Astroglial Connexin 43 Deficiency Protects against LPS-Induced Neuroinflammation: A TSPO Brain µPET Study with [18F]FEPPA

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Reagents for Radiochemistry
2.3. [18F]FEPPA Radiosynthesis and PET/CT Imaging
2.4. Image Analysis and Pharmacokinetic Modeling
2.5. Western Blot
2.6. Fluorescent in Situ Hybridization
2.7. Statistical Analysis
3. Results
3.1. Astrocyte Cx43 Deletion Does Not Alter [18F]FEPPA Brain Distribution
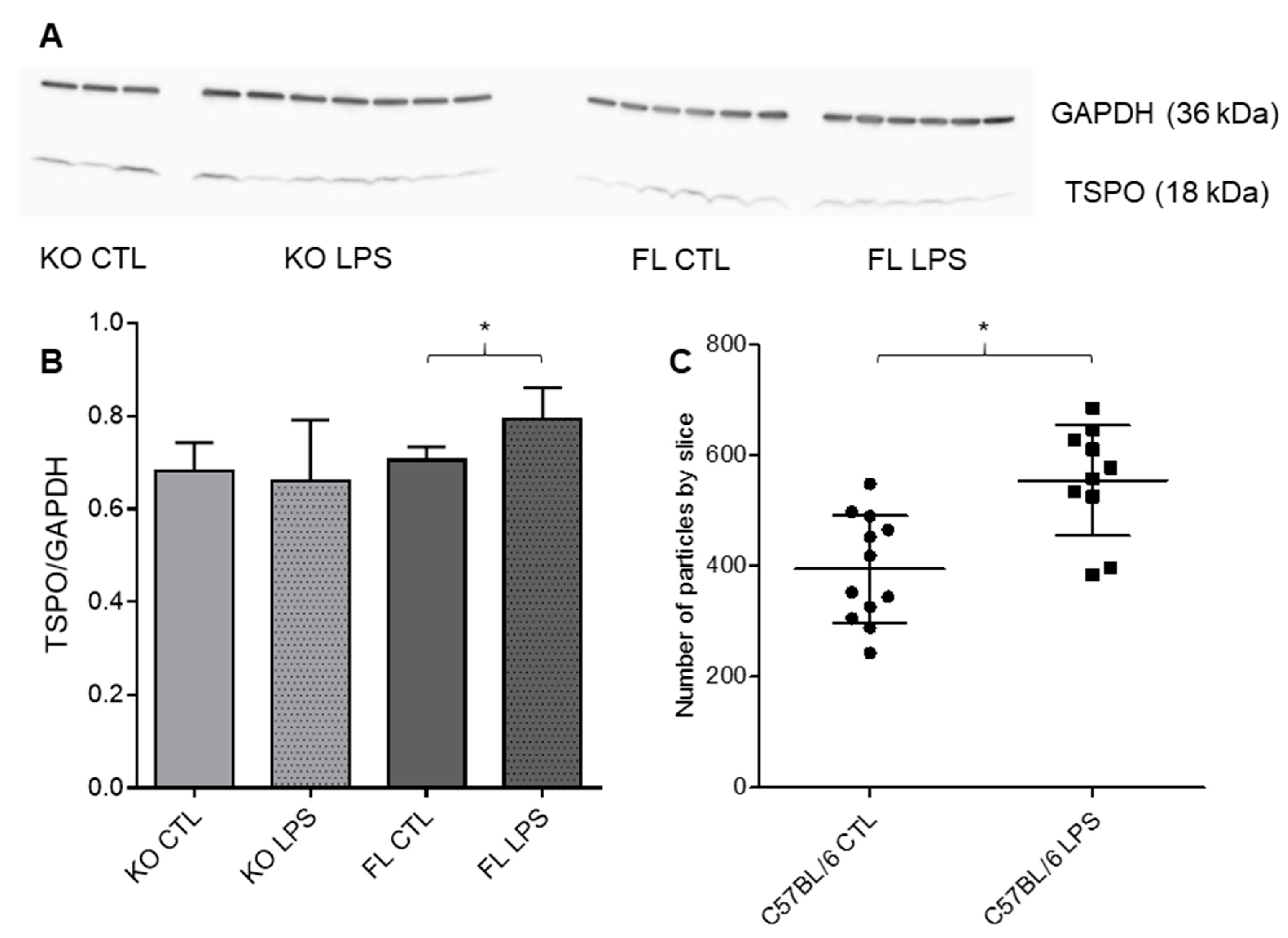
3.2. TSPO Brain Expression in FL Cx43 and KO Cx43 Mice with or without LPS Injection
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Reichenberg, A.; Yirmiya, R.; Schuld, A.; Kraus, T.; Haack, M.; Morag, A.; Pollmächer, T. Cytokine-associated emotional and cognitive disturbances in humans. Arch. Gen. Psychiatry 2001, 58, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Kayser, M.S.; Dalmau, J. Anti-NMDA receptor encephalitis, autoimmunity, and psychosis. Schizophr. Res. 2016, 176, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Goff, D.C.; Henderson, D.C. Inflammation and schizophrenia. Expert Rev. Neurother 2007, 7, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Masdeu, J.C.; Dalmau, J.; Berman, K.F. NMDA Receptor Internalization by Autoantibodies: A Reversible Mechanism Underlying Psychosis? Trends Neurosci. 2016, 39, 300. [Google Scholar] [CrossRef]
- Guilarte, T.R. TSPO in diverse CNS pathologies and psychiatric disease: A critical review and a way forward. Pharmacol. Ther. 2019, 194, 44–58. [Google Scholar] [CrossRef]
- Lagarde, J.; Sarazin, M.; Bottlaender, M. In vivo PET imaging of neuroinflammation in Alzheimer’s disease. J Neural Transm 2018, 125, 847–867. [Google Scholar] [CrossRef]
- Roussakis, A.-A.; Piccini, P. Molecular Imaging of Neuroinflammation in Idiopathic Parkinson’s Disease. Int. Rev. Neurobiol. 2018, 141, 347–363. [Google Scholar]
- Chen, M.-K.; Guilarte, T.R. Imaging the Peripheral Benzodiazepine Receptor Response in Central Nervous System Demyelination and Remyelination. Toxicol. Sci. 2006, 91, 532–539. [Google Scholar] [CrossRef]
- Enache, D.; Pariante, C.M.; Mondelli, V. Markers of central inflammation in major depressive disorder: A systematic review and meta-analysis of studies examining cerebrospinal fluid, positron emission tomography and post-mortem brain tissue. Brain Behav. Immun. 2019, S0889159118312121. [Google Scholar] [CrossRef]
- De Picker, L.J.; Morrens, M.; Chance, S.A.; Boche, D. Microglia and Brain Plasticity in Acute Psychosis and Schizophrenia Illness Course: A Meta-Review. Front. Psychiatry 2017, 8, 238. [Google Scholar] [CrossRef]
- Boulay, A.-C.; Cisternino, S.; Cohen-Salmon, M. Immunoregulation at the gliovascular unit in the healthy brain: A focus on Connexin 43. Brain Behav. Immun. 2016, 56, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Franco-Pérez, J.; Ballesteros-Zebadúa, P.; Fernández-Figueroa, E.A.; Ruiz-Olmedo, I.; Reyes-Grajeda, P.; Paz, C. Sleep deprivation and sleep recovery modifies connexin36 and connexin43 protein levels in rat brain. Neuro. Report 2012, 23, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Bernard, R.; Kerman, I.A.; Thompson, R.C.; Jones, E.G.; Bunney, W.E.; Barchas, J.D.; Schatzberg, A.F.; Myers, R.M.; Akil, H.; Watson, S.J. Altered expression of glutamate signaling, growth factor, and glia genes in the locus coeruleus of patients with major depression. Mol. Psychiatry 2011, 16, 634–646. [Google Scholar] [CrossRef]
- Miguel-Hidalgo, J.J.; Wilson, B.A.; Hussain, S.; Meshram, A.; Rajkowska, G.; Stockmeier, C.A. Reduced connexin 43 immunolabeling in the orbitofrontal cortex in alcohol dependence and depression. J. Psychiatr. Res. 2014, 55, 101–109. [Google Scholar] [CrossRef]
- Boulay, A.-C.; Mazeraud, A.; Cisternino, S.; Saubaméa, B.; Mailly, P.; Jourdren, L.; Blugeon, C.; Mignon, V.; Smirnova, M.; Cavallo, A.; et al. Immune quiescence of the brain is set by astroglial connexin 43. J. Neurosci. 2015, 35, 4427–4439. [Google Scholar] [CrossRef]
- Boulay, A.-C.; Gilbert, A.; Moreira, V.O.; Blugeon, C.; Perrin, S.; Pouch, J.; Crom, S.L.; Ducos, B.; Cohen-Salmon, M. Connexin 43 Controls the Astrocyte Immunoregulatory Phenotype. Brain Sci. 2018, 8, 50. [Google Scholar] [CrossRef]
- Banks, W.A.; Gray, A.M.; Erickson, M.A.; Salameh, T.S.; Damodarasamy, M.; Sheibani, N.; Meabon, J.S.; Wing, E.E.; Morofuji, Y.; Cook, D.G.; et al. Lipopolysaccharide-induced blood-brain barrier disruption: Roles of cyclooxygenase, oxidative stress, neuroinflammation, and elements of the neurovascular unit. J. Neuroinflamm. 2015, 12, 223. [Google Scholar] [CrossRef]
- Theis, M.; Mas, C.; Döring, B.; Krüger, O.; Herrera, P.; Meda, P.; Willecke, K. General and conditional replacement of connexin43-coding DNA by a lacZ reporter gene for cell-autonomous analysis of expression. Cell Commun. Adhes. 2001, 8, 383–386. [Google Scholar] [CrossRef]
- Vignal, N.; Cisternino, S.; Rizzo-Padoin, N.; San, C.; Hontonnou, F.; Gelé, T.; Declèves, X.; Sarda-Mantel, L.; Hosten, B. [18F]FEPPA a TSPO Radioligand: Optimized Radiosynthesis and Evaluation as a PET Radiotracer for Brain Inflammation in a Peripheral LPS-Injected Mouse Model. Molecules 2018, 23, 1375. [Google Scholar] [CrossRef]
- Rizzo, G.; Veronese, M.; Tonietto, M.; Zanotti-Fregonara, P.; Turkheimer, F.E.; Bertoldo, A. Kinetic modeling without accounting for the vascular component impairs the quantification of [11C]PBR28 brain PET data. J. Cereb. Blood Flow Metab. 2014, 34, 1060–1069. [Google Scholar] [CrossRef] [PubMed]
- Veronese, M.; Reis Marques, T.; Bloomfield, P.S.; Rizzo, G.; Singh, N.; Jones, D.; Agushi, E.; Mosses, D.; Bertoldo, A.; Howes, O.; et al. Kinetic modelling of [11C]PBR28 for 18 kDa translocator protein PET data: A validation study of vascular modelling in the brain using XBD173 and tissue analysis. J. Cereb. Blood Flow Metab. 2017. [Google Scholar] [CrossRef] [PubMed]
- Wimberley, C.; Lavisse, S.; Brulon, V.; Peyronneau, M.-A.; Leroy, C.; Bodini, B.; Remy, P.; Stankoff, B.; Buvat, I.; Bottlaender, M. Impact of endothelial TSPO on the quantification of 18F-DPA-714. J. Nucl. Med. 2017. [Google Scholar] [CrossRef]
- Oudart, M.; Tortuyaux, R.; Mailly, P.; Mazaré, N.; Boulay, A.-C.; Cohen-Salmon, M. AstroDot: A new method for studying the spatial distribution of mRNA in astrocytes. bioRxiv 2019. [Google Scholar] [CrossRef]
- Chever, O.; Lee, C.-Y.; Rouach, N. Astroglial connexin43 hemichannels tune basal excitatory synaptic transmission. J. Neurosci. 2014, 34, 11228–11232. [Google Scholar] [CrossRef]
- Rusjan, P.M.; Wilson, A.A.; Bloomfield, P.M.; Vitcu, I.; Meyer, J.H.; Houle, S.; Mizrahi, R. Quantitation of translocator protein binding in human brain with the novel radioligand [18F]-FEPPA and positron emission tomography. J. Cereb. Blood Flow Metab. 2011, 31, 1807–1816. [Google Scholar] [CrossRef]
- Zhou, J.-J.; Cheng, C.; Qiu, Z.; Zhou, W.-H.; Cheng, G.-Q. Decreased connexin 43 in astrocytes inhibits the neuroinflammatory reaction in an acute mouse model of neonatal sepsis. Neurosci Bull. 2015, 31, 763–768. [Google Scholar] [CrossRef][Green Version]
- Auvity, S.; Goutal, S.; Thézé, B.; Chaves, C.; Hosten, B.; Kuhnast, B.; Saba, W.; Boisgard, R.; Buvat, I.; Cisternino, S.; et al. Evaluation of TSPO PET imaging, a marker of glial activation, to study the neuroimmune footprints of morphine exposure and withdrawal. Drug Alcohol Depend. 2017, 170, 43–50. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | K1 (mL × cm−3 × min−1) | k2 (min−1) | k3/k4 | Kb (min−1) | VT (mL × cm−3) |
|---|---|---|---|---|---|
| FL Cx43 | 1.31 ± 0.43 | 0.70 ± 0.34 | 0.65 ± 0.35 | 0.77 ± 0.12 | 3.24 ± 0.67 |
| FL Cx43 + LPS | 1.70 ± 0.44 | 0.70 ± 0.23 | 0.35 ± 0.24 | 1.11 ± 0.59 | 4.34 ± 0.68 |
| p value | ns | ns | ns | ns | 0.0204 * |
| KO Cx43 | 1.31 ± 0.48 | 0.75 ± 0.39 | 0.25 ± 0.33 | 0.99 ± 0.23 | 3.18 ± 0.63 |
| KO Cx43 + LPS | 1.32 ± 0.19 | 0.60 ± 0.25 | 0.19 ± 0.18 | 0.96 ± 0.22 | 3.95 ± 1.14 |
| p value | ns | ns | ns | ns | ns |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vignal, N.; Boulay, A.-C.; San, C.; Cohen-Salmon, M.; Rizzo-Padoin, N.; Sarda-Mantel, L.; Declèves, X.; Cisternino, S.; Hosten, B. Astroglial Connexin 43 Deficiency Protects against LPS-Induced Neuroinflammation: A TSPO Brain µPET Study with [18F]FEPPA. Cells 2020, 9, 389. https://doi.org/10.3390/cells9020389
Vignal N, Boulay A-C, San C, Cohen-Salmon M, Rizzo-Padoin N, Sarda-Mantel L, Declèves X, Cisternino S, Hosten B. Astroglial Connexin 43 Deficiency Protects against LPS-Induced Neuroinflammation: A TSPO Brain µPET Study with [18F]FEPPA. Cells. 2020; 9(2):389. https://doi.org/10.3390/cells9020389
Chicago/Turabian StyleVignal, Nicolas, Anne-Cécile Boulay, Carine San, Martine Cohen-Salmon, Nathalie Rizzo-Padoin, Laure Sarda-Mantel, Xavier Declèves, Salvatore Cisternino, and Benoît Hosten. 2020. "Astroglial Connexin 43 Deficiency Protects against LPS-Induced Neuroinflammation: A TSPO Brain µPET Study with [18F]FEPPA" Cells 9, no. 2: 389. https://doi.org/10.3390/cells9020389
APA StyleVignal, N., Boulay, A.-C., San, C., Cohen-Salmon, M., Rizzo-Padoin, N., Sarda-Mantel, L., Declèves, X., Cisternino, S., & Hosten, B. (2020). Astroglial Connexin 43 Deficiency Protects against LPS-Induced Neuroinflammation: A TSPO Brain µPET Study with [18F]FEPPA. Cells, 9(2), 389. https://doi.org/10.3390/cells9020389