 subtypes required or redundant? J. Cardiovasc. Pharmacol. Ther. 2012, 17, 21–33. [Google Scholar] [CrossRef]
- Kirchhof, P.; Fabritz, L.; Fortmüller, L.; Matherne, G.P.; Lankford, A.; Baba, H.A.; Schmitz, W.; Breithardt, G.; Neumann, J.; Boknik, P. Altered sinus nodal and atrioventricular nodal function in freely moving mice overexpressing the A1 adenosine receptor. Am. J. Physiol. Heart Circ. Physiol. 2003, 285, H145–H153. [Google Scholar] [CrossRef] [PubMed]
- Ellenbogen, K.A.; O’Neill, G.; Prystowsky, E.N.; Camm, J.A.; Meng, L.; Lieu, H.D.; Jerling, M.; Shreeniwas, R.; Belardinelli, L.; Wolff, A.A.; et al. Trial to evaluate the management of paroxysmal supraventricular tachycardia during an electrophysiology study with tecadenoson. Circulation 2005, 111, 3202–3208. [Google Scholar] [CrossRef] [PubMed]
- Mason, P.K.; DiMarco, J.P. New pharmacological agents for arrhythmias. Circ. Arrhythmia Electrophysiol. 2009, 2, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Szentmiklosi, A.; Galajda, Z.; Cseppento, A.; Gesztelyi, R.; Susan, Z.; Hegyi, B.; Nanasi, P. The Janus Face of Adenosine: Antiarrhythmic and Proarrhythmic Actions. Curr. Pharm. Des. 2015, 21, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.G.; Saggau, P. Presynaptic inhibition of elicited neurotransmitter release. Trends Neurosci. 1997, 20, 204–212. [Google Scholar] [CrossRef]
- Stone, T.W.; Ceruti, S.; Abbracchio, M.P. Adenosine receptors and neurological disease: Neuroprotection and neurodegeneration. In Adenosine Receptors in Health and Disease; Wilson, C.N., Mustafa, S.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 193, pp. 535–587. [Google Scholar]
- Dunwiddie, T.V.; Masino, S.A. The Role and Regulation of Adenosine in the Central Nervous System. Annu. Rev. Neurosci. 2001, 24, 31–55. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.A.; Sebastião, A.M.; De Mendonça, A. Adenosine receptors in the nervous system: Pathophysiological implications. Prog. Neurobiol. 2002, 68, 377–392. [Google Scholar] [CrossRef]
- Stockwell, J.; Jakova, E.; Cayabyab, F.S. Adenosine A1 and A2A receptors in the brain: Current research and their role in neurodegeneration. Molecules 2017, 22, 676. [Google Scholar] [CrossRef]
- Borea, P.A.; Gessi, S.; Merighi, S.; Varani, K. Adenosine as a Multi-Signalling Guardian Angel in Human Diseases: When, Where and How Does it Exert its Protective Effects? Trends Pharmacol. Sci. 2016, 37, 419–434. [Google Scholar] [CrossRef]
- Dickenson, A.H.; Suzuki, R.; Reeve, A.J. Adenosine as a potential analgesic target in inflammatory and neuropathic pains. CNS Drugs 2000, 13, 77–85. [Google Scholar] [CrossRef]
- Sneyd, R.J.; Langton, J.A.; Allan, L.G.; Peacock, J.E.; Rowbotham, D.J. Multicentre evaluation of the adenosine agonist GR79236X in patients with dental pain after third molar extraction. Br. J. Anaesth. 2007, 98, 672–676. [Google Scholar] [CrossRef]
- Luongo, L.; Petrelli, R.; Gatta, L.; Giordano, C.; Guida, F.; Vita, P.; Franchetti, P.; Grifantini, M.; De Novellis, V.; Cappellacci, L.; et al. 5′-Chloro-5′-deoxy-(±)-ENBA, a potent and selective adenosine A1 receptor agonist, alleviates neuropathic pain in mice through functional glial and microglial changes without affecting motor or cardiovascular functions. Molecules 2012, 17, 13712–13726. [Google Scholar] [CrossRef] [PubMed]
- Tosh, D.K.; Rao, H.; Bitant, A.; Salmaso, V.; Mannes, P.; Lieberman, D.I.; Vaughan, K.L.; Mattison, J.A.; Rothwell, A.C.; Auchampach, J.A.; et al. Design and in Vivo Characterization of A 1 Adenosine Receptor Agonists in the Native Ribose and Conformationally Constrained (N)-Methanocarba Series. J. Med. Chem. 2019, 62, 1502–1522. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.G.; Jacobson, K.A. Emerging adenosine receptor agonists an update. Expert Opin. Emerg. Drugs 2011, 16, 597–602. [Google Scholar] [CrossRef]
- Fedele, D.E.; Li, T.; Lan, J.Q.; Fredholm, B.B.; Boison, D. Adenosine A1 receptors are crucial in keeping an epileptic focus localized. Exp. Neurol. 2006, 200, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Benarroch, E.E. Adenosine and its receptors: Multiple modulatory functions and potential therapeutic targets for neurologic disease. Neurology 2008, 70, 231–236. [Google Scholar] [CrossRef]
- Hargus, N.J.; Jennings, C.; Perez-Reyes, E.; Bertram, E.H.; Patel, M.K. Enhanced actions of adenosine in medial entorhinal cortex layer II stellate neurons in temporal lobe epilepsy are mediated via A 1-receptor activation. Epilepsia 2012, 53, 168–176. [Google Scholar] [CrossRef]
- Masino, S.A.; Kawamura, M.; Ruskin, D.N. Adenosine receptors and epilepsy. Current evidence and future potential. Int. Rev. Neurobiol. 2014, 119, 233–255. [Google Scholar]
- Kovács, Z.; D’Agostino, D.P.; Dobolyi, A.; Ari, C. Adenosine A1 Receptor Antagonism Abolished the Anti-seizure Effects of Exogenous Ketone Supplementation in Wistar Albino Glaxo Rijswijk Rats. Front. Mol. Neurosci. 2017, 10, 235. [Google Scholar] [CrossRef]
- Johansson, B.; Halldner, L.; Dunwiddie, T.V.; Masino, S.A.; Poelchen, W.; Giménez-Llort, L.; Escorihuela, R.M.; Fernández-Teruel, A.; Wiesenfeld-Hallin, Z.; Xu, X.J.; et al. Hyperalgesia, anxiety, and decreased hypoxic neuroprotection in mice lacking the adenosine A1 receptor. Proc. Natl. Acad. Sci. USA 2001, 98, 9407–9412. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; Fernández-Teruel, A.; Escorihuela, R.M.; Fredholm, B.B.; Tobeña, A.; Pekny, M.; Johansson, B. Mice lacking the adenosine A1 receptor are anxious and aggressive, but are normal learners with reduced muscle strength and survival rate. Eur. J. Neurosci. 2002, 16, 547–550. [Google Scholar] [CrossRef] [PubMed]
- Prediger, R.D.S.; Da Silva, G.E.; Batista, L.C.; Bittencourt, A.L.; Takahashi, R.N. Activation of adenosine A1 receptors reduces anxiety-like behavior during acute ethanol withdrawal (hangover) in mice. Neuropsychopharmacology 2006, 31, 2210–2220. [Google Scholar] [CrossRef] [PubMed]
- Vincenzi, F.; Borea, P.A.; Varani, K. Anxiolytic properties of A1 adenosine receptor PAMs. Oncotarget 2017, 8, 7216–7217. [Google Scholar] [CrossRef] [PubMed]
- Serchov, T.; Clement, H.W.; Schwarz, M.K.; Iasevoli, F.; Tosh, D.K.; Idzko, M.; Jacobson, K.A.; de Bartolomeis, A.; Normann, C.; Biber, K.; et al. Increased Signaling via Adenosine A1Receptors, Sleep Deprivation, Imipramine, and Ketamine Inhibit Depressive-like Behavior via Induction of Homer1a. Neuron 2015, 87, 549–562. [Google Scholar] [CrossRef]
- Sigworth, L.A.; Rea, M.A. Adenosine A1 receptors regulate the response of the mouse circadian clock to light. Brain Res. 2003, 960, 246–251. [Google Scholar] [CrossRef]
- Elmenhorst, D.; Meyer, P.T.; Winz, O.H.; Matusch, A.; Ermert, J.; Coenen, H.H.; Basheer, R.; Haas, H.L.; Zilles, K.; Bauer, A. Sleep deprivation increases A1 adenosine receptor binding in the human brain: A positron emission tomography study. J. Neurosci. 2007, 27, 2410–2415. [Google Scholar] [CrossRef]
- Basheer, R.; Strecker, R.E.; Thakkar, M.M.; McCarley, R.W. Adenosine and sleep-wake regulation. Prog. Neurobiol. 2004, 73, 379–396. [Google Scholar] [CrossRef]
- Loram, L.C.; Harrison, J.A.; Sloane, E.M.; Hutchinson, M.R.; Sholar, P.; Taylor, F.R.; Berkelhammer, D.; Coats, B.D.; Poole, S.; Milligan, E.D.; et al. Enduring reversal of neuropathic pain by a single intrathecal injection of adenosine 2A receptor agonists: A novel therapy for neuropathic pain. J. Neurosci. 2009, 29, 14015–14025. [Google Scholar] [CrossRef]
- De Mendonça, A.; Ribeiro, J.A. Adenosine and synaptic plasticity. Drug Dev. Res. 2001, 52, 283–290. [Google Scholar] [CrossRef]
- De Mendonça, A.; Ribeiro, J.A. Endogenous adenosine modulates long-term potentiation in the hippocampus. Neuroscience 1994, 62, 385–390. [Google Scholar] [CrossRef]
- De Mendonça, A.; Almeida, T.; Bashir, Z.I.; Ribeiro, J.A. Endogenous adenosine attenuates long-term depression and depotentiation in the CA1 region of the rat hippocampus. Neuropharmacology 1997, 36, 161–167. [Google Scholar] [CrossRef]
- Mihara, T.; Mihara, K.; Yarimizu, J.; Mitani, Y.; Matsuda, R.; Yamamoto, H.; Aoki, S.; Akahane, A.; Iwashita, A.; Matsuoka, N. Pharmacological characterization of a novel, potent adenosine A1 and A2A receptor dual antagonist, 5-[5-amino-3-(4-fluorophenyl) pyrazin-2-yl]-1-isopropylpyridine-2(1H)-one (ASP5854), in models of Parkinson’s disease and cognition. J. Pharmacol. Exp. Ther. 2007, 323, 708–719. [Google Scholar] [CrossRef] [PubMed]
- Lopes, L.V.; Sebastiao, A.M.; Ribeiro, J.A. Adenosine and Related Drugs in Brain Diseases: Present and Future in Clinical Trials. Curr. Top. Med. Chem. 2011, 11, 1087–1101. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Ferré, S.; Genedani, S.; Franco, R.; Agnati, L.F. Adenosine receptor-dopamine receptor interactions in the basal ganglia and their relevance for brain function. Physiol. Behav. 2007, 92, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Blum, D.; Hourez, R.; Galas, M.C.; Popoli, P.; Schiffmann, S.N. Adenosine receptors and Huntington’s disease: Implications for pathogenesis and therapeutics. Lancet Neurol. 2003, 2, 366–374. [Google Scholar] [CrossRef]
- Sebastião, A.M.; Ribeiro, J.A. Adenosine receptors and the central nervous system. In Adenosine Receptors in Health and Disease; Wilson, C.N., Mustafa, S.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 193, pp. 471–534. [Google Scholar]
- Peleli, M.; Carlstrom, M. Adenosine signaling in diabetes mellitus and associated cardiovascular and renal complications. Mol. Aspects Med. 2017, 55, 62–74. [Google Scholar] [CrossRef]
- Kiesman, W.F.; Elzein, E.; Zablocki, J. A1 adenosine receptor antagonists, agonists, and allosteric enhancers. In Adenosine Receptors in Health and Disease; Wilson, C.N., Mustafa, S.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 193, pp. 25–58. [Google Scholar]
- Elzein, E.; Zablocki, J. A1 adenosine receptor agonists and their potential therapeutic applications. Expert Opin. Investig. Drugs 2008, 17, 1901–1910. [Google Scholar] [CrossRef]
- Staehr, P.M.; Dhalla, A.K.; Zack, J.; Wang, X.; Ho, Y.L.; Bingham, J.; Belardinelli, L. Reduction of free fatty acids, safety, and pharmacokinetics of oral GS-9667, an a1 adenosine receptor partial agonist. J. Clin. Pharmacol. 2013, 53, 385–392. [Google Scholar] [CrossRef]
- Dhalla, A.K.; Chisholm, J.W.; Reaven, G.M.; Belardinelli, L. A1 adenosine receptor: Role in diabetes and obesity. In Adenosine Receptors in Health and Disease; Wilson, C.N., Mustafa, S.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 193, pp. 271–295. [Google Scholar]
- Hasko, G.; Pacher, P. A2A receptors in inflammation and injury: Lessons learned from transgenic animals. J. Leukoc. Biol. 2008, 83, 447–455. [Google Scholar] [CrossRef]
- Link, A.A.; Kino, T.; Worth, J.A.; McGuire, J.L.; Crane, M.L.; Chrousos, G.P.; Wilder, R.L.; Elenkov, I.J. Ligand-activation of the adenosine A2a receptors inhibits IL-12 production by human monocytes. J. Immunol. 2000, 164, 436–442. [Google Scholar] [CrossRef]
- Bouma, M.G.; Stad, R.K.; van den Wildenberg, F.A.; Buurman, W.A. Differential regulatory effects of adenosine on cytokine release by activated human monocytes. J. Immunol. 1994, 153, 4159–4168. [Google Scholar]
- Németh, Z.H.; Csóka, B.; Wilmanski, J.; Xu, D.; Lu, Q.; Ledent, C.; Deitch, E.A.; Pacher, P.; Spolarics, Z.; Haskó, G. Adenosine A 2A Receptor Inactivation Increases Survival in Polymicrobial Sepsis. J. Immunol. 2006, 176, 5616–5626. [Google Scholar] [CrossRef]
- Antonioli, L.; Fornai, M.; Blandizzi, C.; Pacher, P.; Haskó, G. Adenosine signaling and the immune system: When a lot could be too much. Immunol. Lett. 2019, 205, 9–15. [Google Scholar] [CrossRef]
- Flögel, U.; Burghoff, S.; Van Lent, P.L.E.M.; Temme, S.; Galbarz, L.; Ding, Z.; El-Tayeb, A.; Huels, S.; Bönner, F.; Borg, N.; et al. Selective activation of adenosine A2A receptors on immune cells by a CD73-dependent prodrug suppresses joint inflammation in experimental rheumatoid arthritis. Sci. Transl. Med. 2012, 4, 146ra108. [Google Scholar] [CrossRef] [PubMed]
- Squadrito, F.; Bitto, A.; Irrera, N.; Pizzino, G.; Pallio, G.; Minutoli, L.; Altavilla, D. Pharmacological Activity and Clinical Use of PDRN. Front. Pharmacol. 2017, 8, 224. [Google Scholar] [CrossRef] [PubMed]
- Field, J.J.; Majerus, E.; Gordeuk, V.R.; Gowhari, M.; Hoppe, C.; Heeney, M.M.; Achebe, M.; George, A.; Chu, H.; Sheehan, B.; et al. Randomized phase 2 trial of regadenoson for treatment of acute vaso-occlusive crises in sickle cell disease. Blood Adv. 2017, 1, 1645–1649. [Google Scholar] [CrossRef] [PubMed]
- Antonioli, L.; Csoka, B.; Fornai, M.; Colucci, R.; Kokai, E.; Blandizzi, C.; Hasko, G. Adenosine and inflammation: what’s new on the horizon? Drug Discov. Today 2014, 19, 1051–1068. [Google Scholar] [CrossRef] [PubMed]
- Peart, J.N.; Headrick, J.P. Adenosinergic cardioprotection: Multiple receptors, multiple pathways. Pharmacol. Ther. 2007, 114, 208–221. [Google Scholar] [CrossRef]
- Dobson, J.G.; Shea, L.G.; Fenton, R.A. Adenosine A2A and β-adrenergic calcium transient and contractile responses in rat ventricular myocytes. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H2364–H2372. [Google Scholar] [CrossRef][Green Version]
- Norton, E.D.; Jackson, E.K.; Turner, M.B.; Virmani, R.; Forman, M.B. The effects of intravenous infusions of selective adenosine A1-receptor and A2-receptor agonists on myocardial reperfusion injury. Am. Heart J. 1992, 123, 332–338. [Google Scholar] [CrossRef]
- Glover, D.K.; Ruiz, M.; Takehana, K.; Petruzella, F.D.; Rieger, J.M.; Macdonald, T.L.; Watson, D.D.; Linden, J.; Beller, G.A. Cardioprotection by adenosine A2A agonists in a canine model of myocardial stunning produced by multiple episodes of transient ischemia. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H3164–H3171. [Google Scholar] [CrossRef] [PubMed]
- Boknik, P.; Drzewiecki, K.; Eskandar, J.; Gergs, U.; Hofmann, B.; Treede, H.; Grote-Wessels, S.; Fabritz, L.; Kirchhof, P.; Fortmüller, L.; et al. Evidence for Arrhythmogenic Effects of A2A-Adenosine Receptors. Front. Pharmacol. 2019, 10, 1051. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekera, P.C.; McIntosh, V.J.; Cao, F.X.; Lasley, R.D. Differential effects of adenosine A2a and A2b receptors on cardiac contractility. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H2082–H2089. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, J.S.; Gabriel-Costa, D.; Sudo, R.T.; Wang, H.; Groban, L.; Ferraz, E.B.; Nascimento, J.H.M.; Fraga, C.A.M.; Barreiro, E.J.; Zapata-Sudo, G. Adenosine A2A receptor agonist prevents cardiac remodeling and dysfunction in spontaneously hypertensive male rats after myocardial infarction. Drug Des. Dev. Ther. 2017, 11, 553–562. [Google Scholar] [CrossRef]
- Mustafa, S.J.; Morrison, R.R.; Teng, B.; Pelleg, A. Adenosine receptors and the heart: Role in regulation of coronary blood flow and cardiac electrophysiology. In Adenosine Receptors in Health and Disease; Wilson, C., Mustafa, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 193, pp. 161–188. [Google Scholar]
- Fuentes, E.; Fuentes, M.; Caballero, J.; Palomo, I.; Hinz, S.; El-Tayeb, A.; Müller, C.E. Adenosine A2A receptor agonists with potent antiplatelet activity. Platelets 2018, 29, 292–300. [Google Scholar] [CrossRef]
- Voloshyna, I.; Littlefield, M.J.; Kaplan, L.; Rieger, J.M.; Figler, R.; Reiss, A.B. Adenosine A2A Receptor Agonists Regulate Cholesterol Homeostasis in Mouse Bone Marrow Derived Macrophages (BMDM). FASEB J. 2013, 27. [Google Scholar] [CrossRef]
- Reiss, A.B.; Grossfeld, D.; Kasselman, L.J.; Renna, H.A.; Vernice, N.A.; Drewes, W.; Konig, J.; Carsons, S.E.; DeLeon, J. Adenosine and the Cardiovascular System. Am. J. Cardiovasc. Drugs 2019, 19, 449–464. [Google Scholar] [CrossRef]
- Koupenova, M.; Johnston-Cox, H.; Vezeridis, A.; Gavras, H.; Yang, D.; Zannis, V.; Ravid, K. A2b adenosine receptor regulates hyperlipidemia and atherosclerosis. Circulation 2012, 125, 354–363. [Google Scholar] [CrossRef]
- Tian, Y.; Piras, B.A.; Kron, I.L.; French, B.A.; Yang, Z. Adenosine 2B Receptor Activation Reduces Myocardial Reperfusion Injury by Promoting Anti-Inflammatory Macrophages Differentiation via PI3K/Akt Pathway. Oxid. Med. Cell. Longev. 2015, 2015, 585297. [Google Scholar] [CrossRef]
- Toldo, S.; Zhong, H.; Mezzaroma, E.; Van Tassell, B.W.; Kannan, H.; Zeng, D.; Belardinelli, L.; Voelkel, N.F.; Abbate, A. GS-6201, a selective blocker of the A2B adenosine receptor, attenuates cardiac remodeling after acute myocardial infarction in the mouse. J. Pharmacol. Exp. Ther. 2012, 343, 587–595. [Google Scholar] [CrossRef]
- Németh, Z.H.; Bleich, D.; Csóka, B.; Pacher, P.; Mabley, J.G.; Himer, L.; Vizi, E.S.; Deitch, E.A.; Szabó, C.; Cronstein, B.N.; et al. Adenosine receptor activation ameliorates type 1 diabetes. FASEB J. 2007, 21, 2379–2388. [Google Scholar] [CrossRef] [PubMed]
- Figler, R.A.; Wang, G.; Srinivasan, S.; Jung, D.Y.; Zhang, Z.; Pankow, J.S.; Ravid, K.; Fredholm, B.; Hedrick, C.C.; Rich, S.S.; et al. Links between Insulin resistance, adenosine A2B receptors, and inflammatory markers in mice and humans. Diabetes 2011, 60, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Valladares, D.; Quezada, C.; Montecinos, P.; Concha, I.I.; Yañez, A.J.; Sobrevia, L.; Martín, R.S. Adenosine A2B receptor mediates an increase on VEGF-A production in rat kidney glomeruli. Biochem. Biophys. Res. Commun. 2008, 366, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Dorotea, D.; Cho, A.; Lee, G.; Kwon, G.; Lee, J.; Sahu, P.K.; Jeong, L.S.; Cha, D.R.; Ha, H. Orally active, species-independent novel A3 adenosine receptor antagonist protects against kidney injury in db/db mice. Exp. Mol. Med. 2018, 50, 38. [Google Scholar] [CrossRef]
- Park, J.G.; Jeong, S.J.; Yu, J.; Kim, G.; Jeong, L.S.; Oh, G.T. LJ-1888, a selective antagonist for the A3 adenosine receptor, ameliorates the development of atherosclerosis and hypercholesterolemia in apolipoprotein E knock-out mice. BMB Rep. 2018, 51, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Auchampach, J.A.; Ge, Z.-D.; Wan, T.C.; Moore, J.; Gross, G.J. A3 adenosine receptor agonist IB-MECA reduces myocardial ischemia-reperfusion injury in dogs. Am. J. Physiol. Heart Circ. Physiol. 2003, 285, H607–H613. [Google Scholar] [CrossRef]
- Ge, Z.D.; Peart, J.N.; Kreckler, L.M.; Wan, T.C.; Jacobson, M.A.; Gross, G.J.; Auchampach, J.A. Cl-IB-MECA [2-chloro-N6-(3-iodobenzyl)adenosine-5′-N-methylcarboxamide] reduces ischemia/reperfusion injury in mice by activating the A3 adenosine receptor. J. Pharmacol. Exp. Ther. 2006, 319, 1200–1210. [Google Scholar] [CrossRef]
- Wan, T.C.; Ge, Z.D.; Tampo, A.; Mio, Y.; Bienengraeber, M.W.; Tracey, W.R.; Gross, G.J.; Kwok, W.M.; Auchampach, J.A. The A3 adenosine receptor agonist CP-532,903 [N 6-(2,5-dichlorobenzyl)-3′ -aminoadenosine-5′-N- methylcarboxamide] protects against myocardial ischemia/reperfusion injury via the sarcolemmal ATP-sensitive potassium channel. J. Pharmacol. Exp. Ther. 2008, 324, 234–243. [Google Scholar] [CrossRef]
- Mohamed, R.A.; Agha, A.M.; Abdel-Rahman, A.A.; Nassar, N.N. Role of adenosine A2A receptor in cerebral ischemia reperfusion injury: Signaling to phosphorylated extracellular signal-regulated protein kinase (pERK1/2). Neuroscience 2016, 314, 145–159. [Google Scholar] [CrossRef]
- Zeraati, M.; Mirnajafi-Zadeh, J.; Fathollahi, Y.; Namvar, S.; Rezvani, M.E. Adenosine A1 and A2A receptors of hippocampal CA1 region have opposite effects on piriform cortex kindled seizures in rats. Seizure 2006, 15, 41–48. [Google Scholar] [CrossRef]
- Li, X.; Kang, H.; Liu, X.; Liu, Z.; Shu, K.; Chen, X.; Zhu, S. Effect of adenosine A2A receptor antagonist ZM241385 on amygdala-kindled seizures and progression of amygdala kindling. J. Huazhong Univ. Sci. Technol. Med. Sci. 2012, 32, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Rai, S.; Hsieh, K.C.; McGinty, D.; Alam, M.N.; Szymusiak, R. Adenosine A2A receptors regulate the activity of sleep regulatory GABAergic neurons in the preoptic hypothalamus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R31–R41. [Google Scholar] [CrossRef] [PubMed]
- Knezevic, N.N.; Cicmil, N.; Knezevic, I.; Candido, K.D. Discontinued neuropathic pain therapy between 2009-2015. Expert Opin. Investig. Drugs 2015, 24, 1631–1646. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, S.P.; Slager, S.L.; De Leon, A.B.; Heiman, G.A.; Klein, D.F.; Hodge, S.E.; Weissman, M.M.; Fyer, A.J.; Knowles, J.A. Evidence for genetic linkage between a polymorphism in the adenosine 2A receptor and panic disorder. Neuropsychopharmacology 2004, 29, 558–565. [Google Scholar] [CrossRef]
- Yacoubi, M.E.; Ledent, C.; Parmentier, M.; Bertorelli, R.; Ongini, E.; Costentin, J.; Vaugeois, J.M. Adenosine A 2A receptor antagonists are potential antidepressants: Evidence based on pharmacology and A 2A receptor knockout mice. Br. J. Pharmacol. 2001, 134, 68–77. [Google Scholar] [CrossRef]
- El Yacoubi, M.; Costentin, J.; Vaugeois, J.M. Adenosine A2A receptors and depression. Neurology 2003, 61, S82–S87. [Google Scholar] [CrossRef]
- Yamada, K.; Kobayashi, M.; Kanda, T. Involvement of adenosine A2A receptors in depression and anxiety. Int. Rev. Neurobiol. 2014, 119, 373–393. [Google Scholar]
- Wardas, J. Potential role of adenosine A2A receptors in the treatment of schizophrenia. Front. Biosci. 2008, 13, 4071–4096. [Google Scholar] [CrossRef]
- Boison, D.; Singer, P.; Shen, H.-Y.; Feldon, J.; Yee, B.K. Adenosine hypothesis of schizophrenia-opportunities for pharmacotherapy. Neuropharmacology 2012, 62, 1527–1543. [Google Scholar] [CrossRef]
- Popoli, P.; Blum, D.; Domenici, M.; Burnouf, S.; Chern, Y. A Critical Evaluation of Adenosine A2A Receptors as Potentially “Druggable” Targets in Huntingtons Disease. Curr. Pharm. Des. 2008, 14, 1500–1511. [Google Scholar] [CrossRef]
- Faivre, E.; Coelho, J.E.; Zornbach, K.; Malik, E.; Baqi, Y.; Schneider, M.; Cellai, L.; Carvalho, K.; Sebda, S.; Figeac, M.; et al. Beneficial effect of a selective adenosine A2A receptor antagonist in the APPswe/PS1dE9 mouse model of Alzheimer’s disease. Front. Mol. Neurosci. 2018, 11, 235. [Google Scholar] [CrossRef] [PubMed]
- Morelli, M.; Carta, A.R.; Jenner, P. Adenosine A2A receptors and Parkinson’s disease. In Adenosine Receptors in Health and Disease; Wilson, C.N., Mustafa, S.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 193, pp. 589–615. [Google Scholar]
- Ballesteros-Yáñez, I.; Castillo, C.A.; Merighi, S.; Gessi, S. The role of adenosine receptors in psychostimulant addiction. Front. Pharmacol. 2018, 8, 985. [Google Scholar] [CrossRef] [PubMed]
- Knapp, C.M.; Foye, M.M.; Cottam, N.; Ciraulo, D.A.; Kornetsky, C. Adenosine agonists CGS 21680 and NECA inhibit the initiation of cocaine self-administration. Pharmacol. Biochem. Behav. 2001, 68, 797–803. [Google Scholar] [CrossRef]
- Linden, J. New insights into the regulation of inflammation by adenosine. J. Clin. Investig. 2006, 116, 1835–1837. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B. Adenosine, an endogenous distress signal, modulates tissue damage and repair. Cell Death Differ. 2007, 14, 1315–1323. [Google Scholar] [CrossRef] [PubMed]
- Ryzhov, S.; Zaynagetdinov, R.; Goldstein, A.E.; Novitskiy, S.V.; Blackburn, M.R.; Biaggioni, I.; Feoktistov, I. Effect of A2B adenosine receptor gene ablation on adenosine-dependent regulation of proinflammatory cytokines. J. Pharmacol. Exp. Ther. 2008, 324, 694–700. [Google Scholar] [CrossRef]
- Zhong, H.; Wu, Y.; Belardinelli, L.; Zeng, D. A2B adenosine induce IL-19 from bronchial epithelial cells, resulting in TNF-α increase. Am. J. Respir. Cell Mol. Biol. 2006, 35, 587–592. [Google Scholar] [CrossRef]
- Popoli, P.; Blum, D.; Martire, A.; Ledent, C.; Ceruti, S.; Abbracchio, M.P. Functions, dysfunctions and possible therapeutic relevance of adenosine A2A receptors in Huntington’s disease. Prog. Neurobiol. 2007, 81, 331–348. [Google Scholar] [CrossRef]
- Nazario, L.R.; da Silva, R.S.; Bonan, C.D. Targeting adenosine signaling in Parkinson’s disease: From pharmacological to non-pharmacological approaches. Front. Neurosci. 2017, 11, 658. [Google Scholar] [CrossRef]
- Von Lubitz, D.K.; Lin, R.C.; Popik, P.; Carter, M.F.; Jacobson, K.A. Adenosine A3 receptor stimulation and cerebral ischemia. Eur. J. Pharmacol. 1994, 263, 59–67. [Google Scholar] [CrossRef]
- Von Lubitz, D.K.; Simpson, K.L.; Lin, R.C. Right Thing at a Wrong Time? Adenosine A3 Receptors and Cerebroprotection in Stroke. Ann. N. Y. Acad. Sci. 2001, 939, 85–96. [Google Scholar] [CrossRef]
- Roseti, C.; Palma, E.; Martinello, K.; Fucile, S.; Morace, R.; Esposito, V.; Cantore, G.; Arcella, A.; Giangaspero, F.; Aronica, E.; et al. Blockage of A2A and A3 adenosine receptors decreases the desensitization of human GABAA receptors microtransplanted to Xenopus oocytes. Proc. Natl. Acad. Sci. USA 2009, 106, 15927–15931. [Google Scholar] [CrossRef]
- Ryzhov, S.; Zaynagetdinov, R.; Goldstein, A.E.; Novitskiy, S.V.; Dikov, M.M.; Blackburn, M.R.; Biaggioni, I.; Feoktistov, I. Effect of A2B Adenosine Receptor Gene Ablation on Proinflammatory Adenosine Signaling in Mast Cells. J. Immunol. 2008, 180, 7212–7220. [Google Scholar] [CrossRef]
- Carroll, S.H.; Wigner, N.A.; Kulkarni, N.; Johnston-Cox, H.; Gerstenfeld, L.C.; Ravid, K. A2B adenosine receptor promotes mesenchymal stem cell differentiation to osteoblasts and bone formation in vivo. J. Biol. Chem. 2012, 287, 15718–15727. [Google Scholar] [CrossRef]
- He, W.; Mazumder, A.; Wilder, T.; Cronstein, B.N. Adenosine regulates bone metabolism via A1, A2A, and A2B receptors in bone marrow cells from normal humans and patients with multiple myeloma. FASEB J. 2013, 27, 3446–3454. [Google Scholar] [CrossRef]
- Ryzhov, S.; Sung, B.H.; Zhang, Q.; Weaver, A.; Gumina, R.J.; Biaggioni, I.; Feoktistov, I. Role of adenosine A2B receptor signaling in contribution of cardiac mesenchymal stem-like cells to myocardial scar formation. Purinergic Signal. 2014, 10, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Frick, J.S.; MacManus, C.F.; Scully, M.; Glover, L.E.; Eltzschig, H.K.; Colgan, S.P. Contribution of Adenosine A2B Receptors to Inflammatory Parameters of Experimental Colitis. J. Immunol. 2009, 182, 4957–4964. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Du, C.; Lv, J.; Zhao, G.; Li, Z.; Wu, Z.; Haskó, G.; Xie, X. Blocking A 2B Adenosine Receptor Alleviates Pathogenesis of Experimental Autoimmune Encephalomyelitis via Inhibition of IL-6 Production and Th17 Differentiation. J. Immunol. 2013, 190, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.G.; Balasubramanian, R.; Kiselev, E.; Wei, Q.; Jacobson, K.A. Probing biased/partial agonism at the G protein-coupled A2B adenosine receptor. Biochem. Pharmacol. 2014, 90, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Van Den Berge, M.; Hylkema, M.N.; Versluis, M.; Postma, D.S. Role of adenosine receptors in the treatment of asthma and chronic obstructive pulmonary disease: Recent developments. Drugs R D 2007, 8, 13–23. [Google Scholar] [CrossRef]
- Varani, K.; Caramori, G.; Vincenzi, F.; Adcock, I.; Casolari, P.; Leung, E.; Maclennan, S.; Gessi, S.; Morello, S.; Barnes, P.J.; et al. Alteration of Adenosine Receptors in Patients with Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2006, 173, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Cronstein, B.N. Adenosine receptors and fibrosis: A translational review. F1000 Biol. Rep. 2011, 3, 21. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Belardinelli, L.; Maa, T.; Zeng, D. Synergy between A2B adenosine receptors and hypoxia in activating human lung fibroblasts. Am. J. Respir. Cell Mol. Biol. 2005, 32, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Hoegl, S.; Brodsky, K.S.; Blackburn, M.R.; Karmouty-Quintana, H.; Zwissler, B.; Eltzschig, H.K. Alveolar Epithelial A2B Adenosine Receptors in Pulmonary Protection during Acute Lung Injury. J. Immunol. 2015, 195, 1815–1824. [Google Scholar] [CrossRef]
- Zhan, E.; McIntosh, V.J.; Lasley, R.D. Adenosine A2A and A2B receptors are both required for adenosine A1 receptor-mediated cardioprotection. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H1183–H1189. [Google Scholar] [CrossRef]
- Lasley, R.D. Adenosine receptor-mediated cardioprotection-current limitations and future directions. Front. Pharmacol. 2018, 9, 310. [Google Scholar] [CrossRef]
- Eltzschig, H.K.; Bonney, S.K.; Eckle, T. Attenuating myocardial ischemia by targeting A2B adenosine receptors. Trends Mol. Med. 2013, 19, 345–354. [Google Scholar] [CrossRef]
- Maas, J.E.; Wan, T.C.; Figler, R.A.; Gross, G.J.; Auchampach, J.A. Evidence that the acute phase of ischemic preconditioning does not require signaling by the A2B adenosine receptor. J. Mol. Cell. Cardiol. 2010, 49, 886–893. [Google Scholar] [CrossRef]
- Dubey, R.K.; Gillespie, D.G.; Osaka, K.; Suzuki, F.; Jackson, E.K. Adenosine inhibits growth of rat aortic smooth muscle cells. Possible role of A2b receptor. Hypertension 1996, 27, 786–793. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, Y.; Nguyen, H.G.; Koupenova, M.; Chauhan, A.K.; Makitalo, M.; Jones, M.R.; St. Hilaire, C.; Seldin, D.C.; Toselli, P.; et al. The A2B adenosine receptor protects against inflammation and excessive vascular adhesion. J. Clin. Investig. 2006, 116, 1913–1923. [Google Scholar] [CrossRef]
- Jackson, E.K.; Gillespie, D.G.; Mi, Z.; Cheng, D. Adenosine receptors influence hypertension in dahl salt-sensitive rats: Dependence on receptor subtype, salt diet, and sex. Hypertension 2018, 72, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.R.; Zhou, Z.; Teng, B.; Mustafa, S.J.S. Role of A1 and A2B Adenosine receptors in Angiotensin II dependent hypertension in mice. FASEB J. 2018, 32, 715.2. [Google Scholar]
- Eisenstein, A.; Patterson, S.; Ravid, K. The Many Faces of the A2b Adenosine Receptor in Cardiovascular and Metabolic Diseases. J. Cell. Physiol. 2015, 230, 2891–2897. [Google Scholar] [CrossRef] [PubMed]
- Merighi, S.; Borea, P.A.; Gessi, S. Adenosine receptors and diabetes: Focus on the A2B adenosine receptor subtype. Pharmacol. Res. 2015, 99, 229–236. [Google Scholar] [CrossRef]
- Patel, L.; Thaker, A. The effects of adenosine A2B receptor inhibition on VEGF and nitric oxide axis-mediated renal function in diabetic nephropathy. Ren. Fail. 2014, 36, 916–924. [Google Scholar] [CrossRef]
- Tak, E.; Ridyard, D.; Kim, J.H.; Zimmerman, M.; Werner, T.; Wang, X.X.; Shabeka, U.; Seo, S.W.; Christians, U.; Klawitter, J.; et al. CD73-dependent generation of adenosine and endothelial adora2b signaling attenuate diabetic nephropathy. J. Am. Soc. Nephrol. 2014, 25, 547–563. [Google Scholar] [CrossRef]
- Sun, Y.; Huang, P. Adenosine A2B receptor: From cell biology to human diseases. Front. Chem. 2016, 4, 37. [Google Scholar] [CrossRef]
- Antonioli, L.; Blandizzi, C.; Pacher, P.; Haskó, G. Immunity, inflammation and cancer: A leading role for adenosine. Nat. Rev. Cancer 2013, 13, 842–857. [Google Scholar] [CrossRef]
- Cekic, C.; Sag, D.; Li, Y.; Theodorescu, D.; Strieter, R.M.; Linden, J. Adenosine A 2B Receptor Blockade Slows Growth of Bladder and Breast Tumors. J. Immunol. 2012, 188, 198–205. [Google Scholar] [CrossRef]
- Iannone, R.; Miele, L.; Maiolino, P.; Pinto, A.; Morello, S. Blockade of A2b adenosine receptor reduces tumor growth and immune suppression mediated by myeloid-derived suppressor cells in a mouse model of melanoma. Neoplasia 2013, 15, 1400–1409. [Google Scholar] [CrossRef]
- Ryzhov, S.; Novitskiy, S.V.; Zaynagetdinov, R.; Goldstein, A.E.; Carbone, D.P.; Biaggioni, I.; Dikov, M.M.; Feoktistov, I. Host A2B receptors promote carcinoma growth. Neoplasia 2008, 10, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Merighi, S.; Simioni, C.; Gessi, S.; Varani, K.; Mirandola, P.; Tabrizi, M.A.; Baraldi, P.G.; Borea, P.A. A2B and A3 adenosine receptors modulate vascular endothelial growth factor and interleukin-8 expression in human melanoma cells treated with etoposide and doxorubicin. Neoplasia 2009, 11, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Gessi, S.; Merighi, S.; Sacchetto, V.; Simioni, C.; Borea, P.A. Adenosine receptors and cancer. Biochim. Biophys. Acta Biomembr. 2011, 1808, 1400–1412. [Google Scholar] [CrossRef] [PubMed]
- Ramkumar, V.; Stiles, G.L.; Beaven, M.A.; Ali, H. The A3 adenosine receptor is the unique adenosine receptor which facilitates release of allergic mediators in mast cells. J. Biol. Chem. 1993, 268, 16887–16890. [Google Scholar]
- Walker, B.A.; Jacobson, M.A.; Knight, D.A.; Salvatore, C.A.; Weir, T.; Zhou, D.; Bai, T.R. Adenosine A3 receptor expression and function in eosinophils. Am. J. Respir. Cell Mol. Biol. 1997, 16, 531–537. [Google Scholar] [CrossRef]
- Gao, Z.; Li, B.S.; Day, Y.J.; Linden, J. A3 adenosine receptor activation triggers phosphorylation of protein kinase B and protects rat basophilic leukemia 2H3 mast cells from apoptosis. Mol. Pharmacol. 2001, 59, 76–82. [Google Scholar] [CrossRef]
- Spruntulis, L.M.; Broadley, K.J. A3 receptors mediate rapid inflammatory cell influx into the lungs of sensitized guinea-pigs. Clin. Exp. Allergy 2001, 31, 943–951. [Google Scholar] [CrossRef]
- Fossetta, J.; Jackson, J.; Deno, G.; Fan, X.; Du, X.K.; Bober, L.; Soudé-Bermejo, A.; De Bouteiller, O.; Caux, C.; Lunn, C.; et al. Pharmacological analysis of calcium responses mediated by the human A3 adenosine receptor in monocyte-derived dendritic cells and recombinant cells. Mol. Pharmacol. 2003, 63, 342–350. [Google Scholar] [CrossRef]
- Martin, L.; Pingle, S.C.; Hallam, D.M.; Rybak, L.P.; Ramkumar, V. Activation of the adenosine A3 receptor in RAW 264.7 cells inhibits lipopolysaccharide-stimulated tumor necrosis factor-alpha release by reducing calcium-dependent activation of nuclear factor-kappaB and extracellular signal-regulated kinase 1/2. J. Pharmacol. Exp. Ther. 2006, 316, 71–78. [Google Scholar] [CrossRef]
- Lee, J.Y.; Jhun, B.S.; Oh, Y.T.; Lee, J.H.; Choe, W.; Baik, H.H.; Ha, J.; Yoon, K.S.; Kim, S.S.; Kang, I. Activation of adenosine A3 receptor suppresses lipopolysaccharide-induced TNF-alpha production through inhibition of PI 3-kinase/Akt and NF-kappaB activation in murine BV2 microglial cells. Neurosci. Lett. 2006, 396, 1–6. [Google Scholar] [CrossRef]
- Chen, Y.; Corriden, R.; Inoue, Y.; Yip, L.; Hashiguchi, N.; Zinkernagel, A.; Nizet, V.; Insel, P.A.; Junger, W.G. ATP release guides neutrophil chemotaxis via P2Y2 and A3 receptors. Science 2006, 314, 1792–1795. [Google Scholar] [CrossRef] [PubMed]
- Gessi, S.; Varani, K.; Merighi, S.; Cattabriga, E.; Iannotta, V.; Leung, E.; Baraldi, P.G.; Borea, P.A. A(3) adenosine receptors in human neutrophils and promyelocytic HL60 cells: A pharmacological and biochemical study. Mol. Pharmacol. 2002, 61, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Gessi, S.; Varani, K.; Merighi, S.; Cattabriga, E.; Avitabile, A.; Gavioli, R.; Fortini, C.; Leung, E.; Mac Lennan, S.; Borea, P.A. Expression of A3 Adenosine Receptors in Human Lymphocytes: Up-Regulation in T Cell Activation. Mol. Pharmacol. 2004, 65, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Szabo, C.; Scott, G.S.; Virag, L.; Egnaczyk, G.; Salzman, A.L.; Shanley, T.P.; Hasko, G. Suppression of macrophage inflammatory protein (MIP)-1alpha production and collagen-induced arthritis by adenosine receptor agonists. Br. J. Pharmacol. 1998, 125, 379–387. [Google Scholar] [CrossRef]
- Baharav, E.; Bar-Yehuda, S.; Madi, L.; Silberman, D.; Rath-Wolfson, L.; Halpren, M.; Ochaion, A.; Weinberger, A.; Fishman, P. Antiinflammatory effect of A3 adenosine receptor agonists in murine autoimmune arthritis models. J. Rheumatol. 2005, 32, 469–476. [Google Scholar]
- Silverman, M.H.; Strand, V.; Markovits, D.; Nahir, M.; Reitblat, T.; Molad, Y.; Rosner, I.; Rozenbaum, M.; Mader, R.; Adawi, M.; et al. Clinical evidence for utilization of the A3 adenosine receptor as a target to treat rheumatoid arthritis: Data from a phase II clinical trial. J. Rheumatol. 2008, 35, 41–48. [Google Scholar]
- Cohen, S.; Barer, F.; Bar-Yehuda, S.; Ijzerman, A.P.; Jacobson, K.A.; Fishman, P. A3 adenosine receptor allosteric modulator induces an anti-inflammatory effect: In vivo studies and molecular mechanism of action. Mediat. Inflamm. 2014, 3, 708746. [Google Scholar]
- Lee, H.T.; Kim, M.; Joo, J.D.; Gallos, G.; Chen, J.F.; Emala, C.W. A3 adenosine receptor activation decreases mortality and renal and hepatic injury in murine septic peritonitis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R959–R969. [Google Scholar] [CrossRef]
- Matot, I.; Weiniger, C.F.; Zeira, E.; Galun, E.; Joshi, B.V.; Jacobson, K.A. A3 adenosine receptors and mitogen-activated protein kinases in lung injury following in vivo reperfusion. Crit. Care 2006, 10, R65. [Google Scholar] [CrossRef]
- David, M.; Akerman, L.; Ziv, M.; Kadurina, M.; Gospodinov, D.; Pavlotsky, F.; Yankova, R.; Kouzeva, V.; Ramon, M.; Silverman, M.H.; et al. Treatment of plaque-type psoriasis with oral CF101: Data from an exploratory randomized phase 2 clinical trial. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 361–367. [Google Scholar] [CrossRef]
- Yang, H.; Avila, M.Y.; Peterson-Yantorno, K.; Coca-Prados, M.; Stone, R.A.; Jacobson, K.A.; Civan, M.M. The cross-species A3 adenosine-receptor antagonist MRS 1292 inhibits adenosine-triggered human nonpigmented ciliary epithelial cell fluid release and reduces mouse intraocular pressure. Curr. Eye Res. 2005, 30, 747–754. [Google Scholar] [CrossRef]
- Okamura, T.; Kurogi, Y.; Hashimoto, K.; Sato, S.; Nishikawa, H.; Kiryu, K.; Nagao, Y. Structure-activity relationships of adenosine A3 receptor ligands: New potential therapy for the treatment of glaucoma. Bioorg. Med. Chem. Lett. 2004, 14, 3775–3779. [Google Scholar] [CrossRef]
- Jacobson, K.A.; Civan, M.M. Ocular Purine Receptors as Drug Targets in the Eye. J. Ocul. Pharmacol. Ther. 2016, 32, 534–547. [Google Scholar] [CrossRef]
- Hua, X.; Chason, K.D.; Fredholm, B.B.; Deshpande, D.A.; Penn, R.B.; Tilley, S.L. Adenosine induces airway hyperresponsiveness through activation of A3 receptors on mast cells. J. Allergy Clin. Immunol. 2008, 122, 107–113. [Google Scholar] [CrossRef][Green Version]
- Zhong, H.; Shlykov, S.G.; Molina, J.G.; Sanborn, B.M.; Jacobson, M.A.; Tilley, S.L.; Blackburn, M.R. Activation of murine lung mast cells by the adenosine A3 receptor. J. Immunol. 2003, 171, 338–345. [Google Scholar] [CrossRef]
- Rudich, N.; Ravid, K.; Sagi-Eisenberg, R. Mast cell adenosine receptors function: A focus on the A3 adenosine receptor and inflammation. Front. Immunol. 2012, 3, 134. [Google Scholar] [CrossRef]
- Ezeamuzie, C.I.; Philips, E. Adenosine A3 receptors on human eosinophils mediate inhibition of degranulation and superoxide anion release. Br. J. Pharmacol. 1999, 127, 188–194. [Google Scholar] [CrossRef]
- Young, H.W.J.; Molina, J.G.; Dimina, D.; Zhong, H.; Jacobson, M.; Chan, L.N.L.; Chan, T.S.; Lee, J.J.; Blackburn, M.R. A3 adenosine receptor signaling contributes to airway inflammation and mucus production in adenosine deaminase-deficient mice. J. Immunol. 2004, 173, 1380–1389. [Google Scholar] [CrossRef]
- Rimmer, J.; Peake, H.L.; Santos, C.M.C.; Lean, M.; Bardin, P.; Robson, R.; Haumann, B.; Loehrer, F.; Handel, M.L. Targeting adenosine receptors in the treatment of allergic rhinitis: A randomized, double-blind, placebo-controlled study. Clin. Exp. Allergy 2007, 37, 8–14. [Google Scholar] [CrossRef]
- Morschl, E.; Molina, J.G.; Volmer, J.B.; Mohsenin, A.; Pero, R.S.; Hong, J.S.; Kheradmand, F.; Lee, J.J.; Blackburn, M.R. A3 Adenosine Receptor Signaling Influences Pulmonary Inflammation and Fibrosis. Am. J. Respir. Cell Mol. Biol. 2008, 39, 697–705. [Google Scholar] [CrossRef]
- Hussain, A.; Karjian, P.; Maddock, H. The role of nitric oxide in A3 adenosine receptor-mediated cardioprotection. Auton. Autacoid Pharmacol. 2009, 29, 97–104. [Google Scholar] [CrossRef]
- Maddock, H.L.; Mocanu, M.M.; Yellon, D.M. Adenosine A(3) receptor activation protects the myocardium from reperfusion/reoxygenation injury. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H1307–H1313. [Google Scholar] [CrossRef]
- Headrick, J.P.; Peart, J. A3 adenosine receptor-mediated protection of the ischemic heart. Vascul. Pharmacol. 2005, 42, 271–279. [Google Scholar] [CrossRef]
- Black, R.G.J.; Guo, Y.; Ge, Z.D.; Murphree, S.S.; Prabhu, S.D.; Jones, W.K.; Bolli, R.; Auchampach, J.A. Gene dosage-dependent effects of cardiac-specific overexpression of the A3 adenosine receptor. Circ. Res. 2002, 91, 165–172. [Google Scholar] [CrossRef]
- Hinze, A.V.; Mayer, P.; Harst, A.; von Kugelgen, I. Adenosine A(3) receptor-induced proliferation of primary human coronary smooth muscle cells involving the induction of early growth response genes. J. Mol. Cell. Cardiol. 2012, 53, 639–645. [Google Scholar] [CrossRef]
- Lu, Z.; Fassett, J.; Xu, X.; Hu, X.; Zhu, G.; French, J.; Zhang, P.; Schnermann, J.; Bache, R.J.; Chen, Y. Adenosine A3 receptor deficiency exerts unanticipated protective effects on the pressure-overloaded left ventricle. Circulation 2008, 118, 1713–1721. [Google Scholar] [CrossRef]
- Yang, T.; Zollbrecht, C.; Winerdal, M.E.; Zhuge, Z.; Zhang, X.M.; Terrando, N.; Checa, A.; Sallstrom, J.; Wheelock, C.E.; Winqvist, O.; et al. Genetic Abrogation of Adenosine A3 Receptor Prevents Uninephrectomy and High Salt-Induced Hypertension. J. Am. Heart Assoc. 2016, 5, e003868. [Google Scholar] [CrossRef]
- Ansari, H.R.; Nadeem, A.; Tilley, S.L.; Mustafa, S.J. Involvement of COX-1 in A3 adenosine receptor-mediated contraction through endothelium in mice aorta. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H3448–H3455. [Google Scholar] [CrossRef]
- El-Awady, M.S.; Ansari, H.R.; Fil, D.; Tilley, S.L.; Mustafa, S.J. NADPH oxidase pathway is involved in aortic contraction induced by A3 adenosine receptor in mice. J. Pharmacol. Exp. Ther. 2011, 338, 711–717. [Google Scholar] [CrossRef]
- Grandoch, M.; Hoffmann, J.; Rock, K.; Wenzel, F.; Oberhuber, A.; Schelzig, H.; Fischer, J.W. Novel effects of adenosine receptors on pericellular hyaluronan matrix: Implications for human smooth muscle cell phenotype and interactions with monocytes during atherosclerosis. Basic Res. Cardiol. 2013, 108, 340. [Google Scholar] [CrossRef]
- Gessi, S.; Fogli, E.; Sacchetto, V.; Merighi, S.; Varani, K.; Preti, D.; Leung, E.; Maclennan, S.; Borea, P.A. Adenosine modulates HIF-1{alpha}, VEGF, IL-8, and foam cell formation in a human model of hypoxic foam cells. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 90–97. [Google Scholar] [CrossRef]
- Rothermel, B.A.; Hill, J.A. Adenosine A3 receptor and cardioprotection: Enticing, enigmatic, elusive. Circulation 2008, 118, 1691–1693. [Google Scholar] [CrossRef]
- Rivkees, S.A.; Thevananther, S.; Hao, H. Are A3 adenosine receptors expressed in the brain? Neuroreport 2000, 11, 1025–1030. [Google Scholar] [CrossRef]
- Pugliese, A.M.; Coppi, E.; Volpini, R.; Cristalli, G.; Corradetti, R.; Jeong, L.S.; Jacobson, K.A.; Pedata, F. Role of adenosine A3 receptors on CA1 hippocampal neurotransmission during oxygen-glucose deprivation episodes of different duration. Biochem. Pharmacol. 2007, 74, 768–779. [Google Scholar] [CrossRef]
- Sei, Y.; von Lubitz, D.K.J.E.; Abbracchio, M.P.; Ji, X.; Jacobson, K.A. Adenosine A3 receptor agonist-induced neurotoxicity in rat cerebellar granule neurons. Drug Dev. Res. 1997, 40, 267–273. [Google Scholar] [CrossRef]
- Von Lubitz, D.K.; Lin, R.C.; Boyd, M.; Bischofberger, N.; Jacobson, K.A. Chronic administration of adenosine A3 receptor agonist and cerebral ischemia: Neuronal and glial effects. Eur. J. Pharmacol. 1999, 367, 157–163. [Google Scholar] [CrossRef]
- Wittendorp, M.C.; Boddeke, H.W.G.M.; Biber, K. Adenosine A3 receptor-induced CCL2 synthesis in cultured mouse astrocytes. Glia 2004, 46, 410–418. [Google Scholar] [CrossRef]
- Pugliese, A.M.; Coppi, E.; Spalluto, G.; Corradetti, R.; Pedata, F. A3 adenosine receptor antagonists delay irreversible synaptic failure caused by oxygen and glucose deprivation in the rat CA1 hippocampus in vitro. Br. J. Pharmacol. 2006, 147, 524–532. [Google Scholar] [CrossRef]
- Etherington, L.A.V.; Frenguelli, B.G. Endogenous adenosine modulates epileptiform activity in rat hippocampus in a receptor subtype-dependent manner. Eur. J. Neurosci. 2004, 19, 2539–2550. [Google Scholar] [CrossRef]
- Laudadio, M.A.; Psarropoulou, C. The A3 adenosine receptor agonist 2-Cl-IB-MECA facilitates epileptiform discharges in the CA3 area of immature rat hippocampal slices. Epilepsy Res. 2004, 59, 83–94. [Google Scholar] [CrossRef]
- Zhu, C.B.; Steiner, J.A.; Munn, J.L.; Daws, L.C.; Hewlett, W.A.; Blakely, R.D. Rapid stimulation of presynaptic serotonin transport by A(3) adenosine receptors. J. Pharmacol. Exp. Ther. 2007, 322, 332–340. [Google Scholar] [CrossRef]
- Zhu, C.B.; Lindler, K.M.; Campbell, N.G.; Sutcliffe, J.S.; Hewlett, W.A.; Blakely, R.D. Colocalization and regulated physical association of presynaptic serotonin transporters with A(3) adenosine receptors. Mol. Pharmacol. 2011, 80, 458–465. [Google Scholar] [CrossRef]
- Mabley, J.; Soriano, F.; Pacher, P.; Haskó, G.; Marton, A.; Wallace, R.; Salzman, A.; Szabó, C. The adenosine A3 receptor agonist, N6-(3-iodobenzyl)-adenosine-5′-N-methyluronamide, is protective in two murine models of colitis. Eur. J. Pharmacol. 2003, 466, 323–329. [Google Scholar] [CrossRef]
- Ren, T.; Tian, T.; Feng, X.; Ye, S.; Wang, H.; Wu, W.; Qiu, Y.; Yu, C.; He, Y.; Zeng, J.; et al. An adenosine A3 receptor agonist inhibits DSS-induced colitis in mice through modulation of the NF-κB signaling pathway. Sci. Rep. 2015, 5, 9047. [Google Scholar] [CrossRef]
- Lee, H.T.; Ota-Setlik, A.; Xu, H.; D’Agati, V.D.; Jacobson, M.A.; Emala, C.W. A 3 adenosine receptor knockout mice are protected against ischemia- and myoglobinuria-induced renal failure. Am. J. Physiol. Renal. Physiol. 2003, 284, F267–F273. [Google Scholar] [CrossRef]
- Yap, S.C.; Thomas Lee, H. Adenosine and protection from acute kidney injury. Curr. Opin. Nephrol. Hypertens. 2012, 21, 24–32. [Google Scholar] [CrossRef]
- Madi, L.; Ochaion, A.; Rath-Wolfson, L.; Bar-Yehuda, S.; Erlanger, A.; Ohana, G.; Harish, A.; Merimski, O.; Barer, F.; Fishman, P. The A3 adenosine receptor is highly expressed in tumor versus normal cells: Potential target for tumor growth inhibition. Clin. Cancer Res. 2004, 10, 4472–4479. [Google Scholar] [CrossRef]
- Fishman, P.; Bar-Yehuda, S.; Synowitz, M.; Powell, J.D.; Klotz, K.N.; Gessi, S.; Borea, P.A. Adenosine receptors and cancer. In Adenosine Receptors in Health and Disease; Wilson, C.N., Musta, S.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 399–441. [Google Scholar]
- Gessi, S.; Merighi, S.; Varani, K.; Leung, E.; Mac Lennan, S.; Borea, P.A. The A3 adenosine receptor: An enigmatic player in cell biology. Pharmacol. Ther. 2008, 117, 123–140. [Google Scholar] [CrossRef]
- Fishman, P.; Bar-Yehuda, S.; Vagman, L. Adenosine and other low molecular weight factors released by muscle cells inhibit tumor cell growth. Cancer Res. 1998, 58, 3181–3187. [Google Scholar]
- Fishman, P.; Bar-Yehuda, S.; Ohana, G.; Pathak, S.; Wasserman, L.; Barer, F.; Multani, A.S. Adenosine acts as an inhibitor of lymphoma cell growth: A major role for the A3 adenosine receptor. Eur. J. Cancer 2000, 36, 1452–1458. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, M.; Laties, A.M.; Mitchell, C.H. Balance of purines may determine life or death of retinal ganglion cells as A3 adenosine receptors prevent loss following P2X7 receptor stimulation. J. Neurochem. 2006, 98, 566–575. [Google Scholar] [CrossRef]
- Gessi, S.; Merighi, S.; Varani, K.; Cattabriga, E.; Benini, A.; Mirandola, P.; Leung, E.; Mac Lennan, S.; Feo, C.; Baraldi, S.; et al. Adenosine receptors in colon carcinoma tissues and colon tumoral cell lines: Focus on the A(3) adenosine subtype. J. Cell. Physiol. 2007, 211, 826–836. [Google Scholar] [CrossRef]
- Powis, G.; Kirkpatrick, L. Hypoxia inducible factor-1alpha as a cancer drug target. Mol. Cancer Ther. 2004, 3, 647–654. [Google Scholar]
- Merighi, S.; Benini, A.; Mirandola, P.; Gessi, S.; Varani, K.; Leung, E.; MacLennan, S.; Baraldi, P.G.; Borea, P.A. A3 adenosine receptors modulate hypoxia-inducible factor-1α expression in human A375 melanoma cells. Neoplasia 2005, 7, 894–903. [Google Scholar] [CrossRef]
- Merighi, S.; Benini, A.; Mirandola, P.; Gessi, S.; Varani, K.; Leung, E.; Maclennan, S.; Borea, P.A. Adenosine modulates vascular endothelial growth factor expression via hypoxia-inducible factor-1 in human glioblastoma cells. Biochem. Pharmacol. 2006, 72, 19–31. [Google Scholar] [CrossRef]
- Rocha, R.; Torres, Á.; Ojeda, K.; Uribe, D.; Rocha, D.; Erices, J.; Niechi, I.; Ehrenfeld, P.; San Martín, R.; Quezada, C. The Adenosine A₃ Receptor Regulates Differentiation of Glioblastoma Stem-Like Cells to Endothelial Cells under Hypoxia. Int. J. Mol. Sci. 2018, 19, 1228. [Google Scholar] [CrossRef]
- Ohana, G.; Bar-Yehuda, S.; Arich, A.; Madi, L.; Dreznick, Z.; Rath-Wolfson, L.; Silberman, D.; Slosman, G.; Fishman, P. Inhibition of primary colon carcinoma growth and liver metastasis by the A3 adenosine receptor agonist CF101. Br. J. Cancer 2003, 89, 1552–1558. [Google Scholar] [CrossRef]
- Madi, L.; Bar-Yehuda, S.; Barer, F.; Ardon, E.; Ochaion, A.; Fishman, P. A3 adenosine receptor activation in melanoma cells: Association between receptor fate and tumor growth inhibition. J. Biol. Chem. 2003, 278, 42121–42130. [Google Scholar] [CrossRef]
- Fishman, P.; Bar-Yehuda, S.; Ardon, E.; Rath-Wolfson, L.; Barrer, F.; Ochaion, A.; Madi, L. Targeting the A3 adenosine receptor for cancer therapy: Inhibition of prostate carcinoma cell growth by A3AR agonist. Anticancer Res. 2003, 23, 2077–2083. [Google Scholar]
- Bar-Yehuda, S.; Stemmer, S.M.; Madi, L.; Castel, D.; Ochaion, A.; Cohen, S.; Barer, F.; Zabutti, A.; Perez-Liz, G.; Del Valle, L.; et al. The A3 adenosine receptor agonist CF102 induces apoptosis of hepatocellular carcinoma via de-regulation of the Wnt and NF-κB signal transduction pathways. Int. J. Oncol. 2008, 33, 287–295. [Google Scholar]
- Fishman, P.; Bar-Yehuda, S.; Madi, L.; Cohn, I. A3 adenosine receptor as a target for cancer therapy. Anti-Cancer Drugs 2002, 13, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Marucci, G.; Santinelli, C.; Buccioni, M.; Navia, A.M.; Lambertucci, C.; Zhurina, A.; Yli-Harja, O.; Volpini, R.; Kandhavelu, M. Anticancer activity study of A3 adenosine receptor agonists. Life Sci. 2018, 205, 155–163. [Google Scholar] [CrossRef]



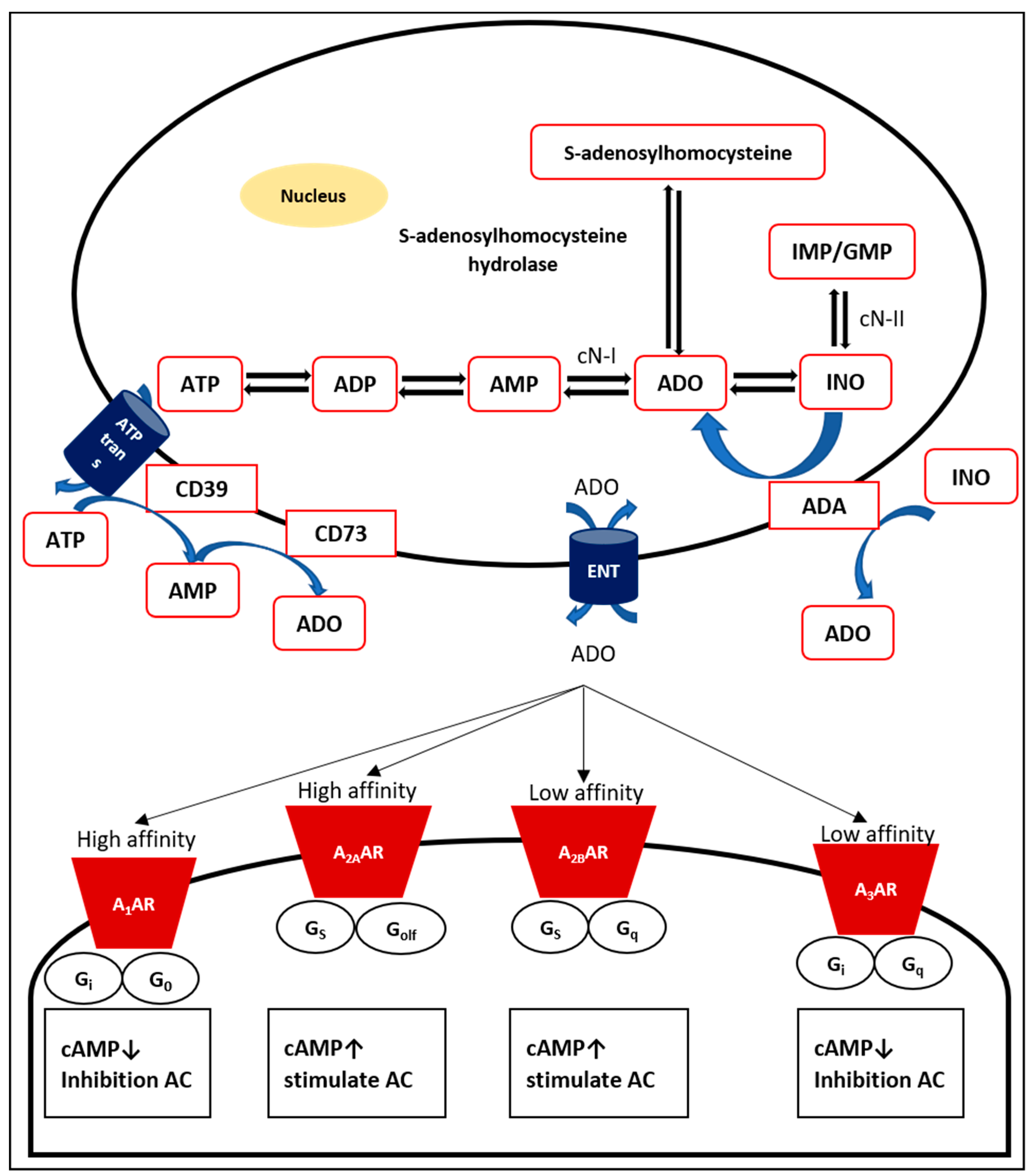{kind=link}
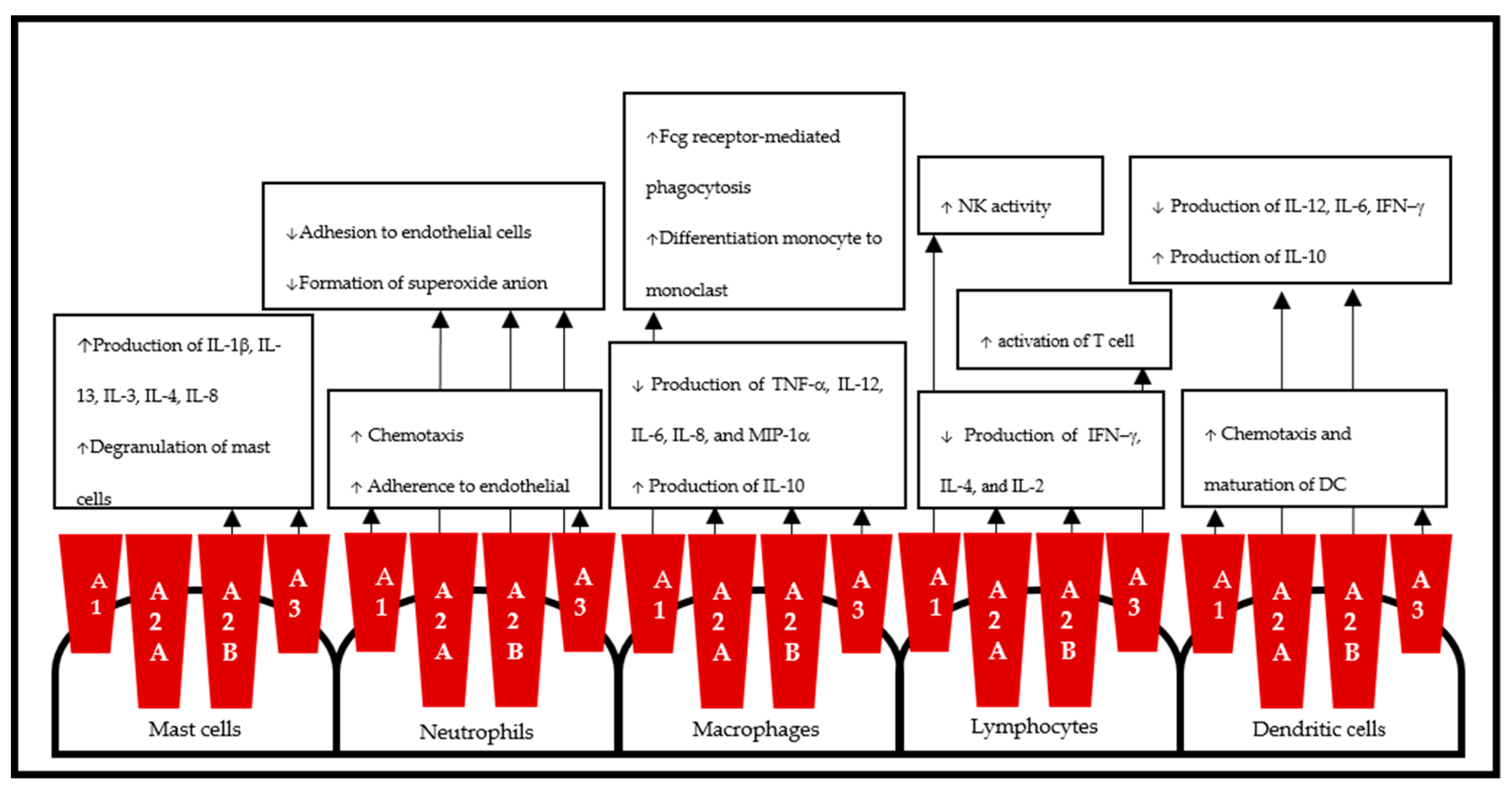{kind=link}
| Receptors | Diseases | Drugs | |
|---|---|---|---|
| Selective Full Agonist | Antagonist/Partial Agonist | ||
| A1AR | COPD and asthma | L-97-1 [87] EPI-2010 [88] | |
| Lung injury | CCPA [91] | ||
| A2AAR | COPD and asthma | CGS-21680 [109] UK371,104 (DC) [110] GW328267X (DC) [111] Regadenoson [112] Apadenoson [59] | |
| Lung injury | CGS-21680 [113] PTX [114] | ||
| A2BAR | COPD and asthma | BAY 60-6853 [115] | CVT-6883 [116] CGS15493, WO-00125210, and ATL-907 [86] |
| Lung injury | BAY 60-6583 [117] | ||
| Fibrosis and interstitial lung diseases | GS-6201 [118] | CVT-6883 [119] QAF 805 [85] | |
| A3AR | COPD and asthma | QAF 805 [85] | |
| Lung injury | IB-MECA [120] | ||
| Receptors | Functions | Diseases | Drugs | ||
|---|---|---|---|---|---|
| Selective Full Agonist | Antagonist/Partial Agonist | ||||
| A1AR | Inotropic and adrenergic control | Negative | Heart failure | CPA [106] | BG9928 [107] Rolofylline [108] Capadenoson [122] Neladenoson [124] |
| Dromotropic and chronotropic control | Negative | Arrhythmia | Tecadenoson (DC)[128] Selodenoson [129] PJ-875 [129] | CVT-2759 [130] | |
| Vascular control | Constriction | Hypertension | |||
| Ischemia | Protection | Angina | Capadenoson [121] | ||
| Adaptation | Hypertrophy | CPA [106] | |||
| Angiogenesis | |||||
| Diabetes mellitus and hyperlipidemia | GR79236 and ARA [165] SDZ WAG994 and RPR749 [123] Tecadenoson [168] | GS-9667/CVT-3619 [167] BW-1433 [168] | |||
| Glaucoma | Trabodenoson (DC) [123] | ||||
| Glomerulonephritis | Metrifudil 2 (DC) [123] | ||||
| A2AAR | Inotropic and adrenergic control | Positive | Heart failure | LASSBio-294 [184] | |
| Vascular control | Dilatation | Imaging | Regadenoson, binodenoson, evodenoson, sonedenoson, and apadenoson [123,185] | ||
| Anti-inflammation | Platelet aggregation | PSB-15826, PSB-12404, and PSB-16301 [186] | |||
| Cholesterol homeostasis | ATL313 [187] | ||||
| Ischemia | Protection | Angina | CGS 21680C [180] ATL-193/ATL-146e [181] | ||
| Adaptation | Hypertrophy | ||||
| Angiogenesis | |||||
| A2BAR | Vascular control | Dilatation | |||
| Anti-inflammation | Atherosclerosis and hyperlipidemia | BAY 60-6853 [189] | |||
| Ischemia | Protection | Angina | BAY 60-6583 [190] | ||
| Adaptation | Hypertrophy | ||||
| Angiogenesis | GS-6201 [191] | ||||
| Diabetes mellitus and hyperlipidemia | NECA [192] ATL-801 [193] MRS-1754 [194] | ||||
| A3AR | Vascular control | Constriction | |||
| Anti-inflammation | Diabetic kidney diseases | LJ-2698 [195] | |||
| Atherosclerosis and hyperlipidemia | LJ-1888 [196] | ||||
| Ischemia | Protection | Angina | IB-MECA [197] CI-IB-MECA [198] CP-532,903 [199] | ||
| Adaptation | Hypertrophy | ||||
| Angiogenesis | |||||
| Receptors | Diseases | Drugs | ||
|---|---|---|---|---|
| Selective Full Agonist | Antagonist/Partial Agonist | Allosteric Modulators | ||
| A1AR | Stroke | NNC-21-0136 [123] | ||
| Sleep | CPA [155] | |||
| Anxiety and depression | MRS5474 [151] | TRR469 [150] | ||
| Cognition and memory | ASP5854 [159] | |||
| Alzheimer’s disease | ||||
| Huntington’s diseases | CPA, CCPA, CHA and ADAC [162] | |||
| Schizophrenia | 2-CLA. NECA, CHA, CPA, PIA, L-PIA [210] | CPT, 8-CPT2, DPCPX, and MSX-3 [210] | ||
| Pain | GR79236X [138] Cl-ENBA [139] MRS7469 [140] SDZ WAG 994, GR79236, GW- 493838 (DC) [141] | T62 (DC) [141] | ||
| Epilepsy | DPCPX [146] | |||
| A2AAR | Stroke | SCH58261 [200] | ||
| Sleep | CGS21680 [133] | |||
| Anxiety and depression | Istradefylline [208] | |||
| Cognition and memory | ASP5854 [159] | |||
| Alzheimer’s disease | MSX-3 [212] | |||
| Huntington’s diseases | ZM241385 [220] | |||
| Parkinson’s diseases | Tozadenant/SYN115, DT1133, ZM241385, ST1535, and istradefylline [61] Preladenant, ASP5854, vipadenant (DC) [61] SCH900800, and BIIB014 [221] KW-6002 [134] | |||
| Schizophrenia | APEC, CGS21680, NECA, CV-1808, and DPMA [210] | MSX-3, DMPX, SCH58261, and ZM241385 [210] | ||
| Pain | CGS21680 [155] Spongosine (DC) [204] | |||
| Epilepsy | ZM241385 [202] | |||
| Drug addiction | CGS21680 and NECA [215] | |||
| A3AR | Stroke | IB-MECA [222] CI-IB-MECA [223] | LJ-1251 [72] | |
| Epilepsy | ANR235 [224] | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Effendi, W.I.; Nagano, T.; Kobayashi, K.; Nishimura, Y. Focusing on Adenosine Receptors as a Potential Targeted Therapy in Human Diseases. Cells 2020, 9, 785. https://doi.org/10.3390/cells9030785
Effendi WI, Nagano T, Kobayashi K, Nishimura Y. Focusing on Adenosine Receptors as a Potential Targeted Therapy in Human Diseases. Cells. 2020; 9(3):785. https://doi.org/10.3390/cells9030785
Chicago/Turabian StyleEffendi, Wiwin Is, Tatsuya Nagano, Kazuyuki Kobayashi, and Yoshihiro Nishimura. 2020. "Focusing on Adenosine Receptors as a Potential Targeted Therapy in Human Diseases" Cells 9, no. 3: 785. https://doi.org/10.3390/cells9030785
APA StyleEffendi, W. I., Nagano, T., Kobayashi, K., & Nishimura, Y. (2020). Focusing on Adenosine Receptors as a Potential Targeted Therapy in Human Diseases. Cells, 9(3), 785. https://doi.org/10.3390/cells9030785