Abstract
The extracellular matrix (ECM) is a dynamic and highly organized tissue structure, providing support and maintaining normal epithelial architecture. In the last decade, increasing evidence has emerged demonstrating that alterations in ECM composition and assembly strongly affect cellular function and behavior. Even though the detailed mechanisms underlying cell-ECM crosstalk are yet to unravel, it is well established that ECM deregulation accompanies the development of many pathological conditions, such as gastric cancer. Notably, gastric cancer remains a worldwide concern, representing the third most frequent cause of cancer-associated deaths. Despite increased surveillance protocols, patients are usually diagnosed at advanced disease stages, urging the identification of novel diagnostic biomarkers and efficient therapeutic strategies. In this review, we provide a comprehensive overview regarding expression patterns of ECM components and cognate receptors described in normal gastric epithelium, pre-malignant lesions, and gastric carcinomas. Important insights are also discussed for the use of ECM-associated molecules as predictive biomarkers of the disease or as potential targets in gastric cancer.
1. Introduction
The extracellular matrix (ECM) is a complex assembly of fibrous proteins, proteoglycans and other molecules, namely cytokines, growth factors and hormones, whose precise composition varies from tissue to tissue [1]. A tight ECM regulation provides for proper architecture, as well as essential cues for mechanosensing and signaling during tissue development and maintenance [1,2]. Indeed, cellular processes such as growth, differentiation, survival, and morphogenesis are highly dependent on the cell-ECM interplay [3,4]. The reciprocal relationship between cells and the ECM is mainly mediated by cell receptors for ECM components, the so-called integrins.
Integrins work as bi-directional molecules that transduce physical and biochemical inputs from the surrounding microenvironment to the cell and vice-versa: transforming intracellular signals into different interactions with their ECM ligands [5,6]. Abnormal ECM and integrin profiles are frequently reported in cancer corroborating the functional relevance and the specificity of both ECM and integrins. Several studies have provided evidence that the ECM contributes to cancer pathogenesis by (i) stimulating integrin-dependent signaling that promotes invasion and proliferation; (ii) promoting an advantageous microenvironmental niche for metastatic cells; (iii) serving as a reservoir of growth factor and cytokines; (iv) interfering with the communication between cancer and immune cells; and (v) forming a physical barrier to anti-cancer agents (Figure 1) [4,6].
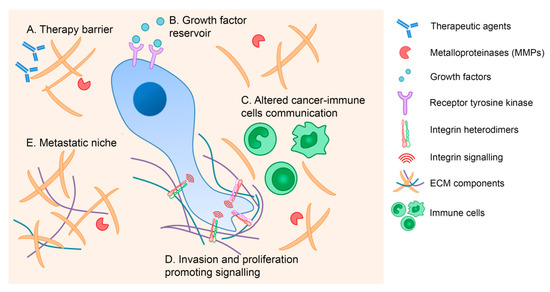
Figure 1.
The extracellular matrix (ECM) contribution to cancer pathogenesis. The ECM mediates cancer development through several mechanisms, including formation of a physical barrier to anti-cancer drugs (A), provision of growth factor and cytokines reserves (B), alteration of immune cell responses (C), stimulation of integrin-dependent signaling that promotes invasion and proliferation (D), and establishment of an advantageous niche for metastatic cells (E).
In gastric cancer, the role of the ECM has been undeniably demonstrated in all steps of the disease, from initiation to metastases. For instance, increased expression of tenascin has been detected in pre-malignant and malignant gastric epithelia [7], whereas collagens have been shown to be deregulated in more advanced stages [8,9,10]. Notably, a subset of collagen genes have been suggested as powerful independent prognostic markers and are able to distinguish pre-malignant from malignant lesions [8,9,10]. It is thus foreseen that ECM components and interactors hold great clinical potential as prognostic biomarkers and pharmacological targets in gastric cancer.
This review comprises a comprehensive analysis of the main studies concerning ECM remodeling and integrin alterations during gastric cancer development. In particular, we highlight the current understanding of key ECM components and adhesion molecules that mediate aberrant cell-ECM crosstalk and postulate their future application in innovative gastric cancer therapies.
2. The Biological Relevance of the ECM in Normal Gastric Tissue
The ECM is a complex three-dimensional network, ubiquitously present in the non-cellular compartment of tissues, providing structural support and maintaining normal tissue architecture, whilst modulating intercellular crosstalk [4,11,12]. As part of every cell’s microenvironment, the ECM is produced and released by local cells such as tissue-specific cells, fibroblasts, and immune cells [1,13,14].
Structurally, the ECM is an organized scaffold assembling a vast number of different macromolecules, namely collagen type-I, -II, -III, -V, and -XI, fibronectin, laminin, vitronectin, elastin, and growth factors, cytokines, and matrix metalloproteinases (MMPs), for which the ECM acts as a reservoir controlling their distribution and availability [12,15]. A complete list of ECM components, their structure, and function are reviewed elsewhere [1,15,16]. Additionally, there are two specialized forms of ECM, the basement membrane, which links the cells to the interstitial matrix, and the interstitial matrix itself, a scaffold in the form of a hydrated gel [11].
It is now well established that the type of components, their proportions, and their assembly determine the rigidity, porosity, and other properties that characterize each tissue [17]. It is thus expected that alterations in ECM composition and assembly, and subsequently on its mechanical and biochemical properties, will strongly affect cellular communication, function, and behavior [4,11,12].
Over the last few years, increasing evidence has emerged on the key role of the ECM in mediating distinct cellular processes including cellular adhesion, polarity, migration, differentiation, proliferation, and survival [14]. Cells are able to sense ECM signals mainly through integrins, a family of cell heterodimeric transmembrane proteins, known to modulate the cytoskeleton and transduce mechanical forces into biochemical events, thus inducing the activation of particular signaling cascades [16,18]. Conversely, cells can also remodel the ECM through the synthesis of new matrix components, alteration of their assembly, or production of enzymes that disrupt the ECM in response to intra- or extracellular stimuli as those occurring during cancer initiation and progression [19,20].
In epithelial tissues, as in gastric epithelia, the basal surface of cells is surrounded by the basement membrane, which is mainly composed of laminins, collagen type-IV, entactin, nidogen, proteoglycans (e.g., perlecan and agrin), and other glycoproteins [11,21,22]. This structure separates the epithelium from the surrounding stroma and provides support and cues for epithelial cell orientation which will help in the establishment and maintenance of the apicobasal polarity [4,23]. While some components are ubiquitously present in the basement membrane scaffold, a wide range of other constituents are unique, depending on the tissue function and origin [24]. In contrast, the interstitial matrix consists mainly of macromolecules such as fibrillar collagens (collagen type-I), fibronectin and proteoglycans, serving a wide variety of purposes, for instance buffering, hydration, binding, and force-resistance of the tissue. [11]. In the human gastric mucosa, aside from the above mentioned common constituents, a specific spatiotemporal distribution of ECM components is known to exist and to differ from that of other epithelial tissues [13]. In the early stages of gastric tissue formation, in which the gastric epithelium is mainly composed of undifferentiated cells, the glycoprotein tenascin was shown to be co-expressed with fibronectin [25]. This expression profile suggests a possible differential role of these ECM components during the morphogenesis of human gastric mucosa [25]. In contrast, other ECM components, for instance, basement membrane heparan sulfate proteoglycan (HSPG) core protein, collagen type-IV, and laminin α1, β1, and γ1 chains were systematically and uniformly detected [25]. More recently, tenascin-X was shown to be critical for normal gastric function as tenascin-X-deficient patients display upper gastric dysfunction and significantly greater symptoms of reflux, indigestion, and abdominal pain [26].
In addition, it was demonstrated that distinct chains of collagen type-IV are differentially distributed in human gastrointestinal tissues [27]. Briefly, collagen, the most abundant ECM fibrous protein, is formed by polypeptide α-chains that assemble into homo- or hetero-trimers and to date up to 28 different types of collagen have been described [28]. Whereas α1(IV), α2(IV), α5(IV), and α6(IV) collagen chains were found in the subepithelial basement membrane of all tissues, the α3(IV) and α4(IV) chains were restricted to specific regions of the gastric and intestinal epithelium, directly facing the lumen of the gastrointestinal tract [27]. This was suggestive of a protective effect against chemical and physical stress, given that α3(IV) and α4(IV) are considered to form physically stronger networks than other collagen type-IV molecules [27,29].
Laminin, another major component of the epithelial basement membrane, is a heterotrimeric protein containing α-, β-, and γ-chains [30]. In human gastric mucosa, laminin chains were shown to have differential expression, with laminin α1 chain found at the basement membrane of both surface and glandular epithelia, while laminin α2 and α3 chains were grossly mutually exclusive. Specifically, α2 chains were mostly detected in the glandular basement membrane and α3 at basement membranes underneath the surface epithelium [31]. The composition of the surface epithelial basement membrane was associated with rapid recovery capacity of the gastric surface epithelium following chemical injury, but also suggested specific programs towards surface or glandular cell differentiation [31].
Another ubiquitous component of the ECM is fibronectin, a dimeric glycoprotein involved in normal cell adhesion and growth, also shown to be central for tissue development and wound healing [32]. Fibronectin modulates the organization of interstitial ECM and is key for cell-ECM interaction as a ligand for many molecules, among which are other ECM components and integrins [11,33]. Supporting its involvement in epithelial healing, fibronectin was found to be markedly increased in the submucosa of healing gastric ulcers when compared to normal controls [34].
Overall, although many aspects of the ECM are still to unravel, it is well established that a tightly controlled ECM composition and cell-ECM interactions are required for normal gastric epithelial function. Deregulation of the ECM will thus result in the development of many pathological conditions, as is the case of gastric cancer.
3. ECM Deregulation in Pre-Malignant Lesions and Gastric Cancer
In cancer, transformed cells are able to leave the tissue of origin, invade and home to metastatic niches—a process in which the ECM plays a fundamental role [35]. It is known that, in parallel to alterations in cancer cells, ECM remodeling enzymes (including MMPs) increase their levels; cancer-associated fibroblasts (CAFs), immune cells, and other stromal cells are recruited; various growth factors are secreted, and collagen deposition is induced, all contributing to ECM remodeling at the tumor site [36,37]. These series of events lead to increased ECM stiffness, abnormal cell-cell adhesion, up-regulation of integrin signaling and subsequent activation of downstream cascades, promoting tumor growth and progression [17,36,37].
In addition to recognized cancer-associated ECM alterations, pre-malignant lesions already display ECM deregulation, with major implications for prognosis purposes and therapeutic strategies. In the particular case of human gastric epithelia, it is possible to distinguish two main histological types of gastric adenocarcinoma: the diffuse and the intestinal-type [38]. In the diffuse-type gastric cancer, no pre-malignant lesion is known and, microscopically, it appears to lack glandular structures, consisting of isolated or small groups of poorly cohesive cells [39,40,41]. These cells diffusely infiltrate the gastric wall leading to its widespread thickening and rigidity, known as linitis plastic [42]. In the distinct intestinal-type, a cascade of precancerous lesions—known as the Correa cascade—precedes gastric cancer [43,44]. In this histological type, normal gastric epithelia can evolve into non-atrophic chronic gastritis with active chronic inflammation and advance to multifocal atrophic gastritis, followed by intestinal metaplasia and dysplasia, culminating in gastric adenocarcinoma [43,44]. In the context of pre-malignant lesions and cancer development, tenascin was shown to be differentially expressed in inflammatory, dysplastic and neoplastic lesions of the human stomach (Table 1) [7]. A slight increase of tenascin was observed in superficial inflammation and early cancer, whereas a marked increase was detected in ulcers and invasive tumors of both diffuse and intestinal types, implicating tenascin in malignant growth and lesions undergoing repair and remodeling [7]. Furthermore, a subset of collagen genes was found to be differentially expressed in lesions of the human stomach and able to distinguish malignant from pre-malignant lesions, pinpointing these genes, in particular, COL11A1 and COL1A1, as biomarkers for early detection of gastric cancer [10]. Of relevance, MMP proteins, which belong to a family of zinc-dependent proteolytic enzymes, are important for ECM degradation thus contributing to the disruption of the basement membrane [45]. Accordingly, an increase in MMP-2 and MMP-9 production was observed in the gastric mucosa of patients with Helicobacter pylori-associated gastritis when compared with that of uninfected individuals, which indicates that MMP activity is likely to contribute to tissue damage during this process [46]. Subsequent studies have shown that gastric cancer cells infected by Helicobacter pylori increase the activity of MMP-2, MMP-9, and MMP-10 through c-Met- and EGFR-dependent signaling pathways, inducing ECM remodeling and cell invasion [47,48].

Table 1.
ECM components deregulated in gastric cancer. ECM proteins displaying abnormal expression patterns in gastric cancer and associated clinical observations.
Further corroborating their involvement in gastric carcinogenesis, altered levels of distinct MMPs have been widely reported in gastric cancer. For instance, MMP-9 expression was markedly higher in gastric carcinoma tissues than in adjacent healthy tissues, and associated with the depth of cancer invasion, suggesting that MMP-9 may serve as a novel biomarker in the diagnosis and prognosis of gastric carcinoma [53]. Moreover, MMP-9 levels were found to be significantly higher in the serum of gastric cancer patients when compared with those of controls [54]. Evaluation of urine samples from individuals with gastric cancer versus healthy controls also revealed that urinary MMP-9/NGAL complex was a potential biomarker of early-stage gastric cancer [57]. In addition, a number of meta-analyses found that overexpression of MMPs was associated with poor prognosis in gastric cancer patients, as was the case for MMP-9 and MMP-2 [50,51,56]. Interestingly, expression levels of both proteins were significantly higher in intestinal-type gastric cancer than in the diffuse-type [49]. MMP-9 expression, along with COX-2 and VEGF, were also increased in gastrointestinal stromal tumors (GIST), the most common mesenchymal neoplasms of the gastrointestinal tract [55]. Likewise, aggressive tumor phenotype and shorter overall survival in gastric cancer patients have been associated with higher MMP-7 expression [52].
Other ECM components are similarly relevant in gastric cancer development, as demonstrated by a large number of studies. Guszczyn and Sobolewski reported the enhancement of collagen turnover in gastric cancer tissues, which could contribute to disorganization of the ECM [59]. Several collagen genes were found to be overexpressed in gastric cancer and, among these, COL1A1 and COL4A1 were closely associated with overall survival of gastric cancer patients and could be regarded as risk factors for poor prognosis [9]. The expression of COL12A1 was also found upregulated in gastric cancer and positively correlated with tumor invasiveness, metastasis, and advanced clinical stage [8]. In tumor tissues of gastric cancer patients, levels of collagen type-I and -IV, fibronectin, and laminin were markedly higher than those detected in the normal tissues [60].
Furthermore, others and our group have demonstrated the importance of laminin γ2, which is a major component of epithelial basement membranes, in gastric cancer progression. Specifically, Wnt5a was shown to upregulate laminin γ2 promoting gastric cancer cell aggressiveness [61]. Moreover, our group has demonstrated that gastric cancer cells with E-cadherin dysfunction depend on laminin γ2 to survive and invade. We postulate that laminin γ2 upregulation may constitute an adaptive stimulus to allow cells to escape anoikis and invade adjacent tissues, contributing to cancer progression [62].
Deregulation of additional ECM components, namely glycoproteins and proteins of the basement membrane, has also been investigated. Lumican, an ECM proteoglycan, was found to be highly expressed in human gastric CAFs and its expression positively associated with depth of invasion, lymph node metastasis, TNM stage, and poor survival rate of gastric cancer patients [63]. Fibulin 1, which belongs to a family of extracellular glycoproteins, is a structural component of the basement membrane able to interact with other ECM components [71]. Importantly, fibulin 1 was shown to be downregulated through promoter hypermethylation in human gastric carcinoma tissues [64]. The levels of Nidogen-2 were reported to be significantly increased in gastric cancer tissues in comparison with normal controls and positively associated with TNM stage and poor prognosis of gastric cancer patients [65]. Connective tissue growth factor (CTGF) is another matrisome glycoprotein with high expression in tumor tissues and found to be an independent predictor of poor prognosis in gastric cancer patients [66,72]. Periostin, a secretory protein that can alter the remodeling of the ECM, was found to be highly expressed in gastric tumors and to be positively associated with gastric cancer metastasis by promoting tumor metastasis and invasion [67,73]. Of relevance, periostin has been reported to maintain primary tumor growth, as well as to contribute to a “fertile soil” for colonization and proliferation of cancer cells in metastatic niches [73,74]. Two different types of ECM proteoglycans, versican, and decorin, were significantly increased in human gastric carcinoma samples when compared with human normal gastric mucosa specimens [68]. Expression of the proteoglycan biglycan was also described to correlate with aggressiveness and poor prognosis of gastric cancer [69]. Interestingly, a core matrisome gene signature, of nine upregulated ECM genes, was identified in patients with gastric, ovarian, lung, and colon cancers, and was able to predict clinical outcome in these patients [75].
Notably, some ECM molecules have been described to be differentially expressed in distinct types of gastric cancer. Indeed, higher expression levels of galectin-1 and thrombospondin were detected in diffuse gastric cancer in comparison with those found in perifocal and tumor zones of the intestinal-type, which could reflect the dissimilarities of the two histotypes [70]. The expression signature of the diffuse gastric cancer included genes encoding collagens, biglycan, osteoglycin, proteoglycan, MMPs, cadherin 11, Thy-1 SERPINS, and fibrillin, revealing active ECM production and remodeling, as well as signaling linked to regulation of cell proliferation [76]. A comparative study of gene expression profiles from diffuse and intestinal-type gastric cancers demonstrated that the signature of the diffuse-type cancer exhibited altered expression of genes related to ECM components, whereas that of the intestinal-type revealed distinct alterations in cell growth or cell cycle pathways [77,78].
In conclusion, disruption of the tightly orchestrated ECM organization will compromise gastric tissue structure and function, ultimately contributing to gastric cancer progression. In line with this, other key players including integrin receptors that mediate the cell’s interaction with the ECM are crucial determinants for carcinogenesis, as illustrated in Figure 2.
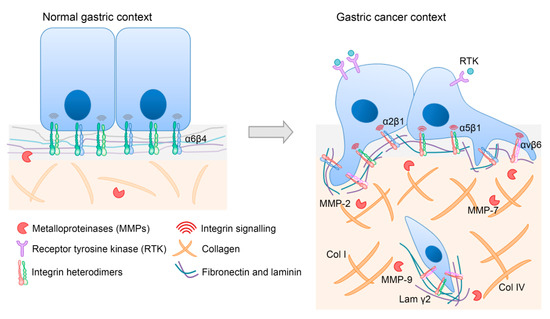
Figure 2.
Representative image of major ECM-integrin alterations in gastric cancer. ECM composition, as well as integrin expression and signaling, are distinct in normal and in gastric cancer contexts. Relevant players for gastric carcinogenesis are depicted to illustrate aberrant features. Briefly, during gastric cancer development, the expression of some integrin heterodimers is lost (α6β4), whereas that of others is increased (α2β1, α5β1, and αvβ6). Moreover, the increased activity of several MMPs (MMP-2, MMP-7, and MMP-9) is associated with cancer cell invasion. Increased levels of collagens (Col I and Col IV) and laminin γ2 are also part of a specific gastric cancer ECM signature.
4. Aberrant Expression of Integrins in Gastric Cancer
Integrins are cell-surface adhesion molecules that sense information within the ECM and translate such signals into cellular responses involving tissue-specific gene regulation [79]. Remarkably, intracellular signal inputs are also reflected in integrin activity and engagement to ECM ligands, highlighting a bidirectional function of integrins as mechanosensors and mechanotransducers [5].
During outside-in signaling, the engagement of integrins with ECM ligands induces a conformational change on the integrin’s cytoplasmic domain and promotes the assembly of macromolecules termed focal adhesions [80,81]. In contrast, molecular interactions with the integrin cytoplasmic domain lead to conformational changes, resulting in receptor activation and increased affinity to ECM ligands (inside-out signaling) [82,83,84].
Integrins are found as heterodimeric combinations of 18 α and 8 β subunits that interact in a restricted manner, generating 24 family members expressed in a cell- and tissue-specific manner [6]. A combination of both subunits determines integrin specificity for its corresponding ligand. Integrins can recognize Arg-Gly-Asp (RGD) peptide motifs or conformational structures encompassing different amino acid rearrangements [85]. More so, a panel of integrins bind to unique ECM ligands (for instance, α5β1 integrin to fibronectin), whereas others present a certain degree of functional redundancy and are able to engage multiple ligands, overlapping with different integrin heterodimers (such as αvβ3 integrin that binds laminin, collagen, fibronectin, and tenascin C) [86,87]. It has been speculated that this integrin redundancy is an adaptive mechanism to allow a prompt response to changes in the microenvironment [84].
Given the relevance of integrin-ECM interactions in determining cell fate, it is not surprising that deregulated integrin expression and activity is a precursor event in the pathogenesis of many human diseases [88]. Indeed, defects in platelet integrin αIIbβ3 (GPIIb-IIIa) can originate Glanzmann thrombasthenia [89], and the leukocyte adhesion deficiency (LAD) is a primary immunodeficiency disorder caused by a mutation on β2 integrin [90,91]. Additionally, a number of skin diseases are frequently associated with mutations on α2, α6 and β4 integrins [92].
Aberrant integrin expression has also been reported in cancer, where it is associated with progression and poor prognosis [3,6]. In vitro and in vivo studies have demonstrated that abnormal integrin levels award cancer cells with increased capabilities to survive and migrate in a hostile microenvironment [3,6]. Accordingly, over the past years, distinct integrins have emerged as prognostic biomarkers and potential therapeutic targets in the oncology field [3,93,94]. For instance, loss of β1 integrin expression precludes tumor progression in different tumor models, including breast cancer and pancreatic tumor β-cells [95,96]. In early-stage non-small cell lung cancer, increased expression of β1 or α5 integrins was found correlated with poor prognosis [97]. In fact, expression of αvβ3, αvβ5, α5β1, α6β4, α4β1, αvβ6, and αvβ8 integrin heterodimers have been shown to correlate with disease progression and poor patient outcome in a myriad of cancer types and, as such, those have become main targets of research and clinical studies [6].
Focusing on the context of gastric cancer, several integrin heterodimers have already been claimed to be involved in the etiology of the disease (Table 2). In particular, the predictive value of αvβ6 integrin has been highlighted, since its increased expression is recurrently associated with lymph node metastases in gastrointestinal cancers and reduced patient survival [98,99,100,101,102,103]. Zhao et al. reported that the invasiveness of gastric cancer cells expressing αvβ6 integrin seems to occur through ECM degradation in a process mediated by the pro-angiogenic growth factor VEGF and enhanced secretion of matrix metalloprotein-9 (MMP-9) [101]. Another study by Gu and colleagues demonstrated a potential link between cancer cell survival and αvβ6/MMP-9 signaling in colon cancer cells [104]. Additionally, increased αvβ6 expression has been correlated with the number of CAFs, awarding αvβ6 a prognostic value in human gastric cancer [102].
Table 2.
Integrins abnormally expressed in gastric cancer. Depiction of reported integrins with aberrant expression pattern in gastric cancer and their clinical relevance.
Integrins α2β1 and α3β1 have also been linked to the metastatic process in human gastric cancer, despite exerting separate functions in this process [105,106]. α2β1 was associated with the presence of lymph node and liver metastases, whereas α3β1 expression correlated with liver and peritoneal metastases [105]. Importantly, a multivariate analysis of both integrins in primary gastric cancer samples associates α3β1 expression with peritoneal metastasis formation and depth of invasion [105]. Later, up-regulation of integrin α2β1 was shown to be essential for peritoneal dissemination of gastric cancer promoted by the interaction with Cysteine-rich 61 [106]. Cysteine-rich 61 is an ECM protein regulating a broad range of cellular activities, including cell adhesion, migration, proliferation, cell survival and angiogenesis [109,110]. In breast cancer, Cysteine-rich 61 supports metastases and mitigates anoikis, which may explain its association with more advanced disease features [111,112].
The classical fibronectin receptor, α5β1 integrin, has also been described as abnormally expressed in gastric cancer [107]. A study involving 186 gastric cancer samples demonstrated that 68.3% of cases presented higher α5β1 integrin expression than that of paired normal mucosa [107]. α5β1 integrin levels were closely related to histological differentiation, lymph node metastases, and tumor recurrence, suggesting α5β1-integrin as a marker of poor prognosis [107]. Further, overall survival and disease-free survival of patients displaying high α5β1-integrin expression were significantly worse than those of patients with low or absent expression [107]. In accordance, it was verified that under controlled in vitro conditions, invasive cancer cells display high α5β1 integrin levels [113]. α5β1 generates higher contractile forces and increased cytoskeletal dynamics, which allow faster and persistent migration [113].
More recently, Boger et al. have evaluated αvβ3 and αvβ5 as prognostic, diagnostic and therapeutic targets in a large cohort of 482 gastric cancer cases [108]. The group observed more often a positive αvβ3 and αvβ5 status in the intestinal-type gastric cancer than in the diffuse-type and, thus proposed that both markers could be helpful in the histological classification of gastric cancer [108]. Moreover, αvβ5 was confirmed to be an independent prognostic factor of intestinal-type gastric cancer, given that patients with absence of αvβ5 on stroma cells had better disease outcome and significantly longer survival [108]. In an in vivo context, blockage of αvβ3 and αvβ5 integrins with monoclonal antibodies resulted in a drastic reduction of tumor growth and metastases through inhibition of focal adhesions and cell motility signals [114,115]. Notably, loss of integrin expression has also been described in gastric cancer. A study by Ishii et al. demonstrates that loss of α6β4 is a biomarker of peritoneal dissemination and poor prognosis of gastric cancer patients [116]. Integrins play an indisputable role in nearly every step of cancer progression, from initiation to metastasis [6]. In gastric cancer, it is also clear that altered integrins mediate an array of cellular effects that culminate in tumor progression. An outstanding challenge in this research field remains the understanding of the signaling following integrin-ECM gastric interactions that trigger tumor-promoting characteristics.
5. ECM-Integrin Signaling in Cancer
Every single cell type displays a specific integrin expression profile that changes in response to cellular or environmental inputs [117,118]. In normal cells, integrin activity is strictly regulated, whereas in cancer cells, abnormal integrin activity promotes the acquisition of oncogenic properties either through ECM remodeling or by interfering with intracellular signaling that may, for instance, lead to oncogene activation [6,117]. Tumor cells are able to switch integrin’s exposure and modify their downstream signaling in order to survive, proliferate and successfully colonize adjacent tissues [119]. In breast cancer, it has been reported that decreased levels of α2β1 and α3β1 integrins potentiate tumor cell dissemination [95,120,121]. In contrast, overexpression of α6β4 and αvβ3 integrins were shown to be correlated with metastasis formation and shorter patient survival [122,123,124]. The processes underlying integrin regulation and signaling in cancer are however complex and highly dependent on the tissue of origin, histological tumor type, and disease stage [119].
As explained in the previous section, integrins hold a unique ability to signal bidirectionally and can thus initiate a cascade of events upon either ligand engagement or intracellular interaction with specific moieties [125,126]. Integrin binding to ECM proteins induces integrin clustering and assembly of focal adhesion complexes at the plasma membrane. The tyrosine-phosphorylated protein Focal Adhesion Kinase (FAK) is a major component of focal adhesions that, upon recruitment, is autophosphorylated at Tyr397 residue, exposing a steroid receptor coactivator (Src) homology 2 (SH2) domain-binding site for Src [127,128]. Src then phosphorylates FAK at the additional Tyr576 and Tyr577 residues, amplifying its catalytic activity [92,129]. FAK phosphorylation mobilizes proteins that contain SH2 domains, such as the growth factor receptor-bound protein 2 (Grb2) and the phosphatidylinositol 3-kinase (PI3K), which in turn activate the Ras-mitogen-activated protein kinase (MAPK/ERK) and PI3K pathways [130,131]. In particular, binding of α5β1 to fibronectin, or binding of the epidermal growth factor (EGF) to its receptor (EGFR), triggers MAPK/ERK signaling that coordinates cell survival, proliferation, motility and metabolism [6,132,133,134]. The PI3K/AKT pathway is preferentially activated in response to αvβ3 integrin, which promotes survival of cancer cells by targeting the pro-apoptotic Bcl-2 related protein [135]. Under conditions of nutrient availability, activation of PI3K/AKT can also involve the mammalian target of rapamycin (mTOR) that controls cell growth and proliferation [136].
Additionally, activated FAK can bind and phosphorylate other molecules, namely paxillin and p130Cas (also known as BCAR1), allowing the subsequent recruitment of a panel of adaptor and signaling molecules [127,137]. Through recruitment and phosphorylation of p130Cas, activated FAK/Src complex stimulates Rac1 activity, a member of the Ras superfamily of GTP-binding proteins that induces protrusion formation and inhibits cytoskeletal contractility, thereby facilitating cell spreading [138].
Corroborating the involvement of integrin-mediated signaling in the genesis and development of gastric cancer, pTyr397 FAK phosphorylation was found to be abundant among patients with gastric carcinomas [139]. Patients with higher levels of pTyr397 FAK displayed increased recurrence following surgical resection and poor 5-year recurrence-free survival [139]. In vitro studies demonstrated that a dominant-negative mutant of FAK (impairing the Tyr397 FAK phosphorylation) reduced the ability of gastric cancer cells to migrate, invade, and proliferate when compared with cells overexpressing wild-type FAK [139]. The pathogen H. pylori has also been shown to activate FAK in gastric epithelial cells, leading to cell scattering and elongation [140]. Upon translocation of the bacterial factor cytotoxin-associated gene A (CagA), FAK activity is modulated by both cortactin and vinculin modifications, which deregulate cell-matrix adhesion [140,141]. Moreover, expression of p130Cas was mainly absent in normal gastric mucosa, whereas it was strongly or moderately positive in gastric carcinoma [142]. A similar tendency was observed for paxillin, which was aberrantly upregulated in gastric cancer tissues and cell lines [143,144]. In fact, Chen and collaborators evaluated a large series of 239 gastric cancer patients and established a direct correlation between paxillin expression and distant metastasis, as well as advanced tumor stage [143]. Protein modulation through overexpression and inhibition approaches revealed that paxillin is a key regulator of proliferation and migration of gastric cancer cells [143].
In contrast with the outside-in cascade of events, inside-out signaling initiates upon binding of integrin-activators like talins and kindlins (kindlin-1, kindlin-2, and kindlin-3) to the intracellular portion of β-integrins [92,145]. This interaction leads to an extended conformation of integrins and, consequently, to their increased affinity for ECM ligands [92,145]. Remarkably, kindlin-2 was upregulated both at RNA and protein levels in gastric cancer [146]. High kindlin-2 expression levels were associated with tumor stromal invasion, lymph node metastasis, and tumor staging, and were considered an independent risk factor of progression-free survival [146]. In this context, kindlin-2 seems to play a pro-invasive function through the activation of β1 and β3 integrins [147].
Aside from its function as an integrin activator, talin is also a critical mediator of mechanotransduction signals [148]. Along with filamin and α-actinin, talin is responsible for the connection between integrins and the actomyosin cytoskeleton [149]. This cytoskeletal bridge is crucial to orchestrate protein trafficking, cell morphology and a myriad of cellular functions, including survival and motility [14]. Unlike talin, kindlins alone are not sufficient to shift integrins to a high-affinity state, despite being required for proper talin function [150]. The mechanism through which kindlins cooperate with talin to support integrin activation remains unclear, although it has been proposed that kindlins recruit talin to integrin β tails, promoting integrin activation [151]. A different explanation is that kindlins and talin synergize in integrin activation and do not interfere with each other´s interaction with integrins [152]. Accordingly, kindlins may co-activate integrin through a mechanism independent of talin recruitment [152].
Despite the increased knowledge of the signaling cascades mediating cell-ECM interactions, there is still a lack of studies focusing on gastric cancer. In the near future, we expect to witness breakthrough research in this topic unraveling disease-associated mechanisms and, ultimately, fostering the emergence of novel therapeutic strategies targeting integrin signaling.
6. Potential Therapeutic Targets and Strategies
Several studies have shown that inhibition of integrin or its downstream effectors could block the major hallmarks of cancer [3,119]. Therefore, integrins and adaptor molecules have soon emerged as potential therapeutic targets for a number of cancer types, including glioblastoma, melanoma and breast cancer [115,153,154,155,156].
Based on integrin expression profiles, two therapeutic strategies have been developed. One involves direct inhibition of integrin function and the other aims at integrin-directed delivery of drugs, with the first concept being employed more often in the clinic, namely in ulcerative colitis, Crohn’s disease, and multiple sclerosis [3,157].
So far, no clinical trials of integrin-based therapies have been carried out for gastric cancer (ClinicalTrials.gov). This is probably due to scarce data regarding the integrin expression profile in gastric carcinoma patients and in normal gastric tissue.
Among the few ECM receptors described as abnormally expressed in gastric cancer, ανβ6 increased expression is associated with reduced survival and it has been suggested as a prognostic marker in early-stage disease [99,100]. As such, ανβ6 could be an attractive target for early intervention and treatment of gastric carcinoma, and to date, several antibodies and small molecules have been developed to inhibit this molecule. Abituzumab (DI17E6, EMD 525797: Merck KgaA)—a humanized monoclonal IgG2 antibody that targets αν heterodimers [158] – yielded clinical benefit in patients with early-stage metastatic colorectal cancer expressing high levels of ανβ6 [159]. Intetumumab (CNTO95) from Centocor is also a pan-αv integrin inhibitor with well-established anti-tumor and anti-angiogenic effects in a human melanoma xenograft model [114,115]. In Phase I trials, it exhibited low toxicity and good tolerance among patients affected by advanced melanoma and castration-resistant prostate cancer [160,161]. Still, the therapeutic effect of Intetumumab in both cancer contexts requires further investigation [160,161]. More recently, Biogen-Idec developed a monoclonal antibody specifically targeting ανβ6, which was shown to inhibit tumor growth in xenografts of human pharyngeal carcinoma cells through regulation of transforming growth factor-beta (TGF-β) [162]. This antibody, STX-100 (BG00011), is currently in clinical trials for treatment of idiopathic pulmonary fibrosis and for nephropathy [157]. Likewise, the first small molecule inhibitor of αvβ6 integrin, GSK3008348, was produced by GlaxoSmithKline Research as an inhaled compound for the treatment of idiopathic pulmonary fibrosis [163].
As described in previous sections, α2β1 integrin was found overexpressed in peritoneal metastases of gastric carcinoma and has been implicated in the dissemination of gastric cancer cells, both in patient samples and xenograft models [164,165]. Aside from its role as a regulator of cancer metastasis, α2β1 was described as a promoter of inflammation, angiogenesis, and chemoresistance [166]. Hence, several clinical programs have been implemented targeting this specific molecule. For instance, the monoclonal blocking antibody Vatelizumab (CHR-1103) was developed by Chromos Molecular systems and Glenmark Pharmaceuticals for the treatment of multiple sclerosis and ulcerative colitis [157,166]. The sulfonamide derivative small molecule E7820, which inhibits α2 gene expression, was used in combination with standard chemotherapy in advanced or refractory solid tumors, namely metastatic colorectal carcinoma [167,168].
α3β1 integrin is a receptor of laminin and its expression is correlated with the depth of gastric cancer invasion (into the muscularis propria or subserosa), as well as with the formation of peritoneal metastases [105]. Interestingly, an existing pan-specific anti-β1 antibody targeting laminin receptors was shown to exert a synergistic anti-tumor effect when combined with cisplatin [169].
Despite encouraging results from in vitro and preclinical studies, the success of integrin-targeted strategies in cancer has been limited [157]. In fact, the applicability of ECM receptors as therapeutic targets is highly dependent on the tumor type and on the disease stage given that the pattern of integrin expression varies between cancer types and during cancer progression [6]. Careful patient stratification and a deep understanding of basic mechanisms of integrin regulation are thus urgent to improve the anti-tumor efficacy of integrin therapies.
In addition to integrin-targeted strategies, ECM remodeling may also be beneficial for cancer treatment. Several cancer types exhibit abnormal accumulation/deposition of particular ECM components and increased ECM stiffness, which impair drug diffusion and, consequently, decreases treatment efficacy [4,170]. Moreover, an increased ECM density perturbs cell-cell adhesion, enhances cell-ECM interaction and increases the proliferation of gastric cancer cells [60]. Therefore, modulation of ECM-related enzymes such as collagenase, MMPs or lysyl oxidases can be a promising therapeutic strategy for gastric cancer. Collagenase disrupts collagen networks and stimulates anti-tumor immune surveillance by increasing the ability of T cells to interact with lung cancer cells [171]. MMP inhibitors, such as Marimastat, Batimastat, and Prinomastat, failed all trials in various cancer types due to the broad-spectrum of these drugs (acting simultaneously in anti- and protumorigenic MMPs) [172,173,174,175]. However, individual MMP-targeting approaches using monoclonal antibodies have been gaining attention and will certainly improve their tolerability and efficacy [176]. In contrast to collagenases and MMPs, which are ECM degrading enzymes, lysyl oxidases generate covalent cross-links between collagen fibers, inducing tissue stiffness and ECM resistance/stability [177,178]. The administration of a monoclonal antibody inhibiting lysyl oxidase-like 2 (LOXL2) reduced lung and liver fibrosis, as well as metastases in xenografted tumors [179]. Those effects were associated with a decrease in activated fibroblasts, reduced production of growth factors and cytokines, and inhibition of TGF-β signaling [179].
Previous studies have demonstrated that TGF-β is increased in gastric tumor tissue when compared with adjacent mucosa [180]. In addition, high TGF-β expression was correlated with worse overall survival of gastric cancer patients [180]. Given the importance of TGF-β in profibrotic activity and in the regulation of ECM synthesis, secretion, and processing [181,182], several studies were designed to evaluate the response of TGF-β inhibitory antibodies [183,184,185]. A specific monoclonal antibody, Fresolimumab (GC1008), was tested in advanced melanoma and renal cell carcinoma patients with acceptable safety and evidence of anti-tumor activity [183]. In systemic sclerosis, Fresolimumab was shown to decrease dermal myofibroblast infiltration and to reduce expression of fibrosis markers [184].
The angiogenic process is also known to involve interactions between endothelial cells and the ECM. Intensive research focusing on this critical interaction has unveiled anti-angiogenic factors with application in cancer therapeutics, namely small peptides derived from naturally occurring proteins. Endostatin is a small fragment of the ECM protein collagen type-XVIII identified as a potent inhibitor of angiogenesis [186]. In gastric cancer, patients displayed higher serum endostatin levels than those of healthy subjects, which were correlated with aggressiveness [187]. Recombinant human endostatin, Endostar, proved to be more effective than single chemotherapy in a plethora of cancers, including gastric cancer [188,189]. Additionally, various sites for angiogenesis have been identified on laminin-1, among which, C16Y was shown to be a potent antagonist to integrins during angiogenesis and has thus been suggested as a potential cancer therapeutic agent [190].
One different anti-angiogenic strategy encompasses therapies targeting the angiogenic VEGF-mediated pathway, which is considered critical not only for the regulation of tumor angiogenesis but also for the degradation and remodeling of the ECM [191]. Significant evidence awards VEGF/VEGFR2 signaling an important role in gastric cancer pathogenesis, and indeed, gastric cancer patients were reported to display significantly higher plasma or serum VEGF levels than healthy control subjects [192]. The monoclonal anti-VEGF antibody Bevacizumab, which was the first drug targeting the VEGF pathway, did not reach promising results in overall survival of gastric cancer patients in the AVAGAST clinical trial, however, it is now approved for first- and/or second-line treatment of a variety of tumors including colorectal cancer [191]. In contrast, the human monoclonal anti-VEGFR2 antibody Ramucirumab yielded significant survival benefits in patients with previously treated advanced gastric cancer or gastroesophageal junction carcinomas in a phase III clinical trial [193]. Accordingly, Ramucirumab is now used to treat gastric cancer patients with advanced or metastatic disease on or after first-line chemotherapy [191]. Currently, there are also a number of clinical trials addressing the inclusion of Apatinib, a highly selective VEGFR2 inhibitor, in gastric cancer treatment regimens, including phase III and IV trials [191,194].
It is clear from the extensive research and clinical trials that despite the enormous potential in modulating the ECM, the majority of agents elicit therapeutic responses in gastric cancer patients that are often too modest. Given that the ECM is actively remodeled, a major challenge in drug development is to identify predictive biomarkers and the correct therapy timing, as well as to design clinical trials with enriched populations, so gastric cancer patients can be offered treatments that will result in a significant increase in overall survival.
7. Conclusions
Increasing evidence has shown that a specific ECM signature is associated with each tissue and its functional features. ECM dynamics, composition, and structure are tightly regulated, and ECM remodeling has proven to have a major impact on cancer progression and prognosis. Experimental and clinical observations strongly indicate that ECM composition and cell exposure of specific integrins are accomplices in the precancerous cascade leading to gastric cancer, namely by promoting proliferation, survival, migration, invasion, and metastasis. Therefore, ECM constituents, receptors and associated signaling molecules should be explored as biomarkers of prognosis and/or therapeutic targets. Innovative and more effective gastric cancer treatments can be achieved by using combined strategies of ECM targeting with RTK inhibitors or immuno-oncology agents.
Author Contributions
J.F. and R.S. were responsible for the study concept and design. A.M.M., J.P., S.M., M.S.F., P.C., and J.F. have drafted the article. J.F., M.S.F., P.C., and R.S. critically reviewed the manuscript for important intellectual content. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by FEDER funds through the Operational Programme for Competitiveness Factors (COMPETE 2020), Programa Operacional de Competitividade e Internacionalização (POCI), Programa Operacional Regional do Norte (Norte 2020) and by National Funds through the Portuguese Foundation for Science and Technology (FCT), under the projects PTDC/MED-GEN/30356/2017, PTDC/BIM-ONC/0171/2012, PTDC/BIM-ONC/0281/2014, NORTE-01–0145-FEDER-000029, and doctoral grants SFRH/BD/114687/2016-AMM, SFRH/BD/143533/2019-JP, and SFRH/BD/108009/2015-SM. We acknowledge the American Association of Patients with Hereditary Gastric Cancer “No Stomach for Cancer” for funding Seruca and Figueiredo’s research.
Conflicts of Interest
The authors declare no competing or financial and non-financial interest.
References
- Mouw, J.K.; Ou, G.; Weaver, V.M. Extracellular matrix assembly: A multiscale deconstruction. Nat. Rev. Mol. Cell Biol. 2014, 15, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, J.D.; Dufresne, E.R.; Schwartz, M.A. Mechanotransduction and extracellular matrix homeostasis. Nat. Rev. Mol. Cell Biol. 2014, 15, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer 2010, 10, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.A. Integrins and extracellular matrix in mechanotransduction. Cold Spring Harb. Perspect. Biol. 2010, 2, a005066. [Google Scholar] [CrossRef] [PubMed]
- Hamidi, H.; Ivaska, J. Every step of the way: Integrins in cancer progression and metastasis. Nat. Rev. Cancer 2018, 18, 533–548. [Google Scholar] [CrossRef]
- Tiitta, O.; Sipponen, P.; Gould, V.; Virtanen, I. Tenascin expression in inflammatory, dysplastic and neoplastic lesions of the human stomach. Virchows Arch. 1994, 425, 369–374. [Google Scholar] [CrossRef]
- Jiang, X.; Wu, M.; Xu, X.; Zhang, L.; Huang, Y.; Xu, Z.; He, K.; Wang, H.; Wang, H.; Teng, L. COL12A1, a novel potential prognostic factor and therapeutic target in gastric cancer. Mol. Med. Rep. 2019, 20, 3103–3112. [Google Scholar] [CrossRef]
- Zhang, Q.N.; Zhu, H.L.; Xia, M.T.; Liao, J.; Huang, X.T.; Xiao, J.W.; Yuan, C. A panel of collagen genes are associated with prognosis of patients with gastric cancer and regulated by microRNA-29c-3p: An integrated bioinformatics analysis and experimental validation. Cancer Manag. Res. 2019, 11, 4757–4772. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhou, T.; Li, A.; Yao, H.; He, F.; Wang, L.; Si, J. A potential role of collagens expression in distinguishing between premalignant and malignant lesions in stomach. Anat. Rec. (Hoboken) 2009, 292, 692–700. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed]
- Rozario, T.; DeSimone, D.W. The extracellular matrix in development and morphogenesis: A dynamic view. Dev. Biol. 2010, 341, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Hussey, G.S.; Keane, T.J.; Badylak, S.F. The extracellular matrix of the gastrointestinal tract: A regenerative medicine platform. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Turnbull, J.; Guimond, S. Extracellular matrix and cell signalling: The dynamic cooperation of integrin, proteoglycan and growth factor receptor. J. Endocrinol. 2011, 209, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Kular, J.K.; Basu, S.; Sharma, R.I. The extracellular matrix: Structure, composition, age-related differences, tools for analysis and applications for tissue engineering. J. Tissue Eng. 2014, 5, 2041731414557112. [Google Scholar] [CrossRef] [PubMed]
- Jarvelainen, H.; Sainio, A.; Koulu, M.; Wight, T.N.; Penttinen, R. Extracellular matrix molecules: Potential targets in pharmacotherapy. Pharm. Rev. 2009, 61, 198–223. [Google Scholar] [CrossRef]
- Lu, P.; Weaver, V.M.; Werb, Z. The extracellular matrix: A dynamic niche in cancer progression. J. Cell Biol. 2012, 196, 395–406. [Google Scholar] [CrossRef]
- Kechagia, J.Z.; Ivaska, J.; Roca-Cusachs, P. Integrins as biomechanical sensors of the microenvironment. Nat. Rev. Mol. Cell Biol. 2019, 20, 457–473. [Google Scholar] [CrossRef]
- Butcher, D.T.; Alliston, T.; Weaver, V.M. A tense situation: Forcing tumour progression. Nat. Rev. Cancer 2009, 9, 108–122. [Google Scholar] [CrossRef]
- Eble, J.A.; Niland, S. The extracellular matrix in tumor progression and metastasis. Clin. Exp. Metastasis 2019, 36, 171–198. [Google Scholar] [CrossRef]
- Egeblad, M.; Nakasone, E.S.; Werb, Z. Tumors as organs: Complex tissues that interface with the entire organism. Dev. Cell 2010, 18, 884–901. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Boudreau, A.; Bissell, M.J. Tissue architecture and function: Dynamic reciprocity via extra- and intra-cellular matrices. Cancer Metastasis Rev. 2009, 28, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Cox, T.R.; Erler, J.T. Remodeling and homeostasis of the extracellular matrix: Implications for fibrotic diseases and cancer. Dis Model. Mech. 2011, 4, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Kruegel, J.; Miosge, N. Basement membrane components are key players in specialized extracellular matrices. Cell Mol. Life Sci. 2010, 67, 2879–2895. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, E.; Menard, D. Differential expression of extracellular matrix components during the morphogenesis of human gastric mucosa. Anat. Rec. 1996, 245, 668–676. [Google Scholar] [CrossRef]
- Aktar, R.; Peiris, M.; Fikree, A.; Eaton, S.; Kritas, S.; Kentish, S.J.; Araujo, E.J.A.; Bacarin, C.; Page, A.J.; Voermans, N.C.; et al. A novel role for the extracellular matrix glycoprotein-Tenascin-X in gastric function. J. Physiol. 2019, 597, 1503–1515. [Google Scholar] [CrossRef]
- Sato, H.; Naito, I.; Momota, R.; Naomoto, Y.; Yamatsuji, T.; Sado, Y.; Ninomiya, Y.; Ohtsuka, A. The differential distribution of type IV collagen alpha chains in the subepithelial basement membrane of the human alimentary canal. Arch. Histol. Cytol. 2007, 70, 313–323. [Google Scholar] [CrossRef]
- Ricard-Blum, S. The collagen family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef]
- Leinonen, A.; Mariyama, M.; Mochizuki, T.; Tryggvason, K.; Reeders, S.T. Complete primary structure of the human type IV collagen alpha 4(IV) chain. Comparison with structure and expression of the other alpha (IV) chains. J. Biol. Chem. 1994, 269, 26172–26177. [Google Scholar]
- Domogatskaya, A.; Rodin, S.; Tryggvason, K. Functional diversity of laminins. Annu Rev. Cell Dev. Biol. 2012, 28, 523–553. [Google Scholar] [CrossRef]
- Virtanen, I.; Tani, T.; Back, N.; Happola, O.; Laitinen, L.; Kiviluoto, T.; Salo, J.; Burgeson, R.E.; Lehto, V.P.; Kivilaakso, E. Differential expression of laminin chains and their integrin receptors in human gastric mucosa. Am. J. Pathol. 1995, 147, 1123–1132. [Google Scholar] [PubMed]
- Mao, Y.; Schwarzbauer, J.E. Fibronectin fibrillogenesis, a cell-mediated matrix assembly process. Matrix Biol. 2005, 24, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Pankov, R.; Yamada, K.M. Fibronectin at a glance. J. Cell Sci. 2002, 115, 3861–3863. [Google Scholar] [CrossRef] [PubMed]
- Gillessen, A.; Shahin, M.; Pohle, T.; Foerster, E.; Domschke, W. Collagen mRNA and fibronectin are increased in healing gastric ulcers in man. J. Physiol. Pharm. 1995, 46, 57–62. [Google Scholar]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Mohan, V.; Das, A.; Sagi, I. Emerging roles of ECM remodeling processes in cancer. Semin. Cancer Biol. 2019. [Google Scholar] [CrossRef]
- Pickup, M.W.; Mouw, J.K.; Weaver, V.M. The extracellular matrix modulates the hallmarks of cancer. EMBO Rep. 2014, 15, 1243–1253. [Google Scholar] [CrossRef]
- Lauren, P. The Two Histological Main Types of Gastric Carcinoma: Diffuse and So-Called Intestinal-Type Carcinoma. An Attempt at a Histo-Clinical Classification. Acta Pathol. Microbiol. Scand. 1965, 64, 31–49. [Google Scholar] [CrossRef]
- Vauhkonen, M.; Vauhkonen, H.; Sipponen, P. Pathology and molecular biology of gastric cancer. Best Pr. Res. Clin. Gastroenterol. 2006, 20, 651–674. [Google Scholar] [CrossRef]
- Hartgrink, H.H.; Jansen, E.P.; van Grieken, N.C.; van de Velde, C.J. Gastric cancer. Lancet 2009, 374, 477–490. [Google Scholar] [CrossRef]
- Carneiro, F.; Seixas, M.; Sobrinho-Simoes, M. New elements for an updated classification of the carcinomas of the stomach. Pathol. Res. Pr. 1995, 191, 571–584. [Google Scholar] [CrossRef]
- Ming, S.C. Gastric carcinoma. A pathobiological classification. Cancer 1977, 39, 2475–2485. [Google Scholar] [CrossRef]
- Correa, P. A human model of gastric carcinogenesis. Cancer Res. 1988, 48, 3554–3560. [Google Scholar] [PubMed]
- Correa, P.; Piazuelo, M.B. The gastric precancerous cascade. J. Dig. Dis. 2012, 13, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Rowe, R.G.; Weiss, S.J. Breaching the basement membrane: Who, when and how? Trends Cell Biol. 2008, 18, 560–574. [Google Scholar] [CrossRef] [PubMed]
- Bergin, P.J.; Anders, E.; Sicheng, W.; Erik, J.; Jennie, A.; Hans, L.; Pierre, M.; Qiang, P.H.; Marianne, Q.J. Increased production of matrix metalloproteinases in Helicobacter pylori-associated human gastritis. Helicobacter 2004, 9, 201–210. [Google Scholar] [CrossRef]
- Costa, A.M.; Ferreira, R.M.; Pinto-Ribeiro, I.; Sougleri, I.S.; Oliveira, M.J.; Carreto, L.; Santos, M.A.; Sgouras, D.N.; Carneiro, F.; Leite, M.; et al. Helicobacter pylori Activates Matrix Metalloproteinase 10 in Gastric Epithelial Cells via EGFR and ERK-mediated Pathways. J. Infect. Dis. 2016, 213, 1767–1776. [Google Scholar] [CrossRef]
- Oliveira, M.J.; Costa, A.C.; Costa, A.M.; Henriques, L.; Suriano, G.; Atherton, J.C.; Machado, J.C.; Carneiro, F.; Seruca, R.; Mareel, M.; et al. Helicobacter pylori induces gastric epithelial cell invasion in a c-Met and type IV secretion system-dependent manner. J. Biol. Chem. 2006, 281, 34888–34896. [Google Scholar] [CrossRef]
- Lee, L.Y.; Wu, C.M.; Wang, C.C.; Yu, J.S.; Liang, Y.; Huang, K.H.; Lo, C.H.; Hwang, T.L. Expression of matrix metalloproteinases MMP-2 and MMP-9 in gastric cancer and their relation to claudin-4 expression. Histol. Histopathol. 2008, 23, 515–521. [Google Scholar]
- Shen, W.; Xi, H.; Wei, B.; Chen, L. The prognostic role of matrix metalloproteinase 2 in gastric cancer: A systematic review with meta-analysis. J. Cancer Res. Clin. Oncol. 2014, 140, 1003–1009. [Google Scholar] [CrossRef]
- Wang, H.L.; Zhou, P.Y.; Zhang, Y.; Liu, P. Relationships between abnormal MMP2 expression and prognosis in gastric cancer: A meta-analysis of cohort studies. Cancer Biother. Radiopharm. 2014, 29, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Soleyman-Jahi, S.; Nedjat, S.; Abdirad, A.; Hoorshad, N.; Heidari, R.; Zendehdel, K. Prognostic significance of matrix metalloproteinase-7 in gastric cancer survival: A meta-analysis. PLoS ONE 2014, 10, e0122316. [Google Scholar] [CrossRef]
- Chen, S.Z.; Yao, H.Q.; Zhu, S.Z.; Li, Q.Y.; Guo, G.H.; Yu, J. Expression levels of matrix metalloproteinase-9 in human gastric carcinoma. Oncol. Lett. 2015, 9, 915–919. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Emara, M.; Cheung, P.Y.; Grabowski, K.; Sawicki, G.; Wozniak, M. Serum levels of matrix metalloproteinase-2 and -9 and conventional tumor markers (CEA and CA 19-9) in patients with colorectal and gastric cancers. Clin. Chem. Lab. Med. 2009, 47, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Huang, J.; Sun, S.; Zhou, Z.; Zhang, J.; Gao, F.; Sun, Q. Expression of matrix metalloproteinase-9, cyclooxygenase-2 and vascular endothelial growth factor are increased in gastrointestinal stromal tumors. Int J. Clin. Exp. Med. 2015, 8, 6495–6501. [Google Scholar]
- Chen, J.; Chen, L.J.; Zhou, H.C.; Yang, R.B.; Lu, Y.; Xia, Y.L.; Wu, W.; Hu, L.W. Prognostic value of matrix metalloproteinase-9 in gastric cancer: A meta-analysis. Hepatogastroenterology 2014, 61, 518–524. [Google Scholar]
- Shimura, T.; Dagher, A.; Sachdev, M.; Ebi, M.; Yamada, T.; Yamada, T.; Joh, T.; Moses, M.A. Urinary ADAM12 and MMP-9/NGAL complex detect the presence of gastric cancer. Cancer Prev. Res. (Phila) 2015, 8, 240–248. [Google Scholar] [CrossRef]
- Xie, X.; Liu, X.; Zhang, Q.; Yu, J. Overexpression of collagen VI alpha3 in gastric cancer. Oncol. Lett. 2014, 7, 1537–1543. [Google Scholar] [CrossRef]
- Guszczyn, T.; Sobolewski, K. Deregulation of collagen metabolism in human stomach cancer. Pathobiology 2004, 71, 308–313. [Google Scholar] [CrossRef]
- Jang, M.; Koh, I.; Lee, J.E.; Lim, J.Y.; Cheong, J.H.; Kim, P. Increased extracellular matrix density disrupts E-cadherin/beta-catenin complex in gastric cancer cells. Biomater. Sci. 2018, 6, 2704–2713. [Google Scholar] [CrossRef]
- Yamamoto, H.; Kitadai, Y.; Yamamoto, H.; Oue, N.; Ohdan, H.; Yasui, W.; Kikuchi, A. Laminin gamma2 mediates Wnt5a-induced invasion of gastric cancer cells. Gastroenterology 2009, 137, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Caldeira, J.; Figueiredo, J.; Bras-Pereira, C.; Carneiro, P.; Moreira, A.M.; Pinto, M.T.; Relvas, J.B.; Carneiro, F.; Barbosa, M.; Casares, F.; et al. E-cadherin-defective gastric cancer cells depend on Laminin to survive and invade. Hum. Mol. Genet. 2015, 24, 5891–5900. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, Q.; Yu, Z.; Wu, X.; Chen, X.; Li, J.; Li, C.; Yan, M.; Zhu, Z.; Liu, B.; et al. Cancer-associated fibroblast-derived Lumican promotes gastric cancer progression via the integrin beta1-FAK signaling pathway. Int. J. Cancer 2017, 141, 998–1010. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.Y.; Jin, H.; Liu, X.; Siu, J.M.; Wong, Y.P.; Ng, E.K.; Yu, J.; Leung, W.K.; Sung, J.J.; Chan, F.K. Fibulin 1 is downregulated through promoter hypermethylation in gastric cancer. Br. J. Cancer 2008, 99, 2083–2087. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.H.; Wang, Y.M.; Jiang, Y.Z.; Ma, S.J.; Zhong, Q.; Wan, Y.Y.; Wang, X.W. NID2 can serve as a potential prognosis prediction biomarker and promotes the invasion and migration of gastric cancer. Pathol. Res. Pr. 2019, 215, 152553. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.Y.; Han, Y.C.; Wu, S.H.; Lv, Z.H. Expression of connective tissue growth factor in tumor tissues is an independent predictor of poor prognosis in patients with gastric cancer. World J. Gastroenterol. 2008, 14, 2110–2114. [Google Scholar] [CrossRef]
- Zhong, H.; Li, X.; Zhang, J.; Wu, X. Overexpression of periostin is positively associated with gastric cancer metastasis through promoting tumor metastasis and invasion. J. Cell Biochem. 2019, 120, 9927–9935. [Google Scholar] [CrossRef] [PubMed]
- Theocharis, A.D.; Vynios, D.H.; Papageorgakopoulou, N.; Skandalis, S.S.; Theocharis, D.A. Altered content composition and structure of glycosaminoglycans and proteoglycans in gastric carcinoma. Int. J. Biochem. Cell Biol. 2003, 35, 376–390. [Google Scholar] [CrossRef]
- Wang, B.; Li, G.X.; Zhang, S.G.; Wang, Q.; Wen, Y.G.; Tang, H.M.; Zhou, C.Z.; Xing, A.Y.; Fan, J.W.; Yan, D.W.; et al. Biglycan expression correlates with aggressiveness and poor prognosis of gastric cancer. Exp. Biol. Med. (Maywood) 2011, 236, 1247–1253. [Google Scholar] [CrossRef]
- Naumova, L.A.; Osipova, O.N.; Klinnikova, M.G. Immunistochemical Analysis of the Expression of TGFbeta, Galectin-1, Vimentin, and Thrombospondin in Gastric Cancer Associated with Systemic Undifferentiated Connective Tissue Dysplasia. Bull. Exp. Biol. Med. 2019, 166, 774–778. [Google Scholar] [CrossRef]
- De Vega, S.; Iwamoto, T.; Yamada, Y. Fibulins: Multiple roles in matrix structures and tissue functions. Cell Mol. Life Sci. 2009, 66, 1890–1902. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O.; Naba, A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb. Perspect. Biol. 2012, 4, a004903. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, Y.; Kunita, A.; Iwata, C.; Komura, D.; Nishiyama, T.; Shimazu, K.; Takeshita, K.; Shibahara, J.; Kii, I.; Morishita, Y.; et al. The niche component periostin is produced by cancer-associated fibroblasts, supporting growth of gastric cancer through ERK activation. Am. J. Pathol. 2014, 184, 859–870. [Google Scholar] [CrossRef]
- Malanchi, I.; Santamaria-Martinez, A.; Susanto, E.; Peng, H.; Lehr, H.A.; Delaloye, J.F.; Huelsken, J. Interactions between cancer stem cells and their niche govern metastatic colonization. Nature 2011, 481, 85–89. [Google Scholar] [CrossRef]
- Yuzhalin, A.E.; Urbonas, T.; Silva, M.A.; Muschel, R.J.; Gordon-Weeks, A.N. A core matrisome gene signature predicts cancer outcome. Br. J. Cancer 2018, 118, 435–440. [Google Scholar] [CrossRef]
- Boussioutas, A.; Li, H.; Liu, J.; Waring, P.; Lade, S.; Holloway, A.J.; Taupin, D.; Gorringe, K.; Haviv, I.; Desmond, P.V.; et al. Distinctive patterns of gene expression in premalignant gastric mucosa and gastric cancer. Cancer Res. 2003, 63, 2569–2577. [Google Scholar] [PubMed]
- Jinawath, N.; Furukawa, Y.; Hasegawa, S.; Li, M.; Tsunoda, T.; Satoh, S.; Yamaguchi, T.; Imamura, H.; Inoue, M.; Shiozaki, H.; et al. Comparison of gene-expression profiles between diffuse- and intestinal-type gastric cancers using a genome-wide cDNA microarray. Oncogene 2004, 23, 6830–6844. [Google Scholar] [CrossRef] [PubMed]
- Ai, K.; Jia, Y.; Li, J.; Wang, C.; Wang, Y. Systematic analysis of multigene predictors in gastric cancer exploiting gene expression signature. J. Cell Biochem. 2018, 120, 8069–8077. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S.; Katz, B.Z.; Lafrenie, R.M.; Yamada, K.M. Fibronectin and integrins in cell adhesion, signaling, and morphogenesis. Ann. N. Y. Acad. Sci. 1998, 857, 119–129. [Google Scholar] [CrossRef]
- Berrier, A.L.; Yamada, K.M. Cell-matrix adhesion. J. Cell Physiol. 2007, 213, 565–573. [Google Scholar] [CrossRef]
- Kim, C.; Ye, F.; Ginsberg, M.H. Regulation of integrin activation. Annu. Rev. Cell Dev. Biol. 2011, 27, 321–345. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhu, J.; Springer, T.A. Complete integrin headpiece opening in eight steps. J. Cell Biol. 2013, 201, 1053–1068. [Google Scholar] [CrossRef] [PubMed]
- Calderwood, D.A. Integrin activation. J. Cell Sci. 2004, 117, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Radisky, D.; Muschler, J.; Bissell, M.J. Order and disorder: The role of extracellular matrix in epithelial cancer. Cancer Invest. 2002, 20, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Ruoslahti, E. RGD and other recognition sequences for integrins. Annu Rev. Cell Dev. Biol. 1996, 12, 697–715. [Google Scholar] [CrossRef]
- Humphries, J.D.; Byron, A.; Humphries, M.J. Integrin ligands at a glance. J. Cell Sci. 2006, 119, 3901–3903. [Google Scholar] [CrossRef]
- Boudreau, N.J.; Jones, P.L. Extracellular matrix and integrin signalling: The shape of things to come. Biochem J. 1999, 339 Pt. 3, 481–488. [Google Scholar] [CrossRef]
- Winograd-Katz, S.E.; Fassler, R.; Geiger, B.; Legate, K.R. The integrin adhesome: From genes and proteins to human disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 273–288. [Google Scholar] [CrossRef]
- Nurden, A.T. Glanzmann thrombasthenia. Orphanet J. Rare Dis. 2006, 1, 10. [Google Scholar] [CrossRef]
- Springer, T.A.; Thompson, W.S.; Miller, L.J.; Schmalstieg, F.C.; Anderson, D.C. Inherited deficiency of the Mac-1, LFA-1, p150,95 glycoprotein family and its molecular basis. J. Exp. Med. 1984, 160, 1901–1918. [Google Scholar] [CrossRef]
- Kishimoto, T.K.; Hollander, N.; Roberts, T.M.; Anderson, D.C.; Springer, T.A. Heterogeneous mutations in the beta subunit common to the LFA-1, Mac-1, and p150,95 glycoproteins cause leukocyte adhesion deficiency. Cell 1987, 50, 193–202. [Google Scholar] [CrossRef]
- Bouvard, D.; Pouwels, J.; De Franceschi, N.; Ivaska, J. Integrin inactivators: Balancing cellular functions in vitro and in vivo. Nat. Rev. Mol. Cell Biol. 2013, 14, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Das, V.; Kalyan, G.; Hazra, S.; Pal, M. Understanding the role of structural integrity and differential expression of integrin profiling to identify potential therapeutic targets in breast cancer. J. Cell Physiol. 2018, 233, 168–185. [Google Scholar] [CrossRef] [PubMed]
- Nieberler, M.; Reuning, U.; Reichart, F.; Notni, J.; Wester, H.J.; Schwaiger, M.; Weinmuller, M.; Rader, A.; Steiger, K.; Kessler, H. Exploring the Role of RGD-Recognizing Integrins in Cancer. Cancers (Basel) 2017, 9, 116. [Google Scholar] [CrossRef] [PubMed]
- Kren, A.; Baeriswyl, V.; Lehembre, F.; Wunderlin, C.; Strittmatter, K.; Antoniadis, H.; Fassler, R.; Cavallaro, U.; Christofori, G. Increased tumor cell dissemination and cellular senescence in the absence of beta1-integrin function. EMBO J. 2007, 26, 2832–2842. [Google Scholar] [CrossRef]
- White, D.E.; Kurpios, N.A.; Zuo, D.; Hassell, J.A.; Blaess, S.; Mueller, U.; Muller, W.J. Targeted disruption of beta1-integrin in a transgenic mouse model of human breast cancer reveals an essential role in mammary tumor induction. Cancer Cell 2004, 6, 159–170. [Google Scholar] [CrossRef]
- Dingemans, A.M.; van den Boogaart, V.; Vosse, B.A.; van Suylen, R.J.; Griffioen, A.W.; Thijssen, V.L. Integrin expression profiling identifies integrin alpha5 and beta1 as prognostic factors in early stage non-small cell lung cancer. Mol. Cancer 2010, 9, 152. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, A.; Tsugawa, S.; Boku, A.; Kobayashi, M.; Minamoto, T.; Nakanishi, I.; Oda, Y. Expression of alphav integrin family in gastric carcinomas: Increased alphavbeta6 is associated with lymph node metastasis. Pathol. Res. Pr. 2003, 199, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Lian, P.L.; Liu, Z.; Yang, G.Y.; Zhao, R.; Zhang, Z.Y.; Chen, Y.G.; Zhuang, Z.N.; Xu, K.S. Integrin alphavbeta6 and matrix metalloproteinase 9 correlate with survival in gastric cancer. World J. Gastroenterol. 2016, 22, 3852–3859. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Xu, K.S.; Wang, J.S.; Yang, G.Y.; Wang, W.; Wang, J.Y.; Niu, W.B.; Liu, E.Y.; Mi, Y.T.; Niu, J. Integrin alphanvbeta6 acts as a prognostic indicator in gastric carcinoma. Clin. Oncol. 2008, 20, 61–66. [Google Scholar] [CrossRef]
- Zhao, R.; Liu, X.Q.; Wu, X.P.; Liu, Y.F.; Zhang, Z.Y.; Yang, G.Y.; Guo, S.; Niu, J.; Wang, J.Y.; Xu, K.S. Vascular endothelial growth factor (VEGF) enhances gastric carcinoma invasiveness via integrin alpha(v)beta6. Cancer Lett. 2010, 287, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.; Zhou, R.; Xu, X.; Tian, T.; Liu, Y.; Liu, Y.; Lian, P.; Wang, J.; Xu, K. Clinical significance of integrin alphavbeta6 expression effects on gastric carcinoma invasiveness and progression via cancer-associated fibroblasts. Med. Oncol. 2013, 30, 580. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.Y.; Xu, K.S.; Pan, Z.Q.; Zhang, Z.Y.; Mi, Y.T.; Wang, J.S.; Chen, R.; Niu, J. Integrin alpha v beta 6 mediates the potential for colon cancer cells to colonize in and metastasize to the liver. Cancer Sci. 2008, 99, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Niu, J.; Dorahy, D.J.; Scott, R.; Agrez, M.V. Integrin alpha(v)beta6-associated ERK2 mediates MMP-9 secretion in colon cancer cells. Br. J. Cancer 2002, 87, 348–351. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ura, H.; Denno, R.; Hirata, K.; Yamaguchi, K.; Yasoshima, T. Separate functions of alpha2beta1 and alpha3beta1 integrins in the metastatic process of human gastric carcinoma. Surg. Today 1998, 28, 1001–1006. [Google Scholar] [CrossRef]
- Lin, M.T.; Chang, C.C.; Lin, B.R.; Yang, H.Y.; Chu, C.Y.; Wu, M.H.; Kuo, M.L. Elevated expression of Cyr61 enhances peritoneal dissemination of gastric cancer cells through integrin alpha2beta1. J. Biol. Chem. 2007, 282, 34594–34604. [Google Scholar] [CrossRef]
- Ren, J.; Xu, S.; Guo, D.; Zhang, J.; Liu, S. Increased expression of alpha5beta1-integrin is a prognostic marker for patients with gastric cancer. Clin. Transl. Oncol. 2014, 16, 668–674. [Google Scholar] [CrossRef]
- Boger, C.; Warneke, V.S.; Behrens, H.M.; Kalthoff, H.; Goodman, S.L.; Becker, T.; Rocken, C. Integrins alphavbeta3 and alphavbeta5 as prognostic, diagnostic, and therapeutic targets in gastric cancer. Gastric Cancer 2015, 18, 784–795. [Google Scholar] [CrossRef]
- Lau, L.F. CCN1/CYR61: The very model of a modern matricellular protein. Cell Mol. Life Sci. 2011, 68, 3149–3163. [Google Scholar] [CrossRef]
- Jun, J.I.; Lau, L.F. Taking aim at the extracellular matrix: CCN proteins as emerging therapeutic targets. Nat. Rev. Drug Discov. 2011, 10, 945–963. [Google Scholar] [CrossRef]
- Lau, L.F. Cell surface receptors for CCN proteins. J. Cell Commun Signal. 2016, 10, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.; Nakachi, K.; Wang, H.; Elashoff, R.; Koeffler, H.P. Elevated levels of connective tissue growth factor, WISP-1, and CYR61 in primary breast cancers associated with more advanced features. Cancer Res. 2001, 61, 8917–8923. [Google Scholar] [PubMed]
- Mierke, C.T.; Frey, B.; Fellner, M.; Herrmann, M.; Fabry, B. Integrin alpha5beta1 facilitates cancer cell invasion through enhanced contractile forces. J. Cell Sci. 2011, 124, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Trikha, M.; Zhou, Z.; Nemeth, J.A.; Chen, Q.; Sharp, C.; Emmell, E.; Giles-Komar, J.; Nakada, M.T. CNTO 95, a fully human monoclonal antibody that inhibits alphav integrins, has antitumor and antiangiogenic activity in vivo. Int. J. Cancer 2004, 110, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Manning, C.D.; Millar, H.; McCabe, F.L.; Ferrante, C.; Sharp, C.; Shahied-Arruda, L.; Doshi, P.; Nakada, M.T.; Anderson, G.M. CNTO 95, a fully human anti alphav integrin antibody, inhibits cell signaling, migration, invasion, and spontaneous metastasis of human breast cancer cells. Clin. Exp. Metastasis 2008, 25, 139–148. [Google Scholar] [CrossRef]
- Ishii, Y.; Ochiai, A.; Yamada, T.; Akimoto, S.; Yanagihara, K.; Kitajima, M.; Hirohashi, S. Integrin alpha6beta4 as a suppressor and a predictive marker for peritoneal dissemination in human gastric cancer. Gastroenterology 2000, 118, 497–506. [Google Scholar] [CrossRef]
- Bianconi, D.; Unseld, M.; Prager, G.W. Integrins in the Spotlight of Cancer. Int. J. Mol. Sci. 2016, 17, 2037. [Google Scholar] [CrossRef]
- Truong, H.; Danen, E.H.J. Integrin switching modulates adhesion dynamics and cell migration. Cell Adhes. Migr. 2009, 3, 179–181. [Google Scholar] [CrossRef]
- Guo, W.; Giancotti, F.G. Integrin signalling during tumour progression. Nat. Rev. Mol. Cell Biol. 2004, 5, 816–826. [Google Scholar] [CrossRef]
- Ramirez, N.E.; Zhang, Z.; Madamanchi, A.; Boyd, K.L.; O’Rear, L.D.; Nashabi, A.; Li, Z.; Dupont, W.D.; Zijlstra, A.; Zutter, M.M. The α2β1 integrin is a metastasis suppressor in mouse models and human cancer. J. Clin. Investig. 2011, 121, 226–237. [Google Scholar] [CrossRef]
- Ramovs, V.; Secades, P.; Song, J.-Y.; Thijssen, B.; Kreft, M.; Sonnenberg, A. Absence of integrin α3β1 promotes the progression of HER2-driven breast cancer in vivo. Breast Cancer Res. Bcr. 2019, 21, 63. [Google Scholar] [CrossRef] [PubMed]
- Friedrichs, K.; Ruiz, P.; Franke, F.; Gille, I.; Terpe, H.J.; Imhof, B.A. High expression level of alpha 6 integrin in human breast carcinoma is correlated with reduced survival. Cancer Res. 1995, 55, 901–906. [Google Scholar] [PubMed]
- Diaz, L.K.; Cristofanilli, M.; Zhou, X.; Welch, K.L.; Smith, T.L.; Yang, Y.; Sneige, N.; Sahin, A.A.; Gilcrease, M.Z. Beta4 integrin subunit gene expression correlates with tumor size and nuclear grade in early breast cancer. Mod. Pathol 2005, 18, 1165–1175. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Bachelier, R.; Treilleux, I.; Pujuguet, P.; Peyruchaud, O.; Baron, R.; Clement-Lacroix, P.; Clezardin, P. Tumor alphavbeta3 integrin is a therapeutic target for breast cancer bone metastases. Cancer Res. 2007, 67, 5821–5830. [Google Scholar] [CrossRef] [PubMed]
- De Franceschi, N.; Hamidi, H.; Alanko, J.; Sahgal, P.; Ivaska, J. Integrin traffic—the update. J. Cell Sci. 2015, 128, 839–852. [Google Scholar] [CrossRef] [PubMed]
- Longmate, W.; DiPersio, C.M. Beyond adhesion: emerging roles for integrins in control of the tumor microenvironment. F1000Research 2017, 6, 1612. [Google Scholar] [CrossRef] [PubMed]
- Horton, E.R.; Byron, A.; Askari, J.A.; Ng, D.H.J.; Millon-Frémillon, A.; Robertson, J.; Koper, E.J.; Paul, N.R.; Warwood, S.; Knight, D.; et al. Definition of a consensus integrin adhesome and its dynamics during adhesion complex assembly and disassembly. Nat. Cell Biol. 2015, 17, 1577–1587. [Google Scholar] [CrossRef]
- Schaller, M.D.; Hildebrand, J.D.; Shannon, J.D.; Fox, J.W.; Vines, R.R.; Parsons, J.T. Autophosphorylation of the focal adhesion kinase, pp125FAK, directs SH2-dependent binding of pp60src. Mol. Cell. Biol. 1994, 14, 1680–1688. [Google Scholar] [CrossRef]
- Schaller, M.D. Biochemical signals and biological responses elicited by the focal adhesion kinase. Biochim. Et Biophys. Acta (Bba) Mol. Cell Res. 2001, 1540, 1–21. [Google Scholar] [CrossRef]
- Guan, J.-L. Role of focal adhesion kinase in integrin signaling. Int. J. Biochem. Cell Biol. 1997, 29, 1085–1096. [Google Scholar] [CrossRef]
- Mitra, S.K.; Schlaepfer, D.D. Integrin-regulated FAK–Src signaling in normal and cancer cells. Curr. Opin. Cell Biol. 2006, 18, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Kuwada, S.K.; Li, X. Integrin alpha5/beta1 mediates fibronectin-dependent epithelial cell proliferation through epidermal growth factor receptor activation. Mol. Biol. Cell 2000, 11, 2485–2496. [Google Scholar] [CrossRef]
- Yee, K.; Weaver, V.; Hammer, D. Integrin-mediated signaling through the MAP-kinase pathway. IET Syst. Biol. 2008, 2, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, M.C.; Er, E.E.; Blenis, J. The Ras-ERK and PI3K-mTOR pathways: Cross-talk and compensation. Trends Biochem. Sci. 2011, 36, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Ouyang, G.; Bao, S. The activation of Akt/PKB signaling pathway and cell survival. J. Cell. Mol. Med. 2005, 9, 59–71. [Google Scholar] [CrossRef]
- Laplante, M.; Sabatini, D.M. mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef]
- Brakebusch, C.; Fässler, R. The integrin-actin connection, an eternal love affair. EMBO J. 2003, 22, 2324–2333. [Google Scholar] [CrossRef]
- Ridley, A.J. Rho GTPases and cell migration. J. Cell Sci. 2001, 114, 2713. [Google Scholar]
- Lai, I.R.; Chu, P.Y.; Lin, H.S.; Liou, J.Y.; Jan, Y.J.; Lee, J.C.; Shen, T.L. Phosphorylation of focal adhesion kinase at Tyr397 in gastric carcinomas and its clinical significance. Am. J. Pathol. 2010, 177, 1629–1637. [Google Scholar] [CrossRef]
- Tegtmeyer, N.; Wittelsberger, R.; Hartig, R.; Wessler, S.; Martinez-Quiles, N.; Backert, S. Serine phosphorylation of cortactin controls focal adhesion kinase activity and cell scattering induced by Helicobacter pylori. Cell Host Microbe 2011, 9, 520–531. [Google Scholar] [CrossRef]
- Moese, S.; Selbach, M.; Brinkmann, V.; Karlas, A.; Haimovich, B.; Backert, S.; Meyer, T.F. The Helicobacter pylori CagA protein disrupts matrix adhesion of gastric epithelial cells by dephosphorylation of vinculin. Cell Microbiol. 2007, 9, 1148–1161. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, J.; Sun, X.; Wang, X. The expressions and interrelation of p130Cas and PTEN in gastric cancer. Chin. Ger. J. Clin. Oncol. 2009, 8, 332–335. [Google Scholar] [CrossRef]
- Chen, D.L.; Wang, Z.Q.; Ren, C.; Zeng, Z.L.; Wang, D.S.; Luo, H.Y.; Wang, F.; Qiu, M.Z.; Bai, L.; Zhang, D.S.; et al. Abnormal expression of paxillin correlates with tumor progression and poor survival in patients with gastric cancer. J. Transl. Med. 2013, 11, 277. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.J.; Zhao, E.H.; Zhao, S.; Zheng, X.; Zheng, H.C.; Takano, Y.; Song, H.R. Paxillin expression is closely linked to the pathogenesis, progression and prognosis of gastric carcinomas. Oncol. Lett. 2014, 7, 189–194. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Springer, T.A.; Dustin, M.L. Integrin inside-out signaling and the immunological synapse. Curr. Opin. Cell Biol. 2012, 24, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Ye, Y.; Dong, L.; Vainionpaa, S.; Mustonen, H.; Puolakkainen, P.; Wang, S. Kindlin-2: A novel adhesion protein related to tumor invasion, lymph node metastasis, and patient outcome in gastric cancer. Am. J. Surg. 2012, 203, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Ye, Y.; Kauttu, T.; Seppanen, H.; Vainionpaa, S.; Wang, S.; Mustonen, H.; Puolakkainen, P. Novel focal adhesion protein kindlin-2 promotes the invasion of gastric cancer cells through phosphorylation of integrin beta1 and beta3. J. Surg. Oncol. 2013, 108, 106–112. [Google Scholar] [CrossRef]
- Sun, Z.; Guo, S.S.; Fässler, R. Integrin-mediated mechanotransduction. J. Cell Biol. 2016, 215, 445. [Google Scholar] [CrossRef]
- Das, M.; Ithychanda, S.; Qin, J.; Plow, E.F. Mechanisms of talin-dependent integrin signaling and crosstalk. Biochim. Et Biophys. Acta 2014, 1838, 579–588. [Google Scholar] [CrossRef]
- Moser, M.; Legate, K.R.; Zent, R.; Fässler, R. The Tail of Integrins, Talin, and Kindlins. Science 2009, 324, 895. [Google Scholar] [CrossRef]
- Ye, F.; Snider, A.K.; Ginsberg, M.H. Talin and kindlin: The one-two punch in integrin activation. Front. Med. 2014, 8, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Kahner, B.N.; Kato, H.; Banno, A.; Ginsberg, M.H.; Shattil, S.J.; Ye, F. Kindlins, Integrin Activation and the Regulation of Talin Recruitment to αIIbβ3. PLoS ONE 2012, 7, e34056. [Google Scholar] [CrossRef] [PubMed]
- Reardon, D.A.; Fink, K.L.; Mikkelsen, T.; Cloughesy, T.F.; O’Neill, A.; Plotkin, S.; Glantz, M.; Ravin, P.; Raizer, J.J.; Rich, K.M.; et al. Randomized phase II study of cilengitide, an integrin-targeting arginine-glycine-aspartic acid peptide, in recurrent glioblastoma multiforme. J. Clin. Oncol. 2008, 26, 5610–5617. [Google Scholar] [CrossRef] [PubMed]
- Delbaldo, C.; Raymond, E.; Vera, K.; Hammershaimb, L.; Kaucic, K.; Lozahic, S.; Marty, M.; Faivre, S. Phase I and pharmacokinetic study of etaracizumab (Abegrin), a humanized monoclonal antibody against alphavbeta3 integrin receptor, in patients with advanced solid tumors. Invest. New Drugs 2008, 26, 35–43. [Google Scholar] [CrossRef]
- Beekman, K.W.; Colevas, A.D.; Cooney, K.; Dipaola, R.; Dunn, R.L.; Gross, M.; Keller, E.T.; Pienta, K.J.; Ryan, C.J.; Smith, D.; et al. Phase II evaluations of cilengitide in asymptomatic patients with androgen-independent prostate cancer: Scientific rationale and study design. Clin. Genitourin. Cancer 2006, 4, 299–302. [Google Scholar] [CrossRef]
- McNeel, D.G.; Eickhoff, J.; Lee, F.T.; King, D.M.; Alberti, D.; Thomas, J.P.; Friedl, A.; Kolesar, J.; Marnocha, R.; Volkman, J.; et al. Phase I trial of a monoclonal antibody specific for alphavbeta3 integrin (MEDI-522) in patients with advanced malignancies, including an assessment of effect on tumor perfusion. Clin. Cancer Res. 2005, 11, 7851–7860. [Google Scholar] [CrossRef]
- Raab-Westphal, S.; Marshall, J.F.; Goodman, S.L. Integrins as Therapeutic Targets: Successes and Cancers. Cancers (Basel) 2017, 9, 110. [Google Scholar] [CrossRef]
- Mitjans, F.; Sander, D.; Adan, J.; Sutter, A.; Martinez, J.M.; Jaggle, C.S.; Moyano, J.M.; Kreysch, H.G.; Piulats, J.; Goodman, S.L. An anti-alpha v-integrin antibody that blocks integrin function inhibits the development of a human melanoma in nude mice. J. Cell Sci. 1995, 108 Pt. 8, 2825–2838. [Google Scholar]
- Elez, E.; Kocakova, I.; Hohler, T.; Martens, U.M.; Bokemeyer, C.; Van Cutsem, E.; Melichar, B.; Smakal, M.; Csoszi, T.; Topuzov, E.; et al. Abituzumab combined with cetuximab plus irinotecan versus cetuximab plus irinotecan alone for patients with KRAS wild-type metastatic colorectal cancer: The randomised phase I/II POSEIDON trial. Ann. Oncol. 2015, 26, 132–140. [Google Scholar] [CrossRef]
- O’Day, S.; Pavlick, A.; Loquai, C.; Lawson, D.; Gutzmer, R.; Richards, J.; Schadendorf, D.; Thompson, J.A.; Gonzalez, R.; Trefzer, U.; et al. phase II study of intetumumab, an anti-alphav-integrin mAb, alone and with dacarbazine in stage IV melanoma. Br. J. Cancer 2011, 105, 346–352. [Google Scholar] [CrossRef]
- Heidenreich, A.; Rawal, S.K.; Szkarlat, K.; Bogdanova, N.; Dirix, L.; Stenzl, A.; Welslau, M.; Wang, G.; Dawkins, F.; de Boer, C.J.; et al. A randomized, double-blind, multicenter, phase 2 study of a human monoclonal antibody to human alphanu integrins (intetumumab) in combination with docetaxel and prednisone for the first-line treatment of patients with metastatic castration-resistant prostate cancer. Ann. Oncol. 2013, 24, 329–336. [Google Scholar] [PubMed]
- Van Aarsen, L.A.; Leone, D.R.; Ho, S.; Dolinski, B.M.; McCoon, P.E.; LePage, D.J.; Kelly, R.; Heaney, G.; Rayhorn, P.; Reid, C.; et al. Antibody-mediated blockade of integrin alpha v beta 6 inhibits tumor progression in vivo by a transforming growth factor-beta-regulated mechanism. Cancer Res. 2008, 68, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Maden, C.H.; Fairman, D.; Chalker, M.; Costa, M.J.; Fahy, W.A.; Garman, N.; Lukey, P.T.; Mant, T.; Parry, S.; Simpson, J.K.; et al. Safety, tolerability and pharmacokinetics of GSK3008348, a novel integrin alphavbeta6 inhibitor, in healthy participants. Eur. J. Clin. Pharm. 2018, 74, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, T.; Yashiro, M.; Nishimura, S.; Inoue, T.; Fujihara, T.; Sawada, T.; Kato, Y.; Seki, S.; Hirakawa-Ys Chung, K. Increased expression of alpha2beta1-integrin in the peritoneal dissemination of human gastric carcinoma. Int. J. Mol. Med. 2000, 5, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, T.; Endo, Y.; Yonemura, Y.; Nojima, N.; Fujita, H.; Fujimura, T.; Obata, T.; Yamaguchi, T.; Sasaki, T. Significance of integrin alpha2/beta1 in peritoneal dissemination of a human gastric cancer xenograft model. Int. J. Oncol. 2001, 18, 809–815. [Google Scholar] [PubMed]
- Naci, D.; Vuori, K.; Aoudjit, F. Alpha2beta1 integrin in cancer development and chemoresistance. Semin. Cancer Biol. 2015, 35, 145–153. [Google Scholar] [CrossRef]
- Funahashi, Y.; Sugi, N.H.; Semba, T.; Yamamoto, Y.; Hamaoka, S.; Tsukahara-Tamai, N.; Ozawa, Y.; Tsuruoka, A.; Nara, K.; Takahashi, K.; et al. Sulfonamide derivative, E7820, is a unique angiogenesis inhibitor suppressing an expression of integrin alpha2 subunit on endothelium. Cancer Res. 2002, 62, 6116–6123. [Google Scholar]
- Milojkovic Kerklaan, B.; Slater, S.; Flynn, M.; Greystoke, A.; Witteveen, P.O.; Megui-Roelvink, M.; de Vos, F.; Dean, E.; Reyderman, L.; Ottesen, L.; et al. A phase I, dose escalation, pharmacodynamic, pharmacokinetic, and food-effect study of alpha2 integrin inhibitor E7820 in patients with advanced solid tumors. Invest. New Drugs 2016, 34, 329–337. [Google Scholar] [CrossRef]
- Kim, M.Y.; Cho, W.D.; Hong, K.P.; Choi da, B.; Hong, J.W.; Kim, S.; Moon, Y.R.; Son, S.M.; Lee, O.J.; Lee, H.C.; et al. Novel monoclonal antibody against beta 1 integrin enhances cisplatin efficacy in human lung adenocarcinoma cells. J. Biomed. Res. 2016, 30, 217–224. [Google Scholar]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef]
- Salmon, H.; Franciszkiewicz, K.; Damotte, D.; Dieu-Nosjean, M.C.; Validire, P.; Trautmann, A.; Mami-Chouaib, F.; Donnadieu, E. Matrix architecture defines the preferential localization and migration of T cells into the stroma of human lung tumors. J. Clin. Investig. 2012, 122, 899–910. [Google Scholar] [CrossRef] [PubMed]
- Hirte, H.; Vergote, I.B.; Jeffrey, J.R.; Grimshaw, R.N.; Coppieters, S.; Schwartz, B.; Tu, D.; Sadura, A.; Brundage, M.; Seymour, L. A phase III randomized trial of BAY 12-9566 (tanomastat) as maintenance therapy in patients with advanced ovarian cancer responsive to primary surgery and paclitaxel/platinum containing chemotherapy: A National Cancer Institute of Canada Clinical Trials Group Study. Gynecol. Oncol. 2006, 102, 300–308. [Google Scholar] [PubMed]
- Bramhall, S.R.; Schulz, J.; Nemunaitis, J.; Brown, P.D.; Baillet, M.; Buckels, J.A. A double-blind placebo-controlled, randomised study comparing gemcitabine and marimastat with gemcitabine and placebo as first line therapy in patients with advanced pancreatic cancer. Br. J. Cancer 2002, 87, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Sparano, J.A.; Bernardo, P.; Stephenson, P.; Gradishar, W.J.; Ingle, J.N.; Zucker, S.; Davidson, N.E. Randomized phase III trial of marimastat versus placebo in patients with metastatic breast cancer who have responding or stable disease after first-line chemotherapy: Eastern Cooperative Oncology Group trial E2196. J. Clin. Oncol. 2004, 22, 4683–4690. [Google Scholar] [CrossRef]
- Parsons, S.L.; Watson, S.A.; Steele, R.J. Phase I/II trial of batimastat, a matrix metalloproteinase inhibitor, in patients with malignant ascites. Eur. J. Surg. Oncol. 1997, 23, 526–531. [Google Scholar] [CrossRef]
- Winer, A.; Adams, S.; Mignatti, P. Matrix Metalloproteinase Inhibitors in Cancer Therapy: Turning Past Failures Into Future Successes. Mol. Cancer 2018, 17, 1147–1155. [Google Scholar] [CrossRef]
- Smith-Mungo, L.I.; Kagan, H.M. Lysyl oxidase: Properties, regulation and multiple functions in biology. Matrix Biol. 1998, 16, 387–398. [Google Scholar] [CrossRef]
- Cox, T.R.; Bird, D.; Baker, A.M.; Barker, H.E.; Ho, M.W.; Lang, G.; Erler, J.T. LOX-mediated collagen crosslinking is responsible for fibrosis-enhanced metastasis. Cancer Res. 2013, 73, 1721–1732. [Google Scholar] [CrossRef]
- Barry-Hamilton, V.; Spangler, R.; Marshall, D.; McCauley, S.; Rodriguez, H.M.; Oyasu, M.; Mikels, A.; Vaysberg, M.; Ghermazien, H.; Wai, C.; et al. Allosteric inhibition of lysyl oxidase-like-2 impedes the development of a pathologic microenvironment. Nat. Med. 2010, 16, 1009–1017. [Google Scholar] [CrossRef]
- Hu, W.Q.; Wang, L.W.; Yuan, J.P.; Yan, S.G.; Li, J.D.; Zhao, H.L.; Peng, C.W.; Yang, G.F.; Li, Y. High expression of transform growth factor beta 1 in gastric cancer confers worse outcome: Results of a cohort study on 184 patients. Hepatogastroenterology 2014, 61, 245–250. [Google Scholar]
- Roberts, A.B.; Heine, U.I.; Flanders, K.C.; Sporn, M.B. Transforming growth factor-beta. Major role in regulation of extracellular matrix. Ann. N. Y. Acad. Sci. 1990, 580, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Penttinen, R.P.; Kobayashi, S.; Bornstein, P. Transforming growth factor beta increases mRNA for matrix proteins both in the presence and in the absence of changes in mRNA stability. Proc. Natl. Acad. Sci. USA 1988, 85, 1105–1108. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.C.; Tan, A.R.; Olencki, T.E.; Shapiro, G.I.; Dezube, B.J.; Reiss, M.; Hsu, F.J.; Berzofsky, J.A.; Lawrence, D.P. Phase I study of GC1008 (fresolimumab): A human anti-transforming growth factor-beta (TGFbeta) monoclonal antibody in patients with advanced malignant melanoma or renal cell carcinoma. PLoS ONE 2014, 9, e90353. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.M.; Padilla, C.M.; McLaughlin, S.R.; Mathes, A.; Ziemek, J.; Goummih, S.; Nakerakanti, S.; York, M.; Farina, G.; Whitfield, M.L.; et al. Fresolimumab treatment decreases biomarkers and improves clinical symptoms in systemic sclerosis patients. J. Clin. Investig. 2015, 125, 2795–2807. [Google Scholar] [CrossRef] [PubMed]
- Vincenti, F.; Fervenza, F.C.; Campbell, K.N.; Diaz, M.; Gesualdo, L.; Nelson, P.; Praga, M.; Radhakrishnan, J.; Sellin, L.; Singh, A.; et al. Focal Segmental Glomerulosclerosis Study, G., A Phase 2, Double-Blind, Placebo-Controlled, Randomized Study of Fresolimumab in Patients With Steroid-Resistant Primary Focal Segmental Glomerulosclerosis. Kidney Int. Rep. 2017, 2, 800–810. [Google Scholar] [CrossRef]
- Folkman, J. Angiogenesis. Annu. Rev. Med. 2006, 57, 1–18. [Google Scholar] [CrossRef]
- Wang, Z.H.; Zhu, Z.T.; Xiao, X.Y.; Sun, J. Correlation of serum levels of endostatin with tumor stage in gastric cancer: A systematic review and meta-analysis. Biomed. Res. Int. 2015, 2015, 623939. [Google Scholar] [CrossRef]
- Gao, S.R.; Li, L.M.; Xia, H.P.; Wang, G.M.; Xu, H.Y.; Wang, A.R. Clinical observation on recombinant human endostatin combined with chemotherapy for advanced gastrointestinal cancer. Asian Pac. J. Cancer Prev. 2015, 16, 4037–4040. [Google Scholar] [CrossRef]
- Yao, J.; Fan, L.; Peng, C.; Huang, A.; Liu, T.; Lin, Z.; Yang, Q.; Zhang, T.; Ma, H. Clinical efficacy of endostar combined with chemotherapy in the treatment of peritoneal carcinomatosis in gastric cancer: Results from a retrospective study. Oncotarget 2017, 8, 70788–70797. [Google Scholar] [CrossRef][Green Version]
- Ponce, M.L.; Hibino, S.; Lebioda, A.M.; Mochizuki, M.; Nomizu, M.; Kleinman, H.K. Identification of a potent peptide antagonist to an active laminin-1 sequence that blocks angiogenesis and tumor growth. Cancer Res. 2003, 63, 5060–5064. [Google Scholar]
- Piazuelo, M.B.; Riechelmann, R.P.; Wilson, K.T.; Algood, H.M.S. Resolution of Gastric Cancer-Promoting Inflammation: A Novel Strategy for Anti-cancer Therapy. Curr Top. Microbiol. Immunol. 2019, 421, 319–359. [Google Scholar]
- Hsieh, H.L.; Tsai, M.M. Tumor progression-dependent angiogenesis in gastric cancer and its potential application. World J. Gastrointest. Oncol. 2019, 11, 686–704. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, C.S.; Tomasek, J.; Yong, C.J.; Dumitru, F.; Passalacqua, R.; Goswami, C.; Safran, H.; Dos Santos, L.V.; Aprile, G.; Ferry, D.R.; et al. Ramucirumab monotherapy for previously treated advanced gastric or gastro-oesophageal junction adenocarcinoma (REGARD): An international, randomised, multicentre, placebo-controlled, phase 3 trial. Lancet 2014, 383, 31–39. [Google Scholar] [CrossRef]
- Hironaka, S. Anti-angiogenic therapies for gastric cancer. Asia Pac. J. Clin. Oncol. 2019, 15, 208–217. [Google Scholar] [CrossRef]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).