PPE51 Is Involved in the Uptake of Disaccharides by Mycobacterium tuberculosis

 , , , ,
, , , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Selection of Mutants and Determination of Mutation Frequency
2.3. Gene Cloning Strategies
2.4. Next-Generation Sequencing
2.5. Labeling of Thio-Disaccharide by Tritiation
2.6. T-6 Accumulation Assay
2.7. Protein Purification
2.8. Production of Anti-PPE51 Rabbit Polyvalent Serum
2.9. PPE51–T-6 Direct Interaction
2.10. CRISPR/Cas9
2.11. qRT-PCR
3. Results
3.1. Mutated PPE51 Affects the Sensitivity of Mtb to Thio-Disaccharide
3.1.1. Selection of T-6 Resistant Mutants
3.1.2. M. tuberculosis T-6 Resistant Mutants Accumulate Mutations in ppe51
3.1.3. PPE51 Mutations Do Not Affect Mtb Resistance to Various Anti-TB Drugs
3.2. PPE51 Is Involved in the Uptake of Thio-Disaccharide by Tubercle Bacilli
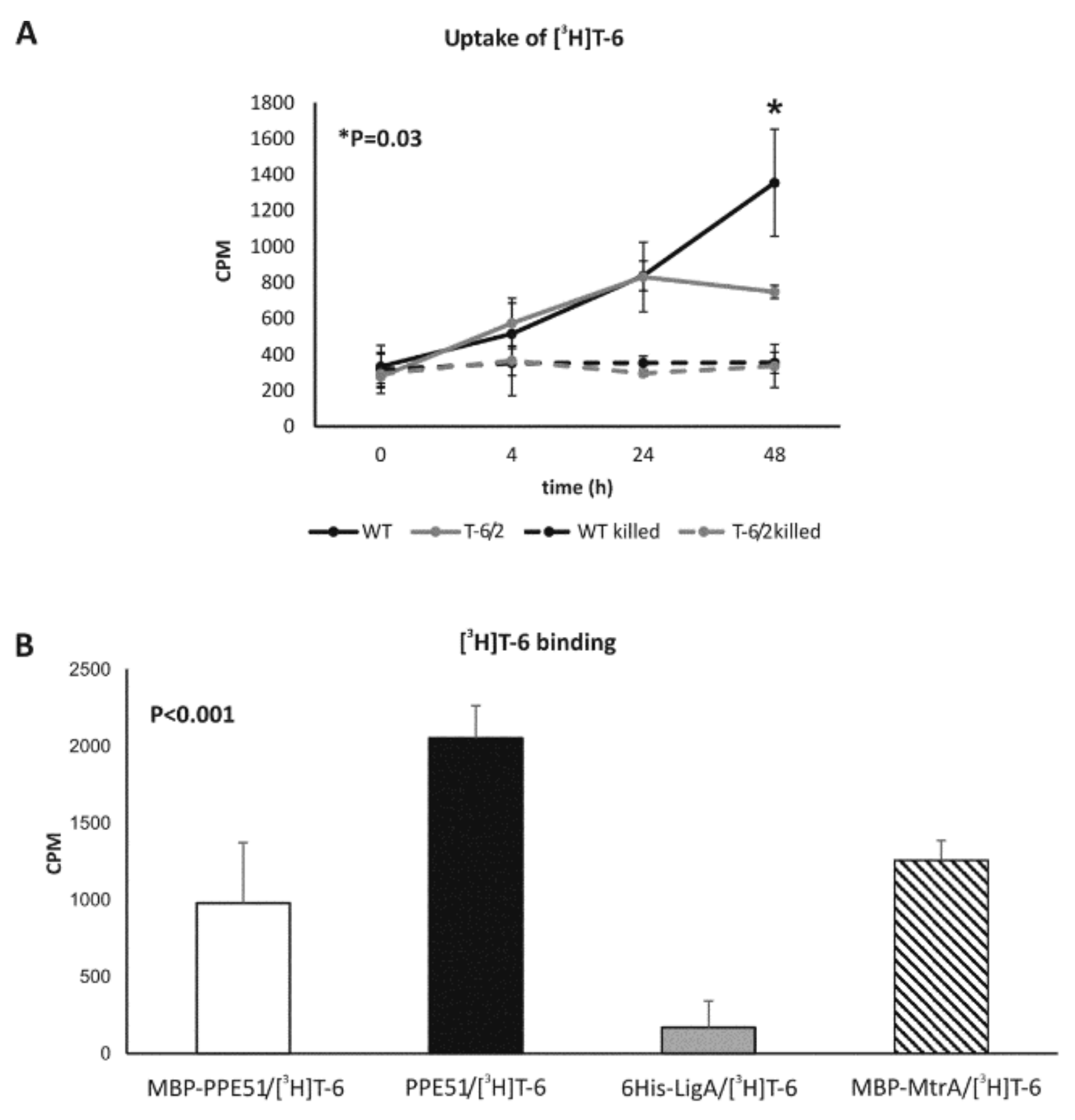
3.2.1. The Uptake of T-6 Is Affected in Mtb ppe51 Mutants
3.2.2. PPE51 Binds to T-6
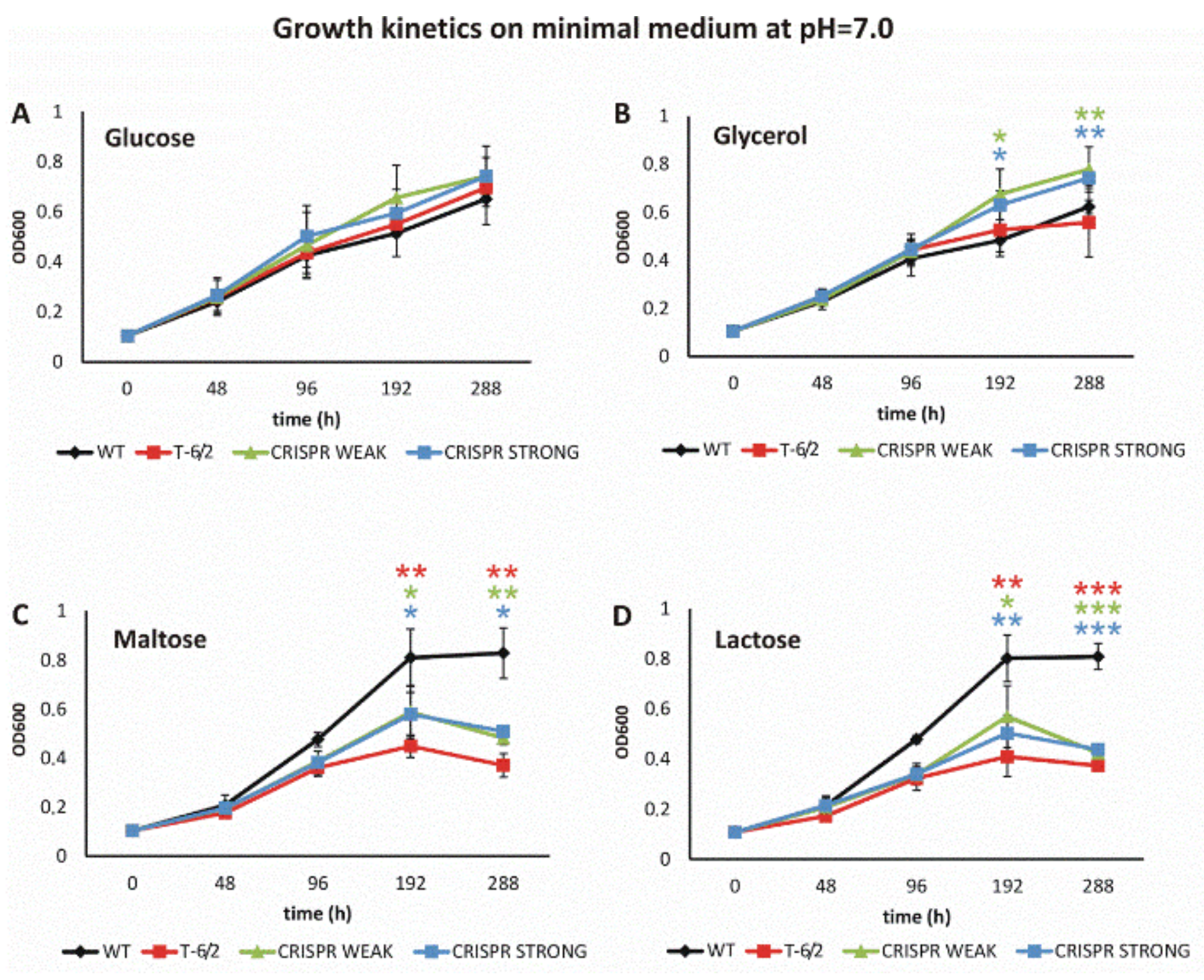
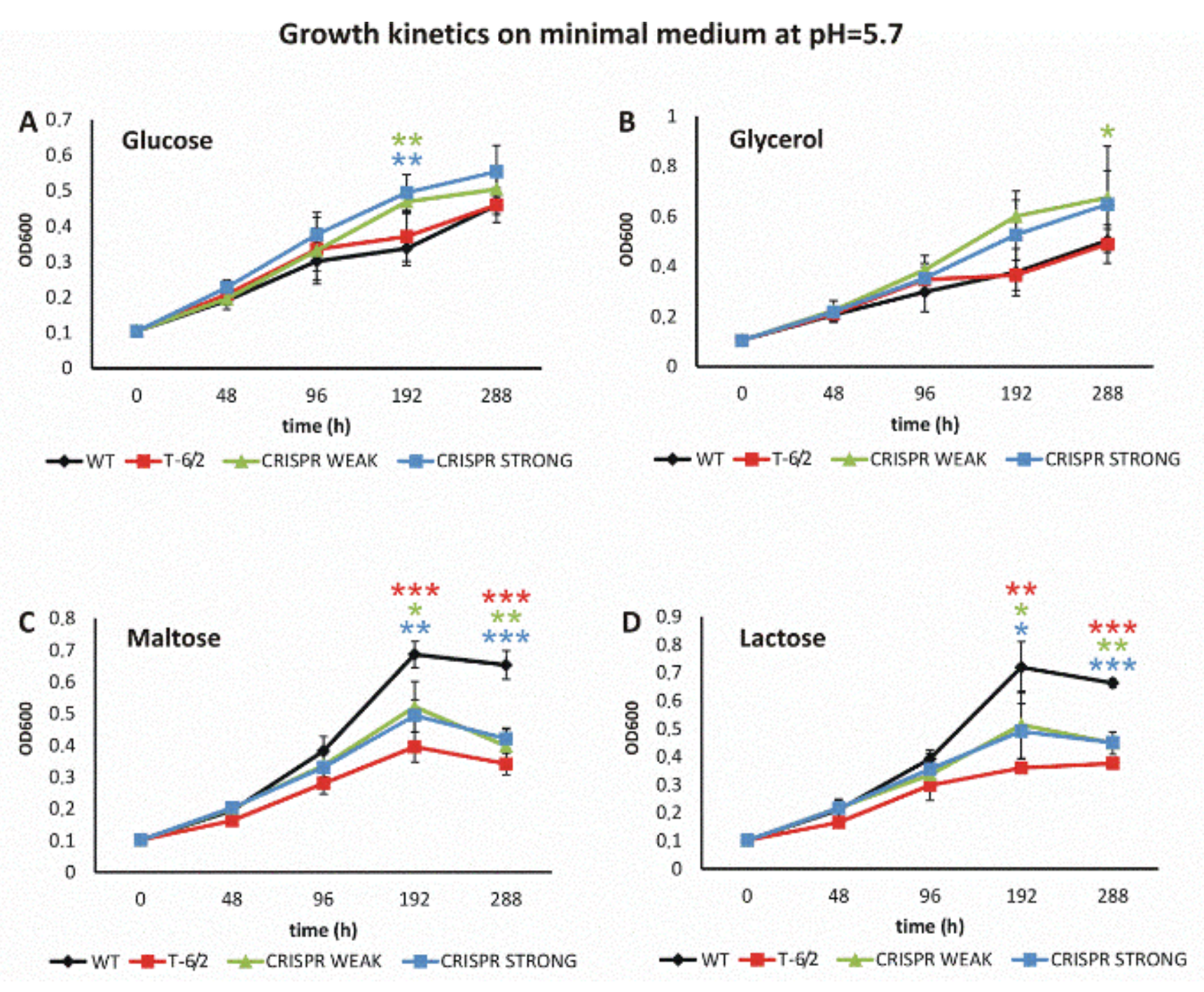
3.3. Depletion of PPE51 Affects the Growth of Mtb in Medium with Disaccharides
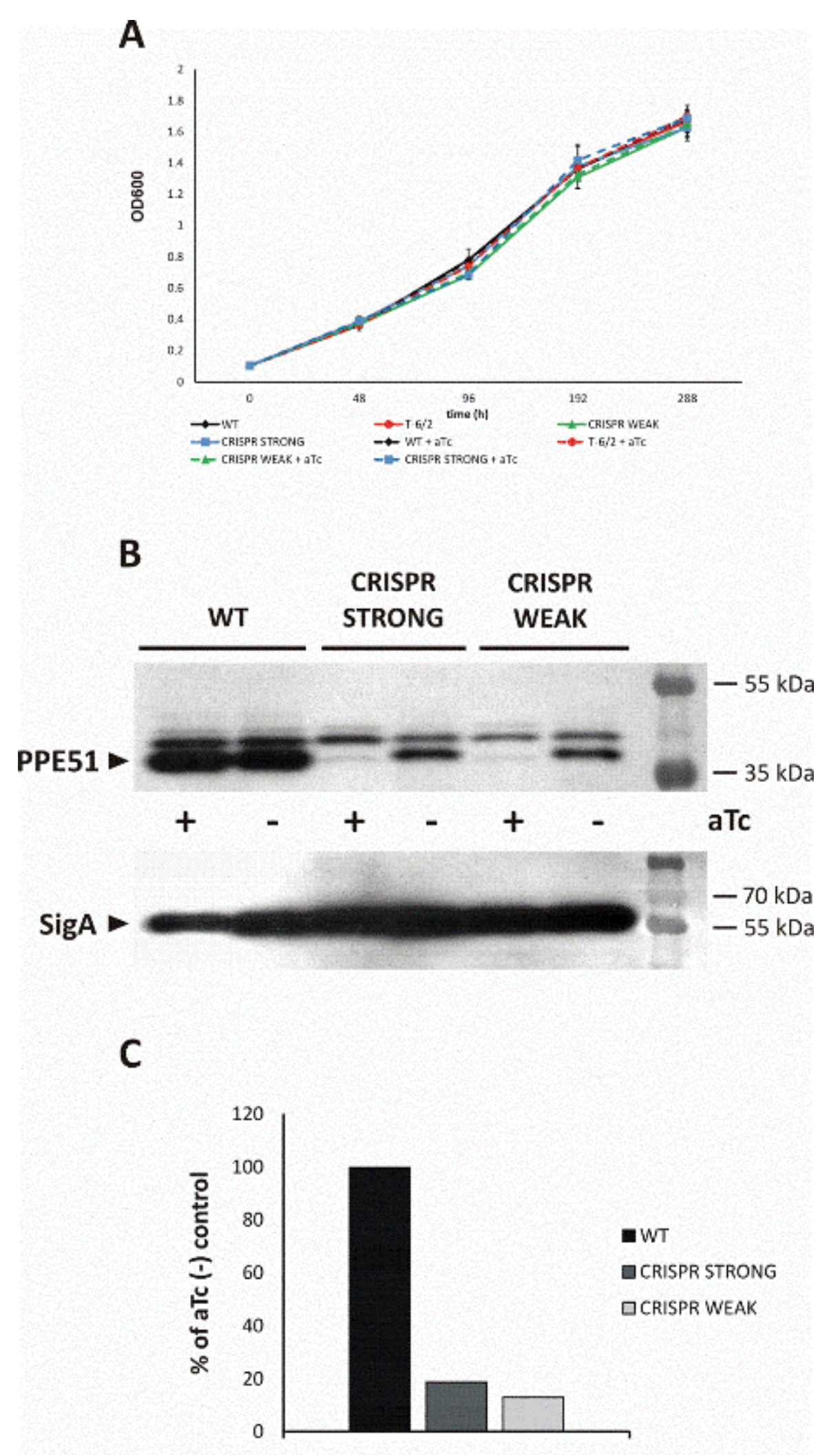
3.3.1. PPE51 Depletion Does Not Affect Mtb Growth in Rich Media
3.3.2. Mutation or Depletion of ppe51 Affect Mtb Growth on Disaccharides
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Global Tuberculosis Report. World Health Organization: Geneva, 2019. Available online: https://apps.who.int/iris/bitstream/handle/10665/329368/9789241565714-eng.pdf?ua=1 (accessed on 17 February 2020).
- WHO. Global Tuberculosis Report. World Health Organization: Geneva, 2017. Available online: http://apps.who.int/medicinedocs/documents/s23360en/s23360en.pdf (accessed on 17 February 2020).
- Jagielski, T.; Minias, A.; van Ingen, J.; Rastogi, N.; Brzostek, A.; Zaczek, A.; Dziadek, J. Methodological and clinical aspects of the molecular epidemiology of mycobacterium tuberculosis and other mycobacteria. Clin. Microbiol. Rev. 2016, 29, 239–290. [Google Scholar] [CrossRef] [Green Version]
- Cole, S.T.; Brosch, R.; Parkhill, J.; Garnier, T.; Churcher, C.; Harris, D.; Gordon, S.V.; Eiglmeier, K.; Gas, S.; Barry, C.E., 3rd.; et al. Deciphering the biology of mycobacterium tuberculosis from the complete genome sequence. Nature 1998, 393, 537–544. [Google Scholar] [CrossRef]
- Sampson, S.L. Mycobacterial pe/ppe proteins at the host-pathogen interface. Clin. Dev. Immunol. 2011, 497203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ates, L.S. New insights into the mycobacterial pe and ppe proteins provide a framework for future research. Mol. Microbiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Camus, J.C.; Pryor, M.J.; Medigue, C.; Cole, S.T. Re-annotation of the genome sequence of mycobacterium tuberculosis h37rv. Microbiol-Sgm 2002, 148, 2967–2973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gey van Pittius, N.C.; Sampson, S.L.; Lee, H.; Kim, Y.; van Helden, P.D.; Warren, R.M. Evolution and expansion of the mycobacterium tuberculosis pe and ppe multigene families and their association with the duplication of the esat-6 (esx) gene cluster regions. BMC Evol. Biol. 2006, 6, 95. [Google Scholar] [CrossRef] [Green Version]
- Delogu, G.; Brennan, M.J.; Manganelli, R. Pe and ppe genes: A tale of conservation and diversity. Adv. Exp. Med. Biol. 2017, 1019, 191–207. [Google Scholar]
- Abdallah, A.M.; Verboom, T.; Weerdenburg, E.M.; Gey van Pittius, N.C.; Mahasha, P.W.; Jimenez, C.; Parra, M.; Cadieux, N.; Brennan, M.J.; Appelmelk, B.J.; et al. Ppe and pe_pgrs proteins of mycobacterium marinum are transported via the type vii secretion system esx-5. Mol. Microbiol. 2009, 73, 329–340. [Google Scholar] [CrossRef]
- Daleke, M.H.; Cascioferro, A.; de Punder, K.; Ummels, R.; Abdallah, A.M.; van der Wel, N.; Peters, P.J.; Luirink, J.; Manganelli, R.; Bitter, W. Conserved pro-glu (pe) and pro-pro-glu (ppe) protein domains target lipy lipases of pathogenic mycobacteria to the cell surface via the esx-5 pathway. J. Biol. Chem. 2011, 286, 19024–19034. [Google Scholar] [CrossRef] [Green Version]
- Ekiert, D.C.; Cox, J.S. Structure of a pe-ppe-espg complex from mycobacterium tuberculosis reveals molecular specificity of esx protein secretion. Proc. Natl. Acad. Sci. USA 2014, 111, 14758–14763. [Google Scholar] [CrossRef] [Green Version]
- Daleke, M.H.; van der Woude, A.D.; Parret, A.H.; Ummels, R.; de Groot, A.M.; Watson, D.; Piersma, S.R.; Jimenez, C.R.; Luirink, J.; Bitter, W.; et al. Specific chaperones for the type vii protein secretion pathway. J. Biol. Chem. 2012, 287, 31939–31947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ates, L.S.; Ummels, R.; Commandeur, S.; van de Weerd, R.; Sparrius, M.; Weerdenburg, E.; Alber, M.; Kalscheuer, R.; Piersma, S.R.; Abdallah, A.M.; et al. Essential role of the esx-5 secretion system in outer membrane permeability of pathogenic mycobacteria. PLoS Genet. 2015, 11, e1005190. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.; Speer, A.; Lin, K.; Ehrt, S.; Niederweis, M. Ppe surface proteins are required for heme utilization by mycobacterium tuberculosis. mBio 2017, 8, e01720-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tullius, M.V.; Nava, S.; Horwitz, M.A. Ppe37 is essential for mycobacterium tuberculosis heme-iron acquisition (hia), and a defective ppe37 in mycobacterium bovis bcg prevents hia. Infect. Immun. 2019, 87, e00540-18. [Google Scholar] [CrossRef] [Green Version]
- Banu, S.; Honore, N.; Saint-Joanis, B.; Philpott, D.; Prevost, M.C.; Cole, S.T. Are the pe-pgrs proteins of mycobacterium tuberculosis variable surface antigens? Mol. Microbiol. 2002, 44, 9–19. [Google Scholar] [CrossRef]
- Brennan, M.J.; Delogu, G.; Chen, Y.P.; Bardarov, S.; Kriakov, J.; Alavi, M.; Jacobs, W.R. Evidence that mycobacterial pe_pgrs proteins are cell surface constituents that influence interactions with other cells. Infect. Immun. 2001, 69, 7326–7333. [Google Scholar] [CrossRef] [Green Version]
- Fishbein, S.; van Wyk, N.; Warren, R.M.; Sampson, S.L. Phylogeny to function: Pe/ppe protein evolution and impact on mycobacterium tuberculosis pathogenicity. Mol. Microbiol. 2015, 96, 901–916. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, B.M.; Kannan, N.; Vemu, L.; Raghunand, T.R. The mycobacterium tuberculosis pe proteins rv0285 and rv1386 modulate innate immunity and mediate bacillary survival in macrophages. PLoS ONE 2012, 7, e51686. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Pathak, S.K.; Banerjee, A.; Pathak, S.; Bhattacharyya, A.; Yang, Z.H.; Talarico, S.; Kundu, M.; Basu, J. Execution of macrophage apoptosis by pe_pgrs33 of mycobacterium tuberculosis is mediated by toll-like receptor 2-dependent release of tumor necrosis factor-alpha. J. Biol. Chem. 2007, 282, 1039–1050. [Google Scholar] [CrossRef] [Green Version]
- Dheenadhayalan, V.; Delogu, G.; Brennan, M.J. Expression of the pe_pgrs 33 protein in mycobacterium smegmatis triggers necrosis in macrophages and enhanced mycobacterial survival. Microbes Infect. 2006, 8, 262–272. [Google Scholar] [CrossRef]
- Bansal, K.; Sinha, A.Y.; Ghorpade, D.S.; Togarsimalemath, S.K.; Patil, S.A.; Kaveri, S.V.; Balaji, K.N.; Bayry, J. Src homology 3-interacting domain of rv1917c of mycobacterium tuberculosis induces selective maturation of human dendritic cells by regulating pi3k-mapk-nf-kappa b signaling and drives th2 immune responses. J. Biol. Chem. 2010, 285, 36511–36522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tundup, S.; Mohareer, K.; Hasnain, S.E. Mycobacterium tuberculosis pe25/ppe41 protein complex induces necrosis in macrophages: Role in virulence and disease reactivation? FEBS Open Biol. 2014, 4, 822–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, X.; Luo, T.; Zhai, X.Q.; Zhang, C.X.; Suo, J.; Ma, P.J.; Wang, C.H.; Bao, L. Ppe11 of mycobacterium tuberculosis can alter host inflammatory response and trigger cell death. Microb. Pathog. 2019, 126, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Bitter, W.; Houben, E.N.G.; Bottai, D.; Brodin, P.; Brown, E.J.; Cox, J.S.; Derbyshire, K.; Fortune, S.M.; Gao, L.Y.; Liu, J.; et al. Systematic genetic nomenclature for type vii secretion systems. PLoS Pathog. 2009, 5, e1000507. [Google Scholar] [CrossRef] [Green Version]
- Houben, E.N.; Bestebroer, J.; Ummels, R.; Wilson, L.; Piersma, S.R.; Jimenez, C.R.; Ottenhoff, T.H.; Luirink, J.; Bitter, W. Composition of the type vii secretion system membrane complex. Mol. Microbiol. 2012, 86, 472–484. [Google Scholar] [CrossRef] [PubMed]
- Groschel, M.I.; Sayes, F.; Simeone, R.; Majlessi, L.; Brosch, R. Esx secretion systems: Mycobacterial evolution to counter host immunity. Nat. Rev. Microbiol. 2016, 14, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Beckham, K.S.; Ciccarelli, L.; Bunduc, C.M.; Mertens, H.D.; Ummels, R.; Lugmayr, W.; Mayr, J.; Rettel, M.; Savitski, M.M.; Svergun, D.I.; et al. Structure of the mycobacterial esx-5 type vii secretion system membrane complex by single-particle analysis. Nat. Microbiol. 2017, 2, 17047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegrist, M.S.; Unnikrishnan, M.; McConnell, M.J.; Borowsky, M.; Cheng, T.Y.; Siddiqi, N.; Fortune, S.M.; Moody, D.B.; Rubin, E.J. Mycobacterial esx-3 is required for mycobactin-mediated iron acquisition. Proc. Natl. Acad. Sci. USA 2009, 106, 18792–18797. [Google Scholar] [CrossRef] [Green Version]
- Tufariello, J.M.; Chapman, J.R.; Kerantzas, C.A.; Wong, K.W.; Vilcheze, C.; Jones, C.M.; Cole, L.E.; Tinaztepe, E.; Thompson, V.; Fenyo, D.; et al. Separable roles for mycobacterium tuberculosis esx-3 effectors in iron acquisition and virulence. Proc. Natl. Acad. Sci. USA 2016, 113, E348–E357. [Google Scholar] [CrossRef] [Green Version]
- Bottai, D.; Di Luca, M.; Majlessi, L.; Frigui, W.; Simeone, R.; Sayes, F.; Bitter, W.; Brennan, M.J.; Leclerc, C.; Batoni, G.; et al. Disruption of the esx-5 system of mycobacterium tuberculosis causes loss of ppe protein secretion, reduction of cell wall integrity and strong attenuation. Mol. Microbiol. 2012, 83, 1195–1209. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.; Cannon, J.R.; Fenselau, C.; Briken, V. A duplicated esat-6 region of esx-5 is involved in protein export and virulence of mycobacteria. Infect. Immun. 2015, 83, 4349–4361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ates, L.S.; Dippenaar, A.; Ummels, R.; Piersma, S.R.; van der Woude, A.D.; van der Kuij, K.; Le Chevalier, F.; Mata-Espinosa, D.; Barrios-Payan, J.; Marquina-Castillo, B.; et al. Mutations in ppe38 block pe_pgrs secretion and increase virulence of mycobacterium tuberculosis. Nat. Microbiol. 2018, 3, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Lovewell, R.R.; Sassetti, C.M.; VanderVen, B.C. Chewing the fat: Lipid metabolism and homeostasis during m. Tuberculosis infection. Curr. Opin. Microbiol. 2016, 29, 30–36. [Google Scholar] [CrossRef]
- Fitzgerald, D.W.; Sterling, T.R.; Haas, D.W. Mycobacterium Tuberculosis. In Principles and Practice of Infectious Diseases; Mandell, G.L., Bennett, J.E., Dolin, R., Eds.; Churchill Livingstone: Philadelphia, PA, USA, 2009. [Google Scholar]
- Von Gottberg, A.; Sacks, L.; Machala, S.; Blumberg, L. Utility of blood cultures and incidence of mycobacteremia in patients with suspected tuberculosis in a south african infectious disease referral hospital. Int. J. Tuberc. Lung Dis. 2001, 5, 80–86. [Google Scholar] [PubMed]
- De Carvalho, L.P.; Fischer, S.M.; Marrero, J.; Nathan, C.; Ehrt, S.; Rhee, K.Y. Metabolomics of mycobacterium tuberculosis reveals compartmentalized co-catabolism of carbon substrates. Chem. Biol. 2010, 17, 1122–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korycka-Machala, M.; Brzostek, A.; Dziadek, B.; Kawka, M.; Poplawski, T.; Witczak, Z.J.; Dziadek, J. Evaluation of the mycobactericidal effect of thio-functionalized carbohydrate derivatives. Molecules 2017, 22, 812–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.; VanderVen, B.C.; Fahey, R.J.; Russell, D.G. Intracellular mycobacterium tuberculosis exploits host-derived fatty acids to limit metabolic stress. J. Biol. Chem. 2013, 288, 6788–6800. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.F.; Russell, D.W. Molecular Coning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Walker, I.H.; Hsieh, P.C.; Riggs, P.D. Mutations in maltose-binding protein that alter affinity and solubility properties. Appl. Microbiol. Biotechnol. 2010, 88, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Sheffield, P.; Gerrard, S.; Derewenda, Z. Overcomming expression and purificatio problems of rhogdi using a family of ∜parallel∝ expression vector. Protein Express. Purif. 1999, 15, 34–39. [Google Scholar] [CrossRef]
- Minias, A.E.; Brzostek, A.M.; Korycka-Machala, M.; Dziadek, B.; Minias, P.; Rajagopalan, M.; Madiraju, M.; Dziadek, J. Rnase hi is essential for survival of mycobacterium smegmatis. PLoS ONE 2015, 10, e0126260. [Google Scholar] [CrossRef] [Green Version]
- Korycka-Machala, M.; Rychta, E.; Brzostek, A.; Sayer, H.R.; Rumijowska-Galewicz, A.; Bowater, R.P.; Dziadek, J. Evaluation of nad(+)-dependent DNA ligase of mycobacteria as a potential target for antibiotics. Antimicrob. Agents Chemother. 2007, 51, 2888–2897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rock, J.M.; Hopkins, F.F.; Chavez, A.; Diallo, M.; Chase, M.R.; Gerrick, E.R.; Pritchard, J.R.; Church, G.M.; Rubin, E.J.; Sassetti, C.M.; et al. Programmable transcriptional repression in mycobacteria using an orthogonal crispr interference platform. Nat. Microbiol. 2017, 2, 16274. [Google Scholar] [CrossRef] [PubMed]
- Plocinski, P.; Macios, M.; Houghton, J.; Niemiec, E.; Plocinska, R.; Brzostek, A.; Slomka, M.; Dziadek, J.; Young, D.; Dziembowski, A. Proteomic and transcriptomic experiments reveal an essential role of rna degradosome complexes in shaping the transcriptome of mycobacterium tuberculosis. Nucleic Acids Res. 2019, 47, 5892–5905. [Google Scholar] [CrossRef] [PubMed]
- Korycka-Machala, M.; Viljoen, A.; Pawelczyk, J.; Borowka, P.; Dziadek, B.; Gobis, K.; Brzostek, A.; Kawka, M.; Blaise, M.; Strapagiel, D.; et al. 1h-benzo[d]imidazole derivatives affect mmpl3 in mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2019, 63, e00441-19. [Google Scholar] [CrossRef] [PubMed]
- Gorla, P.; Plocinska, R.; Sarva, K.; Satsangi, A.T.; Pandeeti, E.; Donnelly, R.; Dziadek, J.; Rajagopalan, M.; Madiraju, M.V. Mtra response regulator controls cell division and cell wall metabolism and affects susceptibility of mycobacteria to the first line antituberculosis drugs. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Ramakrishnan, P.; Aagesen, A.M.; McKinney, J.D.; Tischler, A.D. Mycobacterium tuberculosis resists stress by regulating pe19 expression. Infect. Immun. 2015, 84, 735–746. [Google Scholar] [CrossRef] [Green Version]
- Tischler, A.D.; Leistikow, R.L.; Kirksey, M.A.; Voskuil, M.I.; McKinney, J.D. Mycobacterium tuberculosis requires phosphate-responsive gene regulation to resist host immunity. Infect. Immun. 2013, 81, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Wayne, L.G.; Hayes, L.G. An in vitro model for sequential study of shiftdown of mycobacterium tuberculosis through two stages of nonreplicating persistence. Infect. Immun. 1996, 64, 2062–2069. [Google Scholar] [CrossRef] [Green Version]
- Baker, J.J.; Abramovitch, R.B. Genetic and metabolic regulation of mycobacterium tuberculosis acid growth arrest. Sci. Rep. 2018, 8, 4168. [Google Scholar] [CrossRef] [Green Version]
- Hirayama, B.A.; Loo, D.D.F.; Wright, E.M. Cation effects on protein conformation and transport in the na+/glucose cotransporter. J. Biol. Chem. 1997, 272, 2110–2115. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Thio-Disaccharide − T-6 | ||
|---|---|---|
| Strains | MIC50 (µM) | MIC90 (µM) |
| H37Rv | 50 | 100 |
| PPE51 mutants | ||
| T-6/2 − (A96S) | 550 | 700 |
| T-6/7 − (A66fs) | 550 | 700 |
| T-6/10 − (T83K) | 550 | 700 |
| T-6/22 − (L95P) | 550 | 700 |
| H37Rv + mutated PPE51 | ||
| H37Rv + (A96S) − T-6/2 | 50 | 100 |
| H37Rv + (A66fs) − T-6/7 | 50 | 100 |
| H37Rv + (T83K) − T-6/10 | 50 | 100 |
| H37Rv + (L95P) − T-6/22 | 50 | 100 |
| PPE51 mutants + wild-type PPE51 | ||
| T-6/2 + PPE51 | 50 | 100 |
| T-6/7 + PPE51 | 50 | 100 |
| T-6/10 + PPE51 | 50 | 100 |
| T-6/22 + PPE51 | 50 | 100 |
| Strains | INH | EMB | RIF | STR | TLM |
|---|---|---|---|---|---|
| MIC90 (µM) | |||||
| Rv | 0.365 | 7.2 | 0.6 | 1.4 | 119 |
| PPE51 (A96S) − T-6/2 | 0.365 | 7.2 | 0.6 | 1.4 | 119 |
| PPE51 (A66fs) − T-6/7 | 0.365 | 7.2 | 0.6 | 1.4 | 119 |
| PPE51 (T83K) − T-6/10 | 0.365 | 7.2 | 0.6 | 1.4 | 119 |
| PPE51 (L95P) − T-6/22 | 0.365 | 7.2 | 0.6 | 1.4 | 119 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korycka-Machała, M.; Pawełczyk, J.; Borówka, P.; Dziadek, B.; Brzostek, A.; Kawka, M.; Bekier, A.; Rykowski, S.; Olejniczak, A.B.; Strapagiel, D.; et al. PPE51 Is Involved in the Uptake of Disaccharides by Mycobacterium tuberculosis. Cells 2020, 9, 603. https://doi.org/10.3390/cells9030603
Korycka-Machała M, Pawełczyk J, Borówka P, Dziadek B, Brzostek A, Kawka M, Bekier A, Rykowski S, Olejniczak AB, Strapagiel D, et al. PPE51 Is Involved in the Uptake of Disaccharides by Mycobacterium tuberculosis. Cells. 2020; 9(3):603. https://doi.org/10.3390/cells9030603
Chicago/Turabian StyleKorycka-Machała, Małgorzata, Jakub Pawełczyk, Paulina Borówka, Bożena Dziadek, Anna Brzostek, Malwina Kawka, Adrian Bekier, Sebastian Rykowski, Agnieszka B. Olejniczak, Dominik Strapagiel, and et al. 2020. "PPE51 Is Involved in the Uptake of Disaccharides by Mycobacterium tuberculosis" Cells 9, no. 3: 603. https://doi.org/10.3390/cells9030603
APA StyleKorycka-Machała, M., Pawełczyk, J., Borówka, P., Dziadek, B., Brzostek, A., Kawka, M., Bekier, A., Rykowski, S., Olejniczak, A. B., Strapagiel, D., Witczak, Z., & Dziadek, J. (2020). PPE51 Is Involved in the Uptake of Disaccharides by Mycobacterium tuberculosis. Cells, 9(3), 603. https://doi.org/10.3390/cells9030603