Stress Reshapes the Physiological Response of Halophile Fungi to Salinity



 ,
, 
 , , , and
, , , and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Strain and Culture Conditions
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Molecular Phylogeny
2.4. RNA Extraction, Library Construction, and Sequencing
2.5. Transcriptome Assembly and Transcript Quantification
2.6. Differential Transcript Expression Analysis
2.7. qPCR and Transcriptome Validation
2.8. Electron Microscopy and TEM Image Analysis
2.9. Quantification of Compatible Solutes
2.10. Data Availability
3. Results and Discussion
3.1. Identification of a Terrestrial Isolate of Aspergillus sydowii
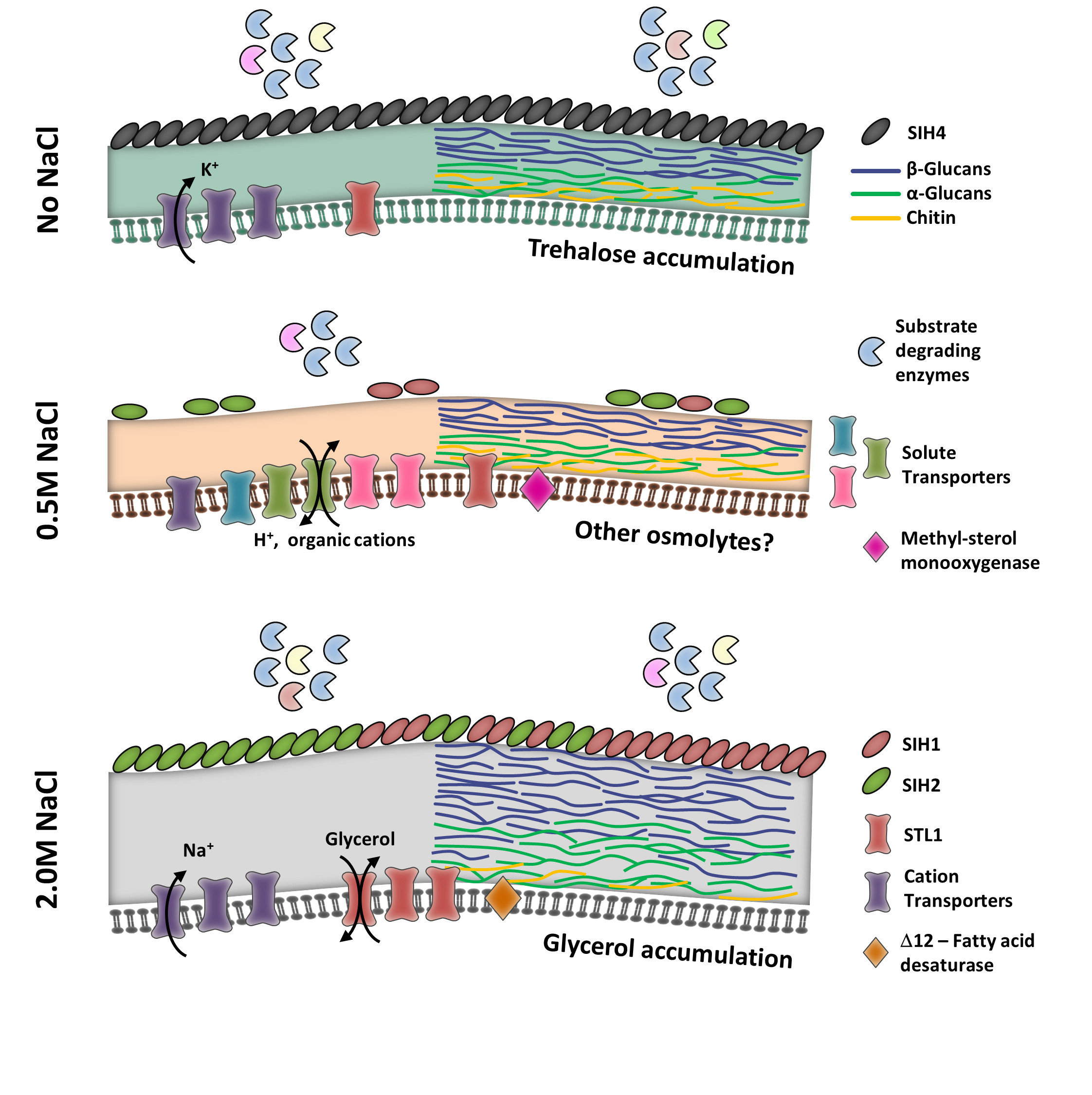
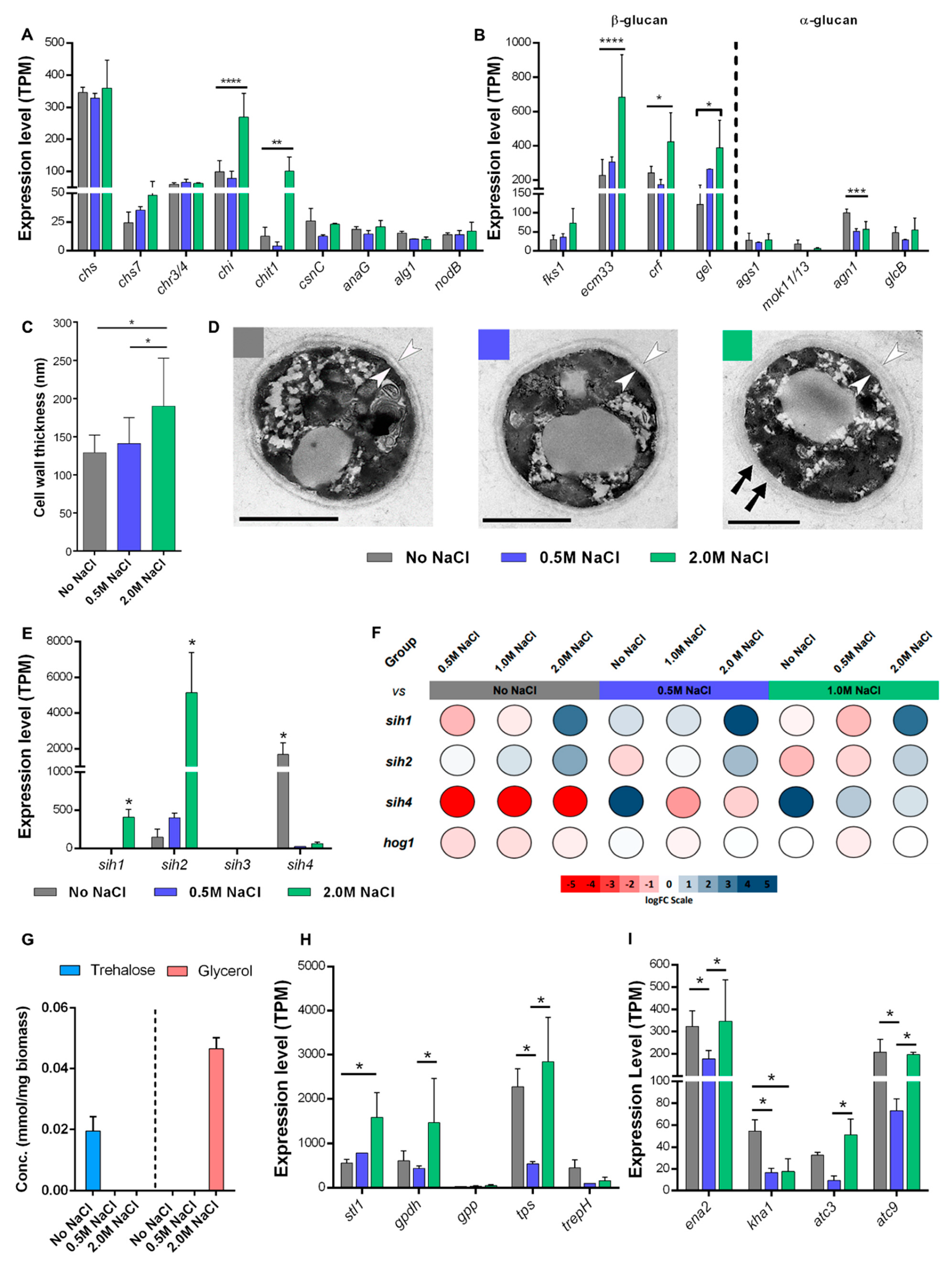
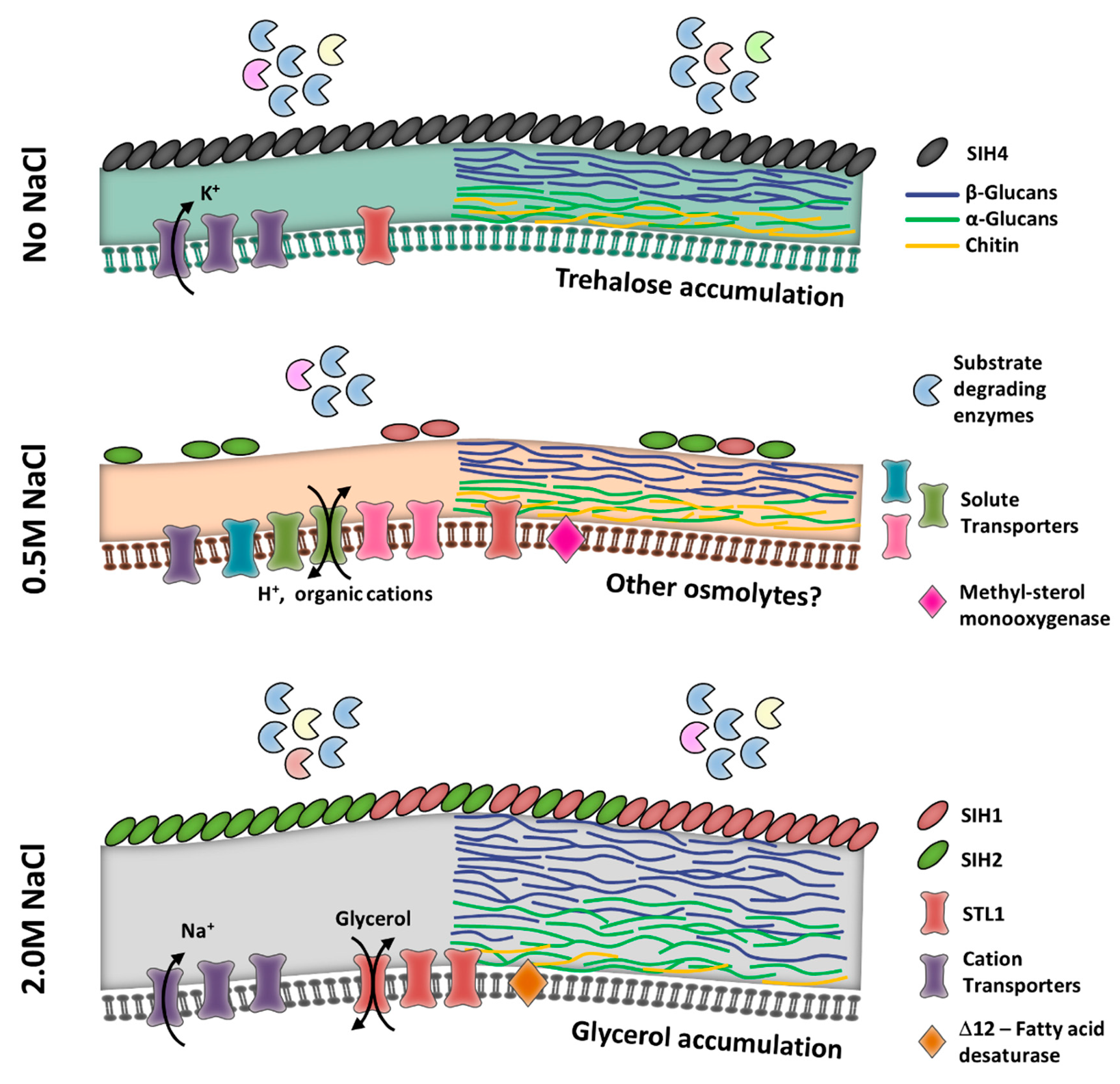
3.2. Thriving and Enduring Require a Different Set of Adaptations
3.3. Membrane and Cell Wall Are the Main Defenses under Saline Stress
3.4. Coating for the Occasion
3.5. Compatible Solutes
3.6. Membrane Transporter Expression Is Not Pivotal for Eluding Ion Toxicity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, G.W.; Ives, L.D.; Nagelkerken, I.A.; Ritchie, K.B. Caribbean sea-fan mortalities. Nature 1996, 383, 487. [Google Scholar] [CrossRef]
- Geiser, D.M.; Taylor, J.W.; Ritchie, K.B.; Smith, G.W. Cause of sea fan death in the West Indies. Nature 1998, 394, 137–138. [Google Scholar] [CrossRef]
- Soler-Hurtado, M.M.; Sandoval-Sierra, V.; Machordom, A.; Dieguez-Uribeondo, J. Aspergillus sydowii and Other Potential Fungal Pathogens in Gorgonian Octocorals of the Ecuadorian Pacific. PLoS ONE 2016, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Butinar, L.; Frisvad, J.C.; Gunde-cimerman, N. Hypersaline waters—A potential source of foodborne toxigenic aspergilli and penicillia. FEMS Microbiol. Lett. 2011, 77, 186–199. [Google Scholar] [CrossRef]
- Zajc, J.; Zalar, P.; Plemenitas, A.; Gunde-Cimerman, N. The mycobiota of the Salterns. In Biology of Marine Fungi; Raghukumar, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 53, pp. 133–158. ISBN 978-3-642-23341-8. [Google Scholar]
- Pitt, J.I.; Hocking, A.D. Spoilage of Stored, Processed and Preserved Foods. In Fungi and Food Spoilage; Pitt, J.I., Hocking, A.D., Eds.; Springer: Boston, MA, USA, 2009; pp. 402–421. ISBN 9780387922065. [Google Scholar]
- Cong, B.; Wang, N.; Liu, S.; Liu, F.; Yin, X.; Shen, J. Isolation, characterization and transcriptome analysis of a novel Antarctic Aspergillus sydowii strain MS-19 as a potential lignocellulosic enzyme source. BMC Microbiol. 2017, 17, 1–14. [Google Scholar] [CrossRef]
- Schmitt, E.K.; Eilinghoff, B.; Olliger, R.; Decker, H.; Kück, U. Development of molecular tools for the mulundocandin producer Aspergillus sydowii: DNA-mediated transformation and reporter gene expression. Appl. Microbiol. Biotechnol. 2002, 58, 625–631. [Google Scholar] [CrossRef]
- Matkar, K.; Chapla, D.; Divecha, J.; Nighojkar, A.; Madamwar, D. Production of cellulase by a newly isolated strain of Aspergillus sydowii and its optimization under submerged fermentation. Int. Biodeterior. Biodegradation 2013, 78, 24–33. [Google Scholar] [CrossRef]
- Alker, A.P.; Smith, G.W.; Kim, K. Characterization of Aspergillus sydowii (Thom et Church), a fungal pathogen of Caribbean sea fan corals. Hydrobiologia 2001, 460, 105–111. [Google Scholar] [CrossRef]
- Wang, J.; Lin, X.; Qin, C.; Liao, S.; Wan, J.; Zhang, T.; Liu, J.; Fredimoses, M.; Chen, H.; Yang, B.; et al. Antimicrobial and antiviral sesquiterpenoids from sponge-associated fungus, Aspergillus sydowii ZSDS1-F6. J. Antibiot. 2014, 67, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.G.; Shankar, R.S. Enzymatic bleaching of kraft pulp by xylanase from Aspergillus sydowii SBS 45. Indian J. Microbiol. 2010, 50, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Madhu, K.M.; Beena, P.S.; Chandrasekaran, M. Extracellular β -glucosidase Production by a Marine Aspergillus sydowii BTMFS 55 under Solid State Fermentation Using Statistical Experimental Design. Biotechnol. Bioprocess Eng. 2009, 14, 457–466. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Plemenitas, A.; Oren, A. Strategies of adaptation of microorganisms of the three domains of life to high salt concentrations. FEMS Microbiol. Rev. 2018, 42, 353–375. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Yu, F.; Tan, Y.; Zhang, X.; Liu, Z. Comparative Transcriptome Sequence Analysis of Sporulation-Related Genes of Aspergillus cristatus in Response to Low and High Osmolarity. Curr. Microbiol. 2017, 74, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.H.; Ding, X.W.; Narsing Rao, M.P.; Zhang, B.; Zhang, Y.G.; Liu, F.H.; Liu, B.B.; Xiao, M.; Li, W.J. Morphological and transcriptomic analysis reveals the osmoadaptive response of endophytic fungus Aspergillus montevidensis ZYD4 to high salt stress. Front. Microbiol. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, J.; Wu, Y.; Ren, Y.; Liu, Q.; Wang, Q.; Zhou, X.; Cai, M.; Zhang, Y. De novo transcriptome sequencing of marine-derived Aspergillus glaucus and comparative analysis of metabolic and developmental variations in response to salt stress. Genes Genom. 2017, 39, 317–329. [Google Scholar] [CrossRef]
- Salamov, A.; Sun, H.; Lipzen, A.; Kis-papo, T.; Weig, A.R.; Riley, R.; Pers, D.; Wasser, S.P.; Rambold, G.; Grigoriev, I.V.; et al. Genomic adaptations of the halophilic Dead Sea filamentous fungus Eurotium rubrum. Nat. Commun. 2014, 5, 1–8. [Google Scholar]
- Saum, S.H.; Müller, V. Regulation of osmoadaptation in the moderate halophile Halobacillus halophilus: Chloride, glutamate and switching osmolyte strategies. Saline Syst. 2008, 4, 1–15. [Google Scholar] [CrossRef]
- Batista-García, R.A.; Balcázar-López, E.; Miranda-Miranda, E.; Sánchez-Reyes, A.; Cuervo-Soto, L.; Aceves-Zamudio, D.; Atriztán-Hernández, K.; Morales-Herrera, C.; Rodríguez-Hernández, R.; Folch-Mallol, J. Characterization of lignocellulolytic activities from a moderate halophile strain of Aspergillus caesiellus isolated from a sugarcane bagasse fermentation. PLoS ONE 2014, 9, 1–12. [Google Scholar]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.-B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Szigeti, G.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 141–173. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed]
- Gouy, M.; Guindon, S.; Gascuel, O. Sea view version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Macmanes, M.D.; et al. De novo transcript sequence reconstruction from RNA-Seq: Reference generation and analysis with Trinity. Nat Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Conesa, A.; Madrigal, P.; Tarazona, S.; Gomez-Cabrero, D.; Cervera, A.; McPherson, A.; Szczesniak, M.W.; Gaffney, D.J.; Elo, L.L.; Zhang, X.; et al. A survey of best practices for RNA-seq data analysis. Genome Biol. 2016, 17, 19. [Google Scholar] [CrossRef]
- Wagner, G.P.; Kin, K.; Lynch, V.J. Measurement of mRNA abundance using RNA-seq data: RPKM measure is inconsistent among samples. Theory Biosci. 2012, 131, 281–285. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-seq data with or without a reference genome. BMC Bioinformatics 2011, 12, 41–74. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2013, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Risso, D.; Ngai, J.; Speed, T.P.; Dudoit, S. Normalization of RNA-seq data using factor analysis of control genes or samples. Nat. Biotechnol. 2014, 32, 896–902. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Bohle, K.; Jungebloud, A.; Dalpiaz, A. Selection of reference genes for normalisation of specific gene quantification data of Aspergillus niger. J. Biotechnol. 2007, 132, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36–e46. [Google Scholar] [CrossRef]
- Kuncic, M.K.; Kogej, T.; Drobne, D.; Gunde-Cimerman, N. Morphological Response of the Halophilic Fungal Genus Wallemia to High Salinity. Appl. Environ. Microbiol. 2010, 76, 329–337. [Google Scholar] [CrossRef]
- Kogej, T.; Stein, M.; Volkmann, M.; Gorbushina, A.A.; Galinski, E.A.; Gunde-Cimerman, N. Osmotic adaptation of the halophilic fungus Hortaea werneckii: Role of osmolytes and melanization. Microbiology 2007, 153, 4261–4273. [Google Scholar] [CrossRef]
- Stielow, J.B.; Lévesque, C.A.; Seifert, K.A.; Meyer, W.; Irinyi, L.; Smits, D.; Renfurm, R.; Verkley, G.J.M.; Groenewald, M.; Chaduli, D.; et al. One fungus, which genes? Development and assessment of universal primers for potential secondary fungal DNA barcodes. Persoonia 2015, 35, 242–263. [Google Scholar] [CrossRef]
- Jurjevic, Z.; Peterson, S.W.; Horn, B.W. Aspergillus section Versicolores: Nine new species and multilocus DNA sequence based phylogeny. IMA Fungus 2012, 3, 59–79. [Google Scholar] [CrossRef]
- Sklenář, F.; Jurjević, Z.; Zalar, P.; Frisvad, J.C.; Visagie, C.M.; Kolařík, M.; Houbraken, J.; Chen, A.J.; Yilmaz, N.; Seifert, K.A.; et al. Phylogeny of xerophilic aspergilli (subgenus Aspergillus) and taxonomic revision of section Restricti. Stud. Mycol. 2017, 88, 161–236. [Google Scholar] [CrossRef]
- Siqueira, J.P.Z.; Sutton, D.A.; García, D.; Gené, J.; Thomson, P.; Wiederhold, N.; Guarro, J. Species diversity of Aspergillus section Versicolores in clinical samples and antifungal susceptibility. Fungal Biol. 2016, 120, 1458–1467. [Google Scholar] [CrossRef] [PubMed]
- Gunde-Cimerman, N.; Turk, M.; Abramovi, Z.; Plemenitas, A. Salt stress and plasma-membrane fluidity in selected extremophilic yeasts and yeast-like fungi. FEMS Yeast Res. 2007, 7, 550–557. [Google Scholar]
- Turk, M.; Mejanelle, L.; Sentjur, M.; Gunde-Cimerman, N.; Grimalt, J.; Plemenitas, A. Salt-induced changes in lipid composition and membrane fluidity of halophilic yeast-like melanized fungi. Extremophiles 2004, 8, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Gow, N.A.R.; Latge, J.; Munro, C.A. The Fungal Cell Wall: Structure, Biosynthesis, and Function. Microbiol Spectr. 2017, 5, 1–25. [Google Scholar]
- Arroyo, J.; Farka, V.; Sanz, A.B.; Cabib, E. Strengthening the fungal cell wall through chitin – glucan cross-links: Effects on morphogenesis and cell integrity. Cell. Microbiol. 2016, 18, 1239–1250. [Google Scholar] [CrossRef]
- Kang, X.; Kirui, A.; Muszyński, A.; Widanage, M.C.D.; Chen, A.; Azadi, P.; Wang, P.; Mentink-Vigier, F.; Wang, T. Molecular architecture of fungal cell walls revealed by solid-state NMR. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Gastebois, A.; Clavaud, C.; Aimanianda, V.; Latgé, J.-P. Aspergillus fumigatus: Cell wall polysaccharides, their biosynthesis and organization. Future Microbiol. 2009, 4, 583–595. [Google Scholar] [CrossRef]
- Mellado, E.; Dubreucq, G.; Mol, P.; Sarfati, J.; Paris, S.; Diaquin, M.; Holden, D.W.; Latg, J.P. Cell wall biogenesis in a double chitin synthase mutant (chsG-/chsE-) of Aspergillus fumigatus. Fungal Genet. Biol. 2003, 38, 98–109. [Google Scholar] [CrossRef]
- Mouyna, I.; Hartl, L.; Latgé, J. β -1, 3-glucan modifying enzymes in Aspergillus fumigatus. Front. Microbiol. 2013, 4, 1–9. [Google Scholar] [CrossRef]
- Beauvais, A.; Latge, J. Functional duality of the cell wall. Curr. Opin. Microbiol. 2014, 11, 11–117. [Google Scholar]
- Chabane, S.; Sarfati, J.; Ibrahim-granet, O.; Du, C.; Schmidt, C.; Mouyna, I.; Prevost, M.; Calderone, R.; Latge, J. Glycosylphosphatidylinositol-Anchored Ecm33p Influences Conidial Cell Wall Biosynthesis in Aspergillus fumigatus. Appl. Environ. Microbiol. 2006, 72, 3259–3267. [Google Scholar] [CrossRef] [PubMed]
- Gil-Bona, A.; Reales-Calderon, J.A.; Parra-Giraldo, C.M.; Martinez-Lopez, R.; Monteoliva, L.; Gil, C. The Cell Wall Protein Ecm33 of Candida albicans is Involved in Chronological Life Span, Morphogenesis, Cell Wall Regeneration, Stress Tolerance, and Host – Cell Interaction. Front. Microbiol. 2016, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhu, J.; Ying, S.-H.; Feng, M.-G. The GPI-anchored protein Ecm33 is vital for conidiation, cell wall integrity, and multi-stress tolerance of two filamentous entomopathogens but not for virulence. Appl. Microbiol. Biotechnol. 2014, 98, 5517–5529. [Google Scholar] [CrossRef] [PubMed]
- Cabib, E. Two Novel Techniques for Determination of Polysaccharide Cross-Links Show that Crh1p and Crh2p Attach Chitin to both b(1-6)- and b(1-3)Glucan in the Saccharomyces cerevisiae Cell Wall. Eukaryot. Cell 2009, 8, 1626–1636. [Google Scholar] [CrossRef] [PubMed]
- Ene, I.V.; Walker, L.A.; Schiavone, M.; Lee, K.K.; Martin-yken, H.; Dague, E. Cell Wall Remodeling Enzymes Modulate Fungal Cell Wall Elasticity and Osmotic Stress Resistance. MBio 2015, 6, 1–15. [Google Scholar] [CrossRef]
- Ao, J.; Free, S.J. Genetic and biochemical characterization of the GH72 family of cell wall transglycosylases in Neurospora crassa. Fungal Genet. Biol. 2017, 101, 46–54. [Google Scholar] [CrossRef]
- Gastebois, A.; Fontaine, T.; Latgé, J.P.; Mouyna, I. b(1-3) glucanosyltransferase Gel4p is essential for Aspergillus fumigatus. Eukaryot. Cell 2010, 9, 1294–1298. [Google Scholar] [CrossRef]
- Mouyna, I.; Morelle, W.; Vai, M.; Monod, M.; Léchenne, B.; Fontaine, T.; Beauvais, A.; Sarfati, J.; Prévost, M.C.; Henry, C.; et al. Deletion of GEL2 encoding for a β(1-3)glucanosyltransferase affects morphogenesis and virulence in Aspergillus fumigatus. Mol. Microbiol. 2005, 56, 1675–1688. [Google Scholar] [CrossRef]
- Valdivieso, M.-H.; Ferrario, L.; Vai, M.; Duran, A.; Popolo, L. Chitin Synthesis in a gas1 Mutant of Saccharomyces cerevisiae. J. Bacteriol. 2000, 182, 4752–4757. [Google Scholar] [CrossRef]
- Wösten, H.A.B. Hydrophobins: Multipurpose Proteins. Annu. Rev. Microbiol. 2001, 55, 625–646. [Google Scholar] [CrossRef]
- Sunde, M.; Kwan, A.H.Y.; Templeton, M.D.; Beever, R.E.; Mackay, J.P. Structural analysis of hydrophobins. Micron 2008, 39, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Zajc, J.; Liu, Y.; Dai, W.; Yang, Z.; Hu, J.; Gostinčar, C.; Gunde-Cimerman, N. Genome and transcriptome sequencing of the halophilic fungus Wallemia ichthyophaga: Haloadaptations present and absent. BMC Genomics 2013, 14, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Plemenitaš, A.; Lenassi, M.; Konte, T.; Kejžar, A.; Zajc, J.; Gostinčar, C.; Gunde-Cimerman, N. Adaptation to high salt concentrations in halotolerant/halophilic fungi: A molecular perspective. Front. Microbiol. 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Littlejohn, K.A.; Hooley, P.; Cox, P.W. Bioinformatics predicts diverse Aspergillus hydrophobins with novel properties. Food Hydrocoll. 2012, 27, 503–516. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Baker, S.; Gamauf, C.; Kenerley, C.M.; Druzhinina, I.S. Purifying selection and birth-and-death evolution in the class II hydrophobin gene families of the ascomycete Trichoderma/Hypocrea. BMC Evol. Biol. 2008, 8, 1–16. [Google Scholar] [CrossRef]
- Krylov, D.M.; Wolf, Y.I.; Rogozin, I.B.; Koonin, E. V Gene Loss, Protein Sequence Divergence, Gene Dispensability, Expression Level, and Interactivity Are Correlated in Eukaryotic Evolution. Genome Res. 2003, 13, 2229–2235. [Google Scholar] [CrossRef]
- Przylucka, A.; Akcapinar, G.B.; Chenthamara, K.; Cai, F.; Grujic, M.; Karpenko, J.; Livoi, M.; Shen, Q.; Kubicek, C.P.; Druzhinina, I.S. HFB7—A novel orphan hydrophobin of the Harzianum and Virens clades of Trichoderma, is involved in response to biotic and abiotic stresses. Fungal Genet. Biol. 2017, 102, 63–76. [Google Scholar] [CrossRef]
- Appels, F.V.W.; Dijksterhuis, J.; Lukasiewicz, C.E.; Jansen, K.M.B.; Wösten, H.A.B.; Krijgsheld, P. Hydrophobin gene deletion and environmental growth conditions impact mechanical properties of mycelium by affecting the density of the material. Sci. Rep. 2018, 8, 1–7. [Google Scholar] [CrossRef]
- Zajc, J.; Kogej, T.; Galinski, E.A.; Gunde-Cimerman, N. Osmoadaptation Strategy of the Most Halophilic Fungus, Wallemia ichthyophaga, Growing Optimally at Salinities above 15% NaCl. Appl. Environ. Microbiol. 2014, 80, 247–256. [Google Scholar] [CrossRef]
- Witteveen, C.F.B.; Visser, J. Polyol pools in Aspergillus niger. FEMS Microbiol. Lett. 1995, 134, 57–62. [Google Scholar] [CrossRef]
- Krijgsheld, P.; Bleichrodt, R.; Van Veluw, G.J.; Wang, F.; Müller, W.H.; Dijksterhuis, J.; Wösten, H.A.B. Development in Aspergillus. Stud. Mycol. 2011, 74, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Ruijter, G.J.G.; Visser, J.; Rinzema, A. Polyol accumulation by Aspergillus oryzae at low water activity in solid-state fermentation. Microbiology 2004, 150, 1095–1101. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, M.R.; Krijgsheld, P.; Bleichrodt, R.; Menke, H.; Stam, H.; Stark, J.; Wösten, H.A.B.; Dijksterhuis, J. Germination of conidia of Aspergillus niger is accompanied by major changes in RNA profiles. Stud. Mycol. 2011, 74, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Fillinger, S.; Chaveroche, M.K.; van Dijck, P.; de Vries, R.; Ruijter, G.; Thevelein, J.; d’Enfert, C. Trehalose is required for the acquisition of tolerance to a variety of stresses in the filamentous fungus Aspergillus nidulans. Microbiology 2001, 147, 1851–1862. [Google Scholar] [CrossRef]
- Gorjan, A.; Gunde-Cimerman, N.; Plemenitas, A.; Gadd, G.M. Adaptation of the glycerol-3-phosphate dehydrogenase Gpd1 to high salinities in the extremely halotolerant Hortaea werneckii and halophilic Wallemia ichthyophaga. Fungal Biol. 2011, 115, 959–970. [Google Scholar]
- Turk, M.; Gostinčar, C. Glycerol metabolism genes in Aureobasidium pullulans and Aureobasidium subglaciale. Fungal Biol. 2018, 122, 63–73. [Google Scholar] [CrossRef]
- Clotet, J.; Posas, F. Control of Cell Cycle in Response to Osmostress: Lessons from Yeast. In Methods in Enzymology; Elsevier Inc.: Amsterdam NX, The Netherlands, 2017; Volume 428, pp. 63–76. [Google Scholar]
- Nadal, E.; Posas, F. EMBO Member ’ s Review Multilayered control of gene expression by stress-activated protein kinases. EMBO J. 2010, 29, 4–13. [Google Scholar] [CrossRef]
- Ramos, J.; Sychrová, H.; Kschischo, M. Yeast Membrane Transport; Ramos, J., Sychrová, H., Kschischo, M., Eds.; Springer International Publishing: AG, Switzerland, 2016; Volume 892, ISBN 9783319253022. [Google Scholar]
- Ramos, J.; Ariño, J.; Sychrova, H. Alkali-metal-cation influx and efflux systems in nonconventional yeast species. FEMS Microbiol. Lett. 2011, 317, 1–8. [Google Scholar] [CrossRef]
- Benito, B.; Garciadeblas, B.; Perez-Martin, J.; Rodriguez-Navarro, A. Growth at High pH and Sodium and Potassium Tolerance in Media above the Cytoplasmic pH Depend on ENA ATPases in Ustilago maydis. Eukaryot. Cell 2009, 8, 821–829. [Google Scholar] [CrossRef]
- Ramírez, J.; Ramírez, O.; Saldaña, C.; Coria, R.; Peña, A. A Saccharomyces cerevisiae Mutant Lacking a K+/H+ Exchanger. J. Bacteriol. 1998, 180, 5860–5865. [Google Scholar] [CrossRef]
- Maresova, L.; Sychrova, H. Physiological characterization of Saccharomyces cerevisiae kha1 deletion mutants. Mol. Microbiol. 2005, 55, 588–600. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence Properties a | Class | Cysteine Spacing | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Length | pI | %Basic | %Acidic | C1-C2 | C3-C4 | C4-C5 | C5-C6 | C7-C8 | ||
| sih1 | 129 | 3.98 | 5.4 | 10.9 | Unclassified | 6 | 35 | 19 | 5 | 19 |
| sih2 | 137 | 4.05 | 3.7 | 7.3 | Class I | 7 | 39 | 18 | 5 | 17 |
| sih3 | 110 | 4.02 | 6.4 | 12.7 | Unclassified | 5 | 32 | 6 | 5 | 13 |
| sih4 | 117 | 3.99 | 7.7 | 17.1 | Unclassified | 6 | 30 | 25 | 8 | 4 |
| Transporter Family | Pfam Number | A. sydowii | A. versicolor | A. nidulans | A. niger | A. fumigatus | W. ichthyophaga | H. werneckii |
|---|---|---|---|---|---|---|---|---|
| Alkali metal cation/H+ antiporter | PF08619 | 1 | 1 | 1 | 1 | 1 | 0 | 2 |
| Sodium: solute symporter | PF00474 | 5 | 7 | 4 | 4 | 3 | 2 | 6 |
| MFS | PF07690 PF00083 | 447 171 | 452 164 | 271 106 | 335 96 | 220 83 | 52 17 | 342 178 |
| Cation transporter/ATPase | PF00690 PF00689 | 21 16 | 16 12 | 13 11 | 13 7 | 12 11 | 7 4 | 14 9 |
| ABC Transporters | PF01061 PF00664 PF00005 PF06422 | 21 22 52 16 | 22 31 75 20 | 18 24 49 14 | 21 32 62 16 | 15 23 52 12 | 3 10 20 2 | 14 32 8 10 |
| AA uptake | PF13520 PF01490 PF00324 | 0 32 86 | 0 31 87 | 0 13 52 | 0 22 61 | 0 16 46 | 0 9 5 | 46 33 27 |
| V-Type ATPase-I | PF01496 | 1 | 1 | 2 | 1 | 1 | 1 | 2 |
| Na+/H+ exchanger family | PF00999 | 8 | 10 | 8 | 7 | 7 | 6 | 14 |
| Na+/Ca2+ exchanger family | PF01699 | 8 | 7 | 6 | 9 | 7 | 3 | 12 |
| Cation transporter family | PF02386 | 6 | 3 | 3 | 4 | 3 | 1 | 8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Llano, Y.; Rodríguez-Pupo, E.C.; Druzhinina, I.S.; Chenthamara, K.; Cai, F.; Gunde-Cimerman, N.; Zalar, P.; Gostinčar, C.; Kostanjšek, R.; Folch-Mallol, J.L.; et al. Stress Reshapes the Physiological Response of Halophile Fungi to Salinity. Cells 2020, 9, 525. https://doi.org/10.3390/cells9030525
Pérez-Llano Y, Rodríguez-Pupo EC, Druzhinina IS, Chenthamara K, Cai F, Gunde-Cimerman N, Zalar P, Gostinčar C, Kostanjšek R, Folch-Mallol JL, et al. Stress Reshapes the Physiological Response of Halophile Fungi to Salinity. Cells. 2020; 9(3):525. https://doi.org/10.3390/cells9030525
Chicago/Turabian StylePérez-Llano, Yordanis, Eya Caridad Rodríguez-Pupo, Irina S. Druzhinina, Komal Chenthamara, Feng Cai, Nina Gunde-Cimerman, Polona Zalar, Cene Gostinčar, Rok Kostanjšek, Jorge Luis Folch-Mallol, and et al. 2020. "Stress Reshapes the Physiological Response of Halophile Fungi to Salinity" Cells 9, no. 3: 525. https://doi.org/10.3390/cells9030525
APA StylePérez-Llano, Y., Rodríguez-Pupo, E. C., Druzhinina, I. S., Chenthamara, K., Cai, F., Gunde-Cimerman, N., Zalar, P., Gostinčar, C., Kostanjšek, R., Folch-Mallol, J. L., Batista-García, R. A., & Sánchez-Carbente, M. d. R. (2020). Stress Reshapes the Physiological Response of Halophile Fungi to Salinity. Cells, 9(3), 525. https://doi.org/10.3390/cells9030525